Alpha-Lactalbumin Enriched Whey Protein Concentrate to Improve Gut, Immunity and Brain Development in Preterm Pigs

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Diets and Nutrition
2.3. Clinical Evaluation and Animal Procedures
2.4. Body Composition and Tissue Collection
2.5. Blood Biochemistry and Blood Analyses
2.6. Hematology and Systemic Immunity
2.7. Gut Morphology
2.8. Gut Function
2.9. Microbiome and Microbial Metabolites
2.10. Physical Activity, Behavior and Cognition
2.11. Statistics
3. Results
3.1. Clinical Characteristics and Growth
3.2. Organ Weights
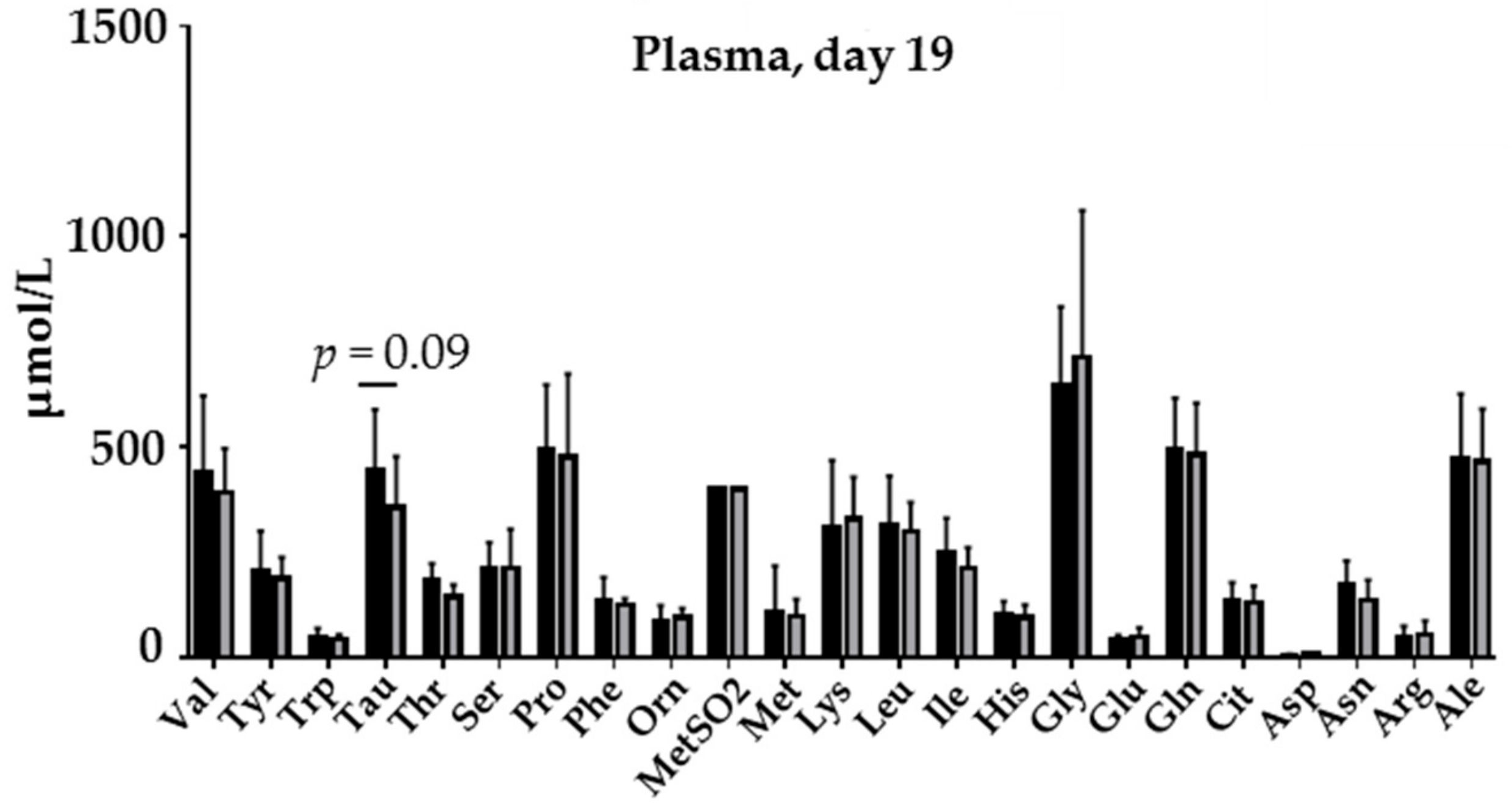
3.3. Biochemistry, Amino Acid Profile and Blood Analyses
3.4. Hematology and Systemic Immunity
3.5. Gut Structure and Function
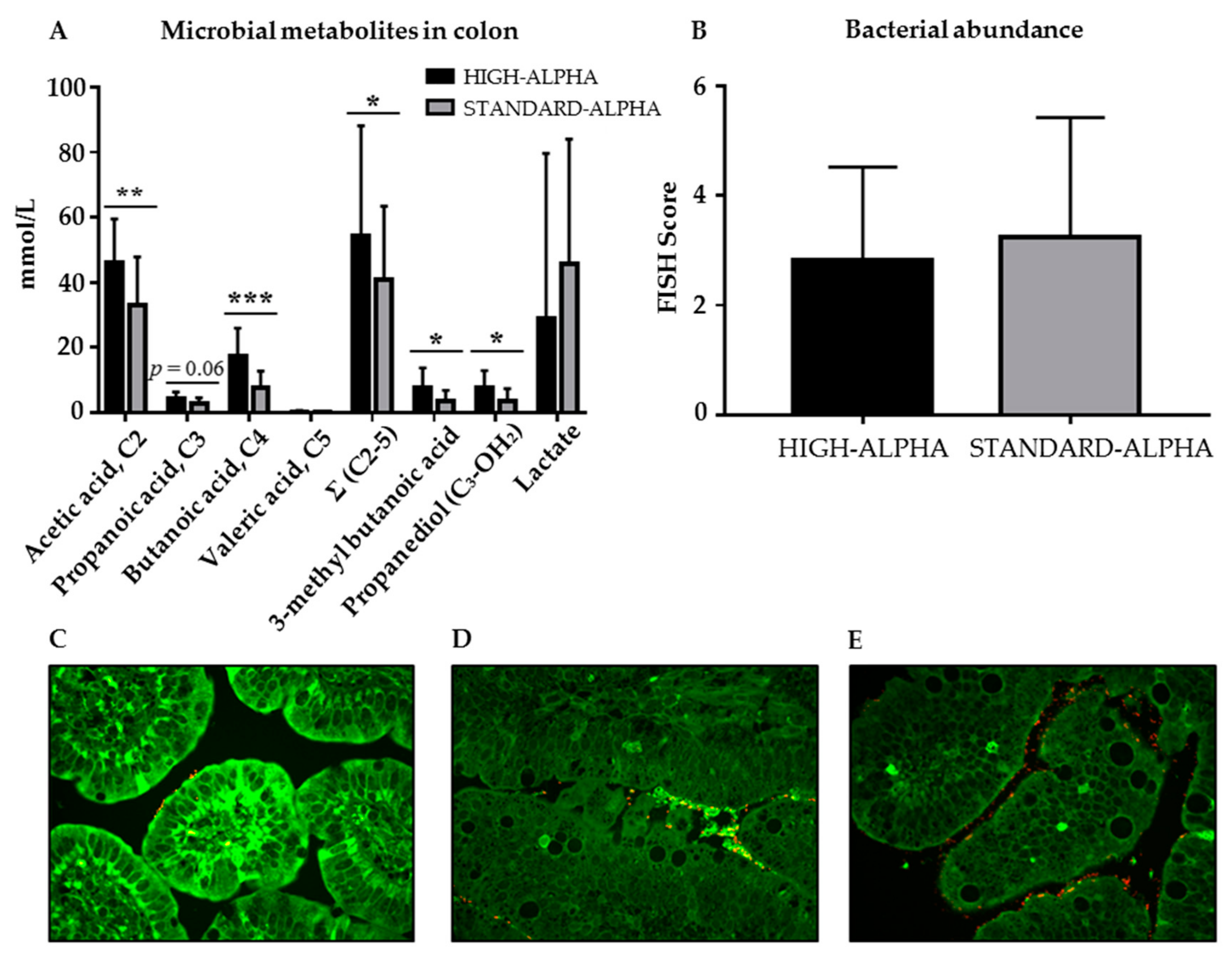
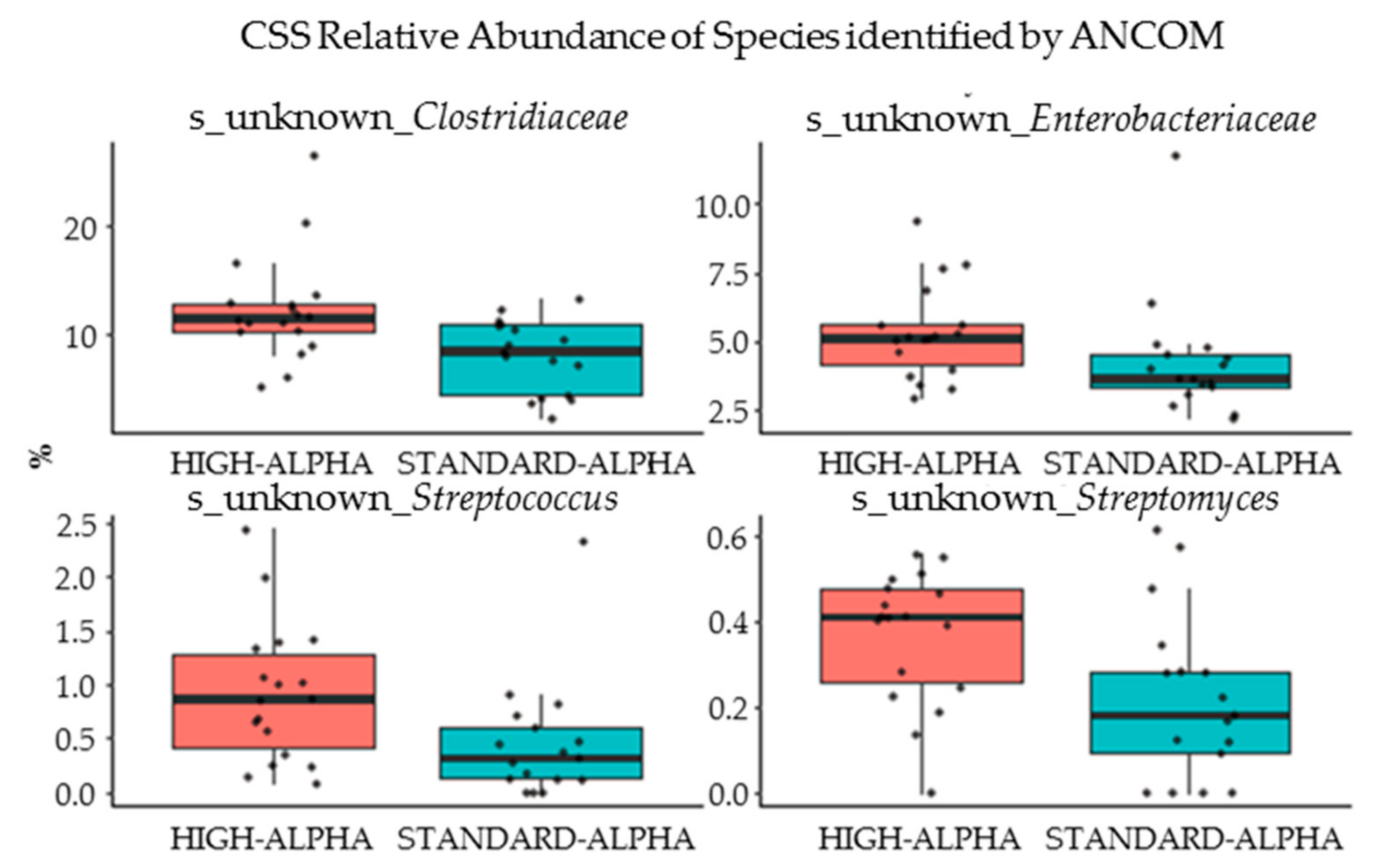
3.6. Microbiome and Microbial Metabolites
3.7. Physical Activity, Behavior and Cognition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blencowe, H.; Cousens, S.; Oestergaard, M.Z.; Chou, D.; Moller, A.-B.; Narwal, R.; Adler, A.; Vera Garcia, C.; Rohde, S.; Say, L.; et al. National, regional, and worldwide estimates of preterm birth rates in the year 2010 with time trends since 1990 for selected countries: A systematic analysis and implications. Lancet 2012, 379, 2162–2172. [Google Scholar] [CrossRef] [Green Version]
- Saigal, S.; Doyle, L.W. An overview of mortality and sequelae of preterm birth from infancy to adulthood. Lancet 2008, 371, 261–269. [Google Scholar] [CrossRef]
- Wang, M.L.; Dorer, D.J.; Fleming, M.P.; Catlin, E.A. Clinical outcomes of near-term infants. Pediatrics 2004, 114, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Fanaro, S. Strategies to improve feeding tolerance in preterm infants. J. Matern. Neonatal. Med. 2012, 25, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.; Cole, T.J. Breast milk and neonatal necrotising enterocolitis. Lancet 1990, 336, 1519–1523. [Google Scholar] [CrossRef]
- Hack, M.; Klein, N.; Taylor, G.H. School-age outcomes of children of extremely low birthweight and gestational age. Semin. Neonatol. 1996, 1, 277–288. [Google Scholar] [CrossRef]
- Finnström, O.; Otterblad Olausson, P.; Sedin, G.; Serenius, F.; Svenningsen, N.; Thiringer, K.; Tunell, R.; Wesström, G. Neurosensory outcome and growth at three years in extremely low birthweight infants: Follow-up results from the Swedish national prospective study. Acta Paediatr. Int. J. Paediatr. 1998, 87, 1055–1060. [Google Scholar] [CrossRef]
- Mathiasen, R.; Hansen, B.M.; Andersen, A.-M.N.N.; Forman, J.L.; Greisen, G. Gestational Age and Basic School Achievements: A National Follow-up Study in Denmark. Pediatrics 2010, 126, e1553–e1561. [Google Scholar] [CrossRef]
- Lawrence, R.M.; Pane, C.A. Human Breast Milk: Current Concepts of Immunology and Infectious Diseases. Curr. Probl. Pediatr. Adolesc. Health Care 2007, 37, 7–36. [Google Scholar] [CrossRef]
- Davis, A.M.; Harris, B.J.; Lien, E.L.; Pramuk, K.; Trabulsi, J. α-Lactalbumin-rich infant formula fed to healthy term infants in a multicenter study: Plasma essential amino acids and gastrointestinal tolerance. Eur. J. Clin. Nutr. 2008, 62, 1294–1301. [Google Scholar] [CrossRef] [Green Version]
- Brew, K.; Castellino, F.J.; Vanaman, T.C.; Hill, R.L. The Complete Amino Acid Sequence of Bovine α-Lactalbumin. J. Biol. Chem. 1970, 245, 4570–4582. [Google Scholar] [PubMed]
- Findlay, J.B.C.; Brew, K. The Complete Amino-Acid Sequence of Human α-Lactalbumin. Eur. J. Biochem. 1972, 27, 65–86. [Google Scholar] [CrossRef] [PubMed]
- Donovan, S.M. Human Milk Proteins: Composition and Physiological Significance. Nestle Nutr. Inst. Workshop Ser. 2019, 90, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruddick, J.P.; Evans, A.K.; Nutt, D.J.; Lightman, S.L.; Rook, G.A.W.; Lowry, C.A. Tryptophan metabolism in the central nervous system: Medical implications. Expert Rev. Mol. Med. 2006, 8, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Markus, C.R.; Olivier, B.; De Haan, E.H.F. Whey protein rich in α-lactalbumin increases the ratio of plasma tryptophan to the sum of the other large neutral amino acids and improves cognitive performance in stress-vulnerable subjects. Am. J. Clin. Nutr. 2002, 75, 1051–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booij, L.; Merens, W.; Markus, C.R.; Van der Does, A.J.W. Diet rich in α-lactalbumin improves memory in unmedicated recovered depressed patients and matched controls. J. Psychopharmacol. 2006, 20, 526–535. [Google Scholar] [CrossRef]
- Minet-Ringuet, J.; Le Ruyet, P.M.; Tomé, D.; Even, P.C. A tryptophan-rich protein diet efficiently restores sleep after food deprivation in the rat. Behav. Brain Res. 2004, 152, 335–340. [Google Scholar] [CrossRef]
- Lien, E.L.; Davis, A.M.; Euler, A.R. Growth and Safety in Term Infants Fed Reduced-Protein Formula with Added Bovine Alpha-Lactalbumin. J. Pediatr. Gastroenterol. Nutr. 2004, 38, 170–176. [Google Scholar] [CrossRef] [Green Version]
- Trabulsi, J.; Capeding, R.; Lebumfacil, J.; Ramanujam, K.; Feng, P.; Mcsweeney, S.; Harris, B.; DeRusso, P. Effect of an α-lactalbumin-enriched infant formula with lower protein on growth. Eur. J. Clin. Nutr. 2011, 65, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Kelleher, S.L.; Chatterton, D.; Nielsen, K.; Lönnerdal, B. Glycomacropeptide and α-lactalbumin supplementation of infant formula affects growth and nutritional status in infant rhesus monkeys1-3. Am. J. Clin. Nutr. 2003, 77, 1261–1268. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, A.; Thomas, U.; Bramaz, N.; Hunziker, P.; von Fellenberg, R. Isolation and identification of three bactericidal domains in the bovine α-lactalbumin molecule. Biochim. Biophys. Acta 1999, 1426, 439–448. [Google Scholar] [CrossRef]
- Brück, W.M.; Graverholt, G.; Gibson, G.R. Use of batch culture and a two-stage continuous culture system to study the effect of supplemental alpha-lactalbumin and glycomacropeptide on mixed populations of human gut bacteria. FEMS Microbiol. Ecol. 2002, 41, 231–237. [Google Scholar] [CrossRef]
- Brück, W.M.; Redgrave, M.; Tuohy, K.M.; Lönnerdal, B.; Graverholt, G.; Hernell, O.; Gibson, G.R. Effects of Bovine α-Lactalbumin and Casein Glycomacropeptide—enriched Infant Formulae on Faecal Microbiota in Healthy Term Infants. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 673–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petschow, B.W.; Talbott, R.D. Response of Bifidobacterium Species to Growth Promoters in Human and Cow Milk. Pediatr. Res. 1991, 29, 208–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Håkansson, A.; Zhivotovsky, B.; Orrenius, S.; Sabharwal, H.; Svanborg, C. Apoptosis induced by a human milk protein. Proc. Natl. Acad. Sci. USA 1995, 92, 8064–8068. [Google Scholar] [CrossRef] [Green Version]
- Jaziri, M.; Migliore-Samour, D.; Casabianca-pignède, M.; Keddad, K.; Morgat, J.L.; Jollès, P. Specific binding sites on human phagocytic blood cells for Gly-Leu-Phe and Val-Glu-Pro-Ile-Pro-Tyr, immunostimulating peptides from human milk proteins. Biochim. Biophys. Acta 1992, 1160, 251–261. [Google Scholar] [CrossRef]
- Sangild, P.T. Gut Responses to Enteral Nutrition in Preterm Infants and Animals. Exp. Biol. Med. 2006, 231, 1695–1711. [Google Scholar] [CrossRef]
- Sangild, P.T.; Thymann, T.; Schmidt, M.; Stoll, B.; Burrin, D.G.; Buddington, R.K. The preterm pig as a model in pediatric gastroenterology. J. Anim. Sci. 2013, 91, 4713–6359. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.D.; Sangild, P.T.; Munch, S.L.; van der Beek, E.M.; Renes, I.B.; Ginneken, C.V.; Greisen, G.O.; Thymann, T. Delayed growth, motor function and learning in preterm pigs during early postnatal life. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R481–R492. [Google Scholar] [CrossRef] [Green Version]
- Dobbing, J.; Sands, J. Comparative aspects of the brain growth spurt. Early Hum. Dev. 1979, 3, 79–83. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, Y.; Pan, X.; Nguyen, D.N.; Brunse, A.; Bojesen, A.M.; Rudloff, S.; Mortensen, M.S.; Burrin, D.G.; Sangild, P.T. Human milk fortification with bovine colostrum is superior to formula-based fortifiers to prevent gut dysfunction, necrotizing enterocolitis, and systemic infection in preterm pigs. J. Parenter. Enter. Nutr. 2019, 43, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, Y.; Nguyen, D.N.; Mortensen, M.S.; van den Akker, C.H.; Skeath, T.; Pors, S.E.; Pankratova, S.; Rudloff, S.; Sørensen, S.J.; et al. Nutrient Fortification of Human Donor Milk Affects Intestinal Function and Protein Metabolism in Preterm Pigs. J. Nutr. 2018, 148, 336–347. [Google Scholar] [CrossRef] [Green Version]
- Stoll, B.; Chang, X.; Fan, M.Z.; Reeds, P.J.; Burrin, D.G. Enteral nutrient intake level determines intestinal protein synthesis and accretion rates in neonatal pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G288–G294. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Hui, Y.; Goericke-Pesch, S.; Pankratova, S.; Kot, W.; Pan, X.; Thymann, T.; Sangild, P.T.; Nguyen, D.N. Gut and immune effects of bioactive milk factors in preterm pigs exposed to prenatal inflammation. Am. J. Physiol. Liver Physiol. 2019, 317, G67–G77. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Jiang, P.; Frøkiær, H.; Heegaard, P.M.H.; Thymann, T.; Sangild, P.T. Delayed development of systemic immunity in preterm pigs as a model for preterm infants. Sci. Rep. 2016, 6, 36816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skovsted Cilieborg, M.; Boye, M.; Thymann, T.; Borg Jensen, B.; Torp Sangild, P. Diet-dependent effects of minimal enteral nutrition on intestinal function and necrotizing enterocolitis in preterm pigs. J. Parenter. Enter. Nutr. 2011, 35, 32–42. [Google Scholar] [CrossRef]
- Thymann, T.; Burrin, D.G.; Tappenden, K.A.; Bjornvad, C.R.; Jensen, S.K.; Sangild, P.T. Formula-feeding reduces lactose digestive capacity in neonatal pigs. Br. J. Nutr. 2006, 95, 1075–1081. [Google Scholar] [CrossRef] [Green Version]
- Ørskov, C.; Rabenhøj, L.; Wettergren, A.; Kofod, H.; Holst, J.J. Tissue and Plasma Concentrations of Amidated and Glycine-Extended Glucagon-Like Peptide I in Humans. Diabetes 1994, 43, 535–539. [Google Scholar] [CrossRef]
- Sangild, P.T.; Siggers, R.H.; Schmidt, M.; Elnif, J.; Bjornvad, C.R.; Thymann, T.; Grondahl, M.L.; Hansen, A.K.; Jensen, S.K.; Boye, M.; et al. Diet- and Colonization-Dependent Intestinal Dysfunction Predisposes to Necrotizing Enterocolitis in Preterm Pigs. Gastroenterology 2006, 130, 1776–1792. [Google Scholar] [CrossRef]
- Cilieborg, M.S.; Sangild, P.T.; Jensen, M.L.; Østergaard, M.V.; Christensen, L.; Rasmussen, S.O.; Mørbak, A.L.; Jørgensen, C.B.; Bering, S.B. α1,2-Fucosyllactose Does Not Improve Intestinal Function or Prevent Escherichia coli F18 Diarrhea in Newborn Pigs. J. Pediatric Gastroenterol. Nutr. 2017, 64, 310–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, T.S.; de Vries, L.; Kot, W.; Hansen, L.H.; Castro-Mejía, J.L.; Vogensen, F.K.; Hansen, A.K.; Nielsen, D.S. Mouse Vendor Influence on the Bacterial and Viral Gut Composition Exceeds the Effect of Diet. Viruses 2019, 11, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, S.O.; Martin, L.; Østergaard, M.V.; Rudloff, S.; Li, Y.; Roggenbuck, M.; Bering, S.B.; Sangild, P.T. Bovine colostrum improves neonatal growth, digestive function and gut immunity relative to donor human milk and infant formula in preterm pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G480–G491. [Google Scholar] [CrossRef] [Green Version]
- Obelitz-Ryom, K.; Rendboe, A.K.; Nguyen, D.N.; Rudloff, S.; Brandt, A.B.; Nielsen, D.S.; Heckmann, A.B.; Chichlowski, M.; Sangild, P.T.; Thymann, T.; et al. Bovine milk oligosaccharides with sialyllactose for preterm piglets. Nutrients 2018, 10, 1489. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.H.; Brandt, A.B.; Thymann, T.; Obelitz-Ryom, K.; Jiang, P.; Hole, C.V.; van Ginneken, C.; Pankratova, S.; Sangild, P.T. Rapid Postnatal Adaptation of Neurodevelopment in Pigs Born Late Preterm. Dev. Neurosci. 2018, 40, 586–600. [Google Scholar] [CrossRef]
- Andersen, A.D.; Nguyen, D.N.; Langhorn, L.; Renes, I.B.; Van Elburg, R.M.; Hartog, A.; Tims, S.; Van De Looij, Y.; Sangild, P.T.; Thymann, T. Synbiotics Combined with Glutamine Stimulate Brain Development and the Immune System in Preterm Pigs. J. Nutr. 2019, 148, 36–45. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.H.; Thomas, P.; Peabody, J. Extrauterine growth restriction remains a serious problem in prematurely born neonates. Pediatrics 2003, 111, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Grier, A.; Qiu, X.; Bandyopadhyay, S.; Holden-wiltse, J.; Kessler, H.A.; Gill, A.L.; Hamilton, B.; Huyck, H.; Misra, S.; Mariani, T.J.; et al. Impact of prematurity and nutrition on the developing gut microbiome and preterm infant growth. Microbiome 2017, 5, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Claud, E.C. Connection between gut microbiome and brain development in preterm infants. Dev. Psychobiol. 2019, 61, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Niño, D.F.; Sodhi, C.P.; Hackam, D.J. Necrotizing enterocolitis: New insights into pathogenesis and mechanisms. Nat. Rev. Gatroenterol. Hepatol. 2016, 13, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.T.; Lacroix, C.; Braegger, C.P.; Chassard, C. Lactate-utilizing community is associated with gut microbiota dysbiosis in colicky infants. Sci. Rep. 2017, 7, 11176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Lu, L.; Yu, Y.; Cluette-brown, J.; Martin, C.R.; Claud, E.C. Effects of Intestinal Microbiota on Brain Development in Humanized Gnotobiotic Mice. Sci. Rep. 2018, 8, 5443. [Google Scholar] [CrossRef] [Green Version]
- Goodman, R.E.; Pickett, M.J. Delayed Lactose Fermentation by Enterobacteriaceae 1. J. Bacteriol. 1966, 92, 318–327. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.A.; Sherman, J.M. The Lactic Acid Fermentation of Streptococci. J. Bacteriol. 1941, 43, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Tang, I.C.; Yang, S.; Okos, M.R. Acetic acid production from whey lactose by the co-culture of Streptococcus lactis and CIostridium formicoaceticum. Appl. Microbiol. Biotechnol. 1988, 28, 138–143. [Google Scholar] [CrossRef]
- Siigur, U.; Ormisson, A.; Tamm, A. Faecal short-chain fatty acids in breast-fed and bottle-fed infants. Acta Paediatr. 1993, 82, 536–538. [Google Scholar] [CrossRef]
- Bourassa, M.W.; Alim, I.; Bultman, S.J.; Ratan, R.R. Butyrate, Neuroepigenetics and the Gut Microbiome: Can a High Fiber Diet Improve Brain Health? Neurosci. Lett. 2016, 118, 6072–6078. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Taliyan, R.; Singh, S. Beneficial effects of sodium butyrate in 6-OHDA induced neurotoxicity and behavioral abnormalities: Modulation of histone deacetylase activity. Behav. Brain Res. 2015, 291, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, N.; Agis-Balboa, R.C.; Walter, J.; Sananbenesi, F.; Fischer, A. Sodium Butyrate Improves Memory Function in an Alzheimer’s Disease Mouse Model When Administered at an Advanced Stage of Disease Progression. J. Alzheimer’s Dis. 2011, 26, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Prentice, R.L.; Aragaki, A.K.; Howard, B.V.; Chlebowski, R.T.; Thomson, C.A.; Van Horn, L.; Tinker, L.F.; Manson, J.E.; Anderson, G.L.; Kuller, L.E.; et al. Low-Fat Dietary Pattern among Postmenopausal Women Influences Long-Term Cancer, Cardiovascular Disease, and Diabetes Outcomes. J. Nutr. 2019, 149, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourcyrous, M.; Nolan, V.G.; Goodwin, A.; Davis, S.L.; Buddington, R.K. Fecal Short-Chain Fatty Acids of Very-Low-Birth-Weight Preterm Infants Fed Expressed Breast Milk or Formula. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 725–731. [Google Scholar] [CrossRef]
- Cekmez, F.; Tanju, I.A.; Canpolat, F.E.; Aydinoz, S.; Aydemir, G.; Karademir, F.; Sarici, S.U. Mean platelet volume in very preterm infants: A predictor of morbidities? Eur. Rev. Med. Pharm. Sci. 2013, 17, 134–137. [Google Scholar] [CrossRef]
- Martin, W.F.; Armstrong, L.E.; Rodriguez, N.R. Dietary protein intake and renal function. Nutr. Metab. 2005, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Markus, C.R.; Olivier, B.; Panhuysen, G.E.M.; Van Der Gugten, J.; Alles, M.S.; Tuiten, A.; Westenberg, H.G.M.; Fekkes, D.; Koppeschaar, H.F.; de Haan, E.E.H.F. The bovine protein α-lactalbumin increases the plasma ratio of tryptophan to the other large neutral amino acids, and in vulnerable subjects raises brain serotonin activity, reduces cortisol concentration, and improves mood under stress 1–3. Am. J. Clin. Nutr. 2000, 71, 1536–1544. [Google Scholar] [CrossRef] [Green Version]
- Fernstrom, J.D.; Wurtman, R.J. Brain Serotonin Content: Physiological Dependence on Plasma Tryptophan Levels. Science 1971, 173, 149–152. [Google Scholar] [CrossRef] [Green Version]
- Dobbing, J.; Sands, J. Quantitative growth and development of human brain. Arch. Dis. Child. 1973, 48, 757–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherjon, S.A.; Smolders-DeHaas, H.; Kok, J.H.; Zondervan, H.A. The “brain-sparing“ effect: Antenatal cerebral Doppler findings in relation to neurologic outcome in very preterm infants. Am. J. Obs. Gynecol. 1993, 169, 169–175. [Google Scholar] [CrossRef]
- Layman, D.K.; Lönnerdal, B.; Fernstrom, J.D. Applications for α-lactalbumin in human nutrition. Nutr. Rev. 2018, 76, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Neville, M.C.; Keller, R.; Seacat, J.; Lutes, V.; Neifert, M.; Casey, C.; Allen, J.; Archer, P. Studies in human lactation: Milk volumes in lactating women during the onset of lactation and full lactation1-3. Am. J. Clin. Nutr. 1988, 48, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Velonà, T.; Abbiati, L.; Beretta, B.; Gaiaschi, A.; Flaúto, U.; Tagliabue, P.; Galli, C.L.; Restani, P. Protein Profiles in Breast Milk from Mothers Delivering Term and Preterm Babies. Pediatr. Res. 1999, 45, 658–663. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Content | HIGH-ALPHA | STANDARD-ALPHA | REF |
|---|---|---|---|
| Energy (kJ/L) | 2902 | 3012 | 2426 |
| Protein (g/L) | 55.0 | 55.0 | 27.0 |
| Carbohydrate (g/L) | 33.0 | 36.0 | 33.0 |
| Fat (g/L) | 38.0 | 40.0 | 38.0 |
| α-lactalbumin (g/L) | 18.0 | 6.3 | 0.7 |
| Osmolality (mOsm/kg) | 200 | 222 | 190 |
| Absolute Weight (g) | Relative Weight (g/kg) | |||||
|---|---|---|---|---|---|---|
| Organ | HIGH-ALPHA | STANDARD-ALPHA | p | HIGH-ALPHA | STANDARD-ALPHA | p |
| Small intestine | 56.1 ± 14 | 65.7 ± 19 | ns | 37.8 ± 5.0 | 41.5 ± 6.5 | * |
| Small intestinal length (cm) | 417 ± 41 | 461 ± 63 | ** | - | - | ns |
| Proximal small intestine | 19.9 ± 5.6 | 23.6 ± 7.0 | ** | 13.3 ± 1.5 | 15.0 ± 2.7 | * |
| Mid small intestine | 17.4 ± 4.4 | 20.3 ± 7.1 | * | 11.8 ± 2.2 | 12.7 ± 2.6 | ns |
| Distal small intestine | 18.8 ± 5.0 | 21.8 ± 6.1 | * | 12.7 ± 1.8 | 13.8 ± 2.3 | ns |
| Stomach | 9.21 ± 2.5 | 9.89 ± 4.0 | ns | 6.16 ± 0.7 | 6.26 ± 1.9 | ns |
| Colon | 40.5 ± 20 | 38.7 ± 19 | ns | 25.4 ± 12 | 25.4 ± 14 | ns |
| Liver | 40.3 ± 12 | 40.1 ± 12 | ns | 26.8 ± 3.3 | 25.2 ± 6.0 | ns |
| Spleen | 5.47 ± 2.3 | 5.83 ± 2.0 | ns | 3.60 ± 1.0 | 3.39 ± 1.1 | ns |
| Heart | 10.2 ± 3.0 | 10.8 ± 2.8 | ns | 6.79 ± 0.9 | 6.89 ± 1.0 | ns |
| Lungs | 34.6 ± 11 | 33.5 ± 10 | ns | 23.3 ± 4.6 | 20.2 ± 7.8 | ns |
| Kidneys | 10.5 ± 2.3 | 11.9 ± 2.2 | 0.08 | 7.10 ± 0.9 | 7.65 ± 1.2 | ns |
| Adrenal glands | 0.37 ± 0.1 | 0.33 ± 0.1 | ns | 0.25 ± 0.1 | 0.21 ± 0.1 | 0.08 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, C.H.; Hui, Y.; Nguyen, D.N.; Ahnfeldt, A.M.; Burrin, D.G.; Hartmann, B.; Heckmann, A.B.; Sangild, P.T.; Thymann, T.; Bering, S.B. Alpha-Lactalbumin Enriched Whey Protein Concentrate to Improve Gut, Immunity and Brain Development in Preterm Pigs. Nutrients 2020, 12, 245. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12010245
Nielsen CH, Hui Y, Nguyen DN, Ahnfeldt AM, Burrin DG, Hartmann B, Heckmann AB, Sangild PT, Thymann T, Bering SB. Alpha-Lactalbumin Enriched Whey Protein Concentrate to Improve Gut, Immunity and Brain Development in Preterm Pigs. Nutrients. 2020; 12(1):245. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12010245
Chicago/Turabian StyleNielsen, Charlotte Holme, Yan Hui, Duc Ninh Nguyen, Agnethe May Ahnfeldt, Douglas G. Burrin, Bolette Hartmann, Anne Birgitte Heckmann, Per Torp Sangild, Thomas Thymann, and Stine Brandt Bering. 2020. "Alpha-Lactalbumin Enriched Whey Protein Concentrate to Improve Gut, Immunity and Brain Development in Preterm Pigs" Nutrients 12, no. 1: 245. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12010245