The Role of Omega-3 Polyunsaturated Fatty Acids from Different Sources in Bone Development


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diet Preparation and Composition
2.3. Lipid Extraction and Analysis by Gas Chromatography (GC)
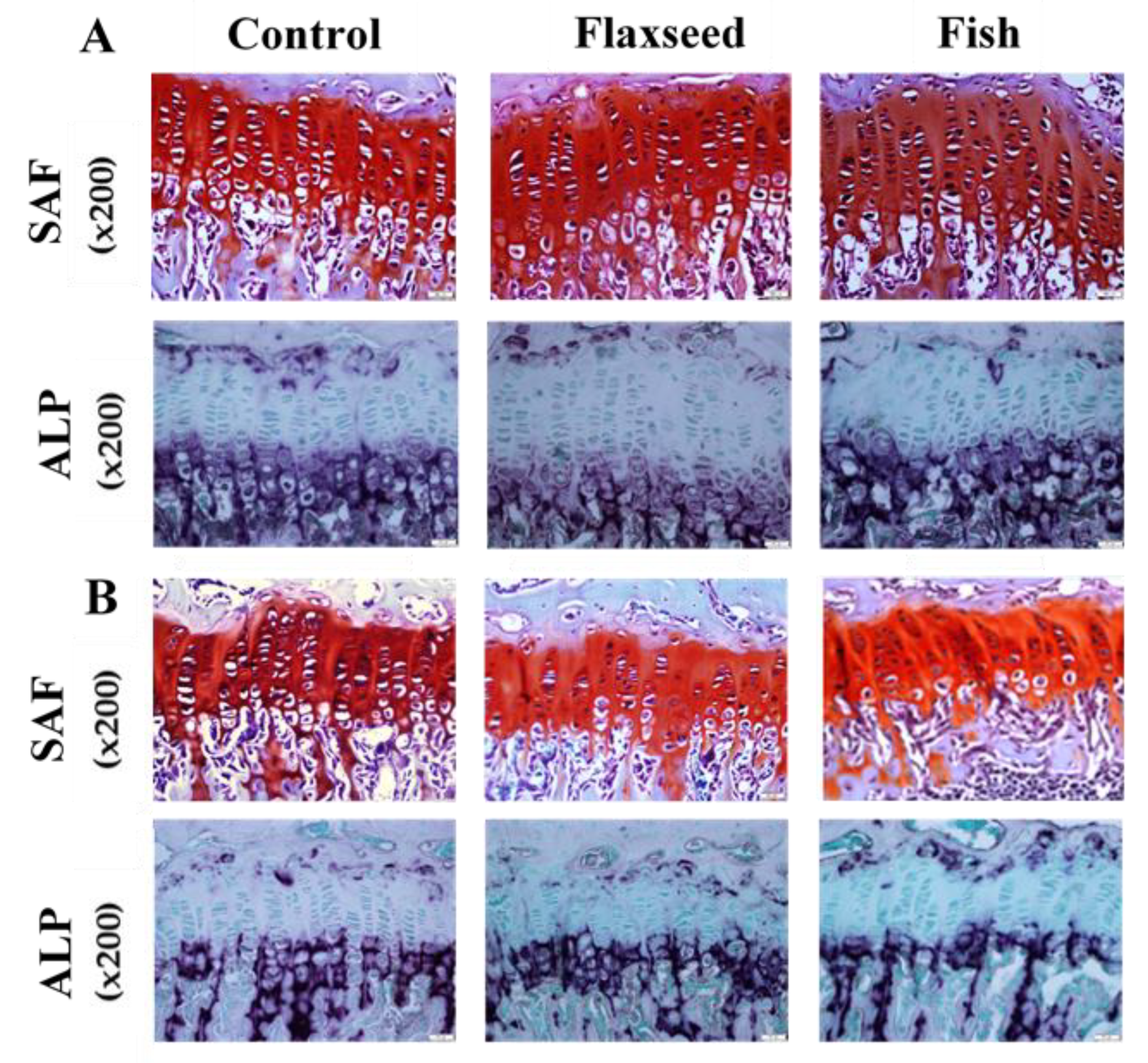
2.4. Histological Staining of Growth-Plate (GP) Sections
2.5. Imaging and Measurement of GPs
2.6. Micro-CT
2.7. Mechanical Testing
2.8. Bone RNA Extraction and RNA-Sequencing
2.8.1. Bioinformatics
2.8.2. Gene-Set Enrichment Analysis
2.9. Statistical Analysis
3. Results
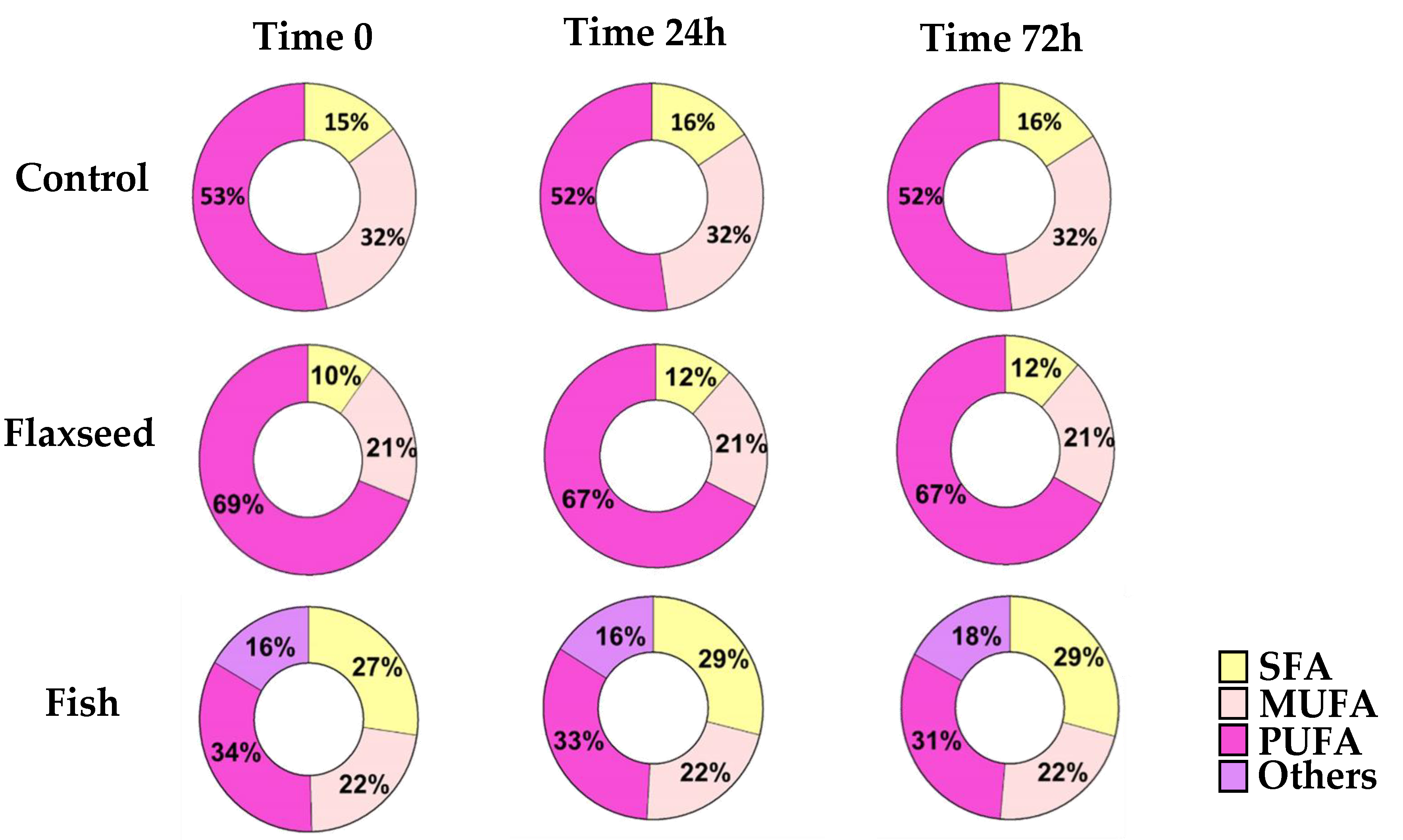
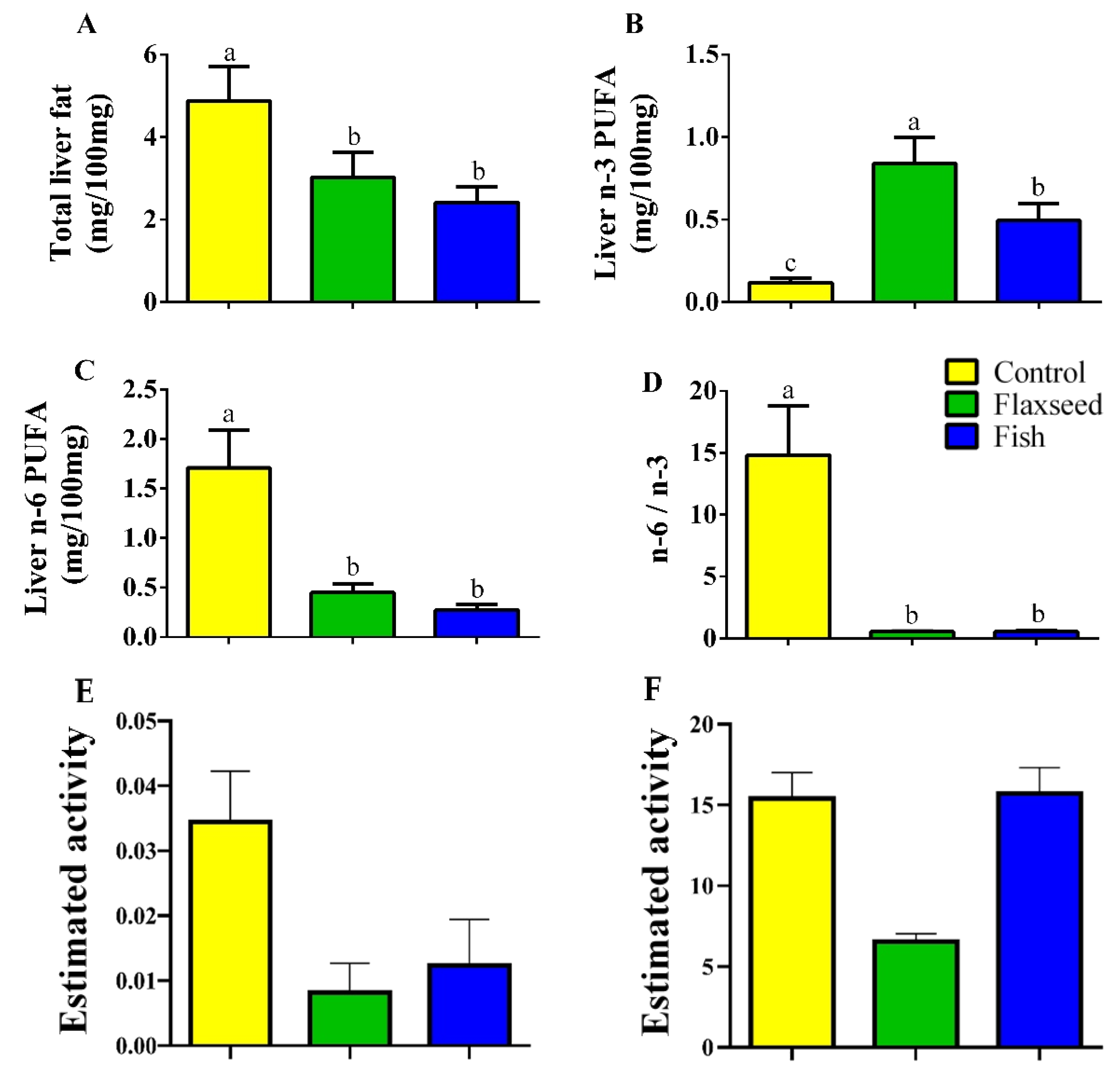
3.1. Dietary n-3 PUFAs Decrease Fat Accumulation and Alter FA Content in Liver and Serum
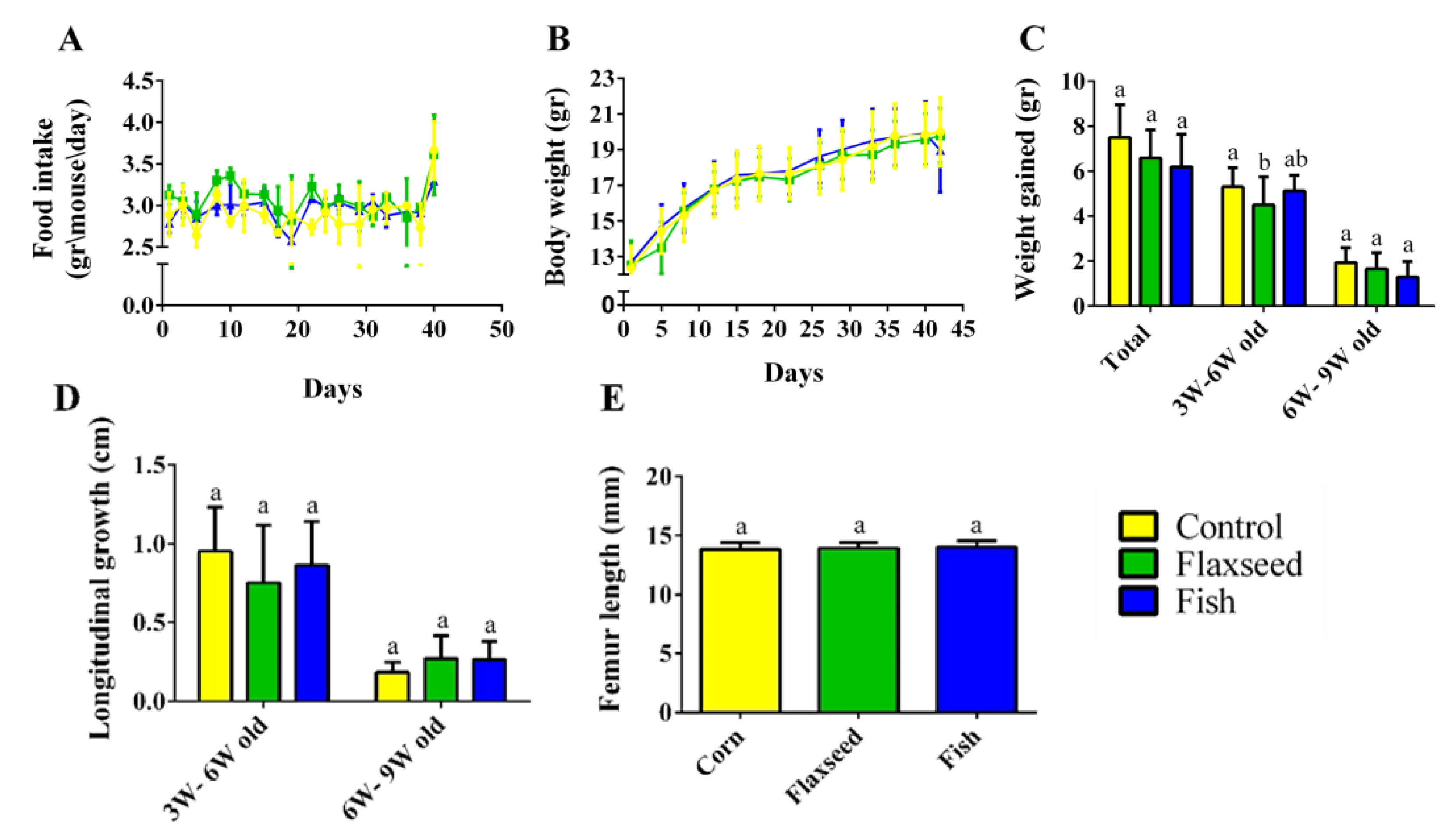
3.2. Effect of Omega-3 from Different Sources on Growth Pattern and Food Consumption
3.3. The Effect of Diets Rich in n-3 PUFAs on Bone Quality
3.3.1. Diets Rich in n-3 PUFAs Improve Bone Mechanical Properties
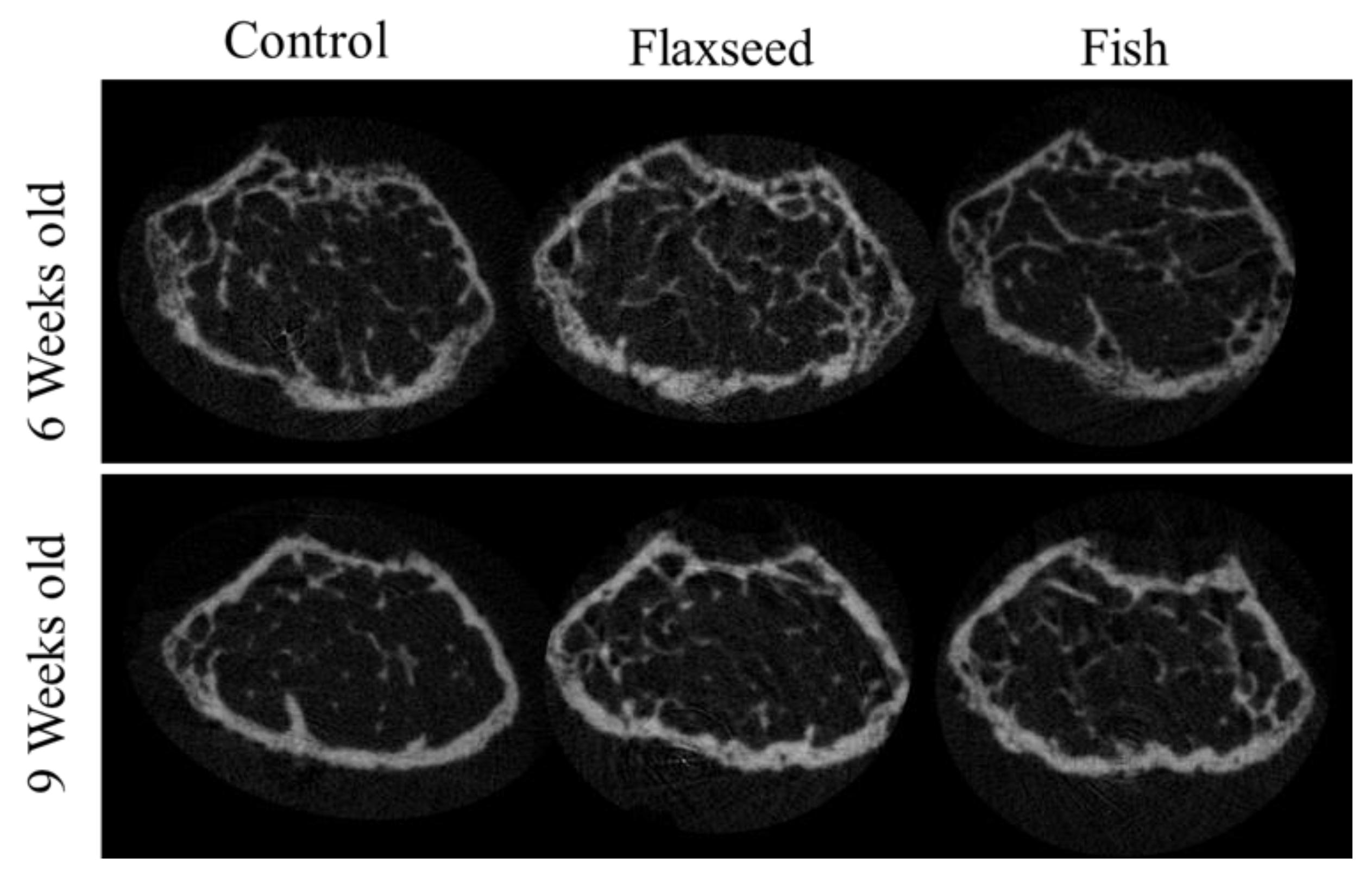
3.3.2. Diet Containing Plant Source of n-3 PUFAs (Flaxseed Oil) Improved Trabecular Bone’s Micro-Architecture
3.3.3. Diet Containing n-3 PUFAs from an Animal Source (Fish Oil) Increased BMD
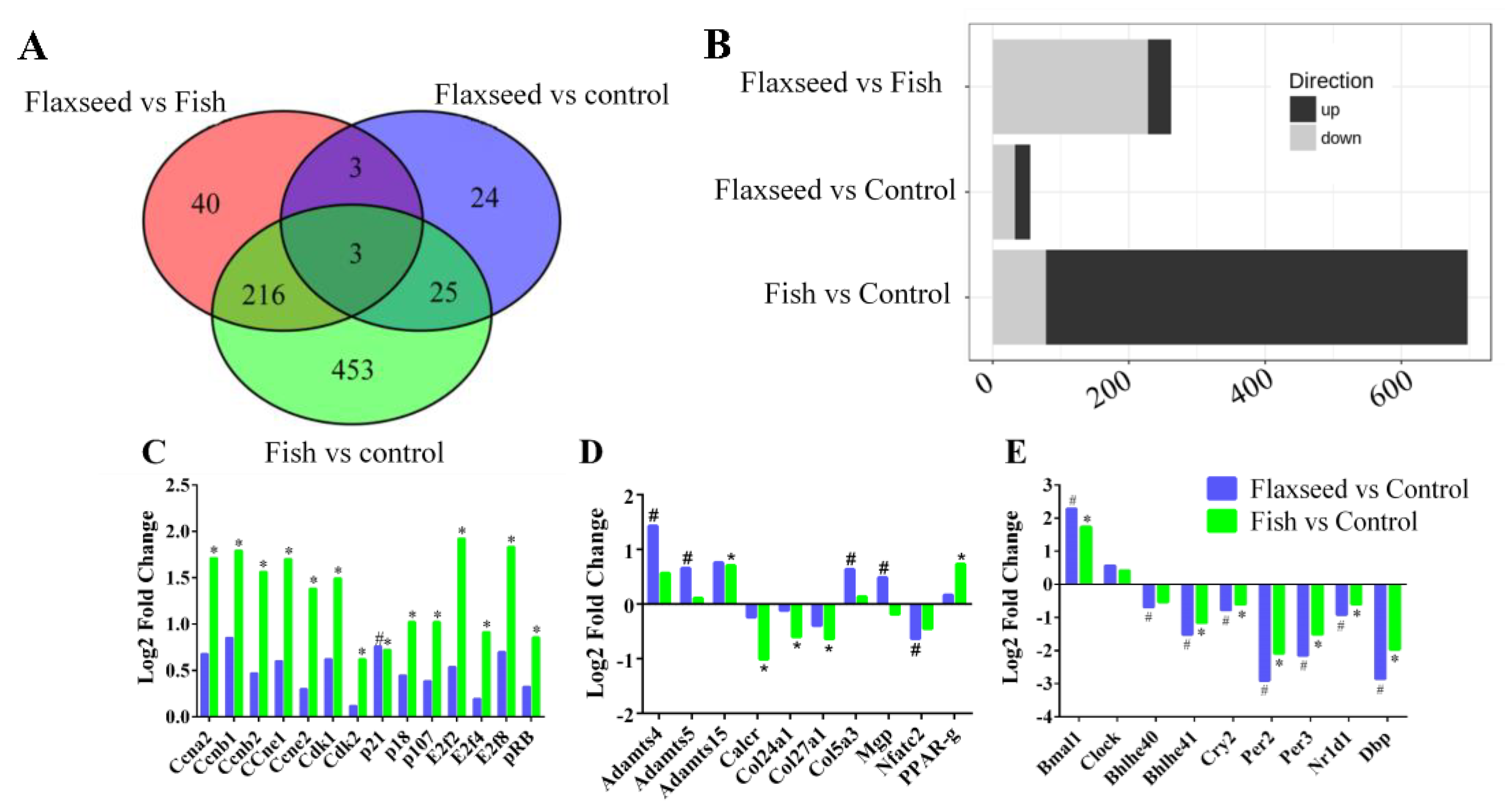
3.4. The Effect of Different Sources of n-3 PUFAs on Bone’s Transcriptional Regulation
IPA Pathway Enrichment Analysis of Genes Related to Dietary n-3 PUFAs
4. Discussion
4.1. Dietary n-3 PUFAs Decrease Hepatic and Serum Fat Levels
4.2. Diet Rich in n-3 PUFAs Affects Postnatal Skeletal Development and Bone Characteristics
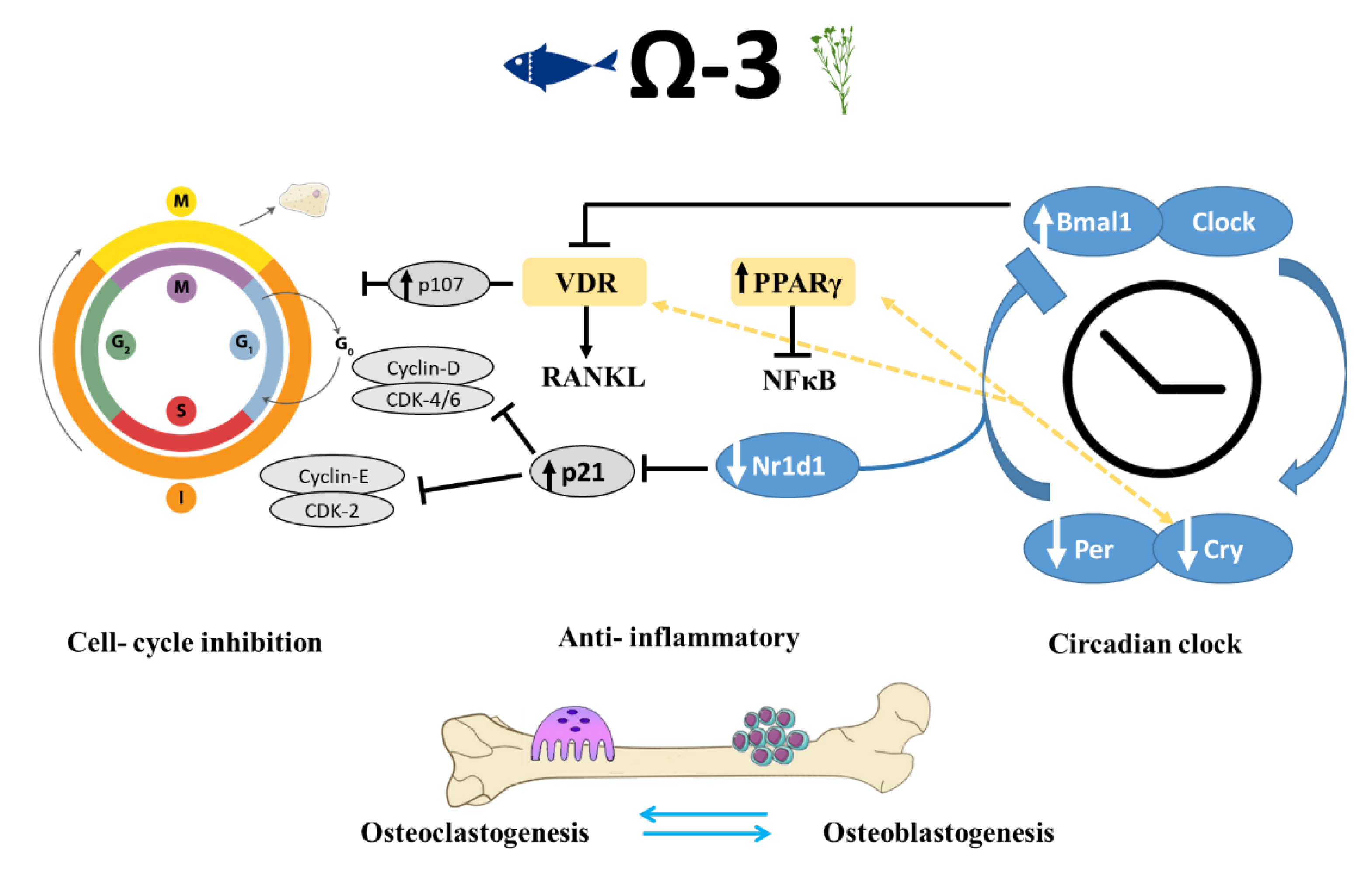
4.3. Transcriptional Regulation of Bone by Different Sources n-3 PUFAs
4.4. Dietary n-3 PUFAs Alter Circadian Clock Gene Expression in Bone
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Karsenty, G. The complexities of skeletal biology. Nature 2003, 423, 316–318. [Google Scholar] [CrossRef] [PubMed]
- Daly, R.M.; Duckham, R.L.; Gianoudis, J. Evidence for an Interaction Between Exercise and Nutrition for Improving Bone and Muscle Health. Curr. Osteoporos. Rep. 2014, 12, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Longo, A.B.; Ward, W.E. PUFAs, Bone Mineral Density, and Fragility Fracture: Findings from Human Studies. Adv. Nutr. 2016, 7, 299–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langdahl, B.; Ferrari, S.; Dempster, D.W. Bone modeling and remodeling: Potential as therapeutic targets for the treatment of osteoporosis. Ther. Adv. Musculoskelet. Dis. 2016, 8, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genuis, S.J.; Schwalfenberg, G.K. Picking a bone with contemporary osteoporosis management: Nutrient strategies to enhance skeletal integrity. Clin. Nutr. 2007, 26, 193–207. [Google Scholar] [CrossRef]
- Högström, M.; Nordström, P.; Nordström, A. n-3 fatty acids are positively associated with peak bone mineral density and bone accrual in healthy men: The NO2 study. Am. J. Clin. Nutr. 2007, 85, 803–807. [Google Scholar] [CrossRef] [Green Version]
- Lukas, R.; Gigliotti, J.C.; Smith, B.J.; Altman, S.; Tou, J.C. Consumption of different sources of omega-3 polyunsaturated fatty acids by growing female rats affects long bone mass and microarchitecture. Bone 2011, 49, 455–462. [Google Scholar] [CrossRef]
- López-vicario, C.; González-périz, A.; Rius, B.; Morán-salvador, E.; García-alonso, V.; Lozano, J.J.; Bataller, R.; Kang, J.X.; Arroyo, V.; Clària, J.; et al. Molecular interplay between Δ5/Δ6 desaturases and long-chain fatty acids in the pathogenesis of non-alcoholic steatohepatitis. Gut 2014, 63, 344–355. [Google Scholar] [CrossRef]
- Poudyal, H.; Panchal, S.K.; Diwan, V.; Brown, L. Omega-3 fatty acids and metabolic syndrome: Effects and emerging mechanisms of action. Prog. Lipid Res. 2011, 50, 372–387. [Google Scholar] [CrossRef]
- Narayan, B.; Miyashita, K.; Hosokawa, M. Physiological effects of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA)—A review. Food Rev. Int. 2006, 22, 291–307. [Google Scholar] [CrossRef]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Kruger, M.C.; Coetzee, M.; Haag, M.; Weiler, H. Long-chain polyunsaturated fatty acids: Selected mechanisms of action on bone. Prog. Lipid Res. 2010, 49, 438–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simopoulos, A.P. The omega-6/omega-3 fatty acid ratio: Health implications. Oléagineux Corps Gras Lipides 2010, 17, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Tur, J.A.; Bibiloni, M.M.; Sureda, A.; Pons, A. Dietary sources of omega 3 fatty acids: Public health risks and benefits. Br. J. Nutr. 2012, 107, S23–S52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koren, N.; Simsa-Maziel, S.; Shahar, R.; Schwartz, B.; Monsonego-Ornan, E. Exposure to omega-3 fatty acids at early age accelerate bone growth and improve bone quality. J. Nutr. Biochem. 2014, 25, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G. Committee Report AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Mesilati-stahy, R.; Mida, K.; Argov-argaman, N. Size-Dependent Lipid Content of Bovine Milk Fat Globule and Membrane Phospholipids. J. Agric. Food Chem. 2011, 59, 7427–7435. [Google Scholar] [CrossRef]
- Simsa-maziel, S.; Zaretsky, J.; Reich, A.; Koren, Y.; Shahar, R.; Monsonego-ornan, E. IL-1RI participates in normal growth plate development and bone modeling. Am. J. Physiol. Metab. 2013, 305, E15–E21. [Google Scholar] [CrossRef] [Green Version]
- Idelevich, A.; Kerschnitzki, M.; Shahar, R.; Monsonego-ornan, E. 1,25(OH)2D3 Alters Growth Plate Maturation and Bone Architecture in Young Rats with Normal Renal Function. PLoS ONE 2011, 6, e20772. [Google Scholar] [CrossRef]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Mu, R. Guidelines for Assessment of Bone Microstructure in Rodents Using Micro—Computed Tomography. J. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef]
- Sharir, A.; Max, M.; Shahar, R. Whole bone mechanics and mechanical testing. Veter. J. 2008, 177, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Ayturk, U.M.; Jacobsen, C.M.; Christodoulou, D.C.; Gorham, J.; Seidman, J.G.; Seidman, C.E.; Robling, G.; Warman, M.L. An RNA-seq Protocol to Identify mRNA Expression Changes in Mouse Diaphyseal Bone: Applications in Mice with Bone Property Altering Lrp5 Mutations. J. Bone Miner. 2013, 28, 2081–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeidipour, B.; Bakhshi, S. The relationship between organizational culture and knowledge management and their simultaneous effects on customer relation management. Adv. Environ. Biol. 2013, 7, 2803–2809. [Google Scholar]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Köster, J.; Rahmann, S. Snakemake—A scalable bioinformatics workflow engine. Bioinformatics 2012, 28, 2520–2522. [Google Scholar] [CrossRef] [Green Version]
- Awada, M.; Soulage, C.O.; Meynier, A.; Debard, C.; Plaisancié, P.; Benoit, B.; Picard, G.; Loizon, E.; Chauvin, M.; Estienne, M.; et al. Dietary oxidized n-3 PUFA induce oxidative stress and infl ammation: Role of intestinal absorption of 4-HHE and reactivity in intestinal cells. J. Lipid Res. 2012, 53, 2069–2080. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.P.; Nakamura, M.T.; Clarke, S.D. Cloning, Expression, and Nutritional Regulation of the Mammalian Δ-6 desaturase. J. Biol. Chem. 1999, 274, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Wit, J.M.; Camacho-Hübner, C. Endocrine regulation of longitudinal bone growth. Endocr. Dev. 2011, 21, 30–41. [Google Scholar] [CrossRef]
- Golub, E.E.; Boesze-battaglia, K. The role of alkaline phosphatase in mineralization. Curr. Opin. Orthop. 2007, 18, 444–448. [Google Scholar] [CrossRef]
- Glatt, V.; Canalis, E.; Stadmeyer, L.; Bouxsein, M.L. Age-related changes in trabecular architecture differ in female and male C57BL/6J mice. J. Bone Miner. Res. 2007, 22, 1197–1207. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.Y.Y.; Ward, W.E.; Kang, J.X.; Ma, D.W.L. Femur EPA and DHA are correlated with femur biomechanical strength in young fat-1 mice. J. Nutr. Biochem. 2009, 20, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Dodin, S.; Lemay, A.; Jacques, H.; Le, F. The Effects of Flaxseed Dietary Supplement on Lipid Profile, Bone Mineral Density, and Symptoms in Menopausal Women: A Randomized, Double-Blind, Wheat Germ Placebo-Controlled Clinical Trial. J. Clin. Endocrinol. Metab. 2015, 90, 1390–1397. [Google Scholar] [CrossRef]
- Lau, B.Y.Y.; Cohen, D.J.A.; Ward, W.E.; Ma, D.W.L. Investigating the Role of Polyunsaturated Fatty Acids in Bone Development Using Animal Models. Molecules 2013, 18, 14203–14227. [Google Scholar] [CrossRef] [Green Version]
- Khadge, S.; Graham, J.; Thiele, G.M.; Mcguire, T.R.; Klassen, L.W.; Duryee, M.J.; Britton, H.C.; Dafferner, A.J.; Beck, J.; Black, P.N.; et al. ScienceDirect Dietary omega-3 and omega-6 polyunsaturated fatty acids modulate hepatic pathology. J. Nutr. Biochem. 2018, 52, 92–102. [Google Scholar] [CrossRef]
- Huang, W. Endogenously elevated n-3 polyunsaturated fatty acids alleviate acute ethanol-induced liver steatosis. BioFactors 2015, 453–462. [Google Scholar] [CrossRef]
- Tou, J.C.; Altman, S.N.; Gigliotti, J.C.; Benedito, V.A.; Cordonier, E.L. Different sources of omega-3 polyunsaturated fatty acids affects apparent digestibility, tissue deposition, and tissue oxidative stability in growing female rats. Lipids Health Dis. 2011, 10, 179. [Google Scholar] [CrossRef] [Green Version]
- Food and Nutrition Board; Institute of Medicine; National Academy of Science. Dietary Reference Intakes (DRIs): Recommended Dietary Allowances and Adequate Intakes, Vitamins; The National Academic Press: Washington, DC, USA, 2015; ISBN 9780323340755. [Google Scholar]
- Wang, M.; Zhang, X.; Ma, L.; Feng, R.; Yan, C.; Su, H.; He, C.; Kang, J.X.; Liu, B.; Wan, J. Omega-3 polyunsaturated fatty acids ameliorate ethanol-induced adipose hyperlipolysis: A mechanism for hepatoprotective e ff ect against alcoholic liver disease. BBA Mol. Basis Dis. 2017, 1863, 3190–3201. [Google Scholar] [CrossRef]
- Peltier, S.; Portois, L.; Malaisse, W.J.; Carpentier, Y.A. Fatty acid profile of plasma and liver lipids in mice depleted in long-chain polyunsaturated (n-3) fatty acids. Int. J. Mol. Med. 2008, 22, 559–563. [Google Scholar] [CrossRef]
- Oh, D.Y.; Walenta, E. Omega-3 fatty acids and FFAR4. Front. Endocrinol. 2014, 5, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Park, S.Y.; Baek, J.E.; Lee, S.Y.; Baek, W.Y.; Lee, S.Y.; Lee, Y.S.; Yoo, H.J.; Kim, H.; Lee, S.H.; et al. Free fatty acid receptor 4 (GPR120) stimulates bone formation and suppresses bone resorption in the presence of elevated n-3 fatty acid levels. Endocrinology 2016, 157, 2621–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, J.H.; Van Der Meulen, M.C.H. Whole bone mechanics and bone quality. Clin. Orthop. Relat. Res. 2011, 469, 2139–2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolfe, R.A.; Nowlan, N.C.; Kenny, E.M.; Cormican, P.; Morris, D.W.; Prendergast, P.J.; Kelly, D.; Murphy, P. Identification of mechanosensitive genes during skeletal development: Alteration of genes associated with cytoskeletal rearrangement and cell signalling pathways. BMC Genom. 2014, 15, 48. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X. Identification of key genes and pathways associated with osteogenic differentiation of adipose stem cells. J. Cell. Physiol. 2018, 233, 9777–9785. [Google Scholar] [CrossRef]
- Ho, X.D.; Phung, P.; Le, V.Q.; Nguyen, V.H.; Reimann, E.; Prans, E.; Kõks, G.; Maasalu, K.; Le, N.T.; Trinh, L.H.; et al. Whole transcriptome analysis identifies differentially regulated networks between osteosarcoma and normal bone samples. Exp. Biol. Med. 2017, 242, 1802–1811. [Google Scholar] [CrossRef]
- Articles, R. Osteoblast Differentiation at a Glance. Med Sci. Monit. Basic Res. 2016, 22, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Blair, H.C.; Larrouture, Q.C.; Li, Y.; Lin, H.; Beer-Stoltz, D.; Liu, L.; Tuan, R.S.; Robinson, L.J.; Schlesinger, P.H.; Nelson, D.J. Osteoblast differentiation and bone matrix formation in vivo and in vitro. Tissue Eng. Part B Rev. 2017, 23, 268–280. [Google Scholar] [CrossRef] [Green Version]
- Varga, T.; Czimmerer, Z.; Nagy, L. PPARs are a unique set of fatty acid regulated transcription factors controlling both lipid metabolism and in fl ammation. BBA Mol. Basis Dis. 2011, 1812, 1007–1022. [Google Scholar] [CrossRef]
- Nakanishi, A.; Tsukamoto, I. N-3 polyunsaturated fatty acids stimulate osteoclastogenesis through PPARγ-mediated enhancement of c-Fos expression, and suppress osteoclastogenesis through PPARγ-dependent inhibition of NFkB activation. J. Nutr. Biochem. 2015, 26, 1317–1327. [Google Scholar] [CrossRef]
- Turner, A.G.; Tjahyono, F.; Chiu, W.S.M.; Skinner, J.; Sawyer, R.; Moore, A.J.; Morris, H.A.; Findlay, D.M.; Zajac, J.D.; Davey, R.A. The role of the calcitonin receptor in protecting against induced hypercalcemia is mediated via its actions in osteoclasts to inhibit bone resorption. Bone 2011, 48, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Magee, P.; Pearson, S.; Whittingham-Dowd, J.; Allen, J. PPARγ as a molecular target of EPA anti-inflammatory activity during TNF-α-impaired skeletal muscle cell differentiation. J. Nutr. Biochem. 2012, 23, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Hirai, T. Circadian clock and bone biology. J. Oral Biosci. 2017, 59, 179–183. [Google Scholar] [CrossRef]
- Oosterman, J.E.; Kalsbeek, A.; Fleur, S.E.; Belsham, D.D. Impact of nutrients on circadian rhythmicity. Am. J. Physiol. Integr. Comp. Physiol. 2015, 308, R337–R350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Ochi, H.; Fukuda, T.; Sato, S.; Sunamura, S.; Takarada, T.; Hinoi, E.; Okawa, A.; Takeda, S. Circadian Clock Regulates Bone Resorption in Mice. J. Bone Miner. Res. 2016, 31, 1344–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takarada, T.; Xu, C.; Ochi, H.; Nakazato, R.; Yamada, D.; Nakamura, S.; Kodama, A.; Shimba, S.; Mieda, M.; Fukasawa, K.; et al. Bone Resorption Is Regulated by Circadian Clock in Osteoblasts. J. Bone Miner. Res. 2017, 32, 872–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreani, T.S.; Itoh, T.Q.; Yildirim, E.; Hwangbo, D.-S.; Allada, R. Genetics of Circadian Rhythms. Sleep Med. Clin. 2015, 10, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Patel, M.S.; Bradley, A.; Wagner, E.F.; Karsenty, G. Leptin-Regulated Bone Formation Bone Disease Program of Texas. Cell 2005, 122, 803–815. [Google Scholar] [CrossRef] [Green Version]
- Goldman, A.L.; Donlon, C.M.; Cook, N.R.; Manson, J.E.; Buring, J.E.; Copeland, T.; Yu, C.Y.; Leboff, M.S. VIT amin D and OmegA-3 TriaL (VITAL) bone health ancillary study: Clinical factors associated with trabecular bone score in women and men. Osteoporos. Int. 2018, 29, 2505–2515. [Google Scholar] [CrossRef]
- Kawai, M.; Rosen, C.J. PPARγ: A circadian transcription factor in adipogenesis and osteogenesis. Nat. Rev. Endocrinol. 2010, 6, 629–636. [Google Scholar] [CrossRef] [Green Version]
- Bunney, P.E.; Zink, A.N.; Holm, A.A.; Billington, C.J.; Kotz, C.M. Orexin activation counteracts decreases in nonexercise activity thermogenesis (NEAT) caused by high-fat diet. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Kriebs, A.; Jordan, S.D.; Soto, E.; Henriksson, E.; Sandate, C.R.; Vaughan, M.E.; Chan, A.B.; Duglan, D.; Papp, S.J.; Huber, A.L.; et al. Circadian repressors CRY1 and CRY2 broadly interact with nuclear receptors and modulate transcriptional activity. Proc. Natl. Acad. Sci. USA 2017, 114, 8776–8781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gérard, C.; Goldbeter, A. Entrainment of the mammalian cell cycle by the circadian clock: Modeling two coupled cellular rhythms. PLoS Comput. Biol. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cho, H.; Yu, R.T.; Atkins, A.R.; Downes, M.; Evans, R.M. Nuclear receptors rock around the clock. EMBO Rep. 2014, 15, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, T. Cell cycle control factors and skeletal development. Jpn. Dent. Sci. Rev. 2013, 49, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Berman, S.D.; Yuan, T.L.; Miller, E.S.; Lee, E.Y.; Caron, A.; Lees, J.A. The Retinoblastoma Protein Tumor Suppressor Is Important for Appropriate Osteoblast Differentiation and Bone Development. Mol. Cancer Res. 2008, 6, 1440–1452. [Google Scholar] [CrossRef] [Green Version]
- Verlinden, L.; Eelen, G.; Beullens, I.; Van Camp, M.; Van Hummelen, P.; Engelen, K.; Van Hellemont, R.; Marchal, K.; De Moor, B.; Foijer, F.; et al. Characterization of the Condensin Component Cnap1 and Protein Kinase Melk as Novel E2F Target Genes Down-regulated by 1, 25-Dihydroxyvitamin D3. J. Biol. Chem. 2005, 280, 37319–37330. [Google Scholar] [CrossRef] [Green Version]
- Riediger, N.D.; Othman, R.A.; Suh, M.; Moghadasian, M.H. A Systemic Review of the Roles of n-3 Fatty Acids in Health and Disease. J. Am. Diet. Assoc. 2009, 109, 668–679. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | g/kg diet | Kcal/kg |
|---|---|---|
| Cornstarch | 397.486 | 1590 |
| Casein (≥85% protein) | 200 | 800 |
| Dextrinized cornstarch (90–94% tetrasaccharides) | 132 | 528 |
| Sucrose | 100 | 400 |
| Corn oil\flaxseed oil\fish oil | 70 | 630 |
| Fiber | 50 | |
| Mineral mix (AIN-93G-MX) | 35 | |
| Vitamin mix (AIN-93-VX) | 10 | |
| L-Cystine | 3 | 12 |
| Choline bitartrate (41.1% choline) | 2.5 | |
| Tert- butylhydroquinone | 0.014 |
| Fatty Acids (gr\mouse) | Diet | ||
|---|---|---|---|
| Control | Flaxseed | Fish | |
| SFAs | 1.68 | 1.05 | 3.49 |
| MUFAs | 3.21 | 1.88 | 2.65 |
| PUFAs | 5.87 | 7.69 | 3.54 |
| Total n-6 PUFAs | 5.74 | 1.77 | 0.5 |
| Total n-3 PUFAs | 0.12 | 5.89 | 3.01 |
| Fatty Acid | Liver | Serum | ||||
|---|---|---|---|---|---|---|
| Control (mg/100 mg) | Flaxseed (mg/100 mg) | Fish (mg/100 mg) | Control (mg/mL) | Flaxseed (mg/mL) | Fish (mg/mL) | |
| Total n-3 | 0.118 (0.02) b | 0.839 (0.15) a | 0.494 (0.10) b | 0.100 (0.03) b | 0.535 (0.07) a | 0.522 (0.09) a |
| 18:3 ALA (n-3) | 0.011 (0) b | 0.539 (0.12) a | 0.015 (0) b | 0.005 (0.00) b | 0.232 (0.03) a | 0.014 (0.00) b |
| 20:5 EPA (n-3) | 0.002 (0) b | 0.158 (0.02) a | 0.157 (0.04) a | 0.003 (0.00) c | 0.184 (0.03) b | 0.241 (0.03) a |
| 22:6 DHA (n-3) | 0.082 (0.02) c | 0.132 (0.02) b | 0.316 (0.05) a | 0.066 (0.03) b | 0.102 (0.01) b | 0.252 (0.05) a |
| Total n-6 | 1.710 (0.38) a | 0.449 (0.08) b | 0.274 (0.05) b | 1.381 (0.36) a | 0.587 (0.08) b | 0.430 (0.09) b |
| 18:2 LA (n-6) | 1.282 (0.34) a | 0.376 (0.07) b | 0.176 (0.04) b | 0.820 (0.31) a | 0.441 (0.07) b | 0.240 (0.06) b |
| 20:4 AA (n-6) | 0.343 (0.05) a | 0.059 (0.01) b | 0.086 (0) b | 0.480 (0.04) a | 0.101 (0.01) c | 0.147 (0.03) b |
| SFA | 1.434 (0.27) a | 0.921 (0.16) b | 1.038 (0.12) b | 0.984 (0.22) a | 0.737 (0.09) b | 0.787 (0.14) ab |
| 14:0 Mytistic | 0.030 (0.01) a | 0.016 (0) b | 0.025 (0) ab | 0.016 (0) a | 0.013 (0) a | 0.016 (0) a |
| 16:0 Palmitic | 1.104 (0.24) a | 0.640 (0.11) b | 0.747 (0.10) b | 0.658 (0.18) a | 0.433 (0.06) b | 0.541 (0.10) ab |
| 18:0 Stearic | 0.294 (0.03) a | 0.261 (0.04) a | 0.261 (0.01) a | 0.300 (0.03) a | 0.270 (0.03) a | 0.220 (0.03) b |
| 20:0 Arachidic | 0.001 (0) a | 0 (0) b | 0.001 (0) ab | 0.002 (0) a | 0 (0) a | 0 (0) a |
| MUFA | 1.619 (0.29) a | 0.815 (0.23) b | 0.615 (0.12)b | 0.582 (0.21) a | 0.372 (0.05) b | 0.389 (0.09) b |
| 16:1 Palmitoleic | 0.187 (0.04) a | 0.108 (0.03) b | 0.132 (0.04) b | 0.061 (0.01) ab | 0.046 (0.01) b | 0.079 (0.02) a |
| 18:1 Vaccenic | 0.003 (0) a | 0 (0) b | 0.002 (0) b | 0.008 (0.01) a | 0.004 (0.01) a | 0.002 (0) a |
| 18:1 Oleic (n-9) | 1.315 (0.24) a | 0.659 (0.18) b | 0.432 (0.06) b | 0.455 (0.18) a | 0.284 (0.03) b | 0.267 (0.06) b |
| Total lipids | 4.873 (0.82) a | 3.020 (0.60) b | 2.418 (0.37) b | 3.023 (0.82) a | 2.216 (0.28) b | 2.116 (0.41) b |
| Control | ||
| 18:3 ALA (n-3) | 1.70 | 0.60 |
| 20:5 EPA (n-3) | 0.05 | 0.11 |
| 22:6 DHA (n-3) | 0.37 | 4.55 |
| 18:2 LA (n-6) | 97.45 | 70.47 |
| 20:4 AA (n-6) | 0.00 | 18.89 |
| Flaxseed | ||
| 18:3 ALA (n-3) | 76.55 | 41.89 |
| 20:5 EPA (n-3) | 0.04 | 12.32 |
| 22:6 DHA (n-3) | 0.09 | 10.25 |
| 18:2 LA (n-6) | 22.76 | 29.20 |
| 20:4 AA (n-6) | 0.01 | 4.63 |
| Fish | ||
| 18:3 ALA (n-3) | 3.07 | 1.98 |
| 20:5 EPA (n-3) | 56.44 | 20.65 |
| 22:6 DHA (n-3) | 25.7 | 41.68 |
| 18:2 LA (n-6) | 8.61 | 23.16 |
| 20:4 AA (n-6) | 3.48 | 11.40 |
| 6 Weeks Old | 9 Weeks Old | |||||
| Width (µm) | Control | Flaxseed | Fish | Control | Flaxseed | Fish |
| Total | 119.9 (11.8) b | 134.0 (19) a | 128.2 (19.5) a | 89.8 (9.4) b | 101.0(7.4) a | 101.1 (10.6) a |
| PZ | 56.3 (9.9) b | 65.1 (13.1) a | 60.1 (13.8) ab | 44.4 (7.9) b | 51.8 (6.6) a | 51.2 (7.7) a |
| HZ | 63.6 (9.3) a | 68.9 (14.2) a | 68.0 (15.9) a | 45.4 (7.6) b | 49.2 (5.9) a | 49.9 (8.1) a |
| No. of cells | Control | Flaxseed | Fish | Control | Flaxseed | Fish |
| Total | 12.2 (0.4) a | 11.9 (0.7) a | 12.2 (1.2) a | 10.4 (0.8) a | 10.3 (0.7) a | 10.4 (0.5) a |
| PZ | 8.6 (0.6) a | 7.9 (0.6) a | 8.3 (0.6) a | 7.1 (0.6) a | 6.6 (0.7) a | 6.9 (0.1) a |
| HZ | 3.6 (0.2) a | 4.0 (0.5) a | 3.9 (0.8) a | 3.3 (0.3) a | 3.6 (0.2) a | 3.5 (0.5) a |
| 6 Weeks Old | 9 Weeks Old | |||||
| Bone Mechanical Properties | Control | Flaxseed | Fish | Control | Flaxseed | Fish |
| Stiffness (N/µ) | 0.031 (0.005) b | 0.038 (0.002) ab | 0.042 (0.009) a | 0.063 (0.013) | 0.062 (0.011) | 0.052 (0.01) |
| Max load (N) | 9.025 (0.82) b | 11.264 (1.566) a | 12.022 (1.137) a | 15.474 (1.826) | 15.082 (0.963) | 14.121 (2.905) |
| Yield point (N) | 5.07 (0.698) | 6.092 (1.537) | 7.228 (2.426) | 6.628 (1.668) | 7.05 (1.657) | 6.395 (1.037) |
| Young’s modulus | 0.405 (0.103) b | 0.522 (0.118) ab | 0.613 (0.178) a | 0.994(0.305) | 0.895(0.304) | 0.72(0.427) |
| Trabecular Bone Microarchitecture | Control | Flaxseed | Fish | Control | Flaxseed | Fish |
| BV/TV % | 8.04 (1.35) b | 10.28 (0.8) a | 8.81 (1.39) ab | 6.02 (1.06) b | 8.10 (1.32) a | 6.87 (1.82) ab |
| Tb.Th (mm) | 0.03 (0) | 0.04 (0) | 0.03 (0) | 0.04 (0) | 0.04 (0) | 0.04 (0) |
| Tb.Sp (mm) | 0.22 (0.01) | 0.20 (0.02) | 0.21 (0.03) | 0.27 (0.03) | 0.24 (0.03) | 0.26 (0.03) |
| Tb.N (1/mm) | 2.21 (0.33) b | 2.75 (0.41) a | 2.48 (0.46) ab | 1.54 (0.25) b | 2.00 (0.32) a | 1.76 (0.46) ab |
| Cortical Bone Microarchitecture | Control | Flaxseed | Fish | Control | Flaxseed | Fish |
| B.Ar/T.Ar (%) | 30.22 (1.88) | 31.67 (1.77) | 30.61 (2.79) | 33.18 (1.89) | 35.66 (2.06) | 34.52 (3.19) |
| M.Ar (mm2) | 1.39 (0.16) | 1.38 (0.11) | 1.36 (0.09) | 1.36 (0.1) | 1.26 (0.1) | 1.37 (0.11) |
| Cs.Th (mm) | 0.11 (0.01) | 0.12 (0) | 0.11 (0.01) | 0.12 (0.01) | 0.13 (0) | 0.13 (0.02) |
| BMD (gr/cm3) | 1.17 (0.07) | 1.21 (0.06) | 1.23 (0.08) | 1.33 (0.05) b | 1.30 (0.05) b | 1.41 (0.06) a |
| Comparison | Pathway | Molecules |
|---|---|---|
| Flaxseed vs. control | Circadian Rhythm Signaling | Bmal, bhlhe40, cry2, nr1d1, per2, per3, dpb |
| Glutathione Redox Reactions | Gstt1, gstt2/gstt2b | |
| Coagulation System | Plat, serpine1 | |
| Adipogenesis pathway | Bmal1, nr1d2, per2 | |
| Role of Osteoblasts, Osteoclasts and Chondrocytes | Adamts4, adamts5, nfatc2 | |
| Fish vs. control | Cell Cycle Control of Replication | Cdc6, cdc7, cdk1, cdk2, cdt1, dbf4, dna2, lig1, mcm2, mcm3, mcm4, mcm5, mcm6, mcm7, orc1, orc2, orc6, pcna, pold1, pole, prim1, top2a |
| Role of BRCA1 in DNA Damage Response | Bard1, blm, brca1, brca2, cdkn1a, e2f1, e2f2, e2f4, e2f7, e2f8, fancd2, fancl, gadd45a, mdc1, plk1, rad51, rb1, rbl1, rfc3, rfc4, topbp1 | |
| Heme Biosynthesis II | Alad, cpox, fech, hmbs, ppox, urod, uros | |
| Circadian Rhythm Signaling | Bmal1, cry2, nr1d1, per2, per3 | |
| Hedgehog Signaling | Ccnb1, cdk1, gli2, prkar2b | |
| Flaxseed vs. Fish | Heme Biosynthesis II | Alad, cpox, fech, ppox, urod, uros |
| Role of BRCA1 in DNA Damage Response | Brca1, brca2, e2f2, e2f4, fancd2, topbp1 | |
| Cell Cycle Control of Replication | Mcm3, mcm5, orc1, pole | |
| Cytokine Signaling | Blvrb, ccr5, il1rl1, socs3 | |
| Hypoxia Signaling | Ube2c, ube2l6, ube2o, ube2t |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozner, R.; Vernikov, J.; Griess-Fishheimer, S.; Travinsky, T.; Penn, S.; Schwartz, B.; Mesilati-Stahy, R.; Argov-Argaman, N.; Shahar, R.; Monsonego-Ornan, E. The Role of Omega-3 Polyunsaturated Fatty Acids from Different Sources in Bone Development. Nutrients 2020, 12, 3494. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113494
Rozner R, Vernikov J, Griess-Fishheimer S, Travinsky T, Penn S, Schwartz B, Mesilati-Stahy R, Argov-Argaman N, Shahar R, Monsonego-Ornan E. The Role of Omega-3 Polyunsaturated Fatty Acids from Different Sources in Bone Development. Nutrients. 2020; 12(11):3494. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113494
Chicago/Turabian StyleRozner, Reut, Janna Vernikov, Shelley Griess-Fishheimer, Tamar Travinsky, Svetlana Penn, Betty Schwartz, Ronit Mesilati-Stahy, Nurit Argov-Argaman, Ron Shahar, and Efrat Monsonego-Ornan. 2020. "The Role of Omega-3 Polyunsaturated Fatty Acids from Different Sources in Bone Development" Nutrients 12, no. 11: 3494. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113494