Effects of Daily Raspberry Consumption on Immune-Metabolic Health in Subjects at Risk of Metabolic Syndrome: A Randomized Controlled Trial


Abstract
:1. Introduction
2. Materials and Methods
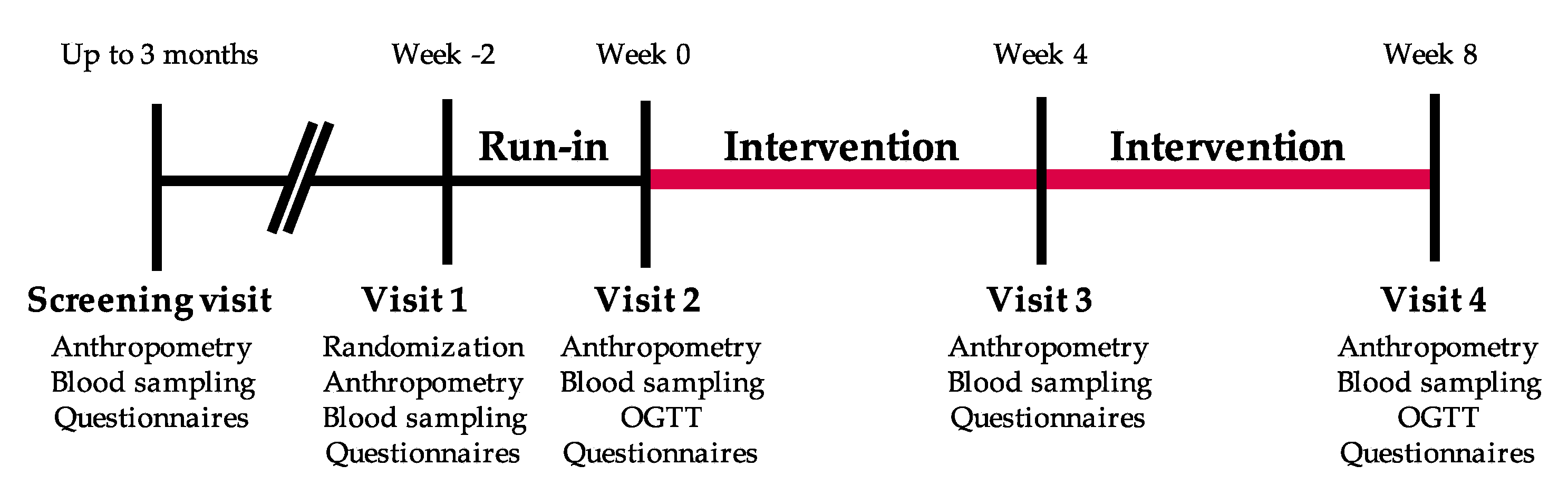
2.1. Study Participants and Eligibility Criteria
2.2. Nutritional Intervention
2.3. Anthropometric Measurements
2.4. Nutritional and Physical Activity Assessments
2.5. Biochemical Analyses
2.6. Transcriptomic Analyses
2.7. Metabolomic Analyses
2.8. Statistical Analyses
3. Results
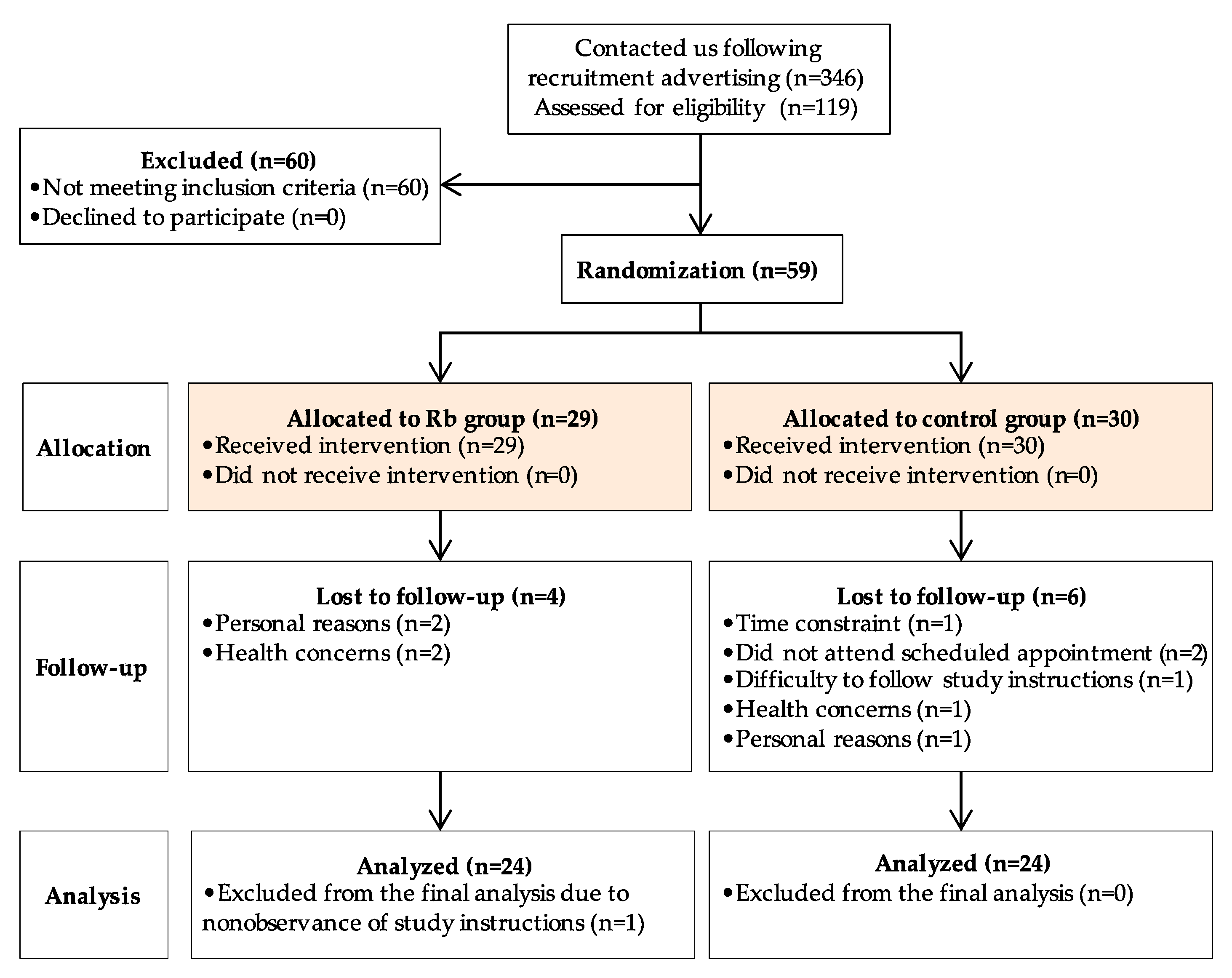
3.1. Trial Flow, Baseline Characteristics, and Compliance
3.2. Food Intake and Physical Activity
3.3. Primary Outcomes
3.4. Secondary Outcomes
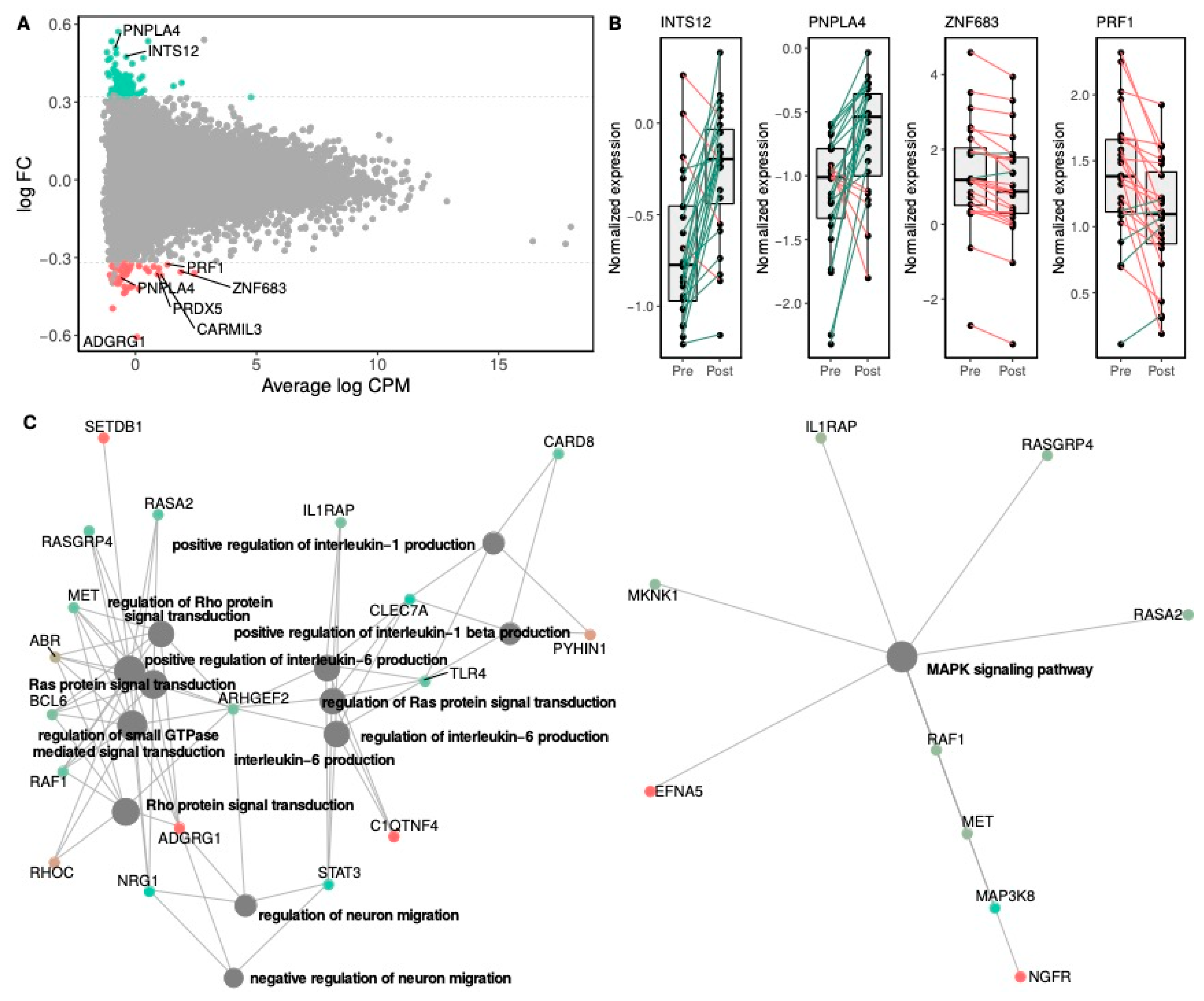
3.4.1. Effects of Rb Supplementation on Gene Expression
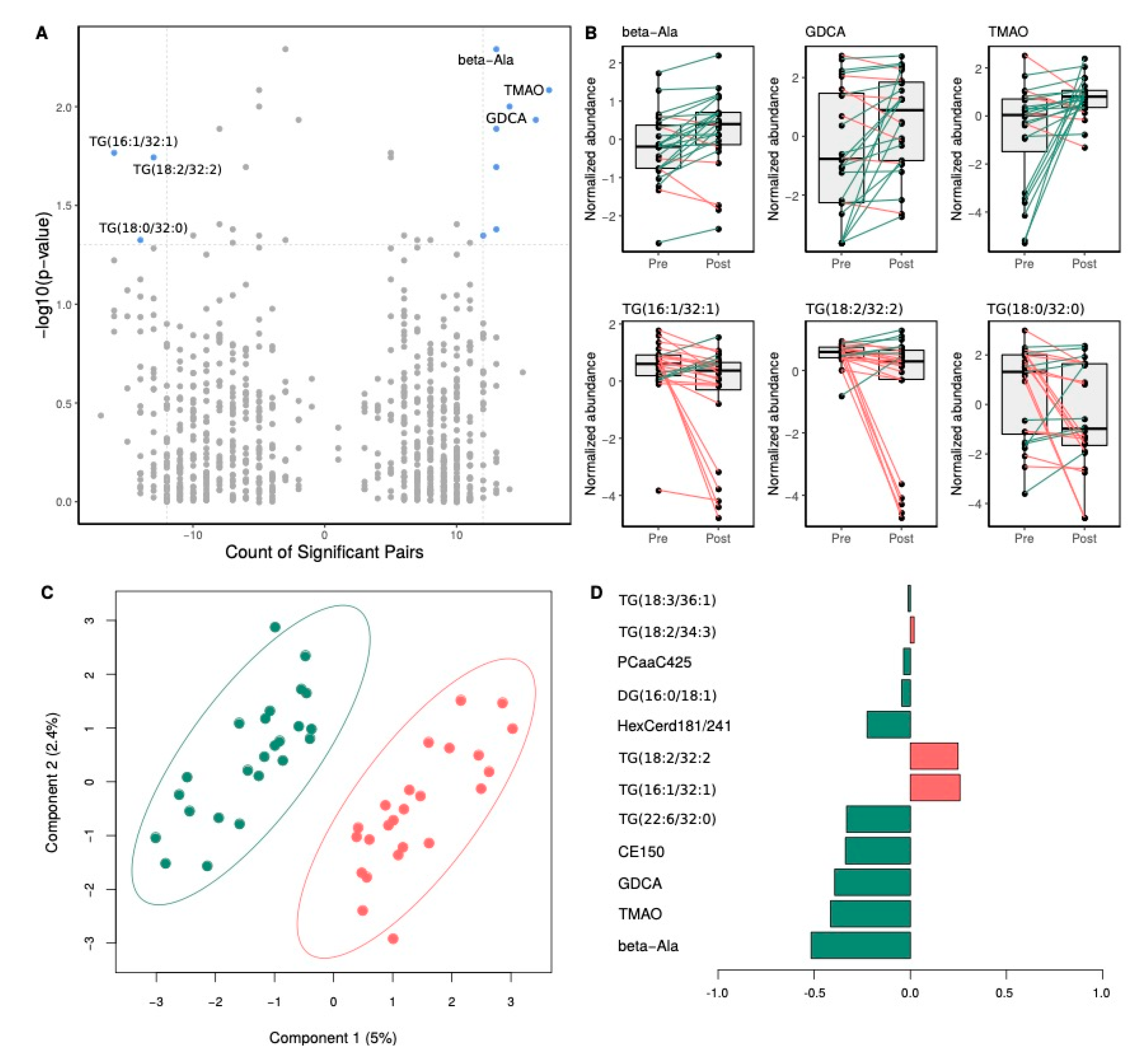
3.4.2. Effects of Rb Supplementation on Metabolite Levels
4. Discussion
Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eichelmann, F.; Schwingshackl, L.; Fedirko, V.; Aleksandrova, K. Effect of plant-based diets on obesity-related inflammatory profiles: A systematic review and meta-analysis of intervention trials. Obes. Rev. 2016, 17, 1067–1079. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Morenga, L.T. Carbohydrate quality and human health: A series of systematic reviews and meta-analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, S.J. The association between dietary fibre deficiency and high-income lifestyle-associated diseases: Burkitt’s hypothesis revisited. Lancet Gastroenterol. Hepatol. 2019, 4, 984–996. [Google Scholar] [CrossRef]
- Alberti, K.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.-C.; James, W.P.T.; Loria, C.M.; Smith, S.C. Harmonizing the Metabolic Syndrome. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [Green Version]
- Health Canada. Canada’s Dietary Guidelines. 2019. Available online: https://food-guide.canada.ca/en/guidelines (accessed on 31 October 2020).
- Shepherd, R. Resistance to Changes in Diet. Proceedings of the Nutrition Society; CABI Publishing: Wallingford, UK, 2002; Volume 61, pp. 267–272. [Google Scholar]
- Luís, Â.; Domingues, F.; Pereira, L. Association between berries intake and cardiovascular diseases risk factors: A systematic review with meta-analysis and trial sequential analysis of randomized controlled trials. Food Funct. 2018, 9, 740–757. [Google Scholar] [CrossRef]
- Rao, A.V.; Snyder, D.M. Raspberries and Human Health: A Review†. J. Agric. Food Chem. 2010, 58, 3871–3883. [Google Scholar] [CrossRef]
- Seeram, N.P.; Adams, L.S.; Zhang, Y.; Lee, R.; Sand, D.; Scheuller, H.S.; Heber, D. Blackberry, Black Raspberry, Blueberry, Cranberry, Red Raspberry, and Strawberry Extracts Inhibit Growth and Stimulate Apoptosis of Human Cancer Cells In Vitro. J. Agric. Food Chem. 2006, 54, 9329–9339. [Google Scholar] [CrossRef]
- Wang, P.-W.; Cheng, Y.-C.; Hung, Y.-C.; Lee, C.-H.; Fang, J.-Y.; Li, W.-T.; Wu, Y.-R.; Pan, T.-L. Red Raspberry Extract Protects the Skin against UVB-Induced Damage with Antioxidative and Anti-inflammatory Properties. Oxidative Med. Cell. Longev. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Tu, L.; Sun, H.; Tang, M.; Zhao, J.; Zhang, Z.; Sun, X.; He, S. Red raspberry extract (Rubus idaeus L shrub) intake ameliorates hyperlipidemia in HFD-induced mice through PPAR signaling pathway. Food Chem. Toxicol. 2019, 133, 110796. [Google Scholar] [CrossRef]
- Noratto, G.; Chew, B.P.; Atienza, L.M. Red raspberry (Rubus idaeus L.) intake decreases oxidative stress in obese diabetic (db/db) mice. Food Chem. 2017, 227, 305–314. [Google Scholar] [CrossRef]
- Kirakosyan, A.; Seymour, E.M.; Kondoleon, N.; Gutierrez-Albanchez, E.; Wolforth, J.; Bolling, S. The intake of red raspberry fruit is inversely related to cardiac risk factors associated with metabolic syndrome. J. Funct. Foods 2018, 41, 83–89. [Google Scholar] [CrossRef]
- Allen, C.T.; Peden-Adams, M.M.; Eudaly, J.; Keil, D.E. Subchronic Exposure to Ellagic Acid Impairs Cytotoxic T-Cell Function and Suppresses Humoral Immunity in Mice. Immunopharmacol. Immunotoxicol. 2003, 25, 409–422. [Google Scholar] [CrossRef]
- Nowak, A.; Sójka, M.; Klewicka, E.; Lipińska, L.; Klewicki, R.; Kołodziejczyk, K. Ellagitannins from Rubus idaeus L. Exert Geno- and Cytotoxic Effects against Human Colon Adenocarcinoma Cell Line Caco-2. J. Agric. Food Chem. 2017, 65, 2947–2955. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Kang, S.; Wang, Y.; Liu, K.; Oshima, K.; Huang, Y.-W.; Zhang, J.; Yearsley, M.; Yu, J.; Wang, L.-S. Black Raspberries Enhance Natural Killer Cell Infiltration into the Colon and Suppress the Progression of Colorectal Cancer. Front. Immunol. 2017, 8, 997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knobloch, T.; Ryan, N.M.; Bruschweiler-Li, L.; Wang, C.; Bernier, M.C.; Somogyi, Á.; Yan, P.; Cooperstone, J.L.; Mo, X.; Brüschweiler, R.; et al. Metabolic Regulation of Glycolysis and AMP Activated Protein Kinase Pathways during Black Raspberry-Mediated Oral Cancer Chemoprevention. Metabolites 2019, 9, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, H.S.; Kim, S.; Hong, S.J.; Choi, S.C.; Choi, J.-H.; Kim, J.-H.; Park, C.-Y.; Cho, J.Y.; Lee, T.-B.; Kwon, J.-W.; et al. Black Raspberry Extract Increased Circulating Endothelial Progenitor Cells and Improved Arterial Stiffness in Patients with Metabolic Syndrome: A Randomized Controlled Trial. J. Med. Food 2016, 19, 346–352. [Google Scholar] [CrossRef]
- Sardo, C.; Kitzmiller, J.P.; Apseloff, G.; Harris, R.B.; Roe, D.J.; Stoner, G.D.; Jacobs, E.T. An Open-Label Randomized Crossover Trial of Lyophilized Black Raspberries on Postprandial Inflammation in Older Overweight Males. Am. J. Ther. 2016, 23, e86–e91. [Google Scholar] [CrossRef]
- Xiao, D.; Zhu, L.; Edirisinghe, I.; Fareed, J.; Brailovsky, Y.; Burton-Freeman, B.M. Attenuation of Postmeal Metabolic Indices with Red Raspberries in Individuals at Risk for Diabetes: A Randomized Controlled Trial. Obesity 2019, 27, 542–550. [Google Scholar] [CrossRef]
- Schell, J.; Betts, N.M.; Lyons, T.J.; Basu, A. Raspberries Improve Postprandial Glucose and Acute and Chronic Inflammation in Adults with Type 2 Diabetes. Ann. Nutr. Metab. 2019, 74, 165–174. [Google Scholar] [CrossRef]
- Istas, G.; Feliciano, R.P.; Weber, T.; Garcia-Villalba, R.; Tomás-Barberán, F.A.; Heiss, C.; Rodriguez-Mateos, A. Plasma urolithin metabolites correlate with improvements in endothelial function after red raspberry consumption: A double-blind randomized controlled trial. Arch. Biochem. Biophys. 2018, 651, 43–51. [Google Scholar] [CrossRef]
- Clegg, M.E.; Pratt, M.; Meade, C.M.; Henry, C.J.K. The addition of raspberries and blueberries to a starch-based food does not alter the glycaemic response. Br. J. Nutr. 2011, 106, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Puupponen-Pimiä, R.; Seppänen-Laakso, T.; Kankainen, M.; Maukonen, J.; Törrönen, R.; Kolehmainen, M.; Leppänen, T.; Moilanen, E.; Nohynek, L.; Aura, A.-M.; et al. Effects of ellagitannin-rich berries on blood lipids, gut microbiota, and urolithin production in human subjects with symptoms of metabolic syndrome. Mol. Nutr. Food Res. 2013, 57, 2258–2263. [Google Scholar] [CrossRef] [PubMed]
- Törrönen, R.; Kolehmainen, M.; Sarkkinen, E.; Poutanen, K.; Mykkänen, H.; Niskanen, L. Berries Reduce Postprandial Insulin Responses to Wheat and Rye Breads in Healthy Women. J. Nutr. 2013, 143, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Scarsella, C.; Alméras, N.; Mauriège, P.; Blanchet, C.; Sauvé, L.; Dewailly, E.; Bergeron, J.; Després, J.-P. Prevalence of metabolic alterations predictive of cardiovascular disease risk in the Québec population. Can. J. Cardiol. 2003, 19, 51–57. [Google Scholar] [PubMed]
- Norgan, N.G. A Review of: Anthropometric Standardization Reference Manual Edited by T. G. LOHMAN, A.F. ROCHE and R. MARTORELL. (Champaign, IL.: Human Kinetics Books, 1988.) ISBN 087322 121 4. Ergonomics 1988, 31, 1493–1494. [Google Scholar] [CrossRef]
- Labonté, M.-È.; Cyr, A.; Baril-Gravel, L.; Royer, M.-M.; Lamarche, B. Validity and reproducibility of a web-based, self-administered food frequency questionnaire. Eur. J. Clin. Nutr. 2011, 66, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Taylor, H.L.; Jacobs, D.R.; Schucker, B.; Knudsen, J.; Leon, A.S.; Debacker, G. A questionnaire for the assessment of leisure time physical activities. J. Chronic Dis. 1978, 31, 741–755. [Google Scholar] [CrossRef]
- Burstein, M.; Samaille, J. Sur un dosage rapide du cholesterol lié aux α-et aux β-lipoprotéines du sérum. Clin. Chim. Acta 1960, 5, 609. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the Concentration of Low-Density Lipoprotein Cholesterol in Plasma, Without Use of the Preparative Ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- LeDue, T.B.; Weiner, D.L.; Sipe, J.D.; Poulin, S.E.; Collins, M.F.; Rifai, N. Analytical evaluation of particle-enhanced immunonephelometric assays for C-reactive protein, serum amyloid A and mannose-binding protein in human serum. Ann. Clin. Biochem. Int. J. Lab. Med. 1998, 35, 745–753. [Google Scholar] [CrossRef]
- Richterich, R.; Dauwalder, H. [Determination of plasma glucose by hexokinase-glucose-6-phosphate dehydrogenase method]. Schweiz. Med. Wochenschr. 1971, 101, 615–618. [Google Scholar] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, M.; DeFronzo, R.A. Insulin sensitivity indices obtained from oral glucose tolerance testing: Comparison with the euglycemic insulin clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Reynés, B.; Priego, T.; Cifre, M.; Oliver, P.; Palou, A. Peripheral Blood Cells, a Transcriptomic Tool in Nutrigenomic and Obesity Studies: Current State of the Art. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1006–1020. [Google Scholar] [CrossRef] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Xia, J. MetaboAnalystR: An R package for flexible and reproducible analysis of metabolomics data. Bioinformatics 2018, 34, 4313–4314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.D.; Giskeødegård, G.F.; Bathen, T.F.; Sitter, B.; Bofin, A.M.; Lønning, P.E.; Lundgren, S.; Gribbestad, I.S. Prognostic value of metabolic response in breast cancer patients receiving neoadjuvant chemotherapy. BMC Cancer 2012, 12, 39. [Google Scholar] [CrossRef] [Green Version]
- Westerhuis, J.A.; Van Velzen, E.J.J.; Hoefsloot, H.C.J.; Smilde, A.K. Multivariate paired data analysis: Multilevel PLSDA versus OPLSDA. Metabolomics 2010, 6, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Le Cao, K.-A.; Boitard, S.; Besse, P. Sparse PLS discriminant analysis: Biologically relevant feature selection and graphical displays for multiclass problems. BMC Bioinform. 2011, 12, 253. [Google Scholar] [CrossRef] [Green Version]
- Rohart, F.; Gautier, B.; Singh, A.; Le Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Post, M.; Cuapio, A.; Osl, M.; Lehmann, D.; Resch, U.; Davies, D.M.; Bilban, M.; Schlechta, B.; Eppel, W.; Nathwani, A.; et al. The Transcription Factor ZNF683/HOBIT Regulates Human NK-Cell Development. Front. Immunol. 2017, 8, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, G.-W.; Hsiao, C.-C.; Peng, Y.-M.; Braga, F.A.V.; Kragten, N.A.; Remmerswaal, E.B.; Van De Garde, M.D.; Straussberg, R.; König, G.M.; Kostenis, E.; et al. The Adhesion G Protein-Coupled Receptor GPR56/ADGRG1 Is an Inhibitory Receptor on Human NK Cells. Cell Rep. 2016, 15, 1757–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debien, E.; Mayol, K.; Biajoux, V.; Daussy, C.; De Agüero, M.G.; Taillardet, M.; Dagany, N.; Brinza, L.; Henry, T.; Dubois, B.; et al. S1PR5 is pivotal for the homeostasis of patrolling monocytes. Eur. J. Immunol. 2013, 43, 1667–1675. [Google Scholar] [CrossRef] [Green Version]
- Drouillard, A.; Mathieu, A.-L.; Marçais, A.; Belot, A.; Viel, S.; Mingueneau, M.; Guckian, K.; Walzer, T. S1PR5 is essential for human natural killer cell migration toward sphingosine-1 phosphate. J. Allergy Clin. Immunol. 2018, 141, 2265–2268.e1. [Google Scholar] [CrossRef] [Green Version]
- Maceyka, M.; Milstien, S.; Spiegel, S. Sphingosine kinases, sphingosine-1-phosphate and sphingolipidomics. Prostaglandins Other Lipid Mediat. 2005, 77, 15–22. [Google Scholar] [CrossRef]
- Iqbal, J.; Walsh, M.T.; Hammad, S.M.; Cuchel, M.; Tarugi, P.; Hegele, R.A.; Davidson, N.O.; Rader, D.J.; Klein, R.L.; Hussain, M.M. Microsomal Triglyceride Transfer Protein Transfers and Determines Plasma Concentrations of Ceramide and Sphingomyelin but Not Glycosylceramide. J. Biol. Chem. 2015, 290, 25863–25875. [Google Scholar] [CrossRef] [Green Version]
- Studer, E.; Zhou, X.; Zhao, R.; Wang, Y.; Takabe, K.; Nagahashi, M.; Pandak, W.M.; Dent, P.; Spiegel, S.; Shi, R.; et al. Conjugated bile acids activate the sphingosine-1-phosphate receptor 2 in primary rodent hepatocytes. Hepatology 2011, 55, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Bryan, A.M.; Del Poeta, M. Sphingosine-1-phosphate receptors and innate immunity. Cell. Microbiol. 2018, 20, e12836. [Google Scholar] [CrossRef] [Green Version]
- Pan, P.; Zhu, Z.; Oshima, K.; Aldakkak, M.; Tsai, S.; Huang, Y.-W.; Dong, W.; Zhang, J.; Lin, C.-W.; Wang, Y.; et al. Black raspberries suppress pancreatic cancer through modulation of NKp46 + CD8 +, and CD11b + immune cells. Food Front. 2020, 1, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Bessler, H. On the Link between Ellagic Acid and the Immune Balance between Human Mononuclear and Colon Carcinoma Cells. Immunol Curr Res 2017, 1, 1. [Google Scholar]
- Wu, F.-S.; Gibbs, T.T.; Farb, D.H. Dual activation of GABAA and glycine receptors by β-alanine: Inverse modulation by progesterone and 5α-pregnan-3α-ol-20-one. Eur. J. Pharmacol. Mol. Pharmacol. 1993, 246, 239–246. [Google Scholar] [CrossRef]
- Jin, Z.; Mendu, S.K.; Birnir, B. GABA is an effective immunomodulatory molecule. Amino Acids 2013, 45, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duthey, B.; Hübner, A.; Diehl, S.; Boehncke, S.; Pfeffer, J.; Boehncke, W.-H. Anti-inflammatory effects of the GABAB receptor agonist baclofen in allergic contact dermatitis. Exp. Dermatol. 2010, 19, 661–666. [Google Scholar] [CrossRef]
- Geng, J.; Yang, C.; Wang, B.; Zhang, X.; Hu, T.; Gu, Y.; Li, J. Trimethylamine N-oxide promotes atherosclerosis via CD36-dependent MAPK/JNK pathway. Biomed. Pharmacother. 2018, 97, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, G.; Maisto, M.; Schisano, C.; Ciampaglia, R.; Narciso, V.; Tenore, G.C.; Novellino, E. Effects of Grape Pomace Polyphenolic Extract (Taurisolo®) in Reducing TMAO Serum Levels in Humans: Preliminary Results from a Randomized, Placebo-Controlled, Cross-Over Study. Nutrients 2019, 11, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bresciani, L.; Dall’Asta, M.; Favari, C.; Calani, L.; Del Rio, D.; Brighenti, F. An in vitro exploratory study of dietary strategies based on polyphenol-rich beverages, fruit juices and oils to control trimethylamine production in the colon. Food Funct. 2018, 9, 6470–6483. [Google Scholar] [CrossRef]
- Mandal, N.; Grambergs, R.; Mondal, K.; Basu, S.K.; Tahia, F.; Dagogo-Jack, S. Role of ceramides in the pathogenesis of diabetes mellitus and its complications. J. Diabetes Complicat. 2020, 10, 107734. [Google Scholar] [CrossRef]
- Cas, M.D.; Ghidoni, R. Cancer Prevention and Therapy with Polyphenols: Sphingolipid-Mediated Mechanisms. Nutrients 2018, 10, 940. [Google Scholar] [CrossRef] [Green Version]
- Si, X.; Tian, J.; Shu, C.; Wang, Y.; Gong, E.S.; Zhang, Y.; Zhang, W.; Cui, H.; Li, B. Serum Ceramide Reduction by Blueberry Anthocyanin-Rich Extract Alleviates Insulin Resistance in Hyperlipidemia Mice. J. Agric. Food Chem. 2020, 68, 8185–8194. [Google Scholar] [CrossRef]
- Gupta, P.; Mohammad, T.; Khan, P.; Alajmi, M.F.; Hussain, A.; Rehman, T.; Hassan, I. Evaluation of ellagic acid as an inhibitor of sphingosine kinase 1: A targeted approach towards anticancer therapy. Biomed. Pharmacother. 2019, 118, 109245. [Google Scholar] [CrossRef] [PubMed]
- Khattab, R.; Celli, G.B.; Ghanem, A.; Brooks, M.S.-L. Effect of frozen storage on polyphenol content and antioxidant activity of haskap berries (Lonicera caerulea L.). J. Berry Res. 2015, 5, 231–242. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | N | Control | N | Rb | p-value | ||||
|---|---|---|---|---|---|---|---|---|---|
| Sex (men/women) | 24 | 9/15 | 24 | 7/17 | 0.54 | ||||
| Age (years) | 24 | 31.92 | ± | 8.05 | 24 | 32.46 | ± | 10.12 | 0.83 |
| BMI (kg/m2) | 24 | 29.38 | ± | 3.94 | 24 | 30.42 | ± | 5.00 | 0.43 |
| Waist circumference (cm) | 24 | 98.10 | ± | 11.81 | 24 | 98.53 | ± | 13.32 | 0.90 |
| Fasting glucose (mmol/L) | 24 | 4.84 | ± | 0.47 | 23 | 4.98 | ± | 0.59 | 0.38 |
| Fasting insulin (pmol/L) | 22 | 73.91 | ± | 33.57 | 19 | 83.11 | ± | 43.54 | 0.21 |
| HbA1C (%) | 24 | 5.05 | ± | 0.29 | 24 | 5.03 | ± | 0.31 | 0.85 |
| Plasma TG (mmol/L) | 24 | 1.56 | ± | 0.78 | 24 | 1.46 | ± | 0.80 | 0.66 |
| ApoB-100 (g/L) | 24 | 0.88 | ± | 0.17 | 24 | 0.92 | ± | 0.24 | 0.48 |
| HDL-C (mmol/L) | 24 | 1.33 | ± | 0.23 | 24 | 1.32 | ± | 0.40 | 0.94 |
| LDL-C (mmol/L) | 24 | 2.55 | ± | 0.74 | 24 | 2.65 | ± | 0.86 | 0.69 |
| CRP (mg/L) | 24 | 2.72 | ± | 2.82 | 23 | 2.68 | ± | 2.34 | 0.95 |
| HOMA-IR | 13 | 1.78 | ± | 0.76 | 11 | 2.84 | ± | 1.73 | 0.06 |
| MATSUDA | 13 | 5.79 | ± | 2.43 | 11 | 4.54 | ± | 2.24 | 0.21 |
| Time | Week 0 | Week 4 | Week 8 | p-Values | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | Control | Rb | Control | Rb | Control | Rb | Time | tx | tx*Time | ||||||||||||
| Energy (kcal) | 2132.8 | ± | 823.2 | 2059.7 | ± | 662.1 | 1992.7 | ± | 692.4 | 1983.3 | ± | 682.1 | 2060.7 | ± | 638.7 | 2075.8 | ± | 623.2 | 0.32 | 0.70 | 0.83 |
| Lipids (g) | 93.5 | ± | 44.9 | 82.7 | ± | 28.6 | 87.7 | ± | 40.9 | 76.4 | ± | 31.8 | 91.4 | ± | 37.7 | 80.9 | ± | 28.7 | 0.25 | 0.60 | 0.99 |
| Proteins (g) | 90.1 | ± | 37.3 | 87.2 | ± | 30.9 | 87.4 | ± | 32.1 | 84.0 | ± | 30.1 | 91.9 | ± | 28.6 | 88.3 | ± | 30.8 | 0.53 | 0.68 | 1.00 |
| Soluble Fiber (g) | 8.3 | ± | 4.1 | 7.7 a | ± | 3.3 | 7.3 x | ± | 3.4 | 8.5 y | ± | 3.1 | 7.5 x | ± | 3.1 | 8.9 by | ± | 3.1 | 0.69 | 0.01 | 0.02 |
| Insoluble Fiber (g) | 15.6 | ± | 7.8 | 15.0 x | ± | 6.8 | 13.8 a | ± | 6.9 | 23.1 by | ± | 6.6 | 14.4 x | ± | 6.2 | 23.8 by | ± | 6.3 | <0.0001 | <0.0001 | <0.0001 |
| Fructose (g) | 24.1 a | ± | 14.4 | 24.9 a | ± | 9.9 | 19.3 bx | ± | 8.9 | 29.7 by | ± | 9.4 | 22.2 x | ± | 9.0 | 29.9 by | ± | 8.9 | 0.41 | <0.0001 | 0.002 |
| Glucose (g) | 23.7 a | ± | 13.3 | 25.1 a | ± | 8.9 | 19.8 bx | ± | 8.8 | 30.2 by | ± | 7.9 | 22.2 x | ± | 8.8 | 30.9 by | ± | 7.8 | 0.18 | <0.0001 | 0.0005 |
| Alcohol (g) | 3.2 | ± | 3.6 | 2.8 | ± | 2.8 | 2.5 | ± | 2.1 | 1.9 | ± | 1.7 | 2.8 | ± | 2.5 | 2.3 | ± | 1.4 | 0.07 | 0.37 | 0.95 |
| Caffeine (mg) | 147.7 | ± | 148.9 | 103.6 | ± | 114.9 | 147.9 | ± | 142.7 | 95.2 | ± | 126.9 | 150.0 | ± | 149.7 | 106.6 | ± | 142.7 | 0.59 | 0.65 | 0.73 |
| Bread and cereals (serving) | 3.9 | ± | 2.2 | 4.6 | ± | 2.3 | 4.0 | ± | 2.0 | 4.1 | ± | 2.6 | 4.1 | ± | 2.3 | 4.1 | ± | 1.9 | 0.45 | 0.36 | 0.26 |
| Fruits (serving) | 2.9 a | ± | 2.6 | 3.1 a | ± | 1.6 | 2.2 bx | ± | 1.7 | 6.0 by | ± | 1.3 | 2.9 x | ± | 1.9 | 5.9 by | ± | 1.3 | <0.0001 | <0.0001 | <0.0001 |
| Vegetables (serving) | 3.6 | ± | 1.9 | 3.6 | ± | 2.3 | 3.3 | ± | 1.6 | 2.6 | ± | 1.6 | 3.4 | ± | 1.6 | 3.2 | ± | 2.1 | 0.02 | 0.10 | 0.41 |
| Dairy products (serving) | 2.0 | ± | 1.7 | 2.2 | ± | 1.1 | 2.0 | ± | 1.3 | 2.3 | ± | 1.3 | 1.8 | ± | 0.9 | 2.2 | ± | 1.4 | 0.69 | 0.51 | 0.74 |
| Animal proteins (serving) | 2.3 | ± | 1.2 | 1.8 | ± | 0.9 | 2.2 | ± | 1.2 | 1.8 | ± | 0.8 | 2.5 | ± | 1.2 | 2.0 | ± | 1.1 | 0.13 | 0.39 | 0.93 |
| Physical activity (AMI) | 298.1 | ± | 147.0 | 259.0 | ± | 131.9 | 280.8 | ± | 190.5 | 218.3 | ± | 159.9 | 0.13 | 0.34 | 0.54 | ||||||
| Variable | N | Control | N | Rb | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| BMI (kg/m2) | 24 | −0.01 | ± | 0.60 | 24 | +0.10 | ± | 0.57 | 0.43 |
| Waist circumference (cm) | 24 | −0.18 | ± | 2.18 | 24 | +0.39 | ± | 3.12 | 0.46 |
| Fasting glucose (mmol/L) | 23 | +0.01 | ± | 0.35 | 22 | −0.09 | ± | 0.29 | 0.27 |
| Fasting insulin (pmol/L) | 19 | +3.10 | ± | 22.98 | 16 | +3.12 | ± | 26.49 | 0.99 |
| HbA1C (%) | 23 | −0.03 | ± | 0.13 | 23 | +0.03 | ± | 0.14 | 0.13 |
| Plasma TG (mmol/L) | 22 | −0.04 | ± | 0.53 | 23 | −0.17 | ± | 0.68 | 0.48 |
| ApoB-100 (g/L) | 23 | +0.01 | ± | 0.09 | 23 | −0.03 | ± | 0.12 | 0.30 |
| HDL-C (mmol/L) | 23 | −0.04 | ± | 0.10 | 23 | −0.01 | ± | 0.18 | 0.43 |
| LDL-C (mmol/L) | 22 | +0.01 | ± | 0.42 | 23 | +0.03 | ± | 0.42 | 0.89 |
| CRP (mg/L) | 20 | +0.90 | ± | 2.33 | 22 | −0.04 | ± | 1.59 | 0.14 |
| HOMA-IR | 13 | +0.11 | ± | 0.74 | 11 | −0.16 | ± | 0.53 | 0.33 |
| Matsuda index | 13 | −0.60 | ± | 2.08 | 11 | +0.14 | ± | 1.43 | 0.33 |
| Time | Week 0 | Week 4 | Week 8 | p-Values | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | N Control | N Rb | Control | Rb | Control | Rb | Time | tx | tx*Time | ||||||||||||||
| SBP (mmHg) | 24 | 110.8 | ± | 11.2 | 24 | 112.8 | ± | 11.0 | 110.7 | ± | 12.9 | 113.1 | ± | 10.4 | 112.8 | ± | 11.9 | 110.9 | ± | 11.0 | 0.99 | 0.37 | 0.03 |
| DBP (mmHg) | 24 | 68.9 | ± | 8.7 | 24 | 71.9 | ± | 8.9 | 68.8 | ± | 10.4 | 71.8 | ± | 9.0 | 69.6 | ± | 11.1 | 70.3 | ± | 9.3 | 0.86 | 0.67 | 0.34 |
| BMI (kg/m2) | 24 | 29.4 | ± | 3.9 | 24 | 30.4 | ± | 4.9 | 29.5 | ± | 3.9 | 30.5 | ± | 5.0 | 29.4 | ± | 3.9 | 30.5 | ± | 5.1 | 0.44 | 0.69 | 0.69 |
| Waist circumference (cm) | 24 | 98.1 | ± | 11.8 | 24 | 98.5 | ± | 13.3 | 98.5 | ± | 12.1 | 98.4 | ± | 13.3 | 97.9 | ± | 12.9 | 98.9 | ± | 14.1 | 0.93 | 0.73 | 0.23 |
| Hips circumference (cm) | 24 | 108.9 | ± | 7.5 | 24 | 112.1 | ± | 10.2 | 109.5 | ± | 8.4 | 112.3 | ± | 10.5 | 109.3 | ± | 8.4 | 112.1 | ± | 10.5 | 0.63 | 0.59 | 0.77 |
| CRP | 20 | 2.50 | ± | 2.50 | 21 | 2.1 | ± | 1.64 | 2.65 | ± | 2.60 | 2.86 | ± | 2.59 | 2.46 | ± | 2.42 | 3.28 | ± | 2.67 | 0.30 | 0.16 | 0.26 |
| ApoB-100 | 23 | 0.86 a | ± | 0.16 | 23 | 0.92 | ± | 0.24 | 0.95 b | ± | 0.17 | 0.92 y | ± | 0.21 | 0.87 a | ± | 0.19 | 0.89 | ± | 0.25 | 0.002 | 0.02 | 0.03 |
| Total-C (mmol/L) | 23 | 4.53 a | ± | 0.79 | 23 | 4.60 | ± | 0.91 | 4.85 b | ± | 0.81 | 4.70 | ± | 0.84 | 4.51 a | ± | 0.88 | 4.54 | ± | 0.86 | 0.001 | 0.14 | 0.22 |
| HDL-C (mmol/L) | 23 | 1.33 | ± | 0.23 | 23 | 1.30 | ± | 0.40 | 1.39 a | ± | 0.29 | 1.32 | ± | 0.35 | 1.29 b | ± | 0.27 | 1.30 | ± | 0.33 | 0.03 | 0.49 | 0.17 |
| LDL-C (mmol/L) | 22 | 2.44 a | ± | 0.67 | 23 | 2.62 | ± | 0.87 | 2.78 b | ± | 0.68 | 2.74 | ± | 0.78 | 2.45 a | ± | 0.55 | 2.65 | ± | 0.82 | 0.001 | 0.42 | 0.14 |
| TG (mmol/L) | 22 | 1.42 | ± | 0.61 | 23 | 1.47 | ± | 0.82 | 1.32 | ± | 0.60 | 1.39 | ± | 0.64 | 1.38 | ± | 0.61 | 1.29 | ± | 0.61 | 0.37 | 0.59 | 0.72 |
| Fasting glucose (mmol/L) | 23 | 4.82 | ± | 0.47 | 22 | 5.00 | ± | 0.59 | 4.85 | ± | 0.49 | 5.11 | ± | 0.57 | 4.83 | ± | 0.52 | 4.91 | ± | 0.57 | 0.12 | 0.99 | 0.25 |
| Fasting insulin (pmol/L) | 17 | 65.3 | ± | 29.0 | 16 | 80.1 | ± | 37.4 | 65.9 | ± | 29.1 | 77.5 | ± | 33.2 | 64.0 | ± | 23.2 | 83.3 | ± | 44.2 | 0.77 | 0.59 | 0.10 |
| HbA1C (%) | 23 | 5.02 | ± | 0.27 | 23 | 5.04 | ± | 0.32 | 4.98 | ± | 0.29 | 5.03 | ± | 0.30 | 5.00 | ± | 0.27 | 5.07 | ± | 0.29 | 0.11 | 0.22 | 0.23 |
| RefSeq | Gene Symbol | Gene Name | Nominal p-Value | FDR | FC |
|---|---|---|---|---|---|
| NM_001114759 | ZNF683 | Zinc finger protein 683 | 4.5 × 10−6 | 0.07 | −1.28 |
| NM_001470 | GABBR1 | Gamma-aminobutyric acid type B receptor subunit 1 | 7.4 × 10−6 | 0.07 | 1.22 |
| NM_033423 | GZMH | Granzyme H | 2.5 × 10−5 | 0.12 | −1.19 |
| NM_031950 | FGFBP2 | Fibroblast growth factor binding protein 2 | 2.6 × 10−5 | 0.12 | −1.20 |
| NM_030760 | S1PR5 | Sphingosine-1-phosphate receptor 5 | 4.8 × 10−5 | 0.18 | −1.21 |
| NM_139355 | MATK | Megakaryocyte-associated tyrosine kinase | 8.4 × 10−5 | 0.26 | −1.16 |
| NM_020395 | INTS12 | Integrator complex subunit 12 | 9.5 × 10−5 | 0.26 | 1.39 |
| NM_001144884 | SLC30A7 | Solute carrier family 30 member 7 | 1.7 × 10−4 | 0.28 | 1.23 |
| NM_005170 | ASCL2 | Achaete-scute family bHLH transcription factor 2 | 1.8 × 10−4 | 0.28 | −1.16 |
| NM_025069 | ZNF703 | Zinc finger protein 703 | 1.9 × 10−4 | 0.28 | −1.14 |
| NM_001083116 | PRF1 | Perforin 1 | 2.1 × 10−4 | 0.28 | −1.25 |
| NM_138360 | CARMIL3 | Capping protein regulator and myosin 1 linker 3 | 2.1 × 10−4 | 0.28 | −1.29 |
| NM_005601 | NKG7 | Natural killer cell granule protein 7 (1) | 2.2 × 10−4 | 0.28 | −1.20 |
| NM_001024401 | SBK1 | SH3 domain binding kinase 1 | 2.2 × 10−4 | 0.28 | −1.12 |
| NM_006653 | FRS3 | Fibroblast growth factor receptor substrate 3 | 2.2 × 10−4 | 0.28 | 1.17 |
| NR_110601 | PGS1 | Phosphatidylglycerophosphate synthase 1 | 2.9 × 10−4 | 0.33 | 1.20 |
| NR_110030 | LINC01215 | Long intergenic non-protein coding RNA 1215 | 3.0 × 10−4 | 0.33 | 1.24 |
| NM_001363693 | NKG7 | Natural killer cell granule protein 7 (2) | 3.2 × 10−4 | 0.33 | −1.24 |
| NM_001145770 | ADGRG1 | Adhesion G protein-coupled receptor G1 | 3.5 × 10−4 | 0.34 | −1.52 |
| NR_024618 | LINC02035 | Long intergenic non-protein coding RNA 2035 | 3.6 × 10−4 | 0.34 | 1.14 |
| NM_000234 | LIG1 | DNA ligase 1 | 3.9 × 10−4 | 0.34 | −1.23 |
| NM_001122630 | CDKN1C | Cyclin dependent kinase inhibitor 1C | 3.9 × 10−4 | 0.34 | -1.18 |
| NM_170783 | ZNRD1 | Zinc ribbon domain containing 1 | 4.1 × 10−4 | 0.34 | −1.22 |
| NM_013432 | TONSL | Tonsoku like, DNA repair protein | 4.8 × 10−4 | 0.36 | −1.15 |
| NM_001145777 | FKBP5 | FKBP prolyl isomerase 5 | 4.9 × 10−4 | 0.36 | 1.19 |
| NM_198053 | CD247 | CD247 molecule | 5.1 × 10−4 | 0.36 | −1.10 |
| NM_032737 | LMNB2 | Lamin B2 | 5.2 × 10−4 | 0.36 | −1.09 |
| NM_004650 | PNPLA4 | Patatin like phospholipase domain containing 4 (1) | 6.2 × 10−4 | 0.37 | −1.30 |
| NM_001271822 | SERPINB6 | Serpin family B member 6 | 6.2 × 10−4 | 0.37 | −1.17 |
| NM_001358511 | PRDX5 | Peroxiredoxin 5 | 6.4 × 10−4 | 0.37 | −1.29 |
| NM_005686 | SOX13 | SRY-box transcription factor 13 | 6.4 × 10−4 | 0.37 | −1.14 |
| NM_017931 | TTC38 | Tetratricopeptide repeat domain 38 | 6.4 × 10−4 | 0.37 | −1.12 |
| NM_001142389 | PNPLA4 | Patatin like phospholipase domain containing 4 (2) | 6.8 × 10−4 | 0.37 | 1.43 |
| NM_012483 | GNLY | Granulysin | 6.9 × 10−4 | 0.37 | −1.18 |
| NM_001004310 | FCRL6 | Fc receptor like 6 | 6.9 × 10−4 | 0.37 | −1.20 |
| NM_024310 | PLEKHF1 | Pleckstrin homology and FYVE domain containing 1 | 7.2 × 10−4 | 0.37 | −1.16 |
| NM_013351 | TBX21 | T-box transcription factor 21 | 7.3 × 10−4 | 0.37 | −1.18 |
| NM_007182 | RASSF1 | Ras association domain family member 1 | 7.6 × 10−4 | 0.37 | −1.14 |
| NM_006056 | NMUR1 | Neuromedin U receptor 1 | 7.7 × 10−4 | 0.37 | −1.13 |
| NM_001110556 | FLNA | Filamin A | 8.9 × 10−4 | 0.42 | −1.12 |
| NM_004669 | CLIC3 | Chloride intracellular channel 3 | 9.3 × 10−4 | 0.42 | −1.22 |
| NM_001136044 | TMUB1 | Transmembrane and ubiquitin-like domain containing 1 | 9.3 × 10−4 | 0.42 | −1.11 |
| NM_001335 | CTSW | Cathepsin W | 9.5 × 10−4 | 0.42 | −1.14 |
| Metabolite Name | Common Name | Super Pathway | p-Value | HMDB |
|---|---|---|---|---|
| beta-Ala | β-Alanine | Biogenic Amines | 0.005 | HMDB0000056 |
| TMAO | Trimethylamine N-oxide | Amine Oxides | 0.008 | HMDB0000925 |
| GDCA | Deoxycholic acid glycine conjugate | Bile Acids | 0.01 | HMDB00631 |
| CE15:0 | Cholesterol 1-pentadecanoate | Cholesterol Esters | 0.01 | HMDB0060057 |
| TG (22:6/32:0) | 1-Palmitoyl-2-palmitoyl-3-docosahexaenoyl-glycerol | Triacylglycerols | 0.01 | HMDB10418 |
| TG (16:1/32:1) | 1-Octadecanoyl-2-(9Z-hexadecenoyl)-3-(9Z-tetradecenoyl)-glycerol | Triacylglycerols | 0.02 | HMDB0044888 |
| TG (18:2/32:2) | 1-Palmitoleoyl-2-palmitoleoyl-3-linoleoyl-glycerol | Triacylglycerols | 0.02 | HMDB05435 |
| HexCer(d18:1/24:1) | Hexosylceramide | Glucosylceramides | 0.02 | - |
| PC aa C42:5 | 1-Arachidonyl-2-docosapentaenoyl-sn-glycero-3-phosphocholine | Glycerophospholipids | 0.04 | HMDB0008287 |
| TG (18:0/32:0) | 1-Octadecanoyl-2-octadecanoyl-3-(9Z-tetradecenoyl)-glycerol | Triacylglycerols | 0.04 | HMDB0044753 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franck, M.; de Toro-Martín, J.; Garneau, V.; Guay, V.; Kearney, M.; Pilon, G.; Roy, D.; Couture, P.; Couillard, C.; Marette, A.; et al. Effects of Daily Raspberry Consumption on Immune-Metabolic Health in Subjects at Risk of Metabolic Syndrome: A Randomized Controlled Trial. Nutrients 2020, 12, 3858. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12123858
Franck M, de Toro-Martín J, Garneau V, Guay V, Kearney M, Pilon G, Roy D, Couture P, Couillard C, Marette A, et al. Effects of Daily Raspberry Consumption on Immune-Metabolic Health in Subjects at Risk of Metabolic Syndrome: A Randomized Controlled Trial. Nutrients. 2020; 12(12):3858. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12123858
Chicago/Turabian StyleFranck, Maximilien, Juan de Toro-Martín, Véronique Garneau, Valérie Guay, Michèle Kearney, Geneviève Pilon, Denis Roy, Patrick Couture, Charles Couillard, André Marette, and et al. 2020. "Effects of Daily Raspberry Consumption on Immune-Metabolic Health in Subjects at Risk of Metabolic Syndrome: A Randomized Controlled Trial" Nutrients 12, no. 12: 3858. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12123858