Effect of a Synbiotic Containing Lactobacillus paracasei and Opuntia humifusa on a Murine Model of Irritable Bowel Syndrome

 , , , and
, , , and 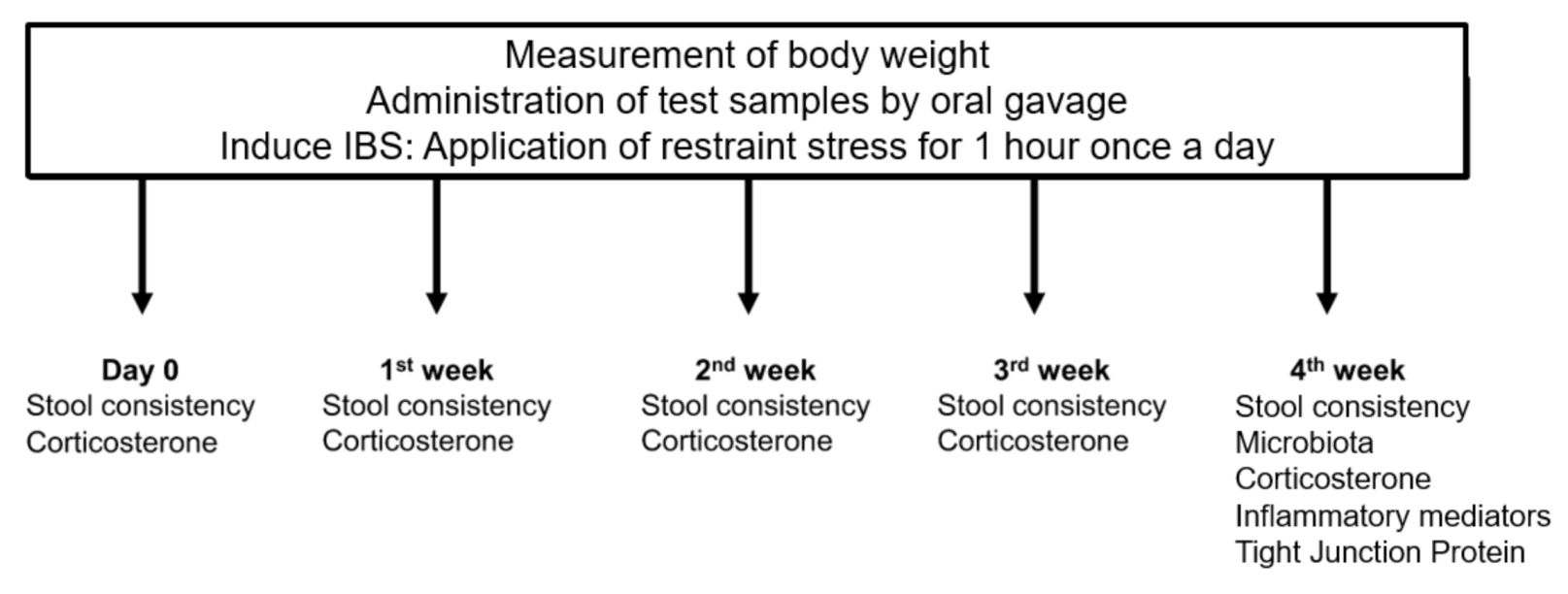{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Bacterial Strains and Materials
2.2. Rats and Study Design
2.3. Fecal Samples and Microbial Analysis
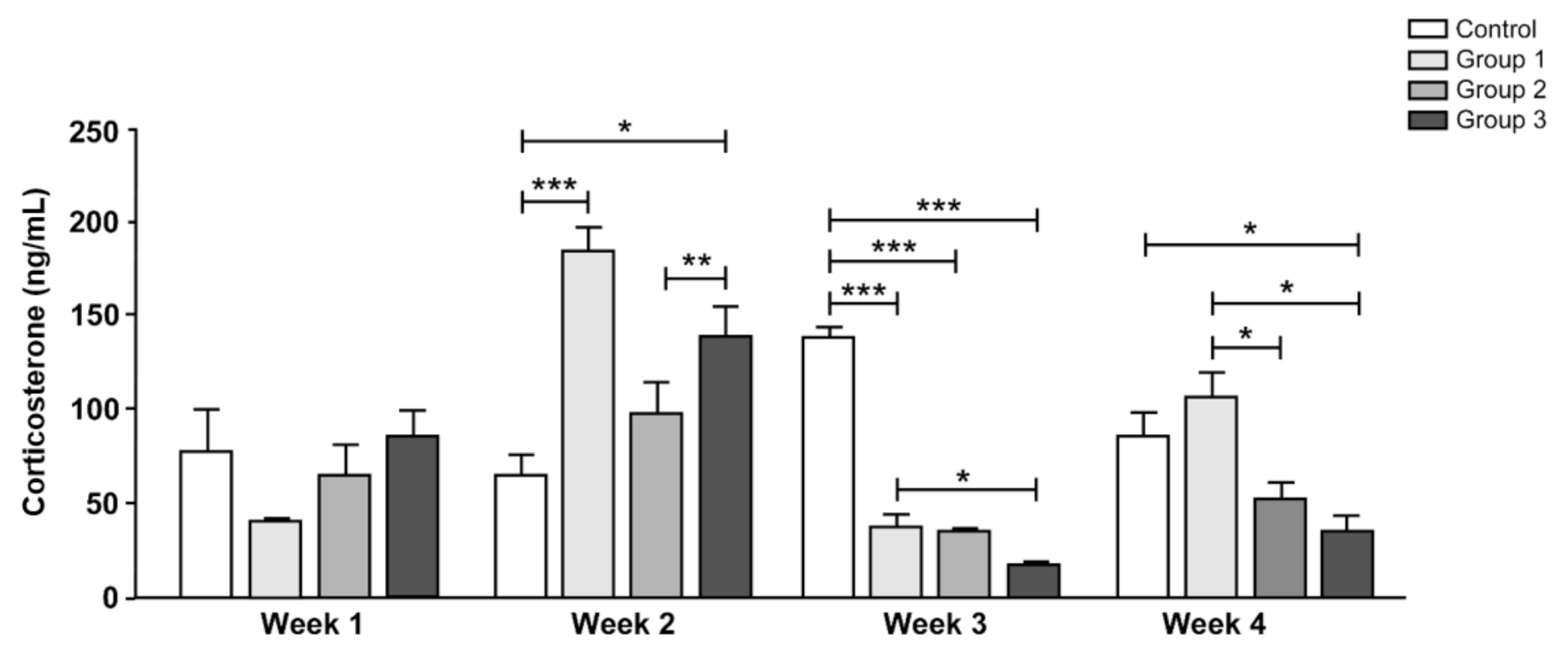
2.4. Serum Corticosterone
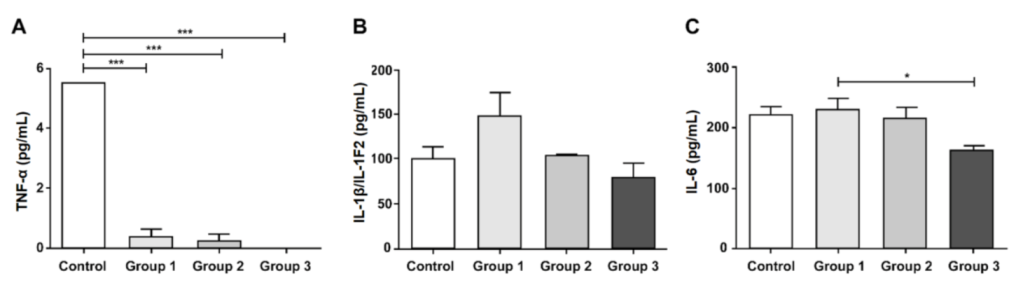
2.5. Assessment of Inflammatory Mediators
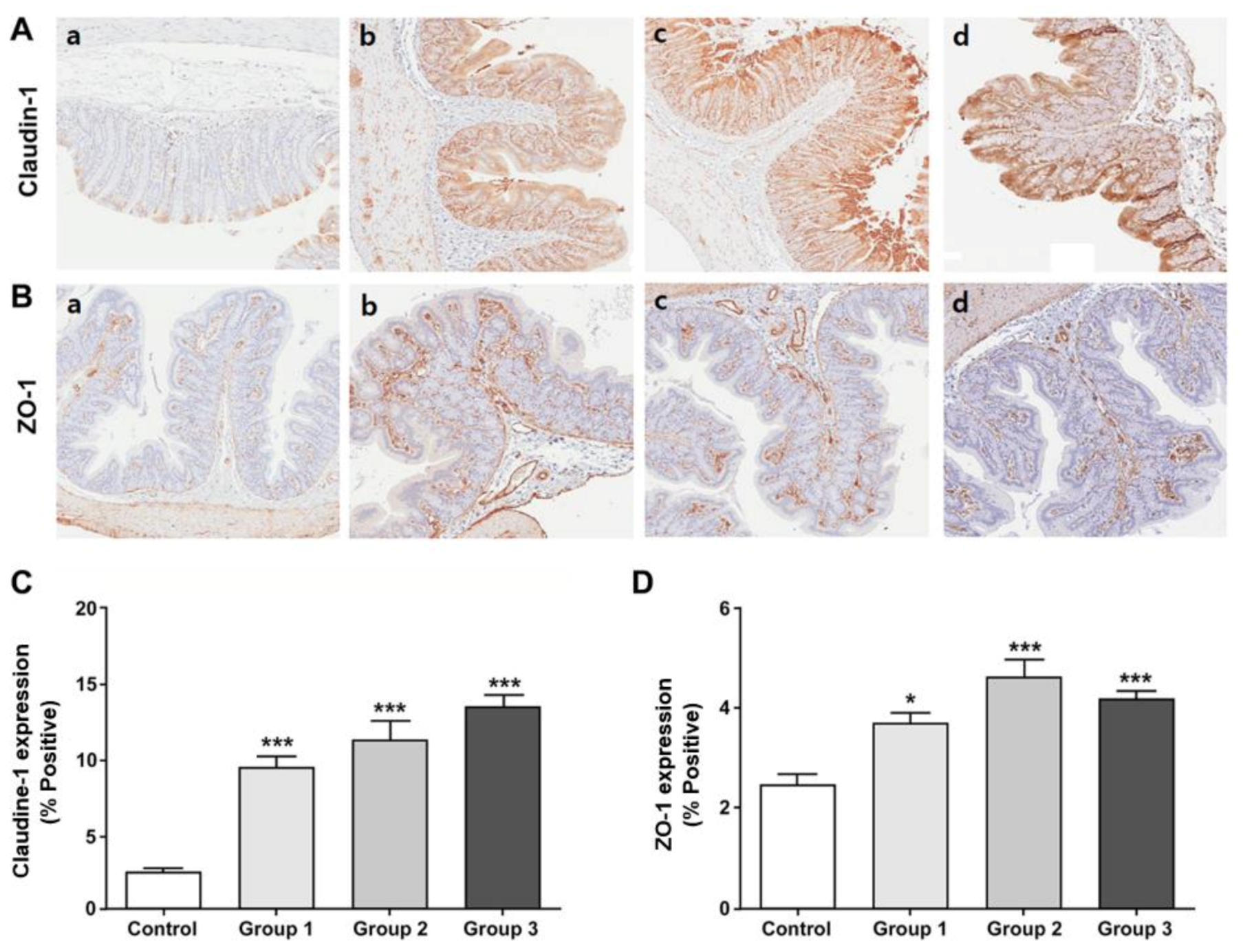
2.6. Tight Junction Proteins
2.7. Statistical Analysis
3. Results
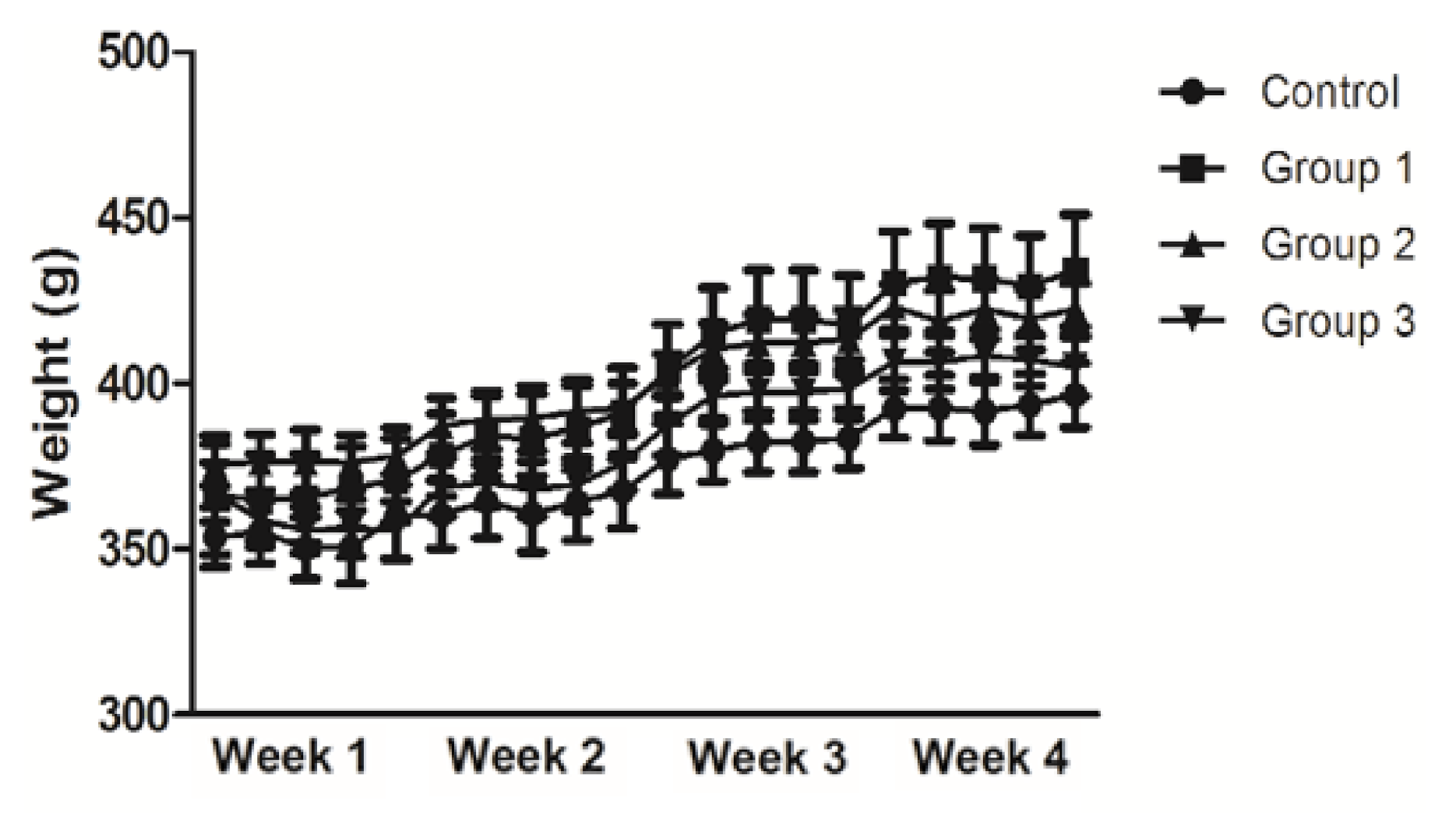
3.1. Analysis of Body Weight
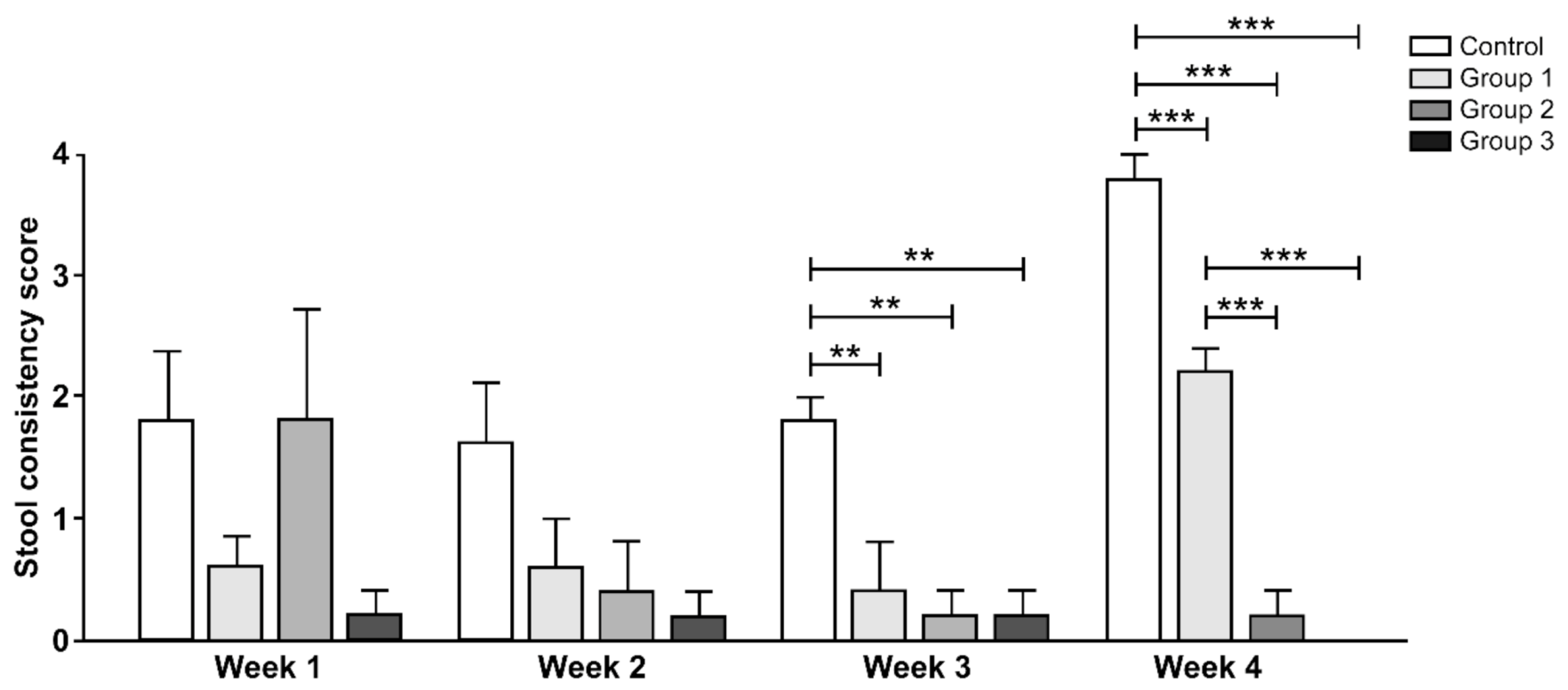
3.2. Stool Consistency
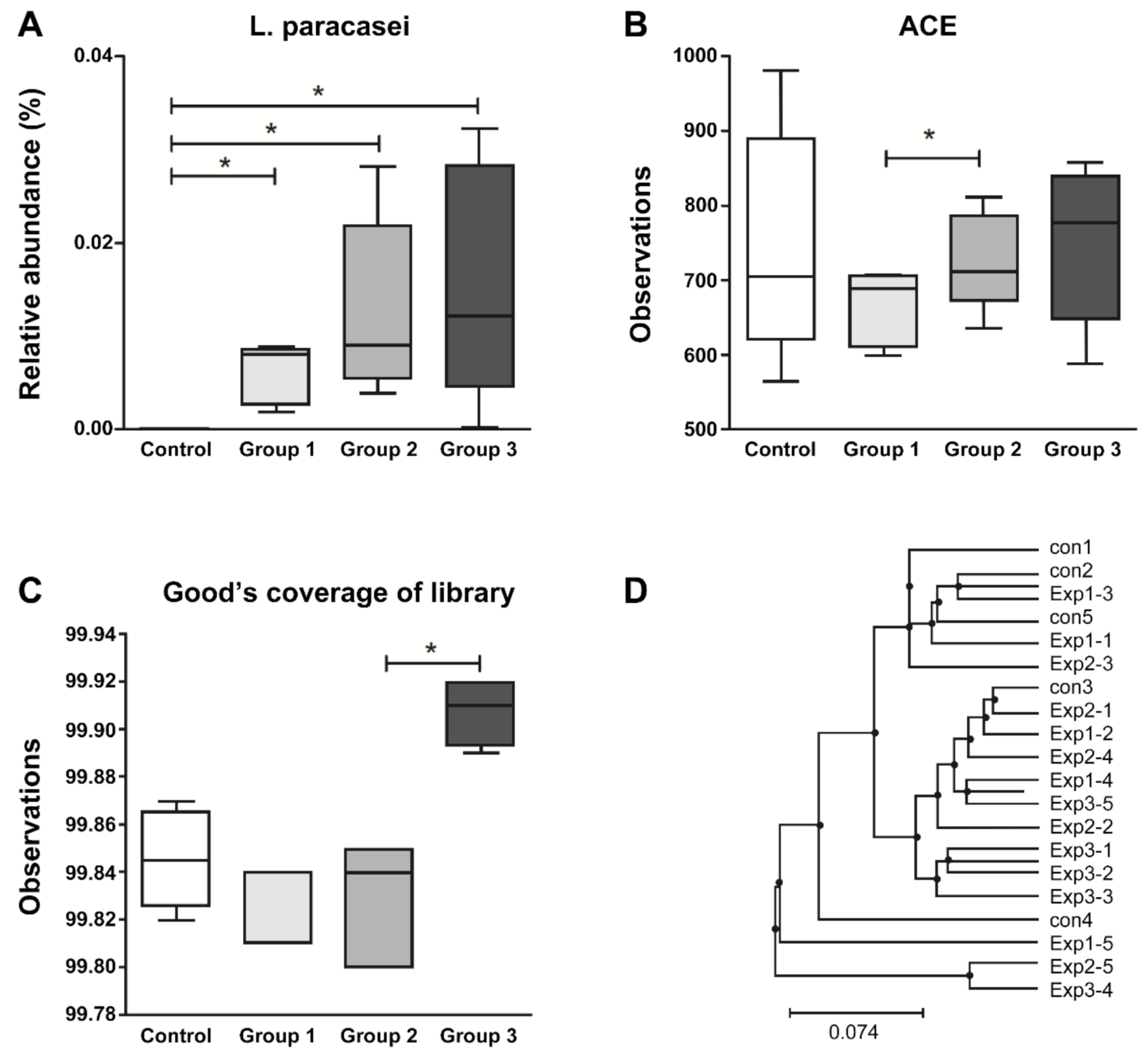
3.3. Fecal Microbial Analysis
3.4. Serum Corticosterone Levels
3.5. Inflammatory Mediator Levels
3.6. Expression of the Tight Junction Proteins in the Colon Tissue
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Song, K.H.; Jung, H.K.; Kim, H.J.; Koo, H.S.; Kwon, Y.H.; Shin, H.D.; Lim, H.C.; Shin, J.E.; Kim, S.E.; Cho, D.H.; et al. Clinical practice guidelines for irritable bowel syndrome in korea, 2017 revised edition. J. Neurogastroenterol. Motil. 2018, 24, 197–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drossman, D.A.; Hasler, W.L. Rome IV—Functional GI Disorders: Disorders of Gut-Brain Interaction. Gastroenterology 2016, 150, 1257–1261. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Savidge, T.; Shulman, R.J. Brain–Gut Microbiome Interactions and Functional Bowel Disorders. Gastroenterology 2014, 146, 1500–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebregts, T.; Adam, B.; Bredack, C.; Röth, A.; Heinzel, S.; Lester, S.; Downie–Doyle, S.; Smith, E.; Drew, P.; Talley, N.J.; et al. Immune Activation in Patients With Irritable Bowel Syndrome. Gastroenterology 2007, 132, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.C.; Talley, N.J. Mucosal inflammation as a potential etiological factor in irritable bowel syndrome: A systematic review. J. Gastroenterol. 2011, 46, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Yao, J.; Wang, C.; Zhang, L.; Kong, W. Molecular and cellular mechanisms of tight junction dysfunction in the irritable bowel syndrome. Mol. Med. Rep. 2015, 12, 3257–3264. [Google Scholar] [CrossRef]
- Kassinen, A.; Krogius-Kurikka, L.; Mäkivuokko, H.; Rinttilä, T.; Paulin, L.; Corander, J.; Malinen, E.; Apajalahti, J.; Palva, A. The Fecal Microbiota of Irritable Bowel Syndrome Patients Differs Significantly From That of Healthy Subjects. Gastroenterology 2007, 133, 24–33. [Google Scholar] [CrossRef]
- Malinen, E.; Krogius-Kurikka, L.; Lyra, A.; Nikkilä, J.; Jääskeläinen, A.; Rinttilä, T.; Vilpponen-Salmela, T.; Von Wright, A.J.; Palva, A. Association of symptoms with gastrointestinal microbiota in irritable bowel syndrome. World J. Gastroenterol. 2010, 16, 4532–4540. [Google Scholar] [CrossRef]
- Tap, J.; Derrien, M.; Toernblom, H.; Brazeilles, R.; Cools-Portier, S.; Doré, J.; Stoersrud, S.; Le Nevé, B.; Oehman, L.; Simrén, M. Identification of an Intestinal Microbiota Signature Associated With Severity of Irritable Bowel Syndrome. Gastroenterology 2017, 152, 111–123.e8. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Pineiro, M.; Asp, N.G.; Reid, G. FAO Technical meeting on prebiotics. J. Clin. Gastroenterol. 2008, 42 (Suppl. 3), S156–S159. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Lara, M.J.; Robles-Sanchez, C.; Ruiz-Ojeda, F.J.; Plaza-Díaz, J.; Gil, Á. Effects of Probiotics and Synbiotics on Obesity, Insulin Resistance Syndrome, Type 2 Diabetes and Non-Alcoholic Fatty Liver Disease: A Review of Human Clinical Trials. Int. J. Mol. Sci. 2016, 17, 928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiller, R.C. Review article: Probiotics and prebiotics in irritable bowel syndrome. Aliment. Pharmacol. Ther. 2008, 28, 385–396. [Google Scholar] [CrossRef]
- O’Mahony, L.; McCarthy, J.; Kelly, P.; Hurley, G.; Luo, F.; Chen, K.; O’Sullivan, G.C.; Kiely, B.; Collins, J.K.; Shanahan, F.; et al. Lactobacillus and bifidobacterium in irritable bowel syndrome: Symptom responses and relationship to cytokine profiles. Gastroenterology 2005, 128, 541–551. [Google Scholar] [CrossRef]
- Ducrotte, P.; Sawant, P.; Jayanthi, V. Clinical trial: Lactobacillus plantarum 299v (DSM 9843) improves symptoms of irritable bowel syndrome. World J. Gastroenterol. 2012, 18, 4012–4018. [Google Scholar] [CrossRef]
- Kim, M.J.; Min, Y.W.; Lee, C.; Jang, Y.S.; Kim, C.H.; Chang, D.K. The effects of three novel probiotics isolated from the Korean fermented food Kimchi on the stress-induced defecation of rats. Precis. Futur. Med. 2020, 4, 31–36. [Google Scholar] [CrossRef]
- Cho, Y.-H.; Hong, S.-M.; Kim, C.-H. Isolation and Characterization of Lactic Acid Bacteria from Kimchi, Korean Traditional Fermented Food to Apply into Fermented Dairy Products. Food Sci. Anim. Resour. 2013, 33, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.G.; Min, Y.W.; Lee, C.; Hong, S.N.; Won, J.Y.; Jang, J.A.; Kim, C.-H.; Chang, D.K. Effects of Novel Probiotics in a Murine Model of Irritable Bowel Syndrome. Korean J. Gastroenterol. 2020, 75, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.Y.; Park, S.-C.; Kim, T.-W.; Kim, K.-S.; Song, J.-C.; Lee, H.-M.; Sung, H.-J.; Rhee, M.H.; Kim, S.-K.; Park, H.-J.; et al. Radical scavenging and anti-inflammatory activity of extracts from Opuntia humifusa raf. J. Pharm. Pharmacol. 2006, 58, 113–119. [Google Scholar] [CrossRef]
- Kang, Y.J.; Kim, H.Y.; Lee, C.; Park, S.Y. Nitric oxide inhibitory constituents from fruits of Opuntia humifusa. Nat. Prod. Sci. 2014, 20, 211–215. [Google Scholar]
- Guevara-Arauza, J.C.; Ornelas-Paz, J.D.J.; Pimentel-González, D.J.; Mendoza, S.R.; Guerra, R.E.S.; Paz-Maldonado, L.M.T. Prebiotic effect of mucilage and pectic-derived oligosaccharides from nopal (Opuntia ficus-indica). Food Sci. Biotechnol. 2012, 21, 997–1003. [Google Scholar] [CrossRef]
- Patel, S. Opuntia cladodes (nopal): Emerging functional food and dietary supplement. Mediterr. J. Nutr. Metab. 2014, 7, 11–19. [Google Scholar] [CrossRef]
- Yu, H.S.; Lee, N.K.; Choi, A.J.; Choe, J.S.; Bae, C.H.; Paik, H.D. Anti-inflammatory potential of probiotic strain weissella cibaria jw15 isolated from kimchi through regulation of nf-kappab and mapks pathways in lps-induced raw 264.7 cells. J. Microbiol. Biotechnol. 2019, 29, 1022–1032. [Google Scholar] [CrossRef]
- Won, J.-Y.; Kim, H.-S.; Jang, J.-A.; Kim, C.-H. Functional Properties of Yogurt Containing Specific Peptides derived from Whey Proteins. J. Milk Sci. Biotechnol. 2017, 35, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Girard, S.-A.; Bah, T.M.; Kaloustian, S.; Lada-Moldovan, L.; Rondeau, I.; Tompkins, T.A.; Godbout, R.; Rousseau, G. Lactobacillus helveticus and Bifidobacterium longum taken in combination reduce the apoptosis propensity in the limbic system after myocardial infarction in a rat model. Br. J. Nutr. 2009, 102, 1420–1425. [Google Scholar] [CrossRef] [Green Version]
- Arseneault-Bréard, J.; Rondeau, I.; Gilbert, K.; Girard, S.-A.; Tompkins, T.A.; Godbout, R.; Rousseau, G. Combination of Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 reduces post-myocardial infarction depression symptoms and restores intestinal permeability in a rat model. Br. J. Nutr. 2011, 107, 1793–1799. [Google Scholar] [CrossRef] [Green Version]
- Dolatabadi, F.; Abdolghaffari, A.H.; Farzaei, M.H.; Baeeri, M.; Ziarani, F.S.; Eslami, M.; Abdollahi, M.; Rahimi, R. The Protective Effect of Melissa officinalis L. in Visceral Hypersensitivity in Rat Using 2 Models of Acid-induced Colitis and Stress-induced Irritable Bowel Syndrome: A Possible Role of Nitric Oxide Pathway. J. Neurogastroenterol. Motil. 2018, 24, 490–501. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumine sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Li, Y.; Ma, C.; Wang, C.; Sun, Z.; Shen, Y.; Liu, L.; Li, S.; Zhang, X.; Cong, B. Restraint Stress Induced Hyperpermeability and Damage of the Blood-Brain Barrier in the Amygdala of Adult Rats. Front. Mol. Neurosci. 2019, 12, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukui, H.; Oshima, T.; Tanaka, Y.; Oikawa, Y.; Makizaki, Y.; Ohno, H.; Tomita, T.; Watari, J.; Miwa, H. Effect of probiotic Bifidobacterium bifidum G9-1 on the relationship between gut microbiota profile and stress sensitivity in maternally separated rats. Sci. Rep. 2018, 8, 12384. [Google Scholar] [CrossRef] [PubMed]
- Bonapersona, V.; Damsteegt, R.; Adams, M.L.; Van Weert, L.T.C.M.; Meijer, O.C.; Joëls, M.; Sarabdjitsingh, R.A. Sex-Dependent Modulation of Acute Stress Reactivity After Early Life Stress in Mice: Relevance of Mineralocorticoid Receptor Expression. Front. Behav. Neurosci. 2019, 13, 181. [Google Scholar] [CrossRef]
- Schemitt, E.; Hartmann, R.M.; Colares, J.R.; Licks, F.; Salvi, J.O.; Marroni, C.A.; Marroni, N.P. Protective action of glutamine in rats with severe acute liver failure. World J. Hepatol. 2019, 11, 273–286. [Google Scholar] [CrossRef]
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight Junction in the Intestinal Epithelium: Its Association with Diseases and Regulation by Phytochemicals. J. Immunol. Res. 2018, 2018, 2645465. [Google Scholar] [CrossRef] [Green Version]
- Vaziri, N.D.; Yuan, J.; Rahimi, A.; Ni, Z.; Said, H.; Subramanian, V.S. Disintegration of colonic epithelial tight junction in uremia: A likely cause of CKD-associated inflammation. Nephrol. Dial. Transplant. 2011, 27, 2686–2693. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.I.; Choi, C.; Shin, S.-W.; Son, A.; Lee, G.-H.; Kim, S.-Y.; Park, H.C. Valproic Acid Sensitizes Hepatocellular Carcinoma Cells to Proton Therapy by Suppressing NRF2 Activation. Sci. Rep. 2017, 7, 14986. [Google Scholar] [CrossRef]
- Mazurak, N.; Broelz, E.; Storr, M.; Enck, P. Probiotic Therapy of the Irritable Bowel Syndrome: Why Is the Evidence Still Poor and What Can Be Done About It? J. Neurogastroenterol. Motil. 2015, 21, 471–485. [Google Scholar] [CrossRef] [Green Version]
- De Vrese, M.; Schrezenmeir, J. Probiotics, prebiotics, and synbiotics. In Food Biotechnology; Stahl, U., Donalies, U.E.B., Nevoigt, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 1–66. [Google Scholar]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Quigley, E.M. Prebiotics and Probiotics in Digestive Health. Clin. Gastroenterol. Hepatol. 2019, 17, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M. Pharmacological actions of Opuntia ficus indica: A Review. J. Appl. Pharm. Sci. 2012, 2, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Gerez, C.; De Valdez, G.F.; Gigante, M.; Grosso, C. Whey protein coating bead improves the survival of the probiotic Lactobacillus rhamnosus CRL 1505 to low pH. Lett. Appl. Microbiol. 2012, 54, 552–556. [Google Scholar] [CrossRef]
- Riddle, M.S.; Welsh, M.; Porter, C.K.; Nieh, C.; Boyko, E.J.; Gackstetter, G.; Hooper, T.I. The Epidemiology of Irritable Bowel Syndrome in the US Military: Findings from the Millennium Cohort Study. Am. J. Gastroenterol. 2016, 111, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Dinan, T.G.; Quigley, E.M.; Ahmed, S.M.; Scully, P.; O’Brien, S.; O’Mahony, L.; O’Mahony, S.; Shanahan, F.; Keeling, P.N. Hypothalamic-Pituitary-Gut Axis Dysregulation in Irritable Bowel Syndrome: Plasma Cytokines as a Potential Biomarker? Gastroenterology 2006, 130, 304–311. [Google Scholar] [CrossRef]
- Zheng, G.; Wu, S.-P.; Hu, Y.; Smith, D.E.; Wiley, J.W.; Hong, S. Corticosterone mediates stress-related increased intestinal permeability in a region-specific manner. Neurogastroenterol. Motil. 2013, 25, e127–e139. [Google Scholar] [CrossRef]
- Holtmann, G.J.; Ford, A.C.; Talley, N.J. Pathophysiology of irritable bowel syndrome. Lancet Gastroenterol. Hepatol. 2016, 1, 133–146. [Google Scholar] [CrossRef]
- Shulman, R.J.; Jarrett, M.E.; Cain, K.C.; Broussard, E.K.; Heitkemper, M.M. Associations among gut permeability, inflammatory markers, and symptoms in patients with irritable bowel syndrome. J. Gastroenterol. 2014, 49, 1467–1476. [Google Scholar] [CrossRef] [Green Version]
- Bertiaux-Vandaële, N.; Youmba, S.B.; Belmonte, L.; Lecleire, S.; Antonietti, M.; Gourcerol, G.; Leroi, A.-M.; Déchelotte, P.; Ménard, J.-F.; Ducrotté, P.; et al. The Expression and the Cellular Distribution of the Tight Junction Proteins Are Altered in Irritable Bowel Syndrome Patients With Differences According to the Disease Subtype. Am. J. Gastroenterol. 2011, 106, 2165–2173. [Google Scholar] [CrossRef]
- Marshall, J.K.; Thabane, M.; Garg, A.X.; Clark, W.; Meddings, J.; Collins, S.M.; WEL Investigators. Intestinal permeability in patients with irritable bowel syndrome after a waterborne outbreak of acute gastroenteritis in Walkerton, Ontario. Aliment. Pharmacol. Ther. 2004, 20, 1317–1322. [Google Scholar] [CrossRef]
- Williams, C.L.; Villar, R.G.; Peterson, J.M.; Burks, T.F. Stress-induced changes in intestinal transit in the rat: A model for irritable bowel syndrome. Gastroenterology 1988, 94, 611–621. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Evangelista, S. Experimental Models of Irritable Bowel Syndrome and the Role of the Enteric Neurotransmission. J. Clin. Med. 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gue, M.; Del Rio-Lacheze, C.; Eutamene, H.; Théodorou, V.; Fioramonti, J.; Bueno, L. Stress-induced visceral hypersensitivity to rectal distension in rats: Role of CRF and mast cells. Neurogastroenterol. Motil. 1997, 9, 271–279. [Google Scholar] [CrossRef]
- Harris, R.B.S. Repeated restraint stress lowers the threshold for response to third ventricle CRF administration. Horm. Behav. 2017, 89, 64–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larauche, M.; Mulak, A.; Taché, Y. Stress and visceral pain: From animal models to clinical therapies. Exp. Neurol. 2012, 233, 49–67. [Google Scholar] [CrossRef] [Green Version]
- Traini, C.; Evangelista, S.; Girod, V.; Faussone-Pellegrini, M.; Vannucchi, M.G. Changes of excitatory and inhibitory neurotransmitters in the colon of rats underwent to the wrap partial restraint stress. Neurogastroenterol. Motil. 2016, 28, 1172–1185. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seong, G.; Lee, S.; Min, Y.W.; Jang, Y.S.; Park, S.-Y.; Kim, C.-H.; Lee, C.; Hong, S.N.; Chang, D.K. Effect of a Synbiotic Containing Lactobacillus paracasei and Opuntia humifusa on a Murine Model of Irritable Bowel Syndrome. Nutrients 2020, 12, 3205. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12103205
Seong G, Lee S, Min YW, Jang YS, Park S-Y, Kim C-H, Lee C, Hong SN, Chang DK. Effect of a Synbiotic Containing Lactobacillus paracasei and Opuntia humifusa on a Murine Model of Irritable Bowel Syndrome. Nutrients. 2020; 12(10):3205. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12103205
Chicago/Turabian StyleSeong, Gyeol, Seungbaek Lee, Yang Won Min, Yeon Sil Jang, So-Young Park, Cheol-Hyun Kim, Chansu Lee, Sung Noh Hong, and Dong Kyung Chang. 2020. "Effect of a Synbiotic Containing Lactobacillus paracasei and Opuntia humifusa on a Murine Model of Irritable Bowel Syndrome" Nutrients 12, no. 10: 3205. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12103205