Rubi Fructus Water Extract Alleviates LPS-Stimulated Macrophage Activation via an ER Stress-Induced Calcium/CHOP Signaling Pathway



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of RF
2.3. Total Flavonoid Content of RF
2.4. Effects of RF on Cell Viability of RAW 264.7
2.5. Effects of RF on NO Production in RAW 264.7 Macrophages Stimulated with LPS
2.6. Effects of RF on Intracellular Calcium Release in RAW 264.7 Stimulated by LPS
2.7. Effects of RF on Cytokine Production in RAW 264.7 Cells Stimulated by LPS
2.8. Effects of RF on mRNA Expression in RAW 264.7 Cells Stimulated by LPS
2.8.1. Isolation of RNA
2.8.2. Determination of RNA Concentration
2.8.3. cDNA Synthesis
2.8.4. RT-qPCR Analysis
2.9. Effects of RF on Phosphorylation of p38 MAPK in RAW 264.7 Cells Stimulated by LPS
2.10. Statistics
3. Results
3.1. Determination of the Total Flavonoid Content of RF
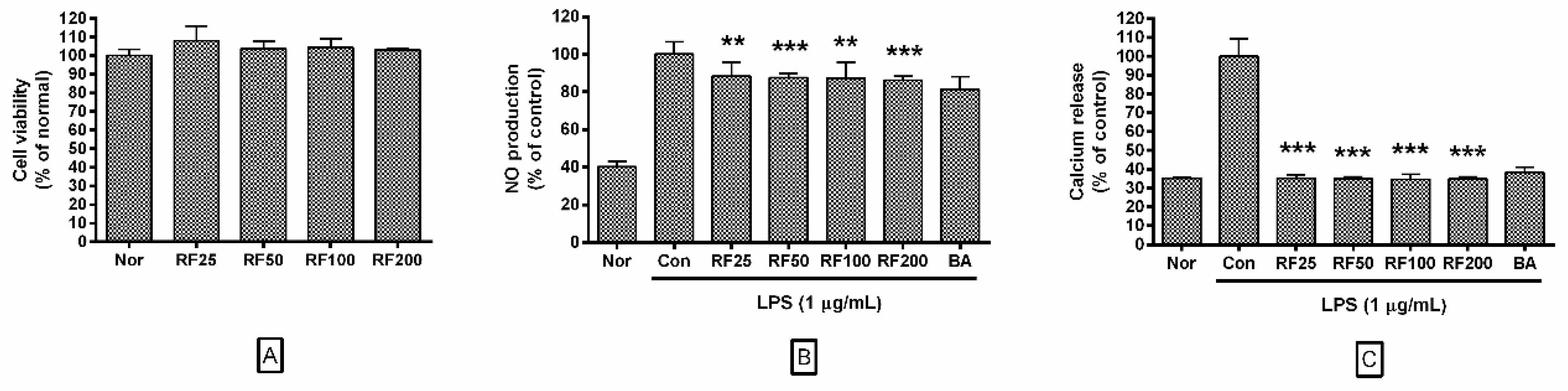
3.2. Effects of RF on Cell Viability
3.3. NO Production in RAW 264.7 Cells
3.4. Calcium Release in RAW 264.7 Cells
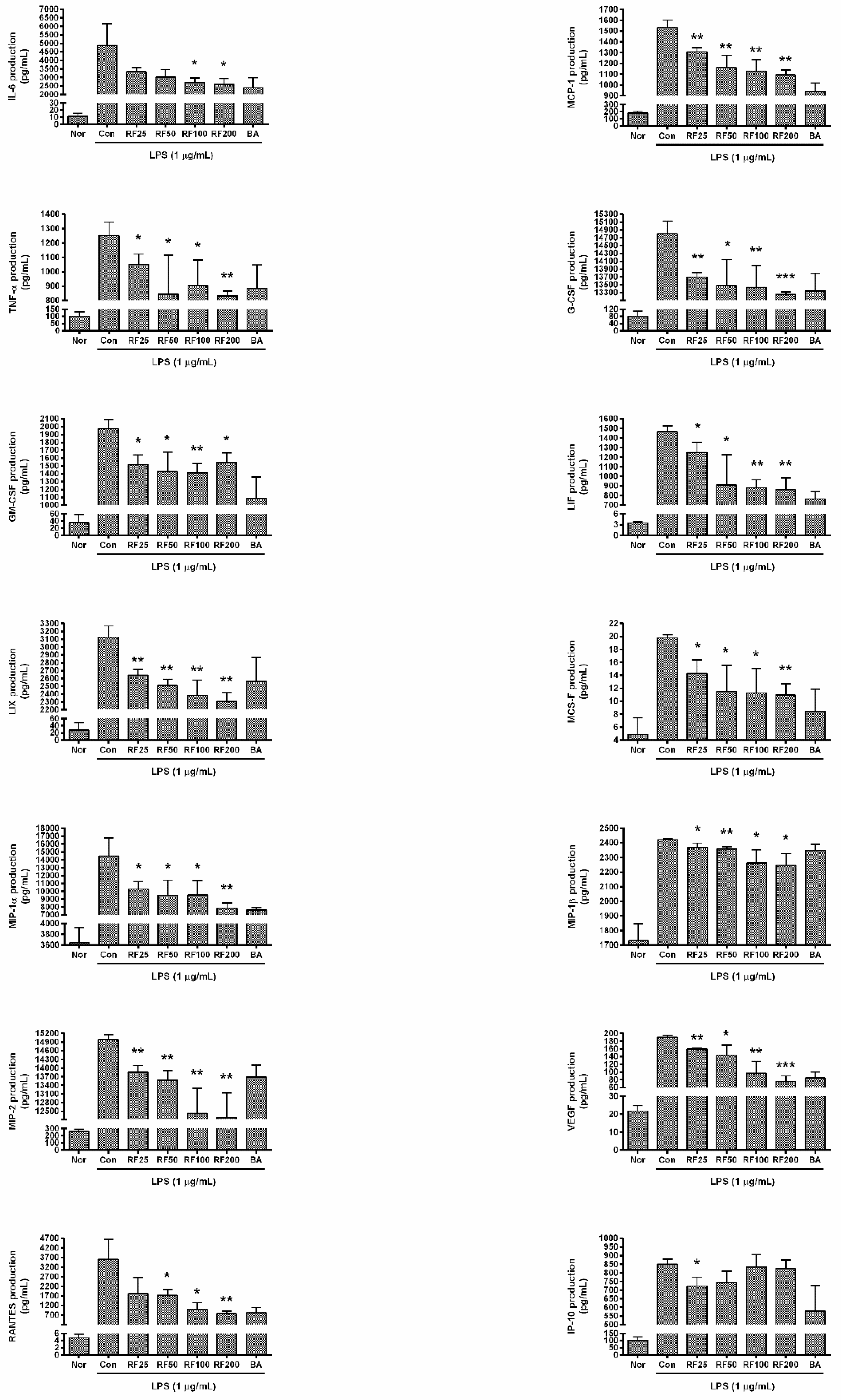
3.5. Cytokine Production in RAW 264.7 Cells
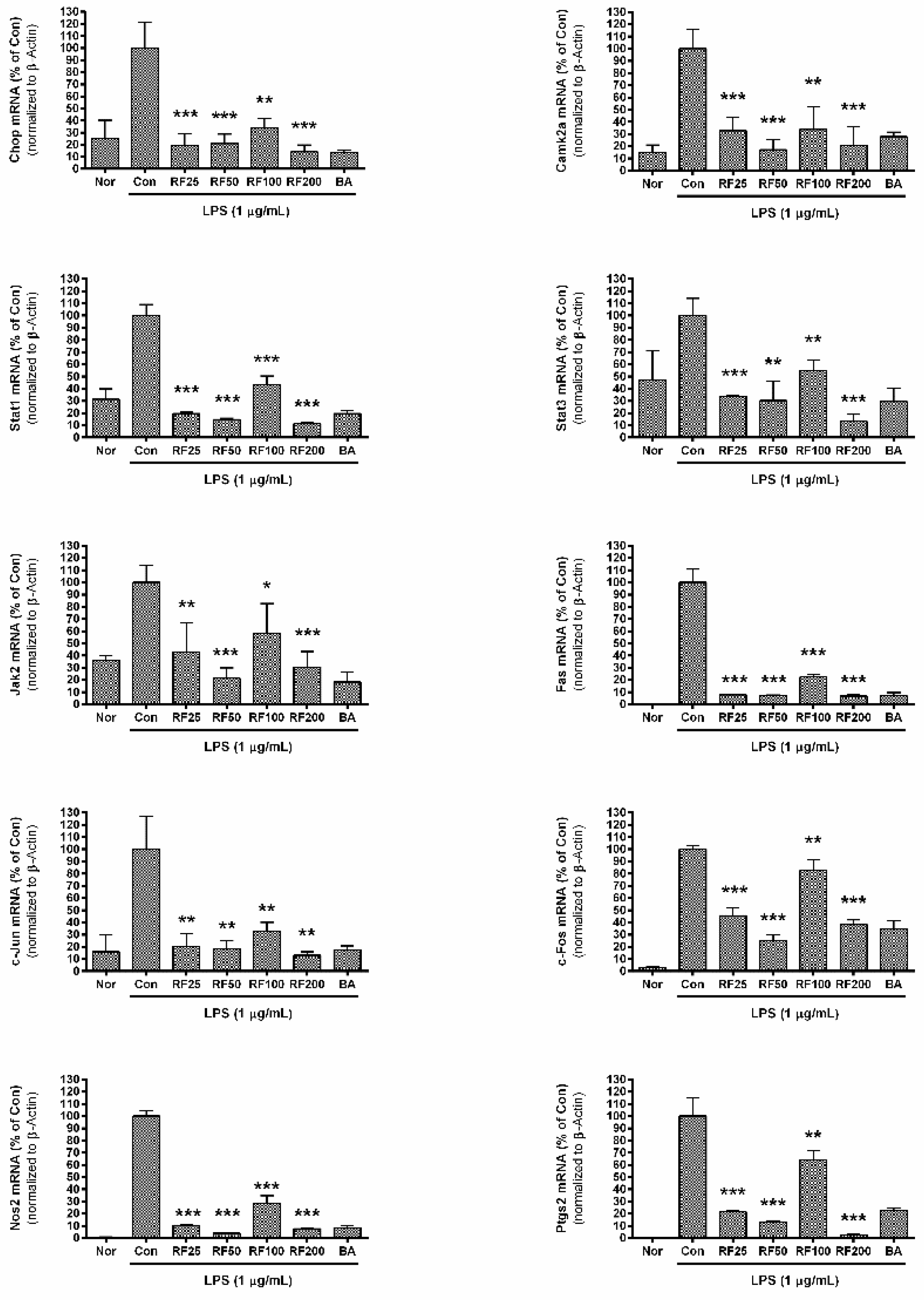
3.6. mRNA Expression in RAW 264.7 Cells
3.7. Phosphorylation of p38 MAPK in RAW 264.7
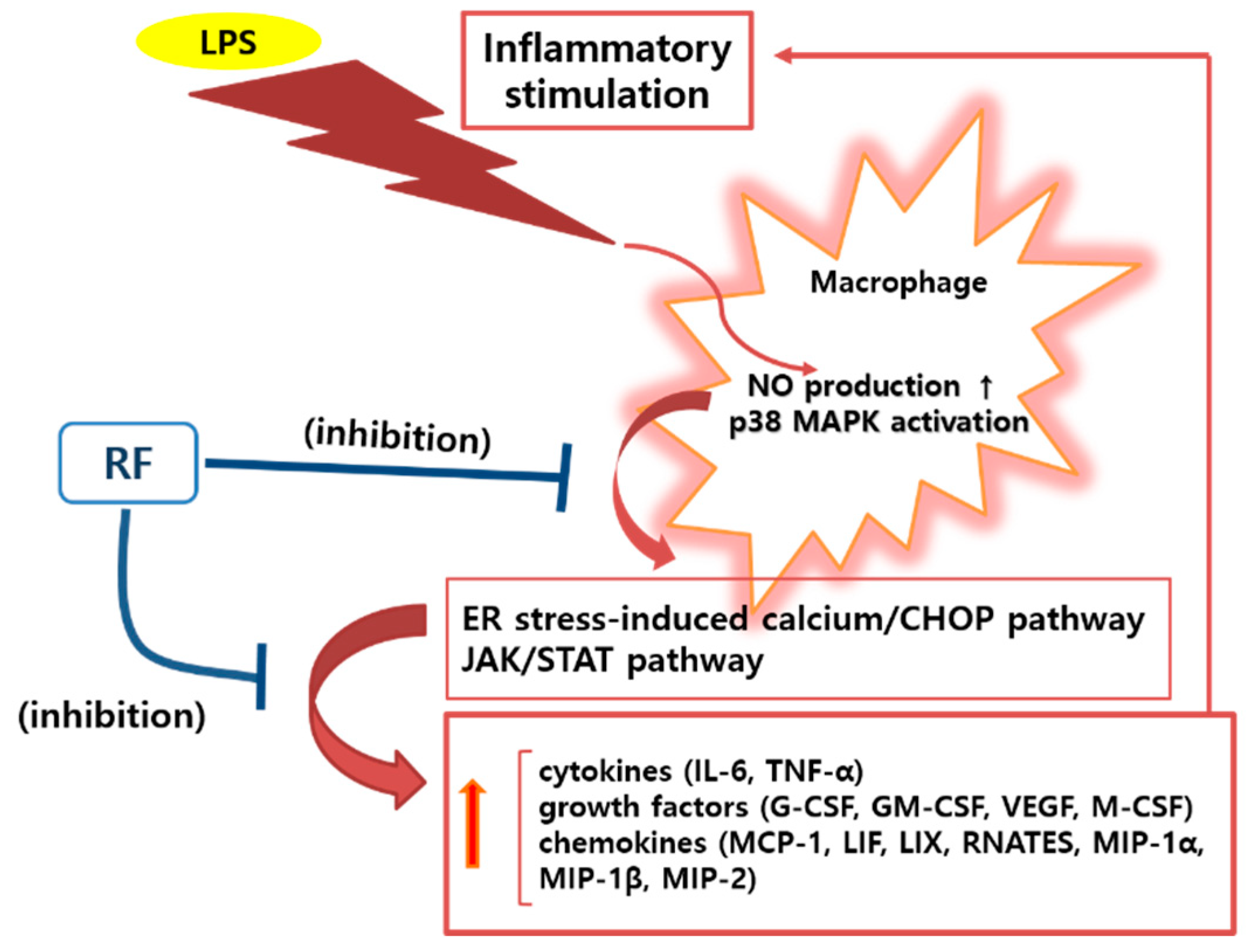
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| RF | Water extract of Rubi Fructus |
| LPS | Lipopolysaccharide |
| NO | Nitric Oxide |
| PGE2 | Prostaglandin E2 |
| IL | Interleukin |
| MCP | Monocyte chemotactic activating factor |
| TNF | Tumor necrosis factor |
| IgG2b | Immunoglobulin G2b |
| OD | Optical density |
| LIX | Lipopolysaccharide-induced CXC chemokine |
| LIF | Leukemia inhibitory factor |
| G-CSF | Granulocyte colony-stimulating factor |
| GM-CSF | Granulocyte macrophage colony-stimulating factor |
| VEGF | Vascular endothelial growth factor |
| M-CSF | Macrophage colony-stimulating factor |
| MIP | Macrophage inflammatory protein |
| RANTES | Regulated on activation, normal T cell expressed and secreted |
| IP | Interferon gamma-induced protein |
| ER | Endoplasmic reticulum |
| CHOP | C/EBP homologous protein |
| CAMK2a | Calcium/calmodulin dependent protein kinase II alpha |
| MAPK | Mitogen-activated protein kinase |
| JAK | Janus kinase |
| STAT | Signal transducers and activators of transcription |
| FAS | First apoptosis signal receptor |
| NOS | Nitric oxide synthase |
| PTGS | Prostaglandin-endoperoxide synthase |
| SIRS | Systemic inflammatory response syndrome |
| RE | Rutin equivalents |
| ERF | Ethanol extract of Rubi Fructus |
| WRF | Water extract of Rubi Fructus |
| DMEM | Dulbecco’s modified Eagle’s medium |
| HMSB | Handheld Magnetic Separation Block |
| SB | Staining Buffer |
| Nor | Normal group |
| MODS | Multiple organ dysfunction syndrome |
| JNK | Jun NH2-terminal kinase |
| NF | Nuclear factor |
| iNOS | Inducible nitric oxide synthase |
| COX-2 | Cyclooxygenase 2 |
| IC50 | Half maximal inhibitory concentration |
| ROS | Reactive oxygen species |
References
- Si-Tahar, M.; Touqui, L.; Chignard, M. Innate Immunity and Inflammation—Two Facets of the Same Anti-Infectious Reaction. Clin. Exp. Immunol. 2009, 156, 194–198. [Google Scholar] [CrossRef]
- Li, H.S.; Watowich, S.S. Innate Immune Regulation by STAT-Mediated Transcriptional Mechanisms. Immunol. Rev. 2014, 261, 84–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Nikrad, M.P.; Travanty, E.A.; Zhou, B.; Phang, T.; Gao, B.; Alford, T.; Ito, Y.; Nahreini, P.; Hartshorn, K.; et al. Innate Immune Response of Human Alveolar Macrophages during Influenza A Infection. PLoS ONE 2012, 7, e29879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arango Duque, G.; Descoteaux, A. Macrophage Cytokines: Involvement in Immunity and Infectious Diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabas, I.; Seimon, T.; Timmins, J.; Li, G.; Lim, W. Macrophage Apoptosis in Advanced Atherosclerosis. Ann. N. Y. Acad. Sci. 2009, 1173 (Suppl. 1), E40–E45. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Ma, S. The Cytokine Storm and Factors Determining the Sequence and Severity of Organ Dysfunction in Multiple Organ Dysfunction Syndrome. Am. J. Emerg. Med. 2008, 26, 711–715. [Google Scholar] [CrossRef]
- Zheng, W.R.; Li, E.C.; Peng, S.; Wang, X.S. Tu Youyou Winning the Nobel Prize: Ethical Research on the Value and Safety of Traditional Chinese Medicine. Bioethics 2020, 34, 166–171. [Google Scholar] [CrossRef]
- Lee, T.; Jung, W.M.; Lee, I.S.; Lee, Y.S.; Lee, H.; Park, H.J.; Kim, N.; Chae, Y. Data Mining of Acupoint Characteristics from the Classical Medical Text: DongUiBoGam of Korean Medicine. Evid. Based Complement. Altern. Med. 2014, 2014, 329563. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.Y.; Zhou, H.; Yang, Y.F.; Sang, B.S.; Liu, L. Current Policies and Measures on the Development of Traditional Chinese Medicine in China. Pharmacol. Res. 2020, 105187. [Google Scholar] [CrossRef]
- Bhandary, B.; Lee, H.Y.; Back, H.I.; Park, S.H.; Kim, M.G.; Kwon, J.W.; Song, J.Y.; Lee, H.K.; Kim, H.R.; Chae, S.W.; et al. Immature Rubus Coreanus Shows a Free Radical-Scavenging Effect and Inhibits Cholesterol Synthesis and Secretion in Liver Cells. Indian J. Pharm. Sci. 2012, 74, 211–216. [Google Scholar]
- Lee, J.H.; Bae, S.Y.; Oh, M.; Seok, J.H.; Kim, S.; Chung, Y.B.; Gowda, K.G.; Mun, J.Y.; Chung, M.S.; Kim, K.H. Antiviral Effects of Black Raspberry (Rubus Coreanus) Seed Extract and its Polyphenolic Compounds on Norovirus Surrogates. Biosci. Biotechnol. Biochem. 2016, 80, 1196–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, K.H.; Lee, J.Y.; Park, J.Y.; Jang, G.Y.; Kim, H.D.; Lee, Y.S.; Kim, D.H. Differences in Anti-Inflammatory Effect of Immature and Mature of Rubus Coreanus Fruits on LPS-Induced RAW 264.7 Macrophages Via NF-kappaB Signal Pathways. BMC Complement. Altern. Med. 2019, 19, 89. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Cho, S.M.; Park, E.; Lee, S.M.; Kim, Y.; Auh, J.H.; Choi, H.K.; Lim, S.; Lee, S.C.; Kim, J.H. Anti-Inflammatory Effects of Rubus Coreanus Miquel through Inhibition of NF-kappaB and MAP Kinase. Nutr. Res. Pract. 2014, 8, 501–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kim, C.K.; Lee, K.S.; Kim, J.H.; Hwang, H.; Jeoung, D.; Choe, J.; Won, M.H.; Lee, H.; Ha, K.S.; et al. Aqueous Extract of Unripe Rubus Coreanus Fruit Attenuates Atherosclerosis by Improving Blood Lipid Profile and Inhibiting NF-kappaB Activation Via Phase II Gene Expression. J. Ethnopharmacol. 2013, 146, 515–524. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, J.Y.; Kim, H.J.; Kim, D.H.; Lee, T.H.; Kang, M.S.; Park, W. Anti-Inflammatory Effects of Angelica Sinensis (Oliv.) Diels Water Extract on RAW 264.7 Induced with Lipopolysaccharide. Nutrients 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Mocan, A.; Vlase, L.; Raita, O.; Hanganu, D.; Păltinean, R.; Dezsi, Ş.; Gheldiu, A.; Oprean, R.; Crişan, G. Comparative Studies on Antioxidant Activity and Polyphenolic Content of Lycium Barbarum L. and Lycium Chinense Mill, Leaves. Pak. J. Pharm. Sci. 2015, 28 (Suppl. 4), 1511–1515. [Google Scholar]
- Shahidullah, A.; Lee, J.Y.; Kim, Y.J.; Halimi, S.M.A.; Rauf, A.; Kim, H.J.; Kim, B.Y.; Park, W. Anti-Inflammatory Effects of Diospyrin on Lipopolysaccharide-Induced Inflammation using RAW 264.7 Mouse Macrophages. Biomedicines 2020, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Park, B.S.; Lee, J.O. Recognition of Lipopolysaccharide Pattern by TLR4 Complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef] [Green Version]
- Cross, A.S. Anti-Endotoxin Vaccines: Back to the Future. Virulence 2014, 5, 219–225. [Google Scholar] [CrossRef]
- Deutschman, C.S.; Tracey, K.J. Sepsis: Current Dogma and New Perspectives. Immunity 2014, 40, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the Eye of the Cytokine Storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujishima, S. Organ Dysfunction as a New Standard for Defining Sepsis. Inflamm. Regen. 2016, 36, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meegan, J.E.; Shaver, C.M.; Putz, N.D.; Jesse, J.J.; Landstreet, S.R.; Lee, H.N.R.; Sidorova, T.N.; McNeil, J.B.; Wynn, J.L.; Cheung-Flynn, J.; et al. Cell-Free Hemoglobin Increases Inflammation, Lung Apoptosis, and Microvascular Permeability in Murine Polymicrobial Sepsis. PLoS ONE 2020, 15, e0228727. [Google Scholar] [CrossRef] [PubMed]
- Hauser, B.; Matejovic, M.; Radermacher, P. Nitric Oxide, Leukocytes and Microvascular Permeability: Causality or Bystanders? Crit. Care 2008, 12, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, B.T.; Chambers, R.C.; Liu, K.D. Acute Respiratory Distress Syndrome. N. Engl. J. Med. 2017, 377, 562–572. [Google Scholar] [CrossRef]
- Kilbourn, R.G.; Traber, D.L.; Szabo, C. Nitric Oxide and Shock. Dis. Mon. 1997, 43, 277–348. [Google Scholar] [CrossRef]
- Teijaro, J.R. Cytokine Storms in Infectious Diseases. Semin. Immunopathol. 2017, 39, 501–503. [Google Scholar] [CrossRef] [Green Version]
- Wahid, F.; Jung, H.; Khan, T.; Hwang, K.H.; Park, J.S.; Chang, S.C.; Khan, M.A.; Kim, Y.Y. Effects of Rubus Coreanus Extract on Visual Processes in Bullfrog’s Eye. J. Ethnopharmacol. 2011, 138, 333–339. [Google Scholar] [CrossRef]
- Kim, C.R.; Choi, S.J.; Oh, S.S.; Kwon, Y.K.; Lee, N.Y.; Park, G.G.; Kim, Y.J.; Heo, H.J.; Jun, W.J.; Park, C.S.; et al. Rubus Coreanus Miquel Inhibits Acetylcholinesterase Activity and Prevents Cognitive Impairment in a Mouse Model of Dementia. J. Med. Food 2013, 16, 785–792. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.; Lee, H.; Lim, H.; Park, S.; Lee, J.; Do, S. Effect of Rubus Coreanus Extracts on Diabetic Osteoporosis by Simultaneous Regulation of Osteoblasts and Osteoclasts. Menopause 2012, 19, 1043–1051. [Google Scholar] [CrossRef]
- Teng, H.; Lin, Q.; Li, K.; Yuan, B.; Song, H.; Peng, H.; Yi, L.; Wei, M.C.; Yang, Y.C.; Battino, M.; et al. Hepatoprotective Effects of Raspberry (Rubus Coreanus Miq.) Seed Oil and its Major Constituents. Food Chem. Toxicol. 2017, 110, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; You, Y.; Yoon, H.G.; Kim, K.; Park, J.; Kim, S.; Ho, J.N.; Lee, J.; Shim, S.; Jun, W. Fatigue-Alleviating Effect on Mice of an Ethanolic Extract from Rubus Coreanus. Biosci. Biotechnol. Biochem. 2011, 75, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.K.; Choi, H.R.; Cho, J.S.; Cho, S.M.; Ha, K.C.; Kim, T.H.; Ryu, H.Y.; Lee, Y.I. Inhibitory Effects of Rubi Fructus Extracts on Hepatic Steatosis Development in High-Fat Diet-Induced Obese Mice. Mol. Med. Rep. 2014, 10, 1821–1827. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, J.; Lee, S.M.; Lee, H.A.; Park, S.; Kim, Y.; Kim, J.H. Chemopreventive Effects of Rubus Coreanus Miquel on Prostate Cancer. Biosci. Biotechnol. Biochem. 2012, 76, 737–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, T.Y.; Kim, S.H.; Lee, E.S.; Eom, D.O.; Kim, H.M. Action of Rubus Coreanus Extract on Systemic and Local Anaphylaxis. Phytother. Res. 2002, 16, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.S.; Yang, W.M.; Chang, M.S.; Park, W.; Kim, D.R.; Lee, H.K.; Kim, W.N.; Park, S.K. Effects of Rubus Coreanus on Sperm Parameters and cAMP-Responsive Element Modulator (CREM) Expression in Rat Testes. J. Ethnopharmacol. 2007, 114, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Oh, S.M.; Lim, S.S.; Lee, Y.S.; Shin, H.K.; Oh, Y.S.; Choe, N.H.; Park, J.H.; Kim, J.K. Induction of Heme Oxygenase-1 Mediates the Anti-Inflammatory Effects of the Ethanol Extract of Rubus Coreanus in Murine Macrophages. Biochem. Biophys. Res. Commun. 2006, 351, 146–152. [Google Scholar] [CrossRef]
- Wang, X.Z.; Ron, D. Stress-Induced Phosphorylation and Activation of the Transcription Factor CHOP (GADD153) by p38 MAP Kinase. Science 1996, 272, 1347–1349. [Google Scholar] [CrossRef] [Green Version]
- Endo, M.; Mori, M.; Akira, S.; Gotoh, T. C/EBP Homologous Protein (CHOP) is Crucial for the Induction of Caspase-11 and the Pathogenesis of Lipopolysaccharide-Induced Inflammation. J. Immunol. 2006, 176, 6245–6253. [Google Scholar] [CrossRef] [Green Version]
- Stout, B.A.; Melendez, K.; Seagrave, J.; Holtzman, M.J.; Wilson, B.; Xiang, J.; Tesfaigzi, Y. STAT1 Activation Causes Translocation of Bax to the Endoplasmic Reticulum during the Resolution of Airway Mucous Cell Hyperplasia by IFN-Gamma. J. Immunol. 2007, 178, 8107–8116. [Google Scholar] [CrossRef] [Green Version]
- Mori, M. Regulation of Nitric Oxide Synthesis and Apoptosis by Arginase and Arginine Recycling. J. Nutr. 2007, 137, 1616S–1620S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmins, J.M.; Ozcan, L.; Seimon, T.A.; Li, G.; Malagelada, C.; Backs, J.; Backs, T.; Bassel-Duby, R.; Olson, E.N.; Anderson, M.E.; et al. Calcium/calmodulin-Dependent Protein Kinase II Links ER Stress with Fas and Mitochondrial Apoptosis Pathways. J. Clin. Investig. 2009, 119, 2925–2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.K.; Cheong, K.J.; Kim, H.Y.; Cheong, J. Endoplasmic Reticulum Stress Induced by Hepatitis B Virus X Protein Enhances Cyclo-Oxygenase 2 Expression Via Activating Transcription Factor 4. Biochem. J. 2011, 435, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Lee, W.S.; Shin, J.S.; Jang, D.S.; Lee, K.T. Xanthotoxin Suppresses LPS-Induced Expression of iNOS, COX-2, TNF-Alpha, and IL-6 Via AP-1, NF-kappaB, and JAK-STAT Inactivation in RAW 264.7 Macrophages. Int. Immunopharmacol. 2017, 49, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Guha, M.; Mackman, N. LPS Induction of Gene Expression in Human Monocytes. Cell. Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name 1 | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Chop | CCACCACACCTGAAAGCAG | TCCTCATACCAGGCTTCCA |
| Camk2a | AGCCATCCTCACCACTAT | ATTCCTTCACGCCATCATT |
| Stat1 | TGAGATGTCCCGGATAGTGG | CGCCAGAGAGAAATTCGTGT |
| Stat3 | GTCTGCAGAGT TCAAGCACCT | TCCTCAGTCACGATCAAGGAG |
| Jak2 | TTGGTTTTGAATTATGGTGTCTGT | TCCAAATTTTACAAATTCTTGAACC |
| Fas | CGCTGTTTTCCCTTGCTG | CCTTGAGTATGAACTCTTAACTGTGAG |
| c-Jun | ACTGGGTTGCGACCTGAC | CAATAGGCCGCTGCTCTC |
| c-Fos | AGAGCGGGAATGGTGAAGA | TCTTCCTCTTCAGGAGATAGCTG |
| Nos2 | TGGAGGTTCTGGATGAGAGC | AATGTCCAGGAAGTAGGTGAGG |
| Ptgs2 | TCAAACAGTTTCTCTACAACAACTCC | ACATTTCTTCCCCCAGCAA |
| β-actin | CTAAGGCCAACCGTGAAAAG | ACCAGAGGCATACAGGGACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-H.; Lee, J.-Y.; Kim, Y.-J.; Kim, H.-J.; Park, W. Rubi Fructus Water Extract Alleviates LPS-Stimulated Macrophage Activation via an ER Stress-Induced Calcium/CHOP Signaling Pathway. Nutrients 2020, 12, 3577. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113577
Kim D-H, Lee J-Y, Kim Y-J, Kim H-J, Park W. Rubi Fructus Water Extract Alleviates LPS-Stimulated Macrophage Activation via an ER Stress-Induced Calcium/CHOP Signaling Pathway. Nutrients. 2020; 12(11):3577. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113577
Chicago/Turabian StyleKim, Do-Hoon, Ji-Young Lee, Young-Jin Kim, Hyun-Ju Kim, and Wansu Park. 2020. "Rubi Fructus Water Extract Alleviates LPS-Stimulated Macrophage Activation via an ER Stress-Induced Calcium/CHOP Signaling Pathway" Nutrients 12, no. 11: 3577. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113577