Functional Deficits in Gut Microbiome of Young and Middle-Aged Adults with Prediabetes Apparent in Metabolizing Bioactive (Poly)phenols



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
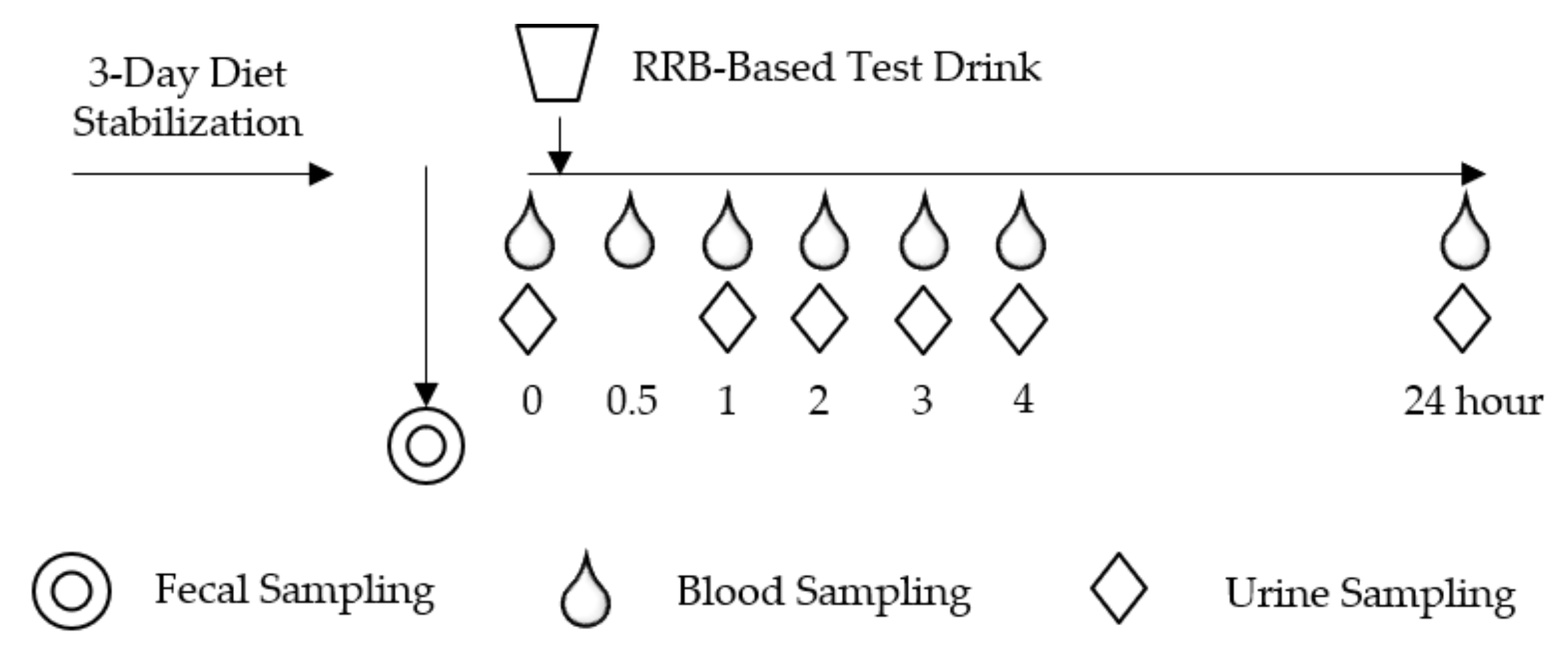
2.2. Study Design and Procedure
2.3. DNA Extraction and Whole Genome Shotgun Sequencing
2.4. Dietary Assessment
2.5. Metabolic Health Indices Analysis
2.6. Blood and Urine Microbial (Poly)phenolic Metabolites Analysis
2.7. Statistical Analysis
3. Results
3.1. Subject Demographics, Dietary Assessment and Metabolic Health Characteristics
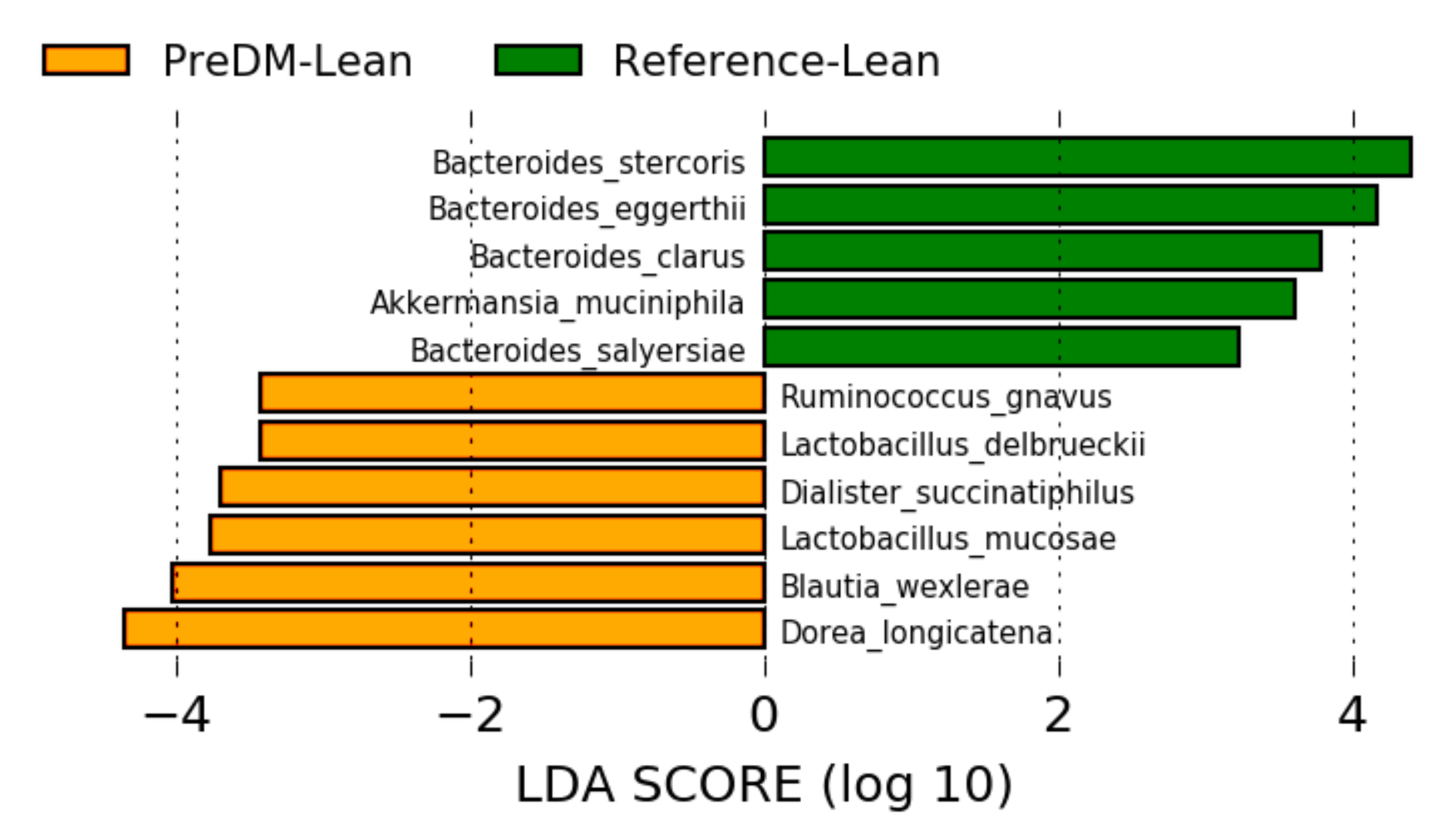
3.2. Gut Microbiome Composition
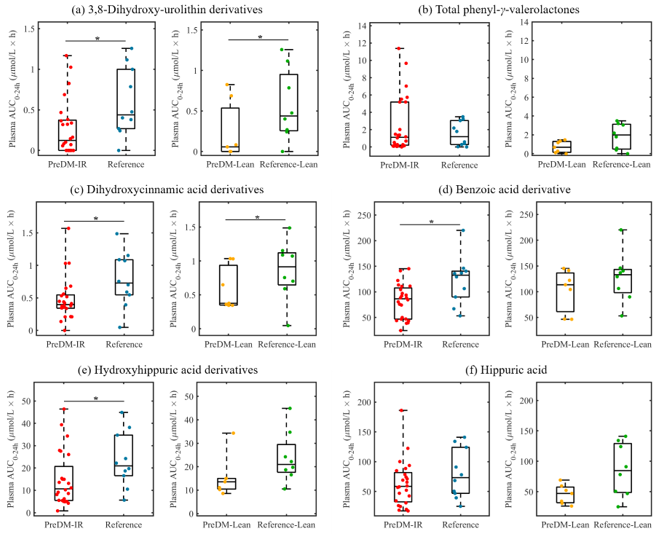
3.3. Predicted and Experimental Analyses of Gut Metabolomics
3.4. Putative Associations between Gut Microbiome Composition and Metabolic Health Indices and Microbial (Poly)phenolic Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federation. International Diabetes Federation: IDF Diabetes Atlas—9th Edition. Available online: https://www.diabetesatlas.org/upload/resources/material/20200302_133351_IDFATLAS9e-final-web.pdf (accessed on 14 September 2020).
- Abdul-Ghani, M.A.; Matsuda, M.; Balas, B.; DeFronzo, R.A. Muscle and Liver Insulin Resistance Indexes Derived From the Oral Glucose Tolerance Test. Diabetes Care 2007, 30, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human Gut Microbiota Changes Reveal the Progression of Glucose Intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef] [PubMed]
- Allin, K.H.; Tremaroli, V.; Caesar, R.; Jensen, B.A.H.; Damgaard, M.T.F.; Bahl, M.I.; Licht, T.R.; Hansen, T.H.; Nielsen, T.; Dantoft, T.M.; et al. Aberrant intestinal microbiota in individuals with prediabetes. Diabetologia 2018, 61, 810–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, H.; Ren, H.; Lu, Y.; Fang, C.; Hou, G.; Yang, Z.; Chen, B.; Yang, F.; Zhao, Y.; Shi, Z.; et al. Distinct gut metagenomics and metaproteomics signatures in prediabetics and treatment-naïve type 2 diabetics. EBioMedicine 2019, 47, 373–383. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. National Diabetes Statistics Report, 2020; US Department of Health and Human Services: Atlanta, GA, USA, 2020. [Google Scholar]
- Castaner, O.; Goday, A.; Park, Y.-M.; Lee, S.-H.; Magkos, F.; Shiow, S.-A.T.E.; Schröder, H. The Gut Microbiome Profile in Obesity: A Systematic Review. Int. J. Endocrinol. 2018, 2018, 4095789. [Google Scholar] [CrossRef]
- Franzosa, E.A.; Sirota-Madi, A.; Avila-Pacheco, J.; Fornelos, N.; Haiser, H.J.; Reinker, S.; Vatanen, T.; Hall, A.B.; Mallick, H.; McIver, L.J.; et al. Gut microbiome structure and metabolic activity in inflammatory bowel disease. Nat. Microbiol. 2019, 4, 293–305. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Mena, P.; Calani, L.; Borges, G.; Pereira-Caro, G.; Bresciani, L.; Del Rio, D.; Lean, M.E.J.; Crozier, A. New insights into the bioavailability of red raspberry anthocyanins and ellagitannins. Free Radic. Biol. Med. 2015, 89, 758–769. [Google Scholar] [CrossRef]
- Zhang, X.; Sandhu, A.; Edirisinghe, I.; Burton-Freeman, B. An exploratory study of red raspberry (Rubus idaeus L.) (poly)phenols/metabolites in human biological samples. Food Funct. 2018, 9, 806–818. [Google Scholar] [CrossRef]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Red Raspberries and Their Bioactive Polyphenols: Cardiometabolic and Neuronal Health Links. Adv. Nutr. 2016, 7, 44–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moazzen, H.; Alizadeh, M. Effects of Pomegranate Juice on Cardiovascular Risk Factors in Patients with Metabolic Syndrome: A Double-Blinded, Randomized Crossover Controlled Trial. Plant Foods Hum. Nutr. 2017, 72, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Betts, N.M.; Nguyen, A.; Newman, E.D.; Fu, D.; Lyons, T.J. Freeze-Dried Strawberries Lower Serum Cholesterol and Lipid Peroxidation in Adults with Abdominal Adiposity and Elevated Serum Lipids. J. Nutr. 2014, 144, 830–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, T.; Wang, B.; Yang, Q.; de Avila, J.M.; Zhu, M.-J.; You, J.; Chen, D.; Du, M. Raspberry promotes brown and beige adipocyte development in mice fed high-fat diet through activation of AMP-activated protein kinase (AMPK) α1. J. Nutr. Biochem. 2018, 55, 157–164. [Google Scholar] [CrossRef]
- Forbes-Hernández, T.Y.; Cianciosi, D.; Ansary, J.; Mezzetti, B.; Bompadre, S.; Quiles, J.L.; Giampieri, F.; Battino, M. Strawberry (Fragaria × ananassa cv. Romina) methanolic extract promotes browning in 3T3-L1 cells. Food Funct. 2020, 11, 297–304. [Google Scholar] [CrossRef]
- De Ferrars, R.M.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.A.; Cassidy, A.; Kay, C.D. The pharmacokinetics of anthocyanins and their metabolites in humans. Br. J. Pharmacol. 2014, 171, 3268–3282. [Google Scholar] [CrossRef] [Green Version]
- Lila, M.A.; Burton-Freeman, B.; Grace, M.; Kalt, W. Unraveling Anthocyanin Bioavailability for Human Health. Annu. Rev. Food Sci. Technol. 2016, 7, 375–393. [Google Scholar] [CrossRef]
- Li, C.; Ford, E.S.; McGuire, L.C.; Mokdad, A.H.; Little, R.R.; Reaven, G.M. Trends in Hyperinsulinemia Among Nondiabetic Adults in the U.S. Diabetes Care 2006, 29, 2396–2402. [Google Scholar] [CrossRef] [Green Version]
- Xiao, D.; Zhu, L.; Edirisinghe, I.; Fareed, J.; Brailovsky, Y.; Burton-Freeman, B. Attenuation of Postmeal Metabolic Indices with Red Raspberries in Individuals at Risk for Diabetes: A Randomized Controlled Trial. Obesity 2019, 27, 542–550. [Google Scholar] [CrossRef]
- Subar, A.F.; Thompson, F.E.; Potischman, N.; Forsyth, B.H.; Buday, R.; Richards, D.; McNutt, S.; Hull, S.G.; Guenther, P.M.; Schatzkin, A.; et al. Formative Research of a Quick List for an Automated Self-Administered 24-Hour Dietary Recall. J. Am. Diet. Assoc. 2007, 107, 1002–1007. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzosa, E.A.; McIver, L.J.; Rahnavard, G.; Thompson, L.R.; Schirmer, M.; Weingart, G.; Lipson, K.S.; Knight, R.; Caporaso, J.G.; Segata, N.; et al. Species-level functional profiling of metagenomes and metatranscriptomes. Nat. Methods 2018, 15, 962–968. [Google Scholar] [CrossRef]
- Leonard, S.R.; Mammel, M.K.; Lacher, D.W.; Elkins, C.A. Application of Metagenomic Sequencing to Food Safety: Detection of Shiga Toxin-Producing Escherichia coli on Fresh Bagged Spinach. Appl. Environ. Microbiol. 2015, 81, 8183–8191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallick, H.; Franzosa, E.A.; Mclver, L.J.; Banerjee, S.; Sirota-Madi, A.; Kostic, A.D.; Clish, C.B.; Vlamakis, H.; Xavier, R.J.; Huttenhower, C. Predictive metabolomic profiling of microbial communities using amplicon or metagenomic sequences. Nat. Commun. 2019, 10, 3136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knopfholz, J.; Disserol, C.C.D.; Pierin, A.J.; Schirr, F.L.; Streisky, L.; Takito, L.L.; Massucheto Ledesma, P.; Faria-Neto, J.R.; Olandoski, M.; da Cunha, C.L.P.; et al. Validation of the Friedewald Formula in Patients with Metabolic Syndrome. Cholesterol 2014, 2014, 261878. [Google Scholar] [CrossRef] [Green Version]
- Cai, G.; Shi, G.; Xue, S.; Lu, W. The atherogenic index of plasma is a strong and independent predictor for coronary artery disease in the Chinese Han population. Medicine 2017, 96, e8058. [Google Scholar] [CrossRef]
- Wallace, T.M.; Levy, J.C.; Matthews, D.R. Use and abuse of HOMA modeling. Diabetes Care 2004, 27, 1487–1495. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Sandhu, A.; Edirisinghe, I.; Burton-Freeman, B.M. Plasma and Urinary (Poly)phenolic Profiles after 4-Week Red Raspberry (Rubus idaeus L.) Intake with or without Fructo-Oligosaccharide Supplementation. Molecules 2020, 25, 4777. [Google Scholar] [CrossRef]
- Kay, C.D.; Clifford, M.N.; Mena, P.; McDougall, G.J.; Andres-Lacueva, C.; Cassidy, A.; Del Rio, D.; Kuhnert, N.; Manach, C.; Pereira-Caro, G.; et al. Recommendations for standardizing nomenclature for dietary (poly)phenol catabolites. Am. J. Clin. Nutr. 2020, 112, 1051–1068. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 9 January 2020).
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanita Body Fat Ranges for Standard Adults. Available online: https://www.tanita.com/data/BodyFatPercentageChart.pdf?rev=CE2E (accessed on 14 September 2020).
- Finucane, M.M.; Sharpton, T.J.; Laurent, T.J.; Pollard, K.S. A Taxonomic Signature of Obesity in the Microbiome? Getting to the Guts of the Matter. PLoS ONE 2014, 9, e84689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, P.W.; Jeffery, I.B. Gut microbiota and aging. Science 2015, 350, 1214–1215. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.S.; Das, M.; Jeffery, I.B.; O’Toole, P.W. Adjusting for age improves identification of gut microbiome alterations in multiple diseases. Elife 2020, 9, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Meslier, V.; Laiola, M.; Roager, H.M.; De Filippis, F.; Roume, H.; Quinquis, B.; Giacco, R.; Mennella, I.; Ferracane, R.; Pons, N.; et al. Mediterranean diet intervention in overweight and obese subjects lowers plasma cholesterol and causes changes in the gut microbiome and metabolome independently of energy intake. Gut 2020, 69, 1258–1268. [Google Scholar] [CrossRef] [Green Version]
- Cox, A.J.; Zhang, P.; Bowden, D.W.; Devereaux, B.; Davoren, P.M.; Cripps, A.W.; West, N.P. Increased intestinal permeability as a risk factor for type 2 diabetes. Diabetes Metab. 2017, 43, 163–166. [Google Scholar] [CrossRef]
- Pedersen, C.; Ijaz, U.Z.; Gallagher, E.; Horton, F.; Ellis, R.J.; Jaiyeola, E.; Duparc, T.; Russell-Jones, D.; Hinton, P.; Cani, P.D.; et al. Fecal Enterobacteriales enrichment is associated with increased in vivo intestinal permeability in humans. Physiol. Rep. 2018, 6, e13649. [Google Scholar] [CrossRef]
- Kang, Y.; Yang, G.; Zhang, S.; Ross, C.F.; Zhu, M.-J. Goji Berry Modulates Gut Microbiota and Alleviates Colitis in IL-10-Deficient Mice. Mol. Nutr. Food Res. 2018, 62, e1800535. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, W.; Wang, J.; Shi, J.; Sun, Y.; Wang, W.; Ning, G.; Liu, R.; Hong, J. Akkermansia muciniphila improves metabolic profiles by reducing inflammation in chow diet-fed mice. J. Mol. Endocrinol. 2017, 58, 1–14. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Geerlings, S.; Kostopoulos, I.; de Vos, W.; Belzer, C. Akkermansia muciniphila in the Human Gastrointestinal Tract: When, Where, and How? Microorganisms 2018, 6, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sovran, B.; Hugenholtz, F.; Elderman, M.; Van Beek, A.A.; Graversen, K.; Huijskes, M.; Boekschoten, M.V.; Savelkoul, H.F.J.; De Vos, P.; Dekker, J.; et al. Age-associated Impairment of the Mucus Barrier Function is Associated with Profound Changes in Microbiota and Immunity. Sci. Rep. 2019, 9, 1437. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; Beltrán, D.; García-Villalba, R.; Espín, J.C.; Tomás-Barberán, F.A. Description of urolithin production capacity from ellagic acid of two human intestinal Gordonibacter species. Food Funct. 2014, 5, 1779–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás-Barberán, F.A.; García-Villalba, R.; González-Sarrías, A.; Selma, M.V.; Espín, J.C. Ellagic Acid Metabolism by Human Gut Microbiota: Consistent Observation of Three Urolithin Phenotypes in Intervention Trials, Independent of Food Source, Age, and Health Status. J. Agric. Food Chem. 2014, 62, 6535–6538. [Google Scholar] [CrossRef]
- Romo-Vaquero, M.; Cortés-Martín, A.; Loria-Kohen, V.; Ramírez-de-Molina, A.; García-Mantrana, I.; Collado, M.C.; Espín, J.C.; Selma, M.V. Deciphering the Human Gut Microbiome of Urolithin Metabotypes: Association with Enterotypes and Potential Cardiometabolic Health Implications. Mol. Nutr. Food Res. 2019, 63, e1800958. [Google Scholar] [CrossRef]
- Selma, M.V.; González-Sarrías, A.; Salas-Salvadó, J.; Andrés-Lacueva, C.; Alasalvar, C.; Örem, A.; Tomás-Barberán, F.A.; Espín, J.C. The gut microbiota metabolism of pomegranate or walnut ellagitannins yields two urolithin-metabotypes that correlate with cardiometabolic risk biomarkers: Comparison between normoweight, overweight-obesity and metabolic syndrome. Clin. Nutr. 2018, 37, 897–905. [Google Scholar] [CrossRef]
- Mena, P.; Bresciani, L.; Brindani, N.; Ludwig, I.A.; Pereira-Caro, G.; Angelino, D.; Llorach, R.; Calani, L.; Brighenti, F.; Clifford, M.N.; et al. Phenyl-γ-valerolactones and phenylvaleric acids, the main colonic metabolites of flavan-3-ols: Synthesis, analysis, bioavailability, and bioactivity. Nat. Prod. Rep. 2019, 36, 714–752. [Google Scholar] [CrossRef]
- Márquez Campos, E.; Stehle, P.; Simon, M.C. Microbial Metabolites of Flavan-3-Ols and Their Biological Activity. Nutrients 2019, 11, 2260. [Google Scholar] [CrossRef] [Green Version]
- Mena, P.; Ludwig, I.A.; Tomatis, V.B.; Acharjee, A.; Calani, L.; Rosi, A.; Brighenti, F.; Ray, S.; Griffin, J.L.; Bluck, L.J.; et al. Inter-individual variability in the production of flavan-3-ol colonic metabolites: Preliminary elucidation of urinary metabotypes. Eur. J. Nutr. 2019, 58, 1529–1543. [Google Scholar] [CrossRef] [Green Version]
- Afshin, A.; Sur, P.J.; Fay, K.A.; Cornaby, L.; Ferrara, G.; Salama, J.S.; Mullany, E.C.; Abate, K.H.; Abbafati, C.; Abebe, Z.; et al. Health effects of dietary risks in 195 countries, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item Name | RRBtest Drink |
|---|---|
| Quantity (g) | 414 |
| Energy (kcal) | 361 |
| Protein (g) | 2.1 |
| Fat (g) | 1.4 |
| Carbohydrate (g) | 85 |
| Fiber (g) | 8 |
| Sugar (g) | 75 |
| RRB sugar | 10 |
| Added sugar | 65 |
| Total (poly)phenols (mg) | 388 |
| Anthocyanin (mg) | 237 |
| Ellagitannin/ellagic acid (mg) | 125 |
| Nutrient and Food Group | PreDM-IR (n = 26) | Reference (n = 10) | p Value |
|---|---|---|---|
| Energy (kcal) | 1917 ± 142 | 1832 ± 193 | NS |
| Protein (g) | 86 ± 6 | 93 ± 14 | NS |
| Total fat (g) | 80 ± 7 | 62 ± 8 | NS |
| Carbohydrate (g) | 213 ± 18 | 233 ± 22 | NS |
| Sugars, total (g) | 84 ± 10 | 70 ± 7 | NS |
| Added sugar (g) | 12 ± 2 | 6 ± 1 | NS |
| Fiber, total dietary (g) | 16 ± 2 | 24 ± 4 | 0.02 |
| Total fruits (cup) | 0.8 ± 0.2 | 1.0 ± 0.3 | NS |
| Total vegetable (cup) | 1.6 ± 0.2 | 2.0 ± 0.5 | NS |
| Total fruit and vegetable (cup) | 2.3 ± 0.3 | 3.0 ± 0.5 | 0.05 |
| Metabolic Health Indices | PreDM-IR (n = 26) | Reference (n = 10) | p-Value 2 | PreDM-Lean (n = 7) 3 | Reference-Lean (n = 8) 3 | p-Value 4 |
|---|---|---|---|---|---|---|
| Age (years) | 34 ± 11 | 31 ± 9 | NS | 28 ± 12 | 30.5 ± 10 | NS |
| Female: Male | 12:14 | 7:3 | NS | 2:5 | 6:2 | NS |
| CAU/AA/AS/HIS | 8:6:9:3 | 3:2:3:2 | NS | 1:1:5:0 | 2:1:3:2 | NS |
| Fasting glucose (mmol/L) | 5.8 ± 0.1 | 4.9 ± 0.1 | <0.0001 | 5.6 ± 0.2 | 4.9 ± 0.1 | 0.008 |
| Fasting insulin (pmol/L) | 88.2 ± 8.7 | 34.0 ± 4.1 | <0.0001 | 67.0 ± 6.8 | 35.0 ± 4.5 | 0.01 |
| HOMA-IR | 3.3 ± 0.3 | 1.1 ± 0.1 | <0.0001 | 2.4 ± 0.3 | 1.1 ± 0.1 | 0.004 |
| HOMA-β% | 115.5 ± 11.3 | 70.1 ± 7.9 | 0.01 | 91.6 ± 9.2 | 72.5 ± 9.8 | NS |
| TC (mmol/L) | 4.7 ± 0.2 | 4.3 ± 0.2 | NS | 4.4 ± 0.1 | 4.4 ± 0.3 | NS |
| HDL-C (mmol/L) | 1.3 ± 0.1 | 1.5 ± 0.1 | NS | 1.3 ± 0.1 | 1.5 ± 0.2 | NS |
| LDL-C (mmol/L) | 2.9 ± 0.1 | 2.5 ± 0.1 | NS | 2.7 ± 0.1 | 2.5 ± 0.2 | NS |
| TG (mmol/L) | 1.1 ± 0.1 | 0.7 ± 0.1 | 0.03 | 0.9 ± 0.1 | 0.7 ± 0.1 | NS |
| TC/HDL-C | 3.7 ± 0.2 | 3.0 ± 0.2 | NS | 3.6 ± 0.4 | 3.0 ± 0.2 | NS |
| LDL-C/HDL-C | 2.3 ± 0.1 | 1.8 ± 0.1 | NS | 2.3 ± 0.3 | 1.8 ± 0.2 | NS |
| Atherogenic Index | 2.7 ± 0.2 | 2.0 ± 0.2 | 0.04 | 2.6 ± 0.4 | 2.0 ± 0.2 | NS |
| Systolic BP (mmHg) | 119.3 ± 2.1 | 110.0 ± 4.0 | 0.07 | 116.3 ± 3.2 | 106.3 ± 3.9 | NS |
| Diastolic BP (mmHg) | 74.9 ± 1.5 | 68.4 ± 3.5 | 0.08 | 70.5 ± 2.2 | 64.9 ± 2.9 | NS |
| Heart rate (beats per minute) | 71.6 ± 1.7 | 65.4 ± 2.6 | NS | 69.4 ± 3.0 | 66.6 ± 2.9 | NS |
| Weight (kg) | 84.2 ± 4.3 | 64.3 ± 4.9 | 0.005 | 66.4 ± 4.2 | 57.7 ± 2.8 | NS |
| BMI (kg·m2) | 28.7 ± 1.2 | 22.5 ± 1.2 | 0.001 | 22.8 ± 0.9 | 20.9 ± 0.7 | NS |
| Waist (cm) | 95.2 ± 3.1 | 78.8 ± 4.4 | 0.002 | 81.6 ± 4.3 | 73.2 ± 2.9 | NS |
| Whole body fat% | 30.1 ± 1.9 | 24.7 ± 3.1 | NS | 19.4 ± 2.2 | 22.3 ± 2.8 | NS |
| Whole body fat mass (kg) | 26.0 ± 2.3 | 16.3 ± 3 | 0.007 | 12.9 ± 1.7 | 12.9 ± 1.8 | NS |
| Fat-free mass (kg) | 57.9 ± 2.9 | 47.6 ± 3.3 | NS | 53.1 ± 3.2 | 44.6 ± 2.7 | NS |
| Trunk fat% | 30.4 ± 1.8 | 23.3 ± 3.1 | 0.05 | 19.4 ± 1.9 | 20.3 ± 2.6 | NS |
| Trunk fat mass (kg) | 14.4 ± 1.3 | 8.7 ± 1.8 | 0.02 | 7.2 ± 1.1 | 6.5 ± 1.0 | NS |
| Trunk fat-free mass (kg) | 31.3 ± 1.4 | 26.2 ± 1.5 | NS | 28.9 ± 1.7 | 24.6 ± 1.1 | NS |
| Predicted muscle mass (kg) | 29.7 ± 1.2 | 25.1 ± 1.5 | NS | 27.8 ± 1.7 | 23.6 ± 1.1 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhao, A.; Sandhu, A.K.; Edirisinghe, I.; Burton-Freeman, B.M. Functional Deficits in Gut Microbiome of Young and Middle-Aged Adults with Prediabetes Apparent in Metabolizing Bioactive (Poly)phenols. Nutrients 2020, 12, 3595. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113595
Zhang X, Zhao A, Sandhu AK, Edirisinghe I, Burton-Freeman BM. Functional Deficits in Gut Microbiome of Young and Middle-Aged Adults with Prediabetes Apparent in Metabolizing Bioactive (Poly)phenols. Nutrients. 2020; 12(11):3595. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113595
Chicago/Turabian StyleZhang, Xuhuiqun, Anqi Zhao, Amandeep K. Sandhu, Indika Edirisinghe, and Britt M. Burton-Freeman. 2020. "Functional Deficits in Gut Microbiome of Young and Middle-Aged Adults with Prediabetes Apparent in Metabolizing Bioactive (Poly)phenols" Nutrients 12, no. 11: 3595. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113595