Seabuckthorn (Hippophaë rhamnoides) Freeze-Dried Powder Protects against High-Fat Diet-Induced Obesity, Lipid Metabolism Disorders by Modulating the Gut Microbiota of Mice


Abstract
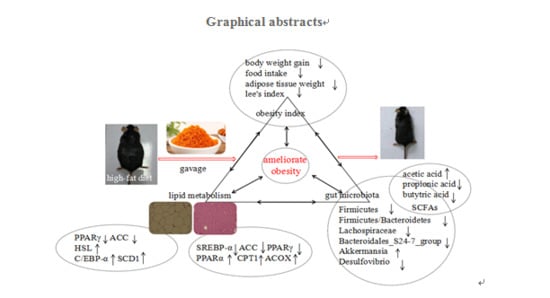
:
1. Introduction
2. Materials and Methods
2.1. Materials and Diets
2.2. Animals Experimental Design and Dietary Treatment
2.3. Sample Collection
2.4. Biochemical Analysis of Serum Parameters
2.5. Histological Analysis of Liver and Adipose Tissues
2.6. Gene Expression Analysis
2.7. Gut Microbiota Analysis
2.8. Gut Metabolites Analysis by Gas Chromatography (GC)
2.9. Statistical Analysis
3. Results
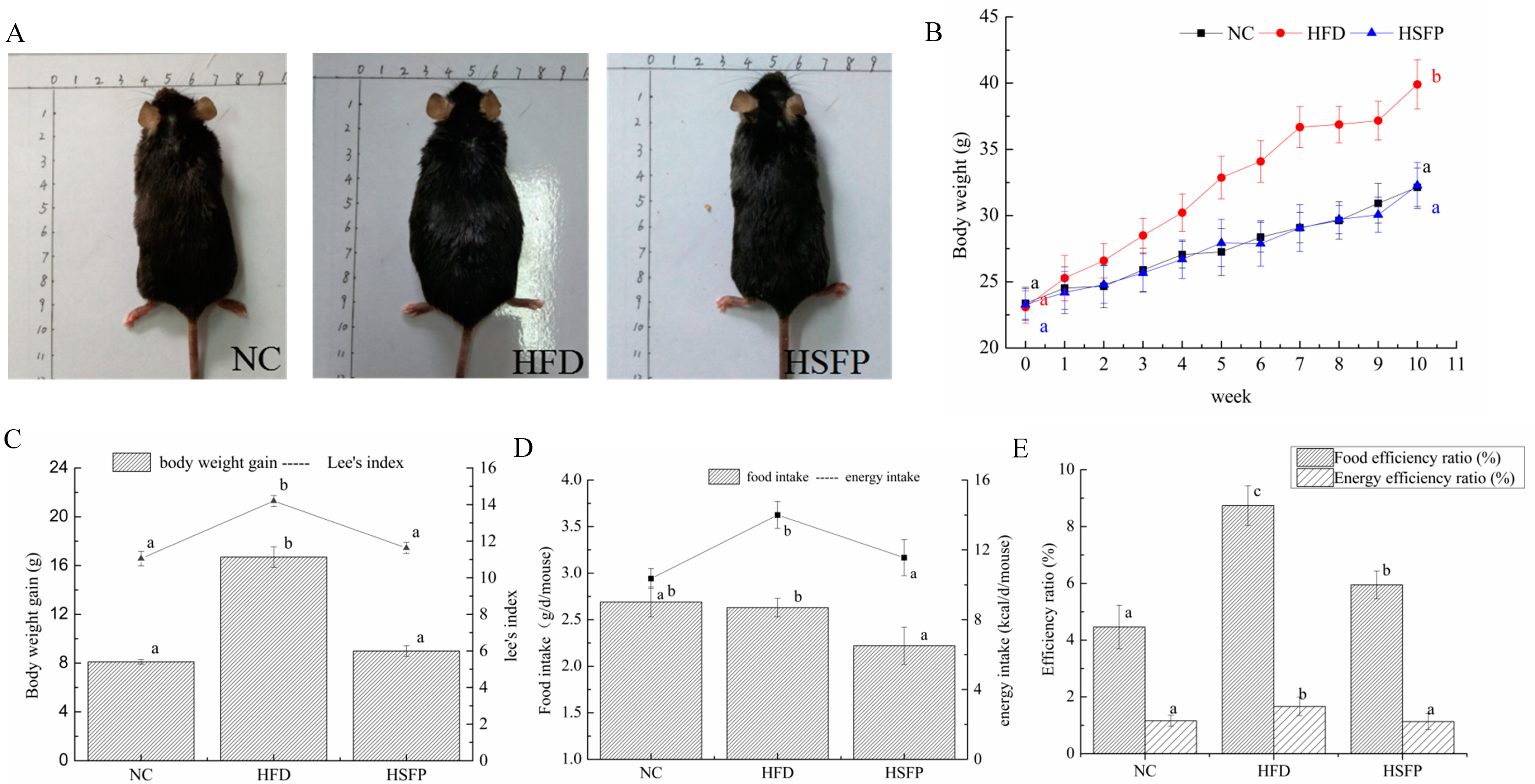
3.1. Seabuckthorn Freeze-Dried Powder Alleviated Obesity in High-Fat Diet-Induced Obese Mice
3.2. Pathological Alterations of Adipose Tissue and Liver
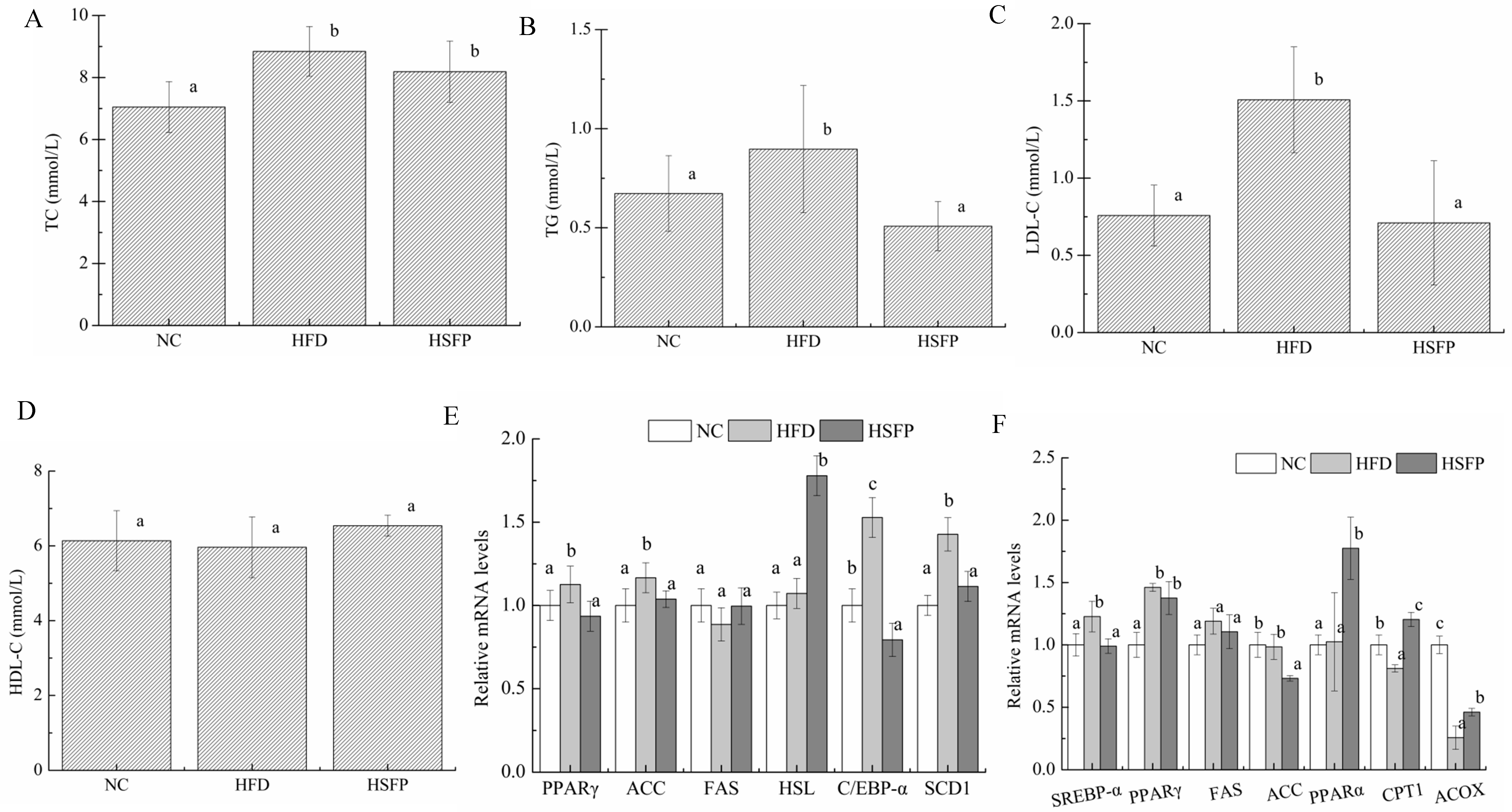
3.3. Seabuckthorn Freeze-Dried Powder Improved Metabolic Disorders in High-Fat Diet-Induced Obese Mice
3.4. Seabuckthorn Freeze-Dried Powder Regulated the Expression of Obesity-Associated Genes in the Mice
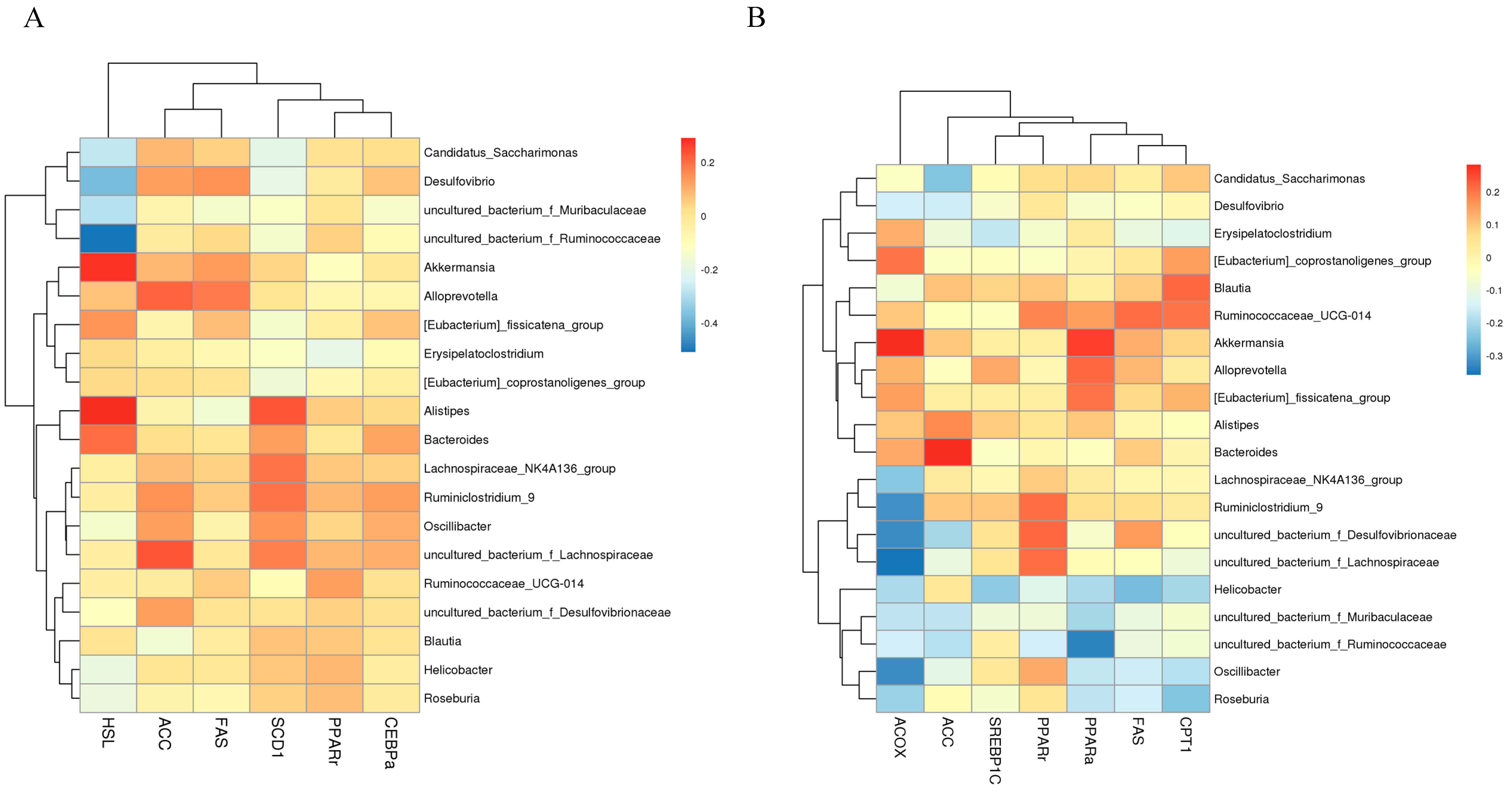
3.5. Seabuckthorn Freeze-Dried Powder Altered the Composition and Structure of the Gut Microbiota
3.6. The Effect of Seabuckthorn Freeze-Dried Powder on Gut Metabolites
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, S.; Hong, J.; Jeon, R.; Kim, H.-S. Adzuki bean ameliorates hepatic lipogenesis and proinflammatory mediator expression in mice fed a high-cholesterol and high-fat diet to induce nonalcoholic fatty liver disease. Nutr. Res. 2016, 36, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, K.; Chiu, H.-F.; Wang, C.-K. Extensive review of popular functional foods and nutraceuticals against obesity and its related complications with a special focus on randomized clinical trials. Food Funct. 2019, 10, 2313–2329. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X.; Wang, S.; Li, H.; Lu, Z.; Shi, J.; Xu, Z. Mannan-oligosaccharide modulates the obesity and gut microbiota in high-fat diet-fed mice. Food Funct. 2018, 9, 3916–3929. [Google Scholar] [CrossRef] [PubMed]
- Han, J.-L.; Lin, H.-L. Intestinal microbiota and type 2 diabetes: From mechanism insights to therapeutic perspective. World J. Gastroenterol. 2014, 20, 17737–17745. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, P.; Shou, Q.Y.; Lu, Y.H.; Wang, G.F.; Qiu, J.N.; Wang, J.; He, L.L.; Chen, J.N.; Jiao, J.J.; Zhang, Y. Arachidonic acid sex-dependently affects obesity through linking gut microbiota-driven inflammation to hypothalamus-adipose-liver axis. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2715–2726. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jin, L.; Li, Y.; Xia, G.; Chen, C.; Zhang, Y. Bamboo-shaving polysaccharide protects against high-diet induced obesity and modulates the gut microbiota of mice. J. Funct. Foods 2018, 49, 20–31. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Z.; Chen, L.; Tang, L.; Wen, S.; Liu, Y.; Yuan, J. Diet induced maternal obesity affects offspring gut microbiota and persists into young adulthood. Food Funct. 2018, 9, 4317–4327. [Google Scholar] [CrossRef]
- Anhe, F.F.; Varin, T.V.; Le Barz, M.; Pilon, G.; Dudonne, S.; Trottier, J.; St-Pierre, P.; Harris, C.S.; Lucas, M.; Lemire, M.; et al. Arctic berry extracts target the gut-liver axis to alleviate metabolic endotoxaemia, insulin resistance and hepatic steatosis in diet-induced obese mice. Diabetologia 2018, 61, 919–931. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Lu, J.; Lin, W.; Wei, X.; Li, H.; Zhao, X.; Jiang, A.; Yuan, J. Effects of a galacto-oligosaccharide-rich diet on fecal microbiota and metabolite profiles in mice. Food Funct. 2018, 9, 1612–1620. [Google Scholar] [CrossRef]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.M.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 17. [Google Scholar] [CrossRef] [Green Version]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nature 2013, 500, 585, Erratum in 2013, 502, 1. [Google Scholar] [CrossRef]
- Song, H.; Han, W.; Yan, F.; Xu, D.; Chu, Q.; Zheng, X. Dietary Phaseolus vulgaris extract alleviated diet-induced obesity, insulin resistance and hepatic steatosis and alters gut microbiota composition in mice. J. Funct. Foods 2016, 20, 236–244. [Google Scholar] [CrossRef]
- Tamchos, S.; Kaul, V. Seabuckthorn: Opportunities and Challenges in Ladakh. Natl. Acad. Sci. Lett. 2019, 42, 175–178. [Google Scholar] [CrossRef]
- Pichiah, P.B.T.; Moon, H.-J.; Park, J.-E.; Moon, Y.-J.; Cha, Y.-S. Ethanolic extract of seabuckthorn (Hippophae rhamnoides L) prevents high-fat diet-induced obesity in mice through down-regulation of adipogenic and lipogenic gene expression. Nutr. Res. 2012, 32, 856–864. [Google Scholar] [CrossRef]
- Hao, W.; He, Z.; Zhu, H.; Liu, J.; Kwek, E.; Zhao, Y.; Ma, K.Y.; He, W.-S.; Chen, Z.-Y. Sea buckthorn seed oil reduces blood cholesterol and modulates gut microbiota. Food Funct. 2019, 10, 5669–5681. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.-Y.; Lee, J.; Kim, Y.J.; Do, A.; Choi, J.-Y.; Cho, S.-J.; Jung, U.J.; Lee, M.-K.; Park, Y.B.; Choi, M.-S. Seabuckthorn Leaves Extract and Flavonoid Glycosides Extract from Seabuckthorn Leaves Ameliorates Adiposity, Hepatic Steatosis, Insulin Resistance, and Inflammation in Diet-Induced Obesity. Nutrients 2017, 9, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.; Meng, L.; Wang, W.; Zhu, X. The effects of Sea buckthorn seed protein on glucose metabolism in streptozotocin-induced diabetic ICR mice. Pak. J. Pharm. Sci. 2019, 32, 2011–2017. [Google Scholar] [PubMed]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.-M.; Zhao, S.; Feng, F.; Liu, Q.; Zhou, X.-L.; Xiao, Y. Modulatory Effect of Set Yogurt with Tartary Buckwheat Juice on Intestinal Flora in Mice. Food Sci. 2019, 40, 123–129. [Google Scholar] [CrossRef]
- Wei, X.-X.; Chen, Q.-S.; Yan, Y.-L.; Zhao, P. Effect of Konjac Glucomannans on Short-Chain Fatty Acids in Cecal Contents of Mice. Food Sci. 2014, 35, 214–219. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, W.; Li, X.; Xin, X.; Liu, D. Daily Supplementation with Fresh Angelica keiskei Juice Alleviates High-Fat Diet-Induced Obesity in Mice by Modulating Gut Microbiota Composition. Mol. Nutr. Food Res. 2019, 63, 1900248. [Google Scholar] [CrossRef]
- Song, H.; Lai, J.; Tang, Q.; Zheng, X. Mulberry ethanol extract attenuates hepatic steatosis and insulin resistance in high-fat diet-fed mice. Nutr. Res. 2016, 36, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-C.; Chang, C.-S.; Chou, C.-H.; Wu, Y.-H.S.; Yang, K.-T.; Tseng, J.-K.; Chang, Y.-Y.; Chen, Y.-C. L-carnitine ameliorates dyslipidemic and hepatic disorders induced by a high-fat diet via regulating lipid metabolism, self-antioxidant capacity, and inflammatory response. J. Funct. Foods 2015, 15, 497–508. [Google Scholar] [CrossRef]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Bloc’h, J.L.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 3, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Lee, S.-J.; Yu, H.-J.; Lee, J.-H.; Cho, H.-Y. Fermentation with Lactobacillus enhances the preventive effect of garlic extract on high fat diet-induced hepatic steatosis in mice. J. Funct. Foods 2017, 30, 125–133. [Google Scholar] [CrossRef]
- Yang, Y.; Li, W.; Liu, Y.; Sun, Y.N.; Li, Y.; Yao, Q.; Li, J.N.; Zhang, Q.; Gao, Y.J.; Gao, L.; et al. Alpha-lipoic acid improves high-fat diet-induced hepatic steatosis by modulating the transcription factors SREBP-1, FoxO1 and Nrf2 via the SIRT1/LKB1/AMPK pathway. J. Nutr. Biochem. 2014, 25, 1207–1217. [Google Scholar] [CrossRef] [Green Version]
- Zang, Y.; Zhang, L.; Igarashi, K.; Yu, C. The anti-obesity and anti-diabetic effects of kaempferol glycosides from unripe soybean leaves in high-fat-diet mice. Food Funct. 2015, 6, 834–841. [Google Scholar] [CrossRef]
- Liu, F.-J.; Liang, D.; Miao, L.-Y.; Li, P.; Li, H.-J. Liver-specific metabolomics characterizes the hepatoprotective effect of saponin-enriched Celosiae Semen extract on mice with nonalcoholic fatty liver disease. J. Funct. Foods 2018, 42, 185–194. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Y.; Song, P.; Wang, H.; Xu, N.; Wang, Y.; Zhang, Z.; Yue, P.; Gao, X. Anti-obesity effects of instant fermented teas in vitro and in mice with high-fat-diet-induced obesity. Food Funct. 2019, 10, 3502–3513. [Google Scholar] [CrossRef]
- Kim, D.-H.; Kim, H.; Jeong, D.; Kang, I.-B.; Chon, J.-W.; Kim, H.-S.; Song, K.-Y.; Seo, K.-H. Kefir alleviates obesity and hepatic steatosis in high-fat diet-fed mice by modulation of gut microbiota and mycobiota: Targeted and untargeted community analysis with correlation of biomarkers. J. Nutr. Biochem. 2017, 44, 35–43. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Guo, C.; Qiao, J.; Zhang, S.; Li, M.; Li, J.; Hatab, S. Elaeagnus mollis Oil Attenuates Non-alcoholic Fatty Disease in High-fat Diet Induced Obese Mice via Modifying the Expression of Lipid Metabolism-related Genes. J. Oleo Sci. 2019, 68, 893–908. [Google Scholar] [CrossRef] [Green Version]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- Tung, Y.-C.; Chang, W.-T.; Li, S.; Wu, J.-C.; Badmeav, V.; Ho, C.-T.; Pan, M.-H. Citrus peel extracts attenuated obesity and modulated gut microbiota in mice with high-fat diet-induced obesity. Food Funct. 2018, 9, 3363–3373. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, J.; Hong, F.; Wang, S.; Jin, X.; Xue, T.; Jia, L.; Zhai, Y. Melatonin prevents obesity through modulation of gut microbiota in mice. J. Pineal Res. 2017, 62, e12399. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Gao, J.; Du, M.; Mao, X. Bovine α-lactalbumin hydrolysates ameliorate obesity-associated endotoxemia and inflammation in high-fat diet-fed mice through modulation of gut microbiota. Food Funct. 2019, 10, 3368–3378. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Anhe, F.F.; Roy, D.; Pilon, G.; Dudonne, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [Green Version]
- Backhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [Green Version]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content |
|---|---|
| Total polyphenols (mg/g) | 6.55 ± 0.29 |
| Total sugar (g/100 g) | 22.83 ± 0.44 |
| Total acid (malic acid as a standard) % | 14.38 ± 0.62 |
| Soluble solid (°Brix) | 62.54 ± 5.30 |
| Soluble dietary fiber (g/100 g) | 26.15 ± 0.53 |
| Insoluble dietary fiber (g/100 g) | 1.51 ± 0.10 |
| Vitamin C (mg/100 g) | 23.24 ± 0.51 |
| Gene | Primer Sequences | Tm | Sequence Number | Product PCR (bp) | |
|---|---|---|---|---|---|
| PPARγ | Forward | TTTTCAAGGGTGCCAGTTTCGATCC | 61 | NM_001127330.1 | 198 |
| Reverse | AATCCTTGGCCCTCTGAGAT | 58 | |||
| ACC | Forward | GGCCAGTGCTATGCTGAGAT | 59 | NM_133360.2 | 108 |
| Reverse | AGGGTCAAGTGCTGCTCCA | 59 | |||
| FAS | Forward | GCTGCGGAAACTTCAGGAAAT | 57 | NM_007988.3 | 84 |
| Reverse | AGAGACGTGTCACTCCTGGACTT | 61 | |||
| SREBP-1c | Forward | CACAGCGGTTTTGAACGACA | 58 | NM_011480.3 | 147 |
| Reverse | CTCTCAGGAGAGTTGGCACC | 58 | |||
| HSL | Forward | CTGGAACTAAGTGGACGCAAG | 56 | NM_001039507 | 91 |
| Reverse | CAGACACACTCCTGCGCATAGAC | 60 | |||
| C/EBP-α | Forward | TCGGTGCGTCTAAGATGAGG | 57 | NM_001287523 | 179 |
| Reverse | TCAAGGCACATTTTTGCTCC | 55 | |||
| SCD1 | Forward | AGAGAACTGGAGACGGGAGT | 57 | NM_009127 | 130 |
| Reverse | AACACCCCGATAGCAATATCCA | 57 | |||
| PPARα | Forward | CAAGGCCTCAGGGTACCACT | 59 | NM_001113418.1 | 111 |
| Reverse | TTGCAGCTCCGATCACACTT | 58 | |||
| CPT-1 | Forward | CTCAGTGGGAGCGACTCTTCA | 59 | NM_013495.2 | 105 |
| Reverse | GGCCTCTGTGGTACACGACAA | 60 | |||
| ACOX | Forward | GCCTTTGTTGTCCCTATCCG | 58 | NM_001271898.1 | 189 |
| Reverse | TACATACGTGCCGTCAGGC | 58 | |||
| β-actin | Forward | CAGGCATTGCTGACAGGATG | 58 | NM_007393 | 156 |
| Reverse | TGCTGATCCACATCTGCTGG | 58 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, C.; Han, L.; Li, M.; Yu, L. Seabuckthorn (Hippophaë rhamnoides) Freeze-Dried Powder Protects against High-Fat Diet-Induced Obesity, Lipid Metabolism Disorders by Modulating the Gut Microbiota of Mice. Nutrients 2020, 12, 265. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12010265
Guo C, Han L, Li M, Yu L. Seabuckthorn (Hippophaë rhamnoides) Freeze-Dried Powder Protects against High-Fat Diet-Induced Obesity, Lipid Metabolism Disorders by Modulating the Gut Microbiota of Mice. Nutrients. 2020; 12(1):265. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12010265
Chicago/Turabian StyleGuo, Caixia, Li Han, Meiping Li, and Ligang Yu. 2020. "Seabuckthorn (Hippophaë rhamnoides) Freeze-Dried Powder Protects against High-Fat Diet-Induced Obesity, Lipid Metabolism Disorders by Modulating the Gut Microbiota of Mice" Nutrients 12, no. 1: 265. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12010265