Identification and Characterization of Edible Cricket Peptides on Hypertensive and Glycemic In Vitro Inhibition and Their Anti-Inflammatory Activity on RAW 264.7 Macrophage Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cricket Protein Hydrolysate Preparation
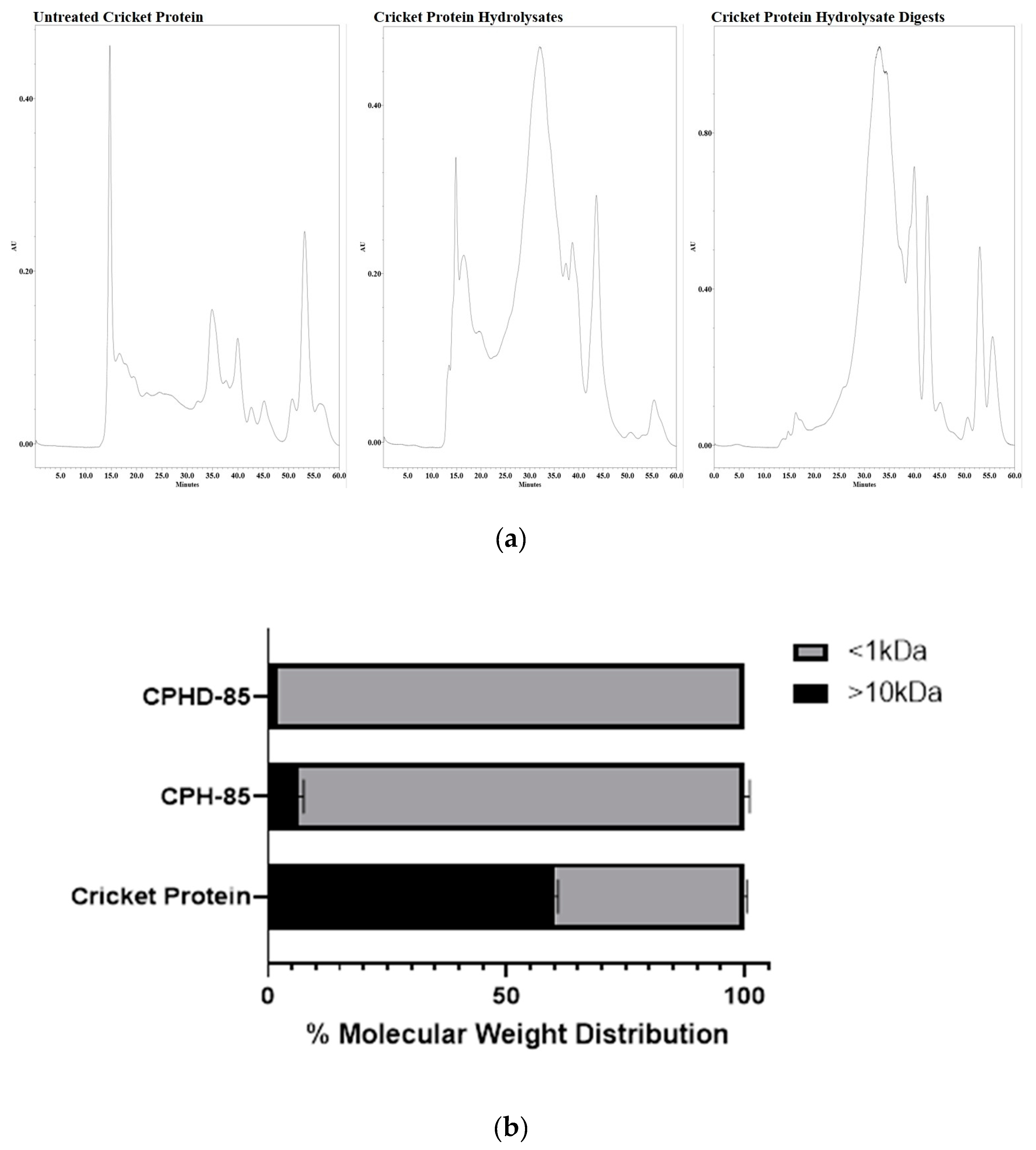
2.3. Molecular Weight Distribution of CPH and CPHD
2.4. CPHD Peptide Fractionation
2.4.1. Size Exclusion Chromatography (SEC)
2.4.2. Reverse-Phased–High-Performance Liquid Chromatography (RP–HPLC)
2.4.3. Ion Exchange Chromatography (IEX)
2.5. Anti-Glycemic, Anti-Hypertensive, and Anti-Inflammatory Assays
2.5.1. Dipeptidyl Peptidase-IV (DPP-IV) Inhibition
2.5.2. Alpha-Amylase Inhibition
2.5.3. Alpha-Glucosidase Inhibition
2.5.4. Angiotensin Converting Enzyme II Inhibition
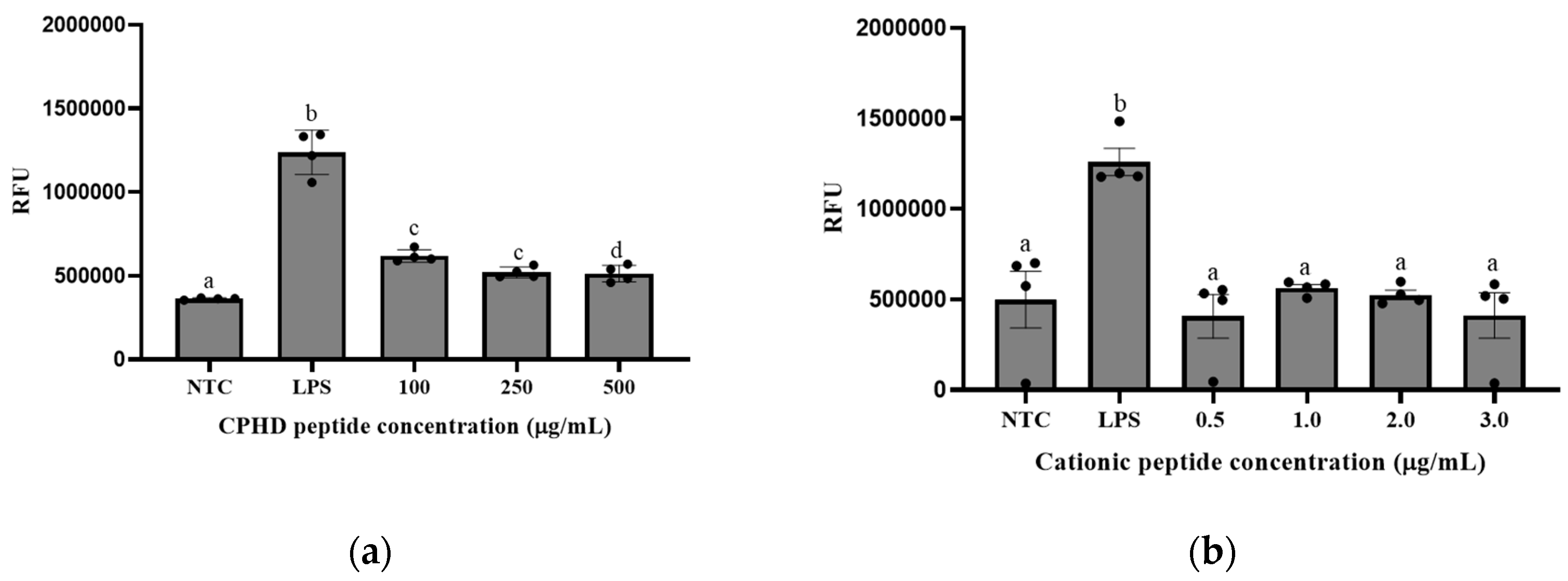
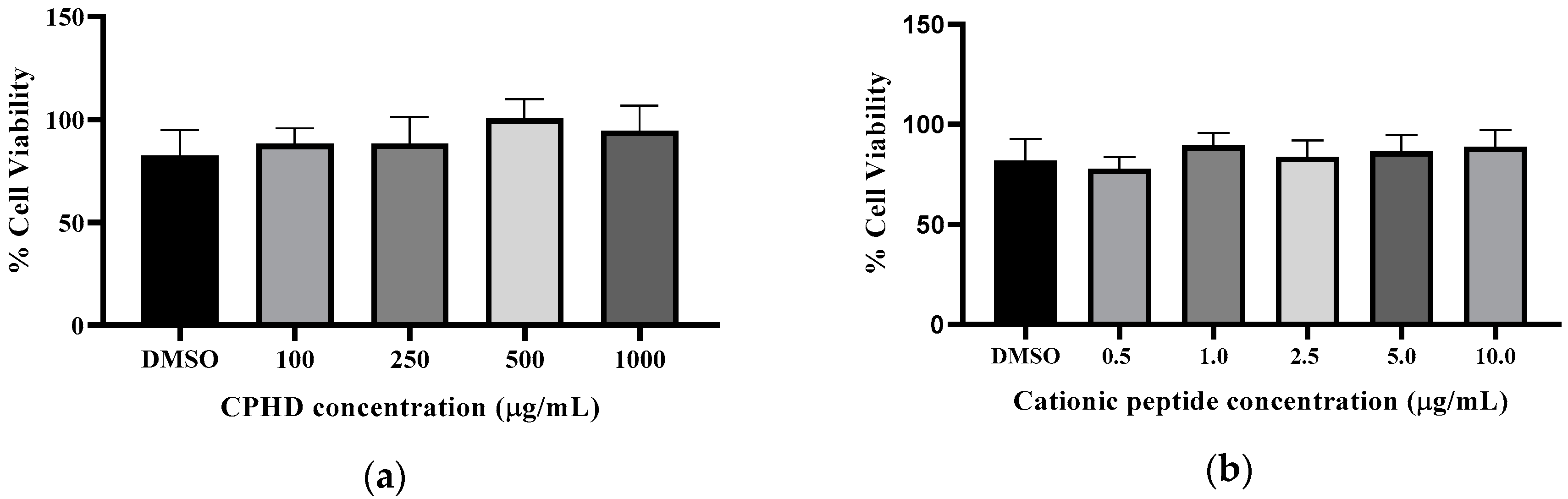
2.5.5. RAW 264.7-NF-κB Cell Culture, Cell Viability, and Anti-inflammatory Response
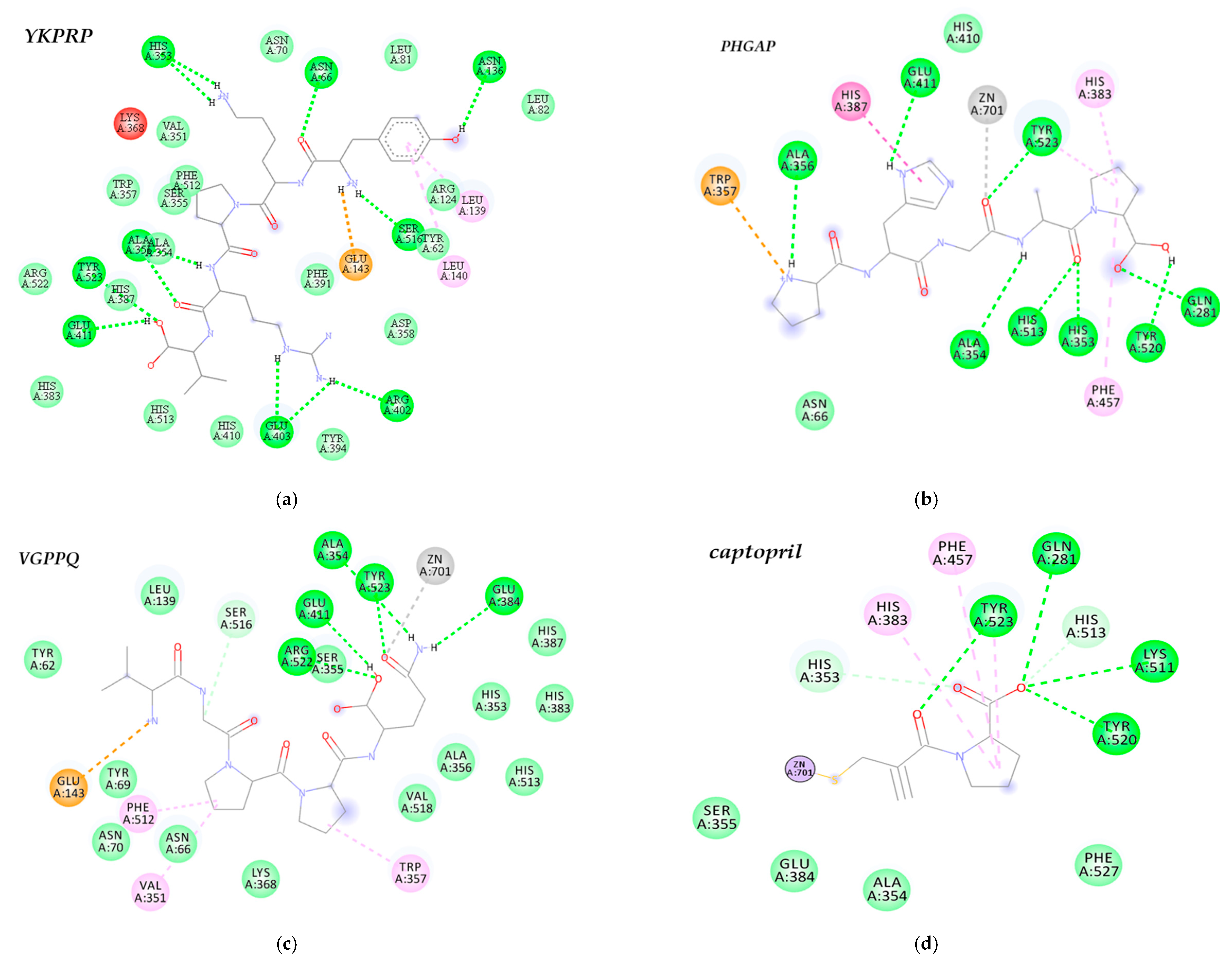
2.6. Identification of Cationic Peptides and Molecular Docking
2.7. Statistical Analysis
3. Results
3.1. Molecular Weight Distribution of CPH and CPHD
3.2. CPHD Peptide Fractionation
3.3. Cationic Peptide Fractions Shows Multifunctional Bioactivity
NF-κB Expression in Macrophage (RAW 264.7) Cells
3.4. Peptide Identification and Molecular Docking against ACE
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Estruch, R.; Martínez-González, M.Á.; Corella, D.; Salas-Salvadó, J.; Ruiz-Gutiérrez, V.; Covas, M.I.; Fiol, M.; Gómez-Gracia, E.; López-Sabater, M.C.; Vinyoles, E.; et al. Effects of a Mediterranean-Style Diet on Cardiovascular Risk Factors: A randomized trial. Ann. Intern. Med. 2006, 145, 1–11. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food Protein-Derived Bioactive Peptides: Production, Processing, and Potential Health Benefits. J. Food Sci. 2011, 77, R11–R24. [Google Scholar] [CrossRef]
- Hall, F.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Luna, G.C.; Martin-Gonzalez, F.S.; Mauer, L.; Liceaga, A.M. Cricket (Acheta domesticus) protein hydrolysates’ impact on the physicochemical, structural and sensory properties of tortillas and tortilla chips. J. Insects Food Feed 2020, 1–12. [Google Scholar] [CrossRef]
- Çabuk, B.; Yılmaz, B. Fortification of traditional egg pasta (erişte) with edible insects: Nutritional quality, cooking properties and sensory characteristics evaluation. J. Food Sci. Technol. 2020, 57, 2750–2757. [Google Scholar] [CrossRef] [PubMed]
- Vercruysse, L.; Smagghe, G.; Matsui, T.; Van Camp, J. Purification and identification of an angiotensin I converting enzyme (ACE) inhibitory peptide from the gastrointestinal hydrolysate of the cotton leafworm, Spodoptera littoralis. Process. Biochem. 2008, 43, 900–904. [Google Scholar] [CrossRef]
- Hall, F.; Johnson, P.; Liceaga, A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. 2018, 262, 39–47. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M. Antioxidant and Anti-Inflammatory Activities of Hydrolysates and Peptide Fractions Obtained by Enzymatic Hydrolysis of Selected Heat-Treated Edible Insects. Nutrients 2017, 9, 970. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; Fitzgerald, R.J. Unlocking the biological potential of proteins from edible insects through enzymatic hydrolysis: A review. Innov. Food Sci. Emerg. Technol. 2017, 43, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, H.J. Food-derived Bioactive Peptides-Opportunities for Designing Future Foods. Curr. Pharm. Des. 2003, 9, 1297–1308. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Vercruysse, L.; Smagghe, G.; Beckers, T.; Van Camp, J. Antioxidative and ACE inhibitory activities in enzymatic hydrolysates of the cotton leafworm, Spodoptera littoralis. Food Chem. 2009, 114, 38–43. [Google Scholar] [CrossRef]
- Vercruysse, L.; Van Camp, J.; Morel, N.; Rougé, P.; Herregods, G.; Smagghe, G. Ala-Val-Phe and Val-Phe: ACE inhibitory peptides derived from insect protein with antihypertensive activity in spontaneously hypertensive rats. Peptides 2010, 31, 482–488. [Google Scholar] [CrossRef]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. Changes in the antioxidant activity of loach (Misgurnus anguillicaudatus) protein hydrolysates during a simulated gastrointestinal digestion. Food Chem. 2010, 120, 810–816. [Google Scholar] [CrossRef]
- Ketnawa, S.; Wickramathilaka, M.; Liceaga, A.M. Changes on antioxidant activity of microwave-treated protein hydrolysates after simulated gastrointestinal digestion: Purification and identification. Food Chem. 2018, 254, 36–46. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Dipeptidyl peptidase-IV inhibitory activity of dairy protein hydrolysates. Int. Dairy J. 2012, 25, 97–102. [Google Scholar] [CrossRef]
- Awosika, T.O.; Aluko, R.E. Inhibition of the in vitro activities of α-amylase, α-glucosidase and pancreatic lipase by yellow field pea (Pisum sativum L.) protein hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Shobana, S.; Sreerama, Y.; Malleshi, N. Composition and enzyme inhibitory properties of finger millet (Eleusine coracana L.) seed coat phenolics: Mode of inhibition of α-glucosidase and pancreatic amylase. Food Chem. 2009, 115, 1268–1273. [Google Scholar] [CrossRef]
- Alvarez, O.M.; Batista, I.; Ramos, C.; Montero, P. Enhancement of ACE and prolyl oligopeptidase inhibitory potency of protein hydrolysates from sardine and tuna by-products by simulated gastrointestinal digestion. Food Funct. 2016, 7, 2066–2073. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 2018, 095505. [Google Scholar] [CrossRef]
- Mosley, A.L.; Florens, L.; Wen, Z.; Washburn, M.P. A label free quantitative proteomic analysis of the Saccharomyces cerevisiae nucleus. J. Proteom. 2009, 72, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Toalá, J.E.; Liceaga, A.M. Identification of chia seed (Salvia hispanica L.) peptides with enzyme inhibition activity towards skin-aging enzymes. Amino Acids 2020, 52, 1149–1159. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Chaudhary, K.; Chauhan, J.S.; Nagpal, G.; Kumar, R.; Sharma, M.; Raghava, G.P. An in silico platform for predicting, screening and designing of antihypertensive peptides. Sci. Rep. 2015, 5, srep12512. [Google Scholar] [CrossRef] [Green Version]
- Amaya, J.A.G.; Cabrera, D.Z.; Matallana, A.M.; Arevalo, K.G.; Guevara-Pulido, J. In-silico design of new enalapril analogs (ACE inhibitors) using QSAR and molecular docking models. Inform. Med. Unlocked 2020, 19, 100336. [Google Scholar] [CrossRef]
- Qian, B.; Tian, C.; Huo, J.; Ding, Z.; Xu, R.; Zhu, J.; Yu, L.; Villarreal, O.D. Design and evaluation of four novel tripeptides as potent angiotensin converting enzyme (ACE) inhibitors with anti-hypertension activity. Peptides 2019, 122, 170171. [Google Scholar] [CrossRef]
- Petrie, J.R.; Guzik, T.J.; Touyz, R.M. Diabetes, Hypertension, and Cardiovascular Disease: Clinical Insights and Vascular Mechanisms. Can. J. Cardiol. 2018, 34, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Sowers, J.R.; Epstein, M.; Frohlich, E.D. Diabetes, Hypertension, and Cardiovascular Disease. Hypertension 2001, 37, 1053–1059. [Google Scholar] [CrossRef] [Green Version]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta Biomembr. 2008, 1778, 357–375. [Google Scholar] [CrossRef] [Green Version]
- Monaco, C. Nuclear factor κB: A potential therapeutic target in atherosclerosis and thrombosis. Cardiovasc. Res. 2004, 61, 671–682. [Google Scholar] [CrossRef]
- Lazennec, G.; Richmond, A. Chemokines and chemokine receptors: New insights into cancer-related inflammation. Trends Mol. Med. 2010, 16, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guha, S.; Majumder, K. Structural-features of food-derived bioactive peptides with anti-inflammatory activity: A brief review. J. Food Biochem. 2018, 43, e12531. [Google Scholar] [CrossRef] [PubMed]
- Danneels, E.L.; Gerlo, S.; Heyninck, K.; Van Craenenbroeck, K.; De Bosscher, K.; Haegeman, G.; De Graaf, D.C. How the Venom from the Ectoparasitoid Wasp Nasonia vitripennis Exhibits Anti-Inflammatory Properties on Mammalian Cell Lines. PLoS ONE 2014, 9, e96825. [Google Scholar] [CrossRef] [Green Version]
- Zielińska, E.; Baraniak, B.; Karaś, M. Identification of antioxidant and anti-inflammatory peptides obtained by simulated gastrointestinal digestion of three edible insects species (Gryllodes sigillatus, Tenebrio molitor, Schistocerca gragaria). Int. J. Food Sci. Technol. 2018, 53, 2542–2551. [Google Scholar] [CrossRef]
- Ng, S.M.S.; Yap, J.M.; Lau, Q.Y.; Ng, F.M.; Ong, E.H.Q.; Barkham, T.; Teo, J.W.P.; Alfatah, M.; Kong, K.W.; Hoon, S.; et al. Structure-activity relationship studies of ultra-short peptides with potent activities against fluconazole-resistant Candida albicans. Eur. J. Med. Chem. 2018, 150, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; He, R.; Aluko, R.E. Kinetics and Molecular Docking Studies of the Inhibitions of Angiotensin Converting Enzyme and Renin Activities by Hemp Seed (Cannabis sativa L.) Peptides. J. Agric. Food Chem. 2014, 62, 4135–4144. [Google Scholar] [CrossRef]
- He, R.; Aluko, R.E.; Ju, X.-R. Evaluating Molecular Mechanism of Hypotensive Peptides Interactions with Renin and Angiotensin Converting Enzyme. PLoS ONE 2014, 9, e91051. [Google Scholar] [CrossRef]
- Howard, J.A.K.; Yufit, D.S.; Chetina, O.V.; Teat, S.J.; Capelli, S.C.; Pattison, P. Crystal structure of the insect neuropeptide proctolin. Org. Biomol. Chem. 2010, 8, 5110–5112. [Google Scholar] [CrossRef]
- Siviter, R.J.; Nachman, R.J.; Dani, M.P.; Keen, J.N.; Shirras, A.D.; Isaac, R.E. Peptidyl dipeptidases (Ance and Acer) of Drosophila melanogaster: Major differences in the substrate specificity of two homologs of human angiotensin I-converting enzyme. Peptides 2002, 23, 2025–2034. [Google Scholar] [CrossRef]
- Hall, F.; Liceaga, A.M. Effect of microwave-assisted enzymatic hydrolysis of cricket (Gryllodes sigillatus) protein on ACE and DPP-IV inhibition and tropomyosin-IgG binding. J. Funct. Foods 2020, 64, 103634. [Google Scholar] [CrossRef]
- Zielińska, E.; Karaś, M.; Baraniak, B.; Jakubczyk, A. Evaluation of ACE, α-glucosidase, and lipase inhibitory activities of peptides obtained by in vitro digestion of selected species of edible insects. Eur. Food Res. Technol. 2020, 246, 1361–1369. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | ACE (%) | DPP-IV (%) | α-Glucosidase (%) | |
|---|---|---|---|---|
| Size exclusion | S1 | 32.3 ± 0.31 a | 19.3 ± 2.52 a | 62.2 ± 1.09 a |
| S2 | 28.8 ± 0.02 b | 16.98 ± 0.30 a | 25.9 ± 2.54 b | |
| S3 | 47.5 ± 0.87 c | 20.6 ± 0.61 a | 83.6 ± 1.14 c | |
| S4 | 73.9 ± 1.12 d | 68.0 ± 2.52 b | 89.5 ± 0.91 d | |
| Reverse-phase | F1 | 25.4 ± 0.6 e | 5.1 ± 0.01 c | 9.4 ± 0.36 e |
| F2 | 29.4 ± 0.04 g | 15.3 ± 2.70 d | 11.6 ± 0.04 f | |
| F3 | 67.2 ± 0.12 h | 18.4 ± 0.59 a | 8.4 ± 0.47 g | |
| F4 | 32.2 ± 0.19 a | 17.1 ± 1.61 a | 7.3 ± 1.51 h | |
| F5 | 47.4 ± 0.5 c | 21.0 ± 1.96 a | 22.3 ± 2.79 i | |
| F6 | 93.4 ± 0.08 i | 26.98 ± 1.63 e | 48.2 ± 5.79 j | |
| Positive inhibitor | Captopril | 98.0 ± 0.01 j | n.a | n.a |
| Ile-Leu-Pro | n.a | 88.9 ±6.05 f | n.a | |
| Acarbose | n.a | n.a | 70.9 ± 0.43 k |
| IEX Fraction | α-Amylase (µg/mL) | α-Glucosidase (µg/mL) | ACE (µg/mL) |
|---|---|---|---|
| Cationic Peptides | 18.537 | 13.902 | 1.922 |
| Anionic Peptides | 5082.75 | 76.623 | 509.062 |
| Ligand | Affinity Energy (kJ/mol) | Zn (II) Coordination | Interaction with ACE Active Site Residues * | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Glu384 | Tyr523 | Ala354 | Gln281 | Tyr520 | Lys511 | His513 | His353 | |||
| Captopril | −24.26 | sulfhydryl group of captopril | + | + | + | + | + | + | + | + |
| YKPRP | −39.75 | None | − | + | + | − | − | − | + | + |
| PHGAP | −36.82 | carboxylic acid group of glycine | − | + | + | + | + | − | + | + |
| VGPPQ | −36.87 | carboxylic acid group of lysine | + | + | + | − | − | − | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hall, F.; Reddivari, L.; Liceaga, A.M. Identification and Characterization of Edible Cricket Peptides on Hypertensive and Glycemic In Vitro Inhibition and Their Anti-Inflammatory Activity on RAW 264.7 Macrophage Cells. Nutrients 2020, 12, 3588. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113588
Hall F, Reddivari L, Liceaga AM. Identification and Characterization of Edible Cricket Peptides on Hypertensive and Glycemic In Vitro Inhibition and Their Anti-Inflammatory Activity on RAW 264.7 Macrophage Cells. Nutrients. 2020; 12(11):3588. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113588
Chicago/Turabian StyleHall, Felicia, Lavanya Reddivari, and Andrea M. Liceaga. 2020. "Identification and Characterization of Edible Cricket Peptides on Hypertensive and Glycemic In Vitro Inhibition and Their Anti-Inflammatory Activity on RAW 264.7 Macrophage Cells" Nutrients 12, no. 11: 3588. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113588