Physiologic Effects of Exogenous Dextrose in Murine Klebsiella pneumoniae Sepsis Vary by Route of Provision

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
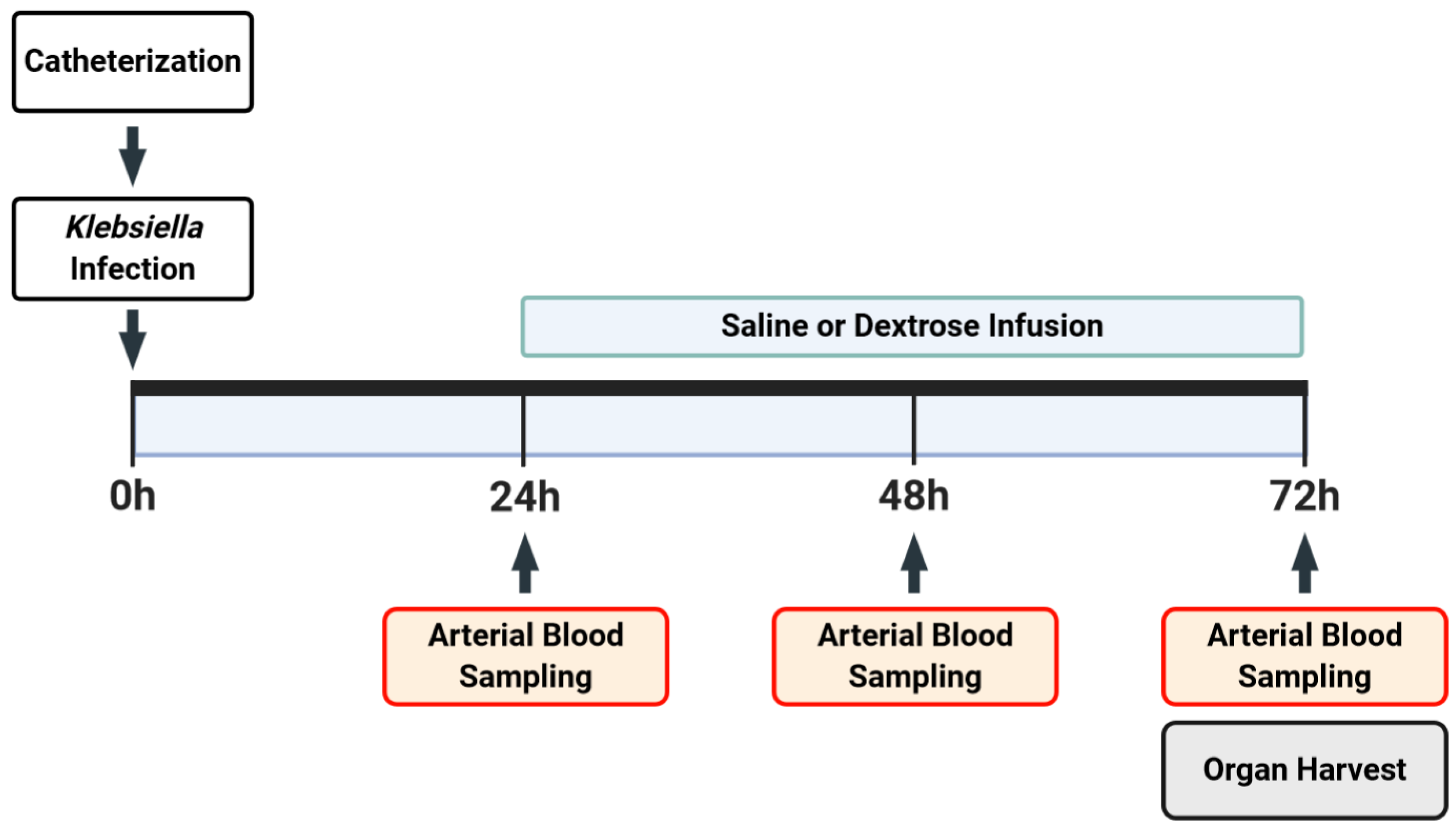
2.2. Experimental Model
2.3. Bronchoalveolar Lavage (BAL)
2.4. Assessment of Bacterial Burden
2.5. Quantification of Hemodynamic Data
2.6. Biochemical Assays
2.7. Lung Injury Scoring
2.8. Statistical Analysis
3. Results
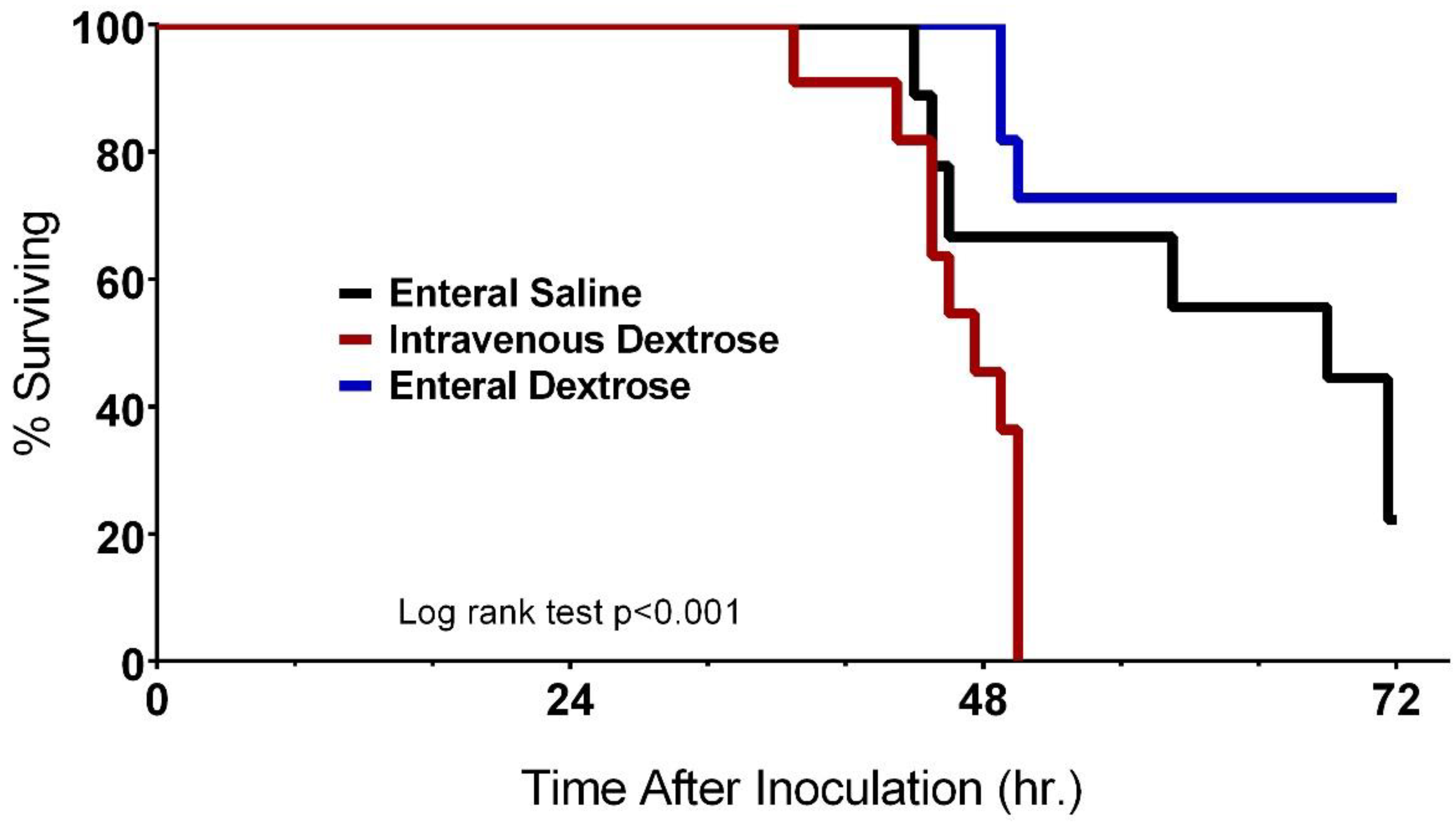
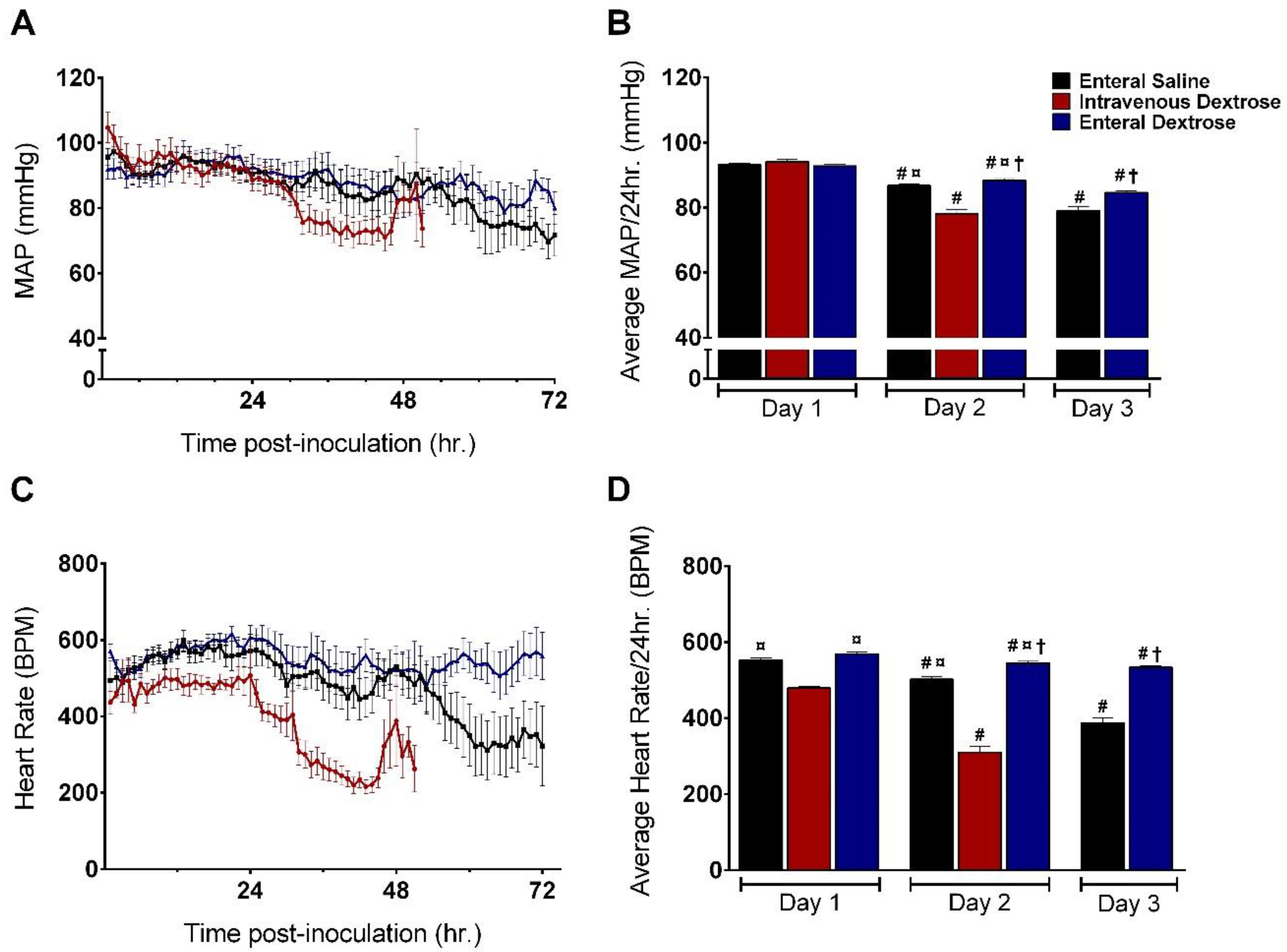
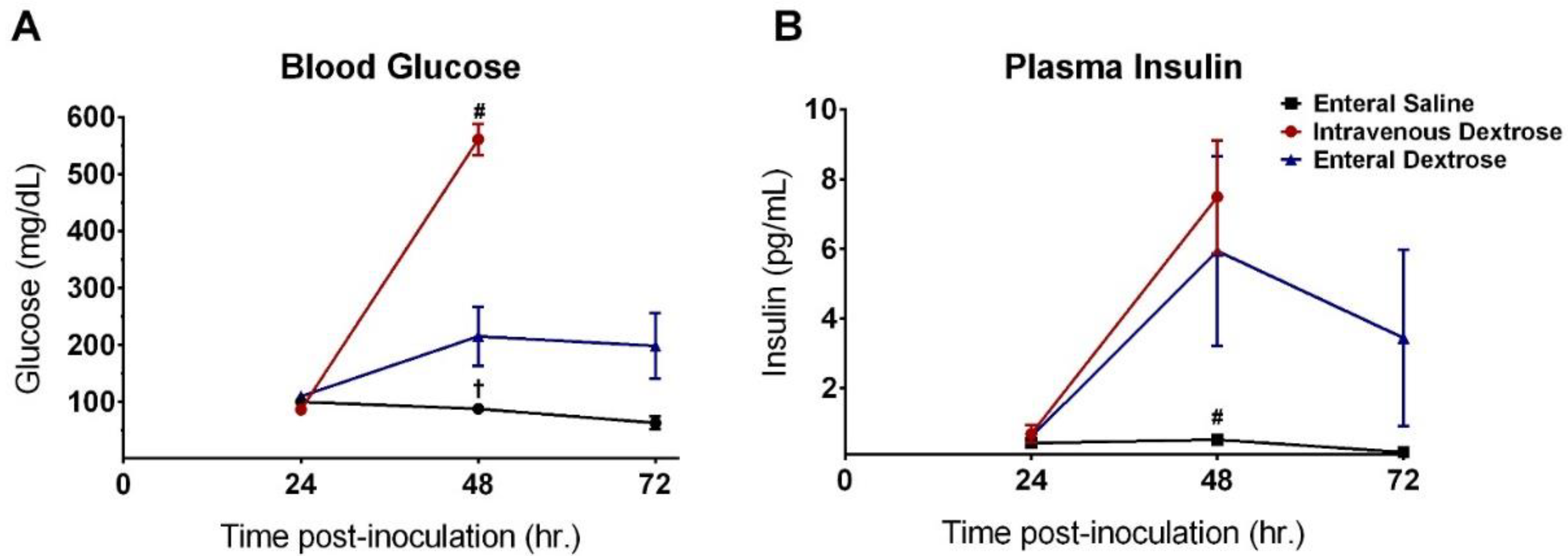
3.1. Physiologic Effects of Early Dextrose Infusion Vary by Route of Administration in a KP Model of Sepsis
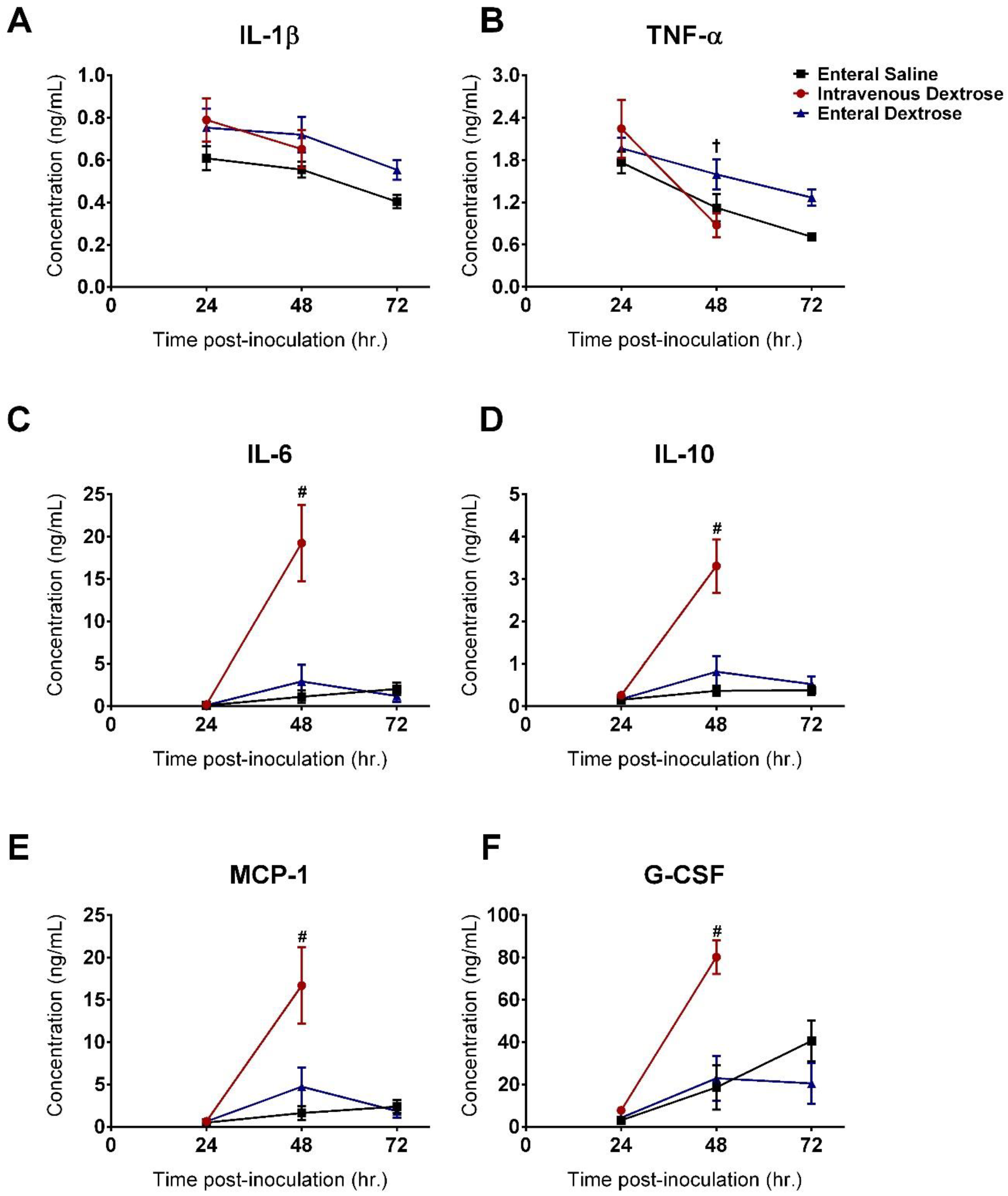
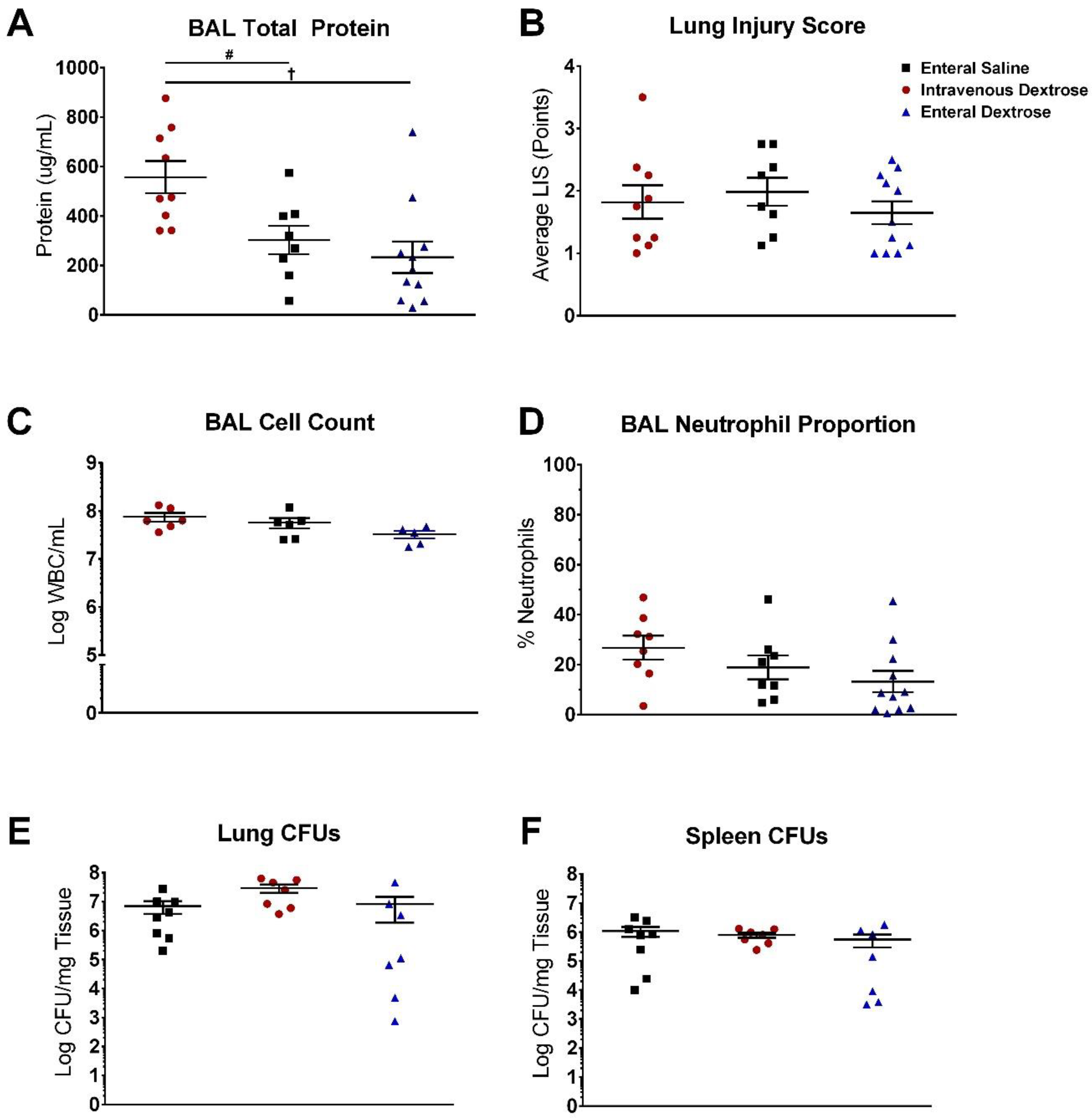
3.2. IV Dextrose Worsens Inflammation and Lung Permeability in a KP Model of Sepsis
3.3. Exogenous Dextrose Does Not Affect Bacterial Burden in a KP Model of Sepsis
3.4. Improvement in Survival with Enteral Dextrose Is Associated with Preservation of Euglycemia
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Callahan, L.A.; Supinski, G.S. Sepsis-induced myopathy. Crit. Care Med. 2009, 37, S354–S367. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, S.N.; Kennedy, D.D.; Ghosh, I.R.; Misra, V.P.; Kiff, K.; Coakley, J.H.; Hinds, C.J. Persistent neuromuscular and neurophysiologic abnormalities in long-term survivors of prolonged critical illness*. Crit. Care Med. 2003, 31, 1012–1016. [Google Scholar] [CrossRef] [PubMed]
- Dinglas, V.D.; Aronson Friedman, L.; Colantuoni, E.; Mendez-Tellez, P.A.; Shanholtz, C.B.; Ciesla, N.D.; Pronovost, P.J.; Needham, D.M. Muscle Weakness and 5-Year Survival in Acute Respiratory Distress Syndrome Survivors. Crit. Care Med. 2017, 45, 446–453. [Google Scholar] [CrossRef] [Green Version]
- Wischmeyer, P.E. Nutrition Therapy in Sepsis. Crit. Care Clin. 2018, 34, 107–125. [Google Scholar] [CrossRef]
- Elke, G.; van Zanten, A.R.H.; Lemieux, M.; McCall, M.; Jeejeebhoy, K.N.; Kott, M.; Jiang, X.; Day, A.G.; Heyland, D.K. Enteral versus parenteral nutrition in critically ill patients: An updated systematic review and meta-analysis of randomized controlled trials. Crit. Care 2016, 20, 117. [Google Scholar] [CrossRef] [Green Version]
- Reignier, J.; Boisramé-Helms, J.; Brisard, L.; Lascarrou, J.-B.; Ait Hssain, A.; Anguel, N.; Argaud, L.; Asehnoune, K.; Asfar, P.; Bellec, F.; et al. Enteral versus parenteral early nutrition in ventilated adults with shock: A randomised, controlled, multicentre, open-label, parallel-group study (NUTRIREA-2). Lancet 2018, 391, 133–143. [Google Scholar] [CrossRef]
- Alberda, C.; Gramlich, L.; Jones, N.; Jeejeebhoy, K.; Day, A.G.; Dhaliwal, R.; Heyland, D.K. The relationship between nutritional intake and clinical outcomes in critically ill patients: Results of an international multicenter observational study. Intensive Care Med. 2009, 35, 1728–1737. [Google Scholar] [CrossRef]
- Wei, X.; Day, A.G.; Ouellette-Kuntz, H.; Heyland, D.K. The Association Between Nutritional Adequacy and Long-Term Outcomes in Critically Ill Patients Requiring Prolonged Mechanical Ventilation: A Multicenter Cohort Study. Crit. Care Med. 2015, 43, 1569–1579. [Google Scholar] [CrossRef]
- Lin, L.Y.; Lin, H.C.; Lee, P.C.; Ma, W.Y.; Lin, H.D. Hyperglycemia correlates with outcomes in patients receiving total parenteral nutrition. Am. J. Med. Sci. 2007, 333, 261–265. [Google Scholar] [CrossRef]
- Pasquel, F.J.; Spiegelman, R.; McCauley, M.; Smiley, D.; Umpierrez, D.; Johnson, R.; Rhee, M.; Gatcliffe, C.; Lin, E.; Umpierrez, E.; et al. Hyperglycemia during total parenteral nutrition: An important marker of poor outcome and mortality in hospitalized patients. Diabetes Care 2010, 33, 739–741. [Google Scholar] [CrossRef] [Green Version]
- Pleva, M.; Mirtallo, J.M.; Steinberg, S.M. Hyperglycemic Events in Non–Intensive Care Unit Patients Receiving Parenteral Nutrition. Nutr. Clin. Pract. 2009, 24, 626–634. [Google Scholar] [CrossRef]
- Pancorbo-Hidalgo, P.L.; García-Fernandez, F.P.; Ramírez-Pérez, C. Complications associated with enteral nutrition by nasogastric tube in an internal medicine unit. J. Clin. Nurs. 2001, 10, 482–490. [Google Scholar] [CrossRef]
- De Waele, E.; Malbrain, M.L.N.G.; Spapen, H. Nutrition in Sepsis: A Bench-to-Bedside Review. Nutrients 2020, 12, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montejo, J.C. Enteral nutrition-related gastrointestinal complications in critically ill patients: A multicenter study. Crit. Care Med. 1999, 27, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Kollef, M.H. Ventilator-associated pneumonia. A multivariate analysis. JAMA 1993, 270, 1965–1970. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, R.N.; Boschert, K.J.; Kudsk, K.A.; Brown, R.O. Hypocaloric enteral tube feeding in critically ill obese patients. Nutrition 2002, 18, 241–246. [Google Scholar] [CrossRef]
- Krishnan, J.A.; Parce, P.B.; Martinez, A.; Diette, G.B.; Brower, R.G. Caloric Intake in Medical ICU Patients: Consistency of Care with Guidelines and Relationship to Clinical Outcomes. Chest 2003, 124, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Hadfield, R.J.; Sinclair, D.G.; Houldsworth, P.E.; Evans, T.W. Effects of enteral and parenteral nutrition on gut mucosal permeability in the critically ill. Am. J. Respir. Crit. Care Med. 1995, 152, 1545–1548. [Google Scholar] [CrossRef]
- van Niekerk, G.; Meaker, C.; Engelbrecht, A.-M. Nutritional support in sepsis: When less may be more. Crit. Care 2020, 24, 53. [Google Scholar] [CrossRef] [Green Version]
- McClave, S.A.; Taylor, B.E.; Martindale, R.G.; Warren, M.M.; Johnson, D.R.; Braunschweig, C.; McCarthy, M.S.; Davanos, E.; Rice, T.W.; Cresci, G.A.; et al. Guidelines for the Provision and Assessment of Nutrition Support Therapy in the Adult Critically Ill Patient. J. Parenter. Enter. Nutr. 2016, 40, 159–211. [Google Scholar] [CrossRef] [PubMed]
- Peter, J.V.; Moran, J.L.; Phillips-Hughes, J. A metaanalysis of treatment outcomes of early enteral versus early parenteral nutrition in hospitalized patients. Crit. Care Med. 2005, 33, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.R.; Schofield-Robinson, O.J.; Alderson, P.; Smith, A.F. Enteral versus parenteral nutrition and enteral versus a combination of enteral and parenteral nutrition for adults in the intensive care unit. Cochrane Database Syst. Rev. 2018, 6, CD012276. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.A.; Singamsetty, S.; Guo, L.; Chuan, B.W.; McDonald, S.; Cooper, B.A.; O’Donnell, B.J.; Stefanovski, D.; Wice, B.; Zhang, Y.; et al. Stimulation of the endogenous incretin glucose-dependent insulinotropic peptide by enteral dextrose improves glucose homeostasis and inflammation in murine endotoxemia. Transl. Res. 2018, 193, 1–12. [Google Scholar] [CrossRef]
- Denstaedt, S.J.; Spencer-Segal, J.L.; Newstead, M.; Laborc, K.; Zeng, X.; Standiford, T.J.; Singer, B.H. Persistent Neuroinflammation and Brain-Specific Immune Priming in a Novel Survival Model of Murine Pneumosepsis. Shock 2020, 54, 78–86. [Google Scholar] [CrossRef]
- Watanabe, Y.; Singamsetty, S.; Zou, B.; Guo, L.; Stefanovski, D.; Alonso, L.C.; Garcia-Ocana, A.; O’Donnell, C.P.; McVerry, B.J. Exogenous Glucose Administration Impairs Glucose Tolerance and Pancreatic Insulin Secretion during Acute Sepsis in Non-Diabetic Mice. PLoS ONE 2013, 8, e67716. [Google Scholar] [CrossRef] [Green Version]
- Singamsetty, S.; Shah, F.A.; Guo, L.; Watanabe, Y.; McDonald, S.; Sharma, R.; Zhang, Y.; Alonso, L.C.; O’Donnell, C.P.; McVerry, B.J. Early initiation of low-level parenteral dextrose induces an accelerated diabetic phenotype in septic C57BL/6J mice. Appl. Physiol. Nutr. Metab. 2016, 41, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Robinson, K.M.; Lee, B.; Scheller, E.V.; Mandalapu, S.; Enelow, R.I.; Kolls, J.K.; Alcorn, J.F. The role of IL-27 in susceptibility to post-influenza Staphylococcus aureus pneumonia. Respir. Res. 2015, 16, 10. [Google Scholar] [CrossRef] [Green Version]
- Chambrier, C.; Laville, M.; Berrada, K.R.; Odeon, M.; BoulÉTreau, P.; Beylot, M. Insulin sensitivity of glucose and fat metabolism in severe sepsis. Clin. Sci. 2000, 99, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Marik, P.E.; Raghavan, M. Stress-hyperglycemia, insulin and immunomodulation in sepsis. Intensive Care Med. 2004, 30, 748–756. [Google Scholar] [CrossRef]
- Van Cromphaut, S.J.; Vanhorebeek, I.; Van den Berghe, G. Glucose metabolism and insulin resistance in sepsis. Curr. Pharm. Des. 2008, 14, 1887–1899. [Google Scholar] [CrossRef]
- van Oss, C.J.; Border, J.R. Influence of intermittent hyperglycemic glucose levels on the phagocytosis of microorganisms by human granulocytes in vitro. Immunol. Commun. 1978, 7, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Jafar, N.; Edriss, H.; Nugent, K. The Effect of Short-Term Hyperglycemia on the Innate Immune System. Am. J. Med. Sci. 2016, 351, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Siu, L.K.; Fung, C.P.; Tsou, H.H.; Wang, J.J.; Chen, C.T.; Wang, S.C.; Chang, F.Y. Impaired phagocytosis of capsular serotypes K1 or K2 Klebsiella pneumoniae in type 2 diabetes mellitus patients with poor glycemic control. J. Clin. Endocrinol. Metab. 2006, 91, 3084–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krinsley, J.S. Association between hyperglycemia and increased hospital mortality in a heterogeneous population of critically ill patients. Mayo Clin. Proc. 2003, 78, 1471–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotchkiss, R.S.; Moldawer, L.L.; Opal, S.M.; Reinhart, K.; Turnbull, I.R.; Vincent, J.-L. Sepsis and septic shock. Nat. Rev. Dis. Primers 2016, 2, 16045. [Google Scholar] [CrossRef] [Green Version]
- Esposito, K.; Nappo, F.; Marfella, R.; Giugliano, G.; Giugliano, F.; Ciotola, M.; Quagliaro, L.; Ceriello, A.; Giugliano, D. Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: Role of oxidative stress. Circulation 2002, 106, 2067–2072. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, N.; Reddy, M.A.; Guha, M.; Natarajan, R. High glucose-induced expression of proinflammatory cytokine and chemokine genes in monocytic cells. Diabetes 2003, 52, 1256–1264. [Google Scholar] [CrossRef] [Green Version]
- Arabi, Y.M.; Dabbagh, O.C.; Tamim, H.M.; Al-Shimemeri, A.A.; Memish, Z.A.; Haddad, S.H.; Syed, S.J.; Giridhar, H.R.; Rishu, A.H.; Al-Daker, M.O.; et al. Intensive versus conventional insulin therapy: A randomized controlled trial in medical and surgical critically ill patients. Crit. Care Med. 2008, 36, 3190–3197. [Google Scholar] [CrossRef]
- De La Rosa, G.D.C.; Donado, J.H.; Restrepo, A.H.; Quintero, A.M.; González, L.G.; Saldarriaga, N.E.; Bedoya, M.; Toro, J.M.; Velásquez, J.B.; Valencia, J.C.; et al. Strict glycaemic control in patients hospitalised in a mixed medical and surgical intensive care unit: A randomised clinical trial. Crit. Care 2008, 12, R120. [Google Scholar] [CrossRef] [Green Version]
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [Green Version]
- Weis, S.; Carlos, A.R.; Moita, M.R.; Singh, S.; Blankenhaus, B.; Cardoso, S.; Larsen, R.; Rebelo, S.; Schäuble, S.; Del Barrio, L.; et al. Metabolic Adaptation Establishes Disease Tolerance to Sepsis. Cell 2017, 169, 1263–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, M.; Demehri, F.R.; Teitelbaum, D.H. Intestine, immunity, and parenteral nutrition in an era of preferred enteral feeding. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 496–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunschweig, C.L.; Levy, P.; Sheean, P.M.; Wang, X. Enteral compared with parenteral nutrition: A meta-analysis. Am. J. Clin. Nutr. 2001, 74, 534–542. [Google Scholar] [CrossRef] [Green Version]
- Gramlich, L.; Kichian, K.; Pinilla, J.; Rodych, N.J.; Dhaliwal, R.; Heyland, D.K. Does enteral nutrition compared to parenteral nutrition result in better outcomes in critically ill adult patients? A systematic review of the literature. Nutrition 2004, 20, 843–848. [Google Scholar] [CrossRef]
- Gungabissoon, U.; Hacquoil, K.; Bains, C.; Irizarry, M.; Dukes, G.; Williamson, R.; Deane, A.M.; Heyland, D.K. Prevalence, risk factors, clinical consequences, and treatment of enteral feed intolerance during critical illness. JPEN J. Parenter. Enteral. Nutr. 2015, 39, 441–448. [Google Scholar] [CrossRef] [PubMed]
- McClave, S.A.; Heyland, D.K. The physiologic response and associated clinical benefits from provision of early enteral nutrition. Nutr. Clin. Pract. 2009, 24, 305–315. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Saline Infused | Dextrose Infused | ||||
|---|---|---|---|---|---|
| Euglycemic | Not Euglycemic | Euglycemic | Not Euglycemic | ||
| Survived | 2 | 0 | Survived | 7 | 1 |
| Died | 0 | 7 | Died | 0 | 14 |
| p-value | 0.028 | p-value | <0.001 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuan, B.; Guo, L.; Cooper, B.; Rawal, S.; Gallego-Martin, T.; Zhang, Y.; McVerry, B.J.; O’Donnell, C.P.; Shah, F.A. Physiologic Effects of Exogenous Dextrose in Murine Klebsiella pneumoniae Sepsis Vary by Route of Provision. Nutrients 2020, 12, 2901. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12102901
Chuan B, Guo L, Cooper B, Rawal S, Gallego-Martin T, Zhang Y, McVerry BJ, O’Donnell CP, Shah FA. Physiologic Effects of Exogenous Dextrose in Murine Klebsiella pneumoniae Sepsis Vary by Route of Provision. Nutrients. 2020; 12(10):2901. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12102901
Chicago/Turabian StyleChuan, Byron, Lanping Guo, Bryce Cooper, Sagar Rawal, Teresa Gallego-Martin, Yingze Zhang, Bryan J. McVerry, Christopher P. O’Donnell, and Faraaz Ali Shah. 2020. "Physiologic Effects of Exogenous Dextrose in Murine Klebsiella pneumoniae Sepsis Vary by Route of Provision" Nutrients 12, no. 10: 2901. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12102901