Effects of Caffeine and Chlorogenic Acid on Nonalcoholic Steatohepatitis in Mice Induced by Choline-Deficient, L-Amino Acid-Defined, High-Fat Diet

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Experimental Protocol
2.2. Assessment of Serum Biochemical Parameters
2.3. Histopathological Analysis
2.4. Immunohistochemistry
2.5. Image Analysis of Histological Slides
2.6. Hepatic Levels of TG and T-Cho
2.7. 4-Hydroxynonenal (4-HNE) Levels in the Liver
2.8. Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR) Analysis
2.9. Statistical Analysis
3. Results
3.1. General Findings
3.2. Biochemical Data of the Serum
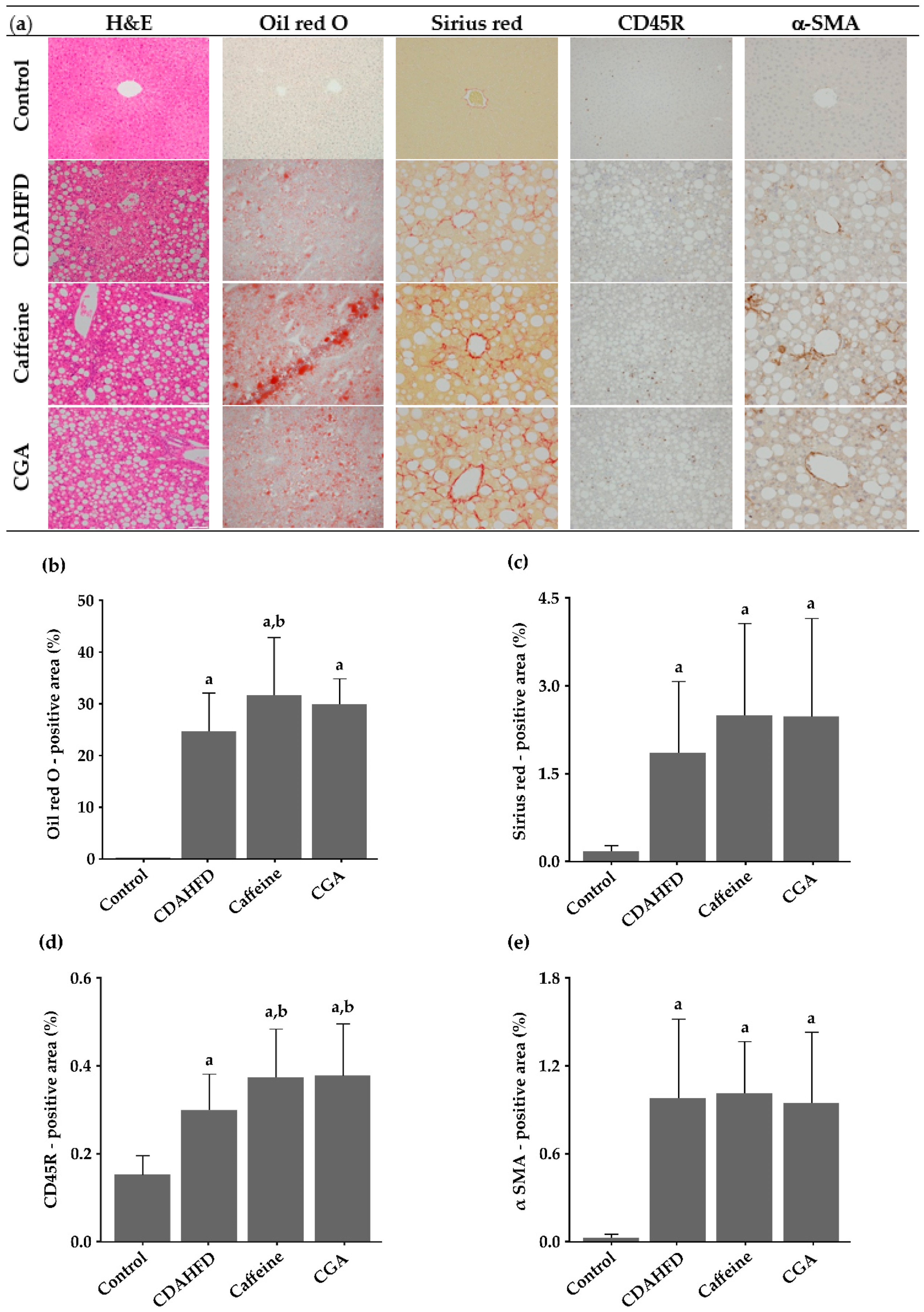
3.3. Histopathological Findings
3.4. Image Analysis of Histological Slides
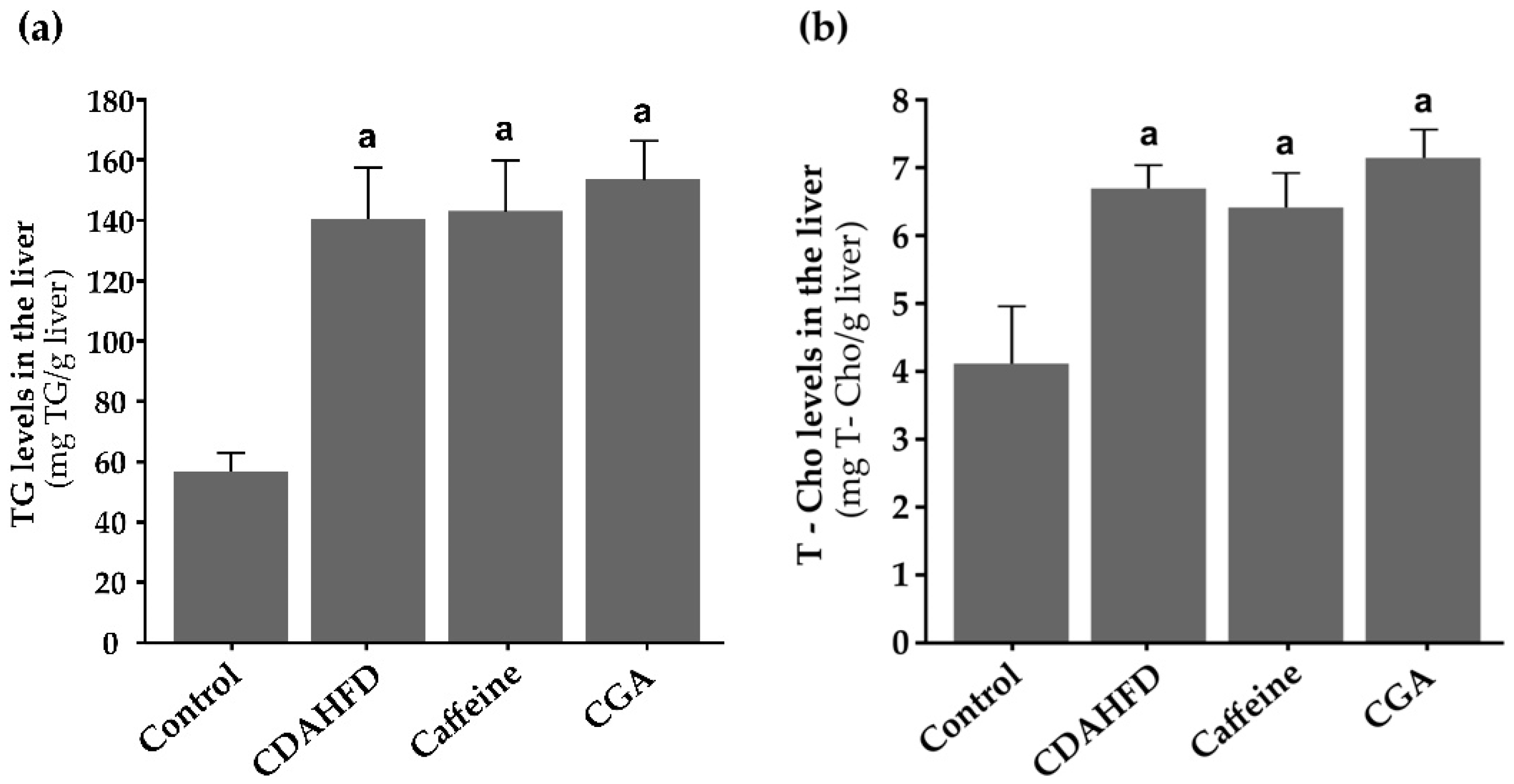
3.5. TG and T-Cho Levels in the Liver
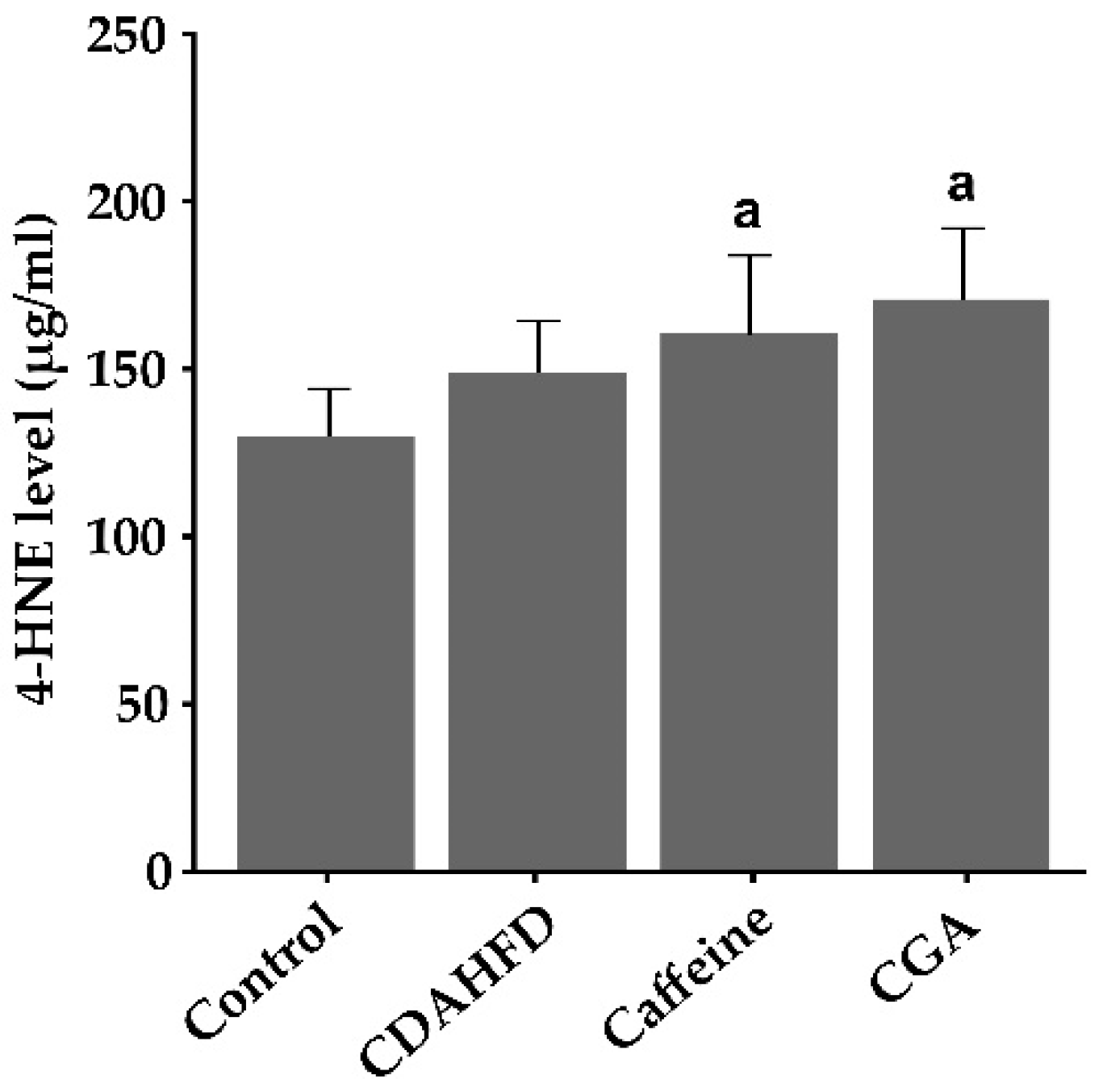
3.6. Hepatic 4-HNE Levels
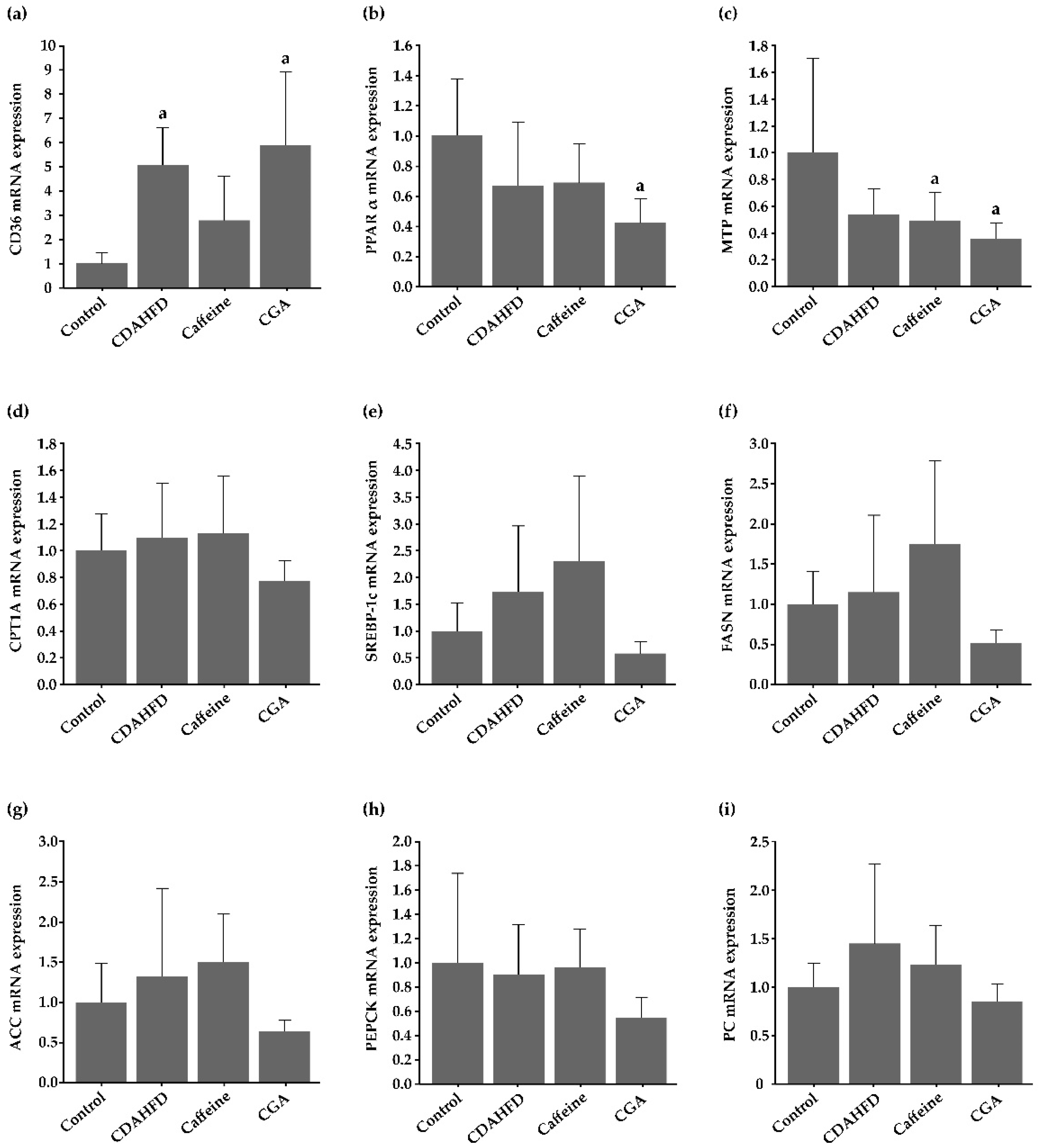
3.7. Gene Expression in the Hepatic Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Charlton, M.; Kasparova, P.; Weston, S.; Lindor, K.; Maor-Kendler, Y.; Wiesner, R.H.; Rosen, C.B.; Batts, K.P. Frequency of nonalcoholic steatohepatitis as a cause of advanced liver disease. Liver Transpl. 2001, 7, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.A.; Lymp, J.F.; St Sauver, J.; Sanderson, S.O.; Lindor, K.D.; Feldstein, A.; Angulo, P. The natural history of nonalcoholic fatty liver disease: A population-based cohort study. Gastroenterology 2005, 129, 113–121. [Google Scholar] [CrossRef]
- Yasui, K.; Hashimoto, E.; Komorizono, Y.; Koike, K.; Arii, S.; Imai, Y.; Shima, T.; Kanbara, Y.; Saibara, T.; Mori, T.; et al. Characteristics of patients with nonalcoholic steatohepatitis who develop hepatocellular carcinoma. Clin. Gastroenterol. Hepatol. 2011, 9, 428–433. [Google Scholar]
- Vernon, G.; Baranova, A.; Younossi, Z.M. Systematic review: The epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults. Aliment. Pharmacol. Ther. 2011, 34, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M. Non-alcoholic fatty liver disease—A global public health perspective. J. Hepatol. 2019, 70, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Butt, M.S.; Sultan, M.T. Coffee and its consumption: Benefits and risks. Crit. Rev. Food Sci. Nutr. 2011, 51, 363–373. [Google Scholar]
- Yesil, A.; Yilmaz, Y. Review article: Coffee consumption, the metabolic syndrome and non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2013, 38, 1038–1044. [Google Scholar] [CrossRef]
- Bhupathiraju, S.N.; Pan, A.; Manson, J.E.; Willett, W.C.; Van Dam, R.M.; Hu, F.B. Changes in coffee intake and subsequent risk of type 2 diabetes: Three large cohorts of US men and women. Diabetologia 2014, 57, 1346–1354. [Google Scholar] [CrossRef] [Green Version]
- Hernan, M.A.; Takkouche, B.; Caamano-Isorna, F.; Gestal-Otero, J.J. A meta-analysis of coffee drinking, cigarette smoking, and the risk of Parkinson’s disease. Ann. Neurol. 2002, 52, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Tran, K.T.; Coleman, H.G.; McMenamin, U.C.; Cardwell, C.R. Coffee consumption by type and risk of digestive cancer: A large prospective cohort study. Br. J. Cancer 2019, 120, 1059–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez-Grobe, Y.; Chavez-Tapia, N.; Sanchez-Valle, V.; Gavilanes-Espinar, J.G.; Ponciano-Rodriguez, G.; Uribe, M.; Mendez-Sanchez, N. High coffee intake is associated with lower grade nonalcoholic fatty liver disease: The role of peripheral antioxidant activity. Ann. Hepatol. 2012, 11, 350–355. [Google Scholar] [CrossRef]
- Bambha, K.; Wilson, L.A.; Unalp, A.; Loomba, R.; Neuschwander-Tetri, B.A.; Brunt, E.M.; Bass, N.M.; Nonalcoholic Steatohepatitis Clinical Research Network (NASH CRN). Coffee consumption in NAFLD patients with lower insulin resistance is associated with lower risk of severe fibrosis. Liver Int. 2014, 34, 1250–1258. [Google Scholar] [CrossRef] [Green Version]
- Vitaglione, P.; Mazzone, G.; Lembo, V.; D’Argenio, G.; Rossi, A.; Guido, M.; Savoia, M.; Salomone, F.; Mennella, I.; De Filippis, F.; et al. Coffee prevents fatty liver disease induced by a high-fat diet by modulating pathways of the gut-liver axis. J. Nutr. Sci. 2019, 8, e15. [Google Scholar] [CrossRef] [Green Version]
- Higdon, J.V.; Frei, B. Coffee and health: A review of recent human research. Crit. Rev. Food Sci. Nutr. 2006, 46, 101–123. [Google Scholar] [CrossRef]
- Astrup, A.; Toubro, S.; Cannon, S.; Hein, P.; Breum, L.; Madsen, J. Caffeine: A double-blind, placebo-controlled study of its thermogenic, metabolic, and cardiovascular effects in healthy volunteers. Am. J. Clin. Nutr. 1990, 51, 759–767. [Google Scholar] [CrossRef]
- Kogure, A.; Sakane, N.; Takakura, Y.; Umekawa, T.; Yoshioka, K.; Nishino, H.; Yamamoto, T.; Kawada, T.; Yoshikawa, T.; Yoshida, T. Effects of caffeine on the uncoupling protein family in obese yellow KK mice. Clin. Exp. Pharmacol. Physiol. 2002, 29, 391–394. [Google Scholar] [CrossRef]
- Yamauchi, R.; Kobayashi, M.; Matsuda, Y.; Ojika, M.; Shigeoka, S.; Yamamoto, Y.; Tou, Y.; Inoue, T.; Katagiri, T.; Murai, A.; et al. Coffee and caffeine ameliorate hyperglycemia, fatty liver, and inflammatory adipocytokine expression in spontaneously diabetic KK-Ay mice. J. Agric. Food Chem. 2010, 58, 5597–5603. [Google Scholar] [CrossRef]
- Barcelos, R.P.; Lima, F.D.; Carvalho, N.R.; Bresciani, G.; Royes, L.F. Caffeine effects on systemic metabolism, oxidative-inflammatory pathways, and exercise performance. Nutr. Res. 2020, 80, 1–17. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.J.; Li, Y.F.; Wang, G.E.; Tan, R.R.; Tsoi, B.; Mao, G.W.; Zhai, Y.J.; Cao, L.F.; Chen, M.; Kurihara, H.; et al. Caffeine ameliorates high energy diet-induced hepatic steatosis: Sirtuin 3 acts as a bridge in the lipid metabolism pathway. Food Funct. 2015, 6, 2578–2587. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Dai, W.; Chen, X.; Wang, K.; Zhang, W.; Liu, L.; Hou, J. Caffeine reduces hepatic lipid accumulation through regulation of lipogenesis and ER stress in zebrafish larvae. J. Biomed. Sci. 2015, 22, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helal, M.G.; Ayoub, S.E.; Elkashefand, W.F.; Ibrahim, T.M. Caffeine affects HFD-induced hepatic steatosis by multifactorial intervention. Hum. Exp. Toxicol. 2018, 37, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Cai, X.; Hayashi, S.; Hao, S.; Sakiyama, H.; Wang, X.; Yang, Q.; Akira, S.; Nishiguchi, S.; Fujiwara, N.; et al. Caffeine-stimulated muscle IL-6 mediates alleviation of non-alcoholic fatty liver disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Xia, L.; Luo, H.; Xu, Y.; Yu, H.; Xu, D.; Wang, H. Prenatal caffeine exposure increases the susceptibility to non-alcoholic fatty liver disease in female offspring rats via activation of GR-C/EBPα-SIRT1 pathway. Toxicology 2019, 417, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.W.; Wong, C.N.Y.; Pin, W.K.; Wong, M.H.Y.; Kwok, C.Y.; Chan, R.Y.K.; Yu, P.H.F.; Chan, S.W. Chlorogenic acid exhibits cholesterol lowering and fatty liver attenuating properties by up-regulating the gene expression of PPAR-α in hypercholesterolemic rats induced with a high-cholesterol diet. Phytother. Res. 2013, 27, 545–551. [Google Scholar] [CrossRef]
- Yan, H.; Gao, Y.Q.; Zhang, Y.; Wang, H.; Liu, G.S.; Lei, J.Y. Chlorogenic acid alleviates autophagy and insulin resistance by suppressing JNK pathway in a rat model of nonalcoholic fatty liver disease. J. Biosci. 2018, 43, 287–294. [Google Scholar] [CrossRef]
- Alqarni, I.; Bassiouni, Y.A.; Badr, A.M.; Ali, R.A. Telmisartan and/or chlorogenic acid attenuates fructose-induced non-alcoholic fatty liver disease in rats: Implications of cross-talk between angiotensin, the sphingosine kinase/spingoine-1-phosphate pathway, and TLR4 receptors. Biochem. Pharmacol. 2019, 164, 252–262. [Google Scholar] [CrossRef]
- Mubarak, A.; Hodgson, J.M.; Considine, M.J.; Croft, K.D.; Matthews, V.B. Supplementation of a high-fat diet with chlorogenic acid is associated with insulin resistance and hepatic lipid accumulation in mice. J. Agric. Food Chem. 2013, 61, 4371–4378. [Google Scholar] [CrossRef]
- Matsumoto, M.; Hada, N.; Sakamaki, Y.; Uno, A.; Shiga, T.; Tanaka, C.; Ito, T.; Katsume, A.; Sudoh, M. An improved mouse model that rapidly develops fibrosis in non-alcoholic steatohepatitis. Int. J. Exp. Pathol. 2013, 94, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Soejima, Y.; Kumagai, A.; Watanabe, M.; Uozaki, H.; Fukusato, T. Inhibitory effects of Japanese herbal medicines sho-saiko-to and juzen-taiho-to on nonalcoholic steatohepatitis in mice. PLoS ONE 2014, 9, e87279. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Watabe, S.; Togashi-Kumagai, A.; Watanabe, M.; Dungubat, E.; Kusano, H.; Kobayashi, Y.; Harada, N.; Yamaji, R.; Sugimoto, K.; et al. Effects of low ethanol consumption on nonalcoholic steatohepatitis in mice. Alcohol 2020, 87, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Birerdinc, A.; Stepanova, M.; Pawloski, L.; Younossi, Z.M. Caffeine is protective in patients with non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2012, 35, 76–82. [Google Scholar] [CrossRef]
- Shen, H.; Rodriguez, A.C.; Shiani, A.; Lipka, S.; Shahzad, G.; Kumar, A.; Mustacchia, P. Association between caffeine consumption and nonalcoholic fatty liver disease: A systemic review and meta-analysis. Therap. Adv. Gastroenterol. 2016, 9, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Josekutty, J.; Iqbal, J.; Iwawaki, T.; Kohno, K.; Hussain, M.M. Microsomal triglyceride transfer protein inhibition induces endoplasmic reticulum stress and increases gene transcription via Ire1α/cJun to enhance plasma ALT/AST. J. Biol. Chem. 2013, 288, 14372–14383. [Google Scholar] [CrossRef] [Green Version]
- Brandt, A.; Nier, A.; Jin, C.J.; Baumann, A.; Jung, F.; Ribas, V.; Garcia-Ruiz, C.; Fernandez-Checa, J.C.; Bergheim, I. Consumption of decaffeinated coffee protects against the development of early non-alcoholic steatohepatitis: Role of intestinal barrier function. Redox Biol. 2019, 21, 101092. [Google Scholar] [CrossRef]
- Zheng, G.; Sayama, K.; Okubo, T.; Juneja, L.R.; Oguni, I. Anti-obesity effects of three major components of green tea, catechins, caffeine and theanine, in mice. In Vivo 2004, 18, 55–62. [Google Scholar]
- Sugiura, C.; Nishimatsu, S.; Moriyama, T.; Ozasa, S.; Kawada, T.; Sayama, K. Catechins and caffeine inhibit fat accumulation in mice through the improvement of hepatic lipid metabolism. J. Obes. 2012, 2012, 520510. [Google Scholar] [CrossRef] [Green Version]
- Okumura, T.; Tsukui, T.; Hosokawa, M.; Miyashita, K. Effect of caffeine and capsaicin on the blood glucose levels of obese/diabetic KK-Ay mice. J. Oleo Sci. 2012, 61, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lam, K.L.; Hu, J.; Ge, S.; Zhou, A.; Zheng, B.; Zeng, S.; Lin, S. Chlorogenic acid alleviates obesity and modulates gut microbiota in high-fat-fed mice. Food Sci. Nutr. 2019, 7, 579–588. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zheng, S.; Sheng, Y.; Miao, T.; Xu, J.; Xu, W.; Huang, K.; Zhao, C. Chlorogenic acid ameliorates obesity by preventing energy balance shift in high-fat diet induced obese mice. J. Sci. Food Agric. 2020, 101, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Soejima, Y.; Fukusato, T. Animal models of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. World J. Gastroenterol. 2012, 18, 2300–2308. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Park, J.E.; Lee, M.; Hardwick, J.P. Hepatic lipid homeostasis by peroxisome proliferator-activated receptor gamma 2. Liver Res. 2018, 2, 209–215. [Google Scholar] [PubMed]
- Ameen, C.; Edvardsson, U.; Ljungberg, A.; Asp, L.; Akerblad, P.; Tuneld, A.; Olofsson, S.O.; Linden, D.; Oscarsson, J. Activation of peroxisome proliferator-activated receptor alpha increases the expression and activity of microsomal triglyceride transfer protein in the liver. J. Biol. Chem. 2005, 280, 1224–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagner, A.; Polizzi, A.; Fouche, E.; Ducheix, S.; Lippi, Y.; Lasserre, F.; Barquissau, V.; Regnier, M.; Lukowicz, C.; Benhamed, F. Liver PPARα is crucial for whole-body fatty acid homeostasis and is protective against NAFLD. Gut 2016, 65, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- Van Herck, M.A.; Vonghia, L.; Francque, S.M. Animal models of nonalcoholic fatty liver disease—A starter’s guide. Nutrients 2017, 9, 1072. [Google Scholar]
- Bertola, A. Rodent models of fatty liver diseases. Liver Res. 2018, 2, 3–13. [Google Scholar] [CrossRef]
- Kawai, D.; Takaki, A.; Nakatsuka, A.; Wada, J.; Tamaki, N.; Yasunaka, T.; Koike, K.; Tsuzaki, R.; Matsumoto, K.; Miyake, Y.; et al. Hydrogen-rich water prevents progression of nonalcoholic steatohepatitis and accompanying hepatocarcinogenesis in mice. Hepatology 2012, 56, 912–921. [Google Scholar] [CrossRef]
- Kim, S.B.; Kang, O.H.; Lee, Y.S.; Han, S.H.; Ahn, Y.S.; Cha, S.W.; Seo, Y.S.; Kong, R.; Kwon, D.Y. Hepatoprotective effect and synergism of Bisdemethoycurcumin against MCD diet-induced nonalcoholic fatty liver disease in mice. PLoS ONE 2016, 11, e0147745. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, S.; Horibe, S.; Sasaki, N.; Tanahashi, T.; Mizuno, S.; Hamaguchi, T.; Rikitake, Y. Inhibitory effects of sodium alginate on hepatic steatosis in mice induced by a methionine- and choline-deficient diet. Mar. Drugs 2019, 17, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohli, R.; Kirby, M.; Xanthakos, S.A.; Softic, S.; Feldstein, A.E.; Saxena, V.; Tang, P.H.; Miles, L.; Miles, M.V.; Balistreri, W.F.; et al. High-fructose, medium chain trans fat diet induces liver fibrosis and elevates plasma coenzyme Q9 in a novel murine model of obesity and nonalcoholic steatohepatitis. Hepatology 2010, 52, 934–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harpaz, E.; Tamir, S.; Weinstein, A.; Weinstein, Y. The effect of caffeine on energy balance. J. Basic Clin. Physiol. Pharmacol. 2017, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.E.; Savani, S.; Battram, D.S.; McLaren, D.H.; Sathasivam, P.; Graham, T.E. Caffeine ingestion before an oral glucose tolerance test impairs blood glucose management in men with type 2 diabetes. J. Nutr. 2004, 134, 2528–2533. [Google Scholar] [CrossRef]
- Beaudoin, M.S.; Allen, B.; Mazzetti, G.; Sullivan, P.J.; Graham, T.E. Caffeine ingestion impairs insulin sensitivity in a dose-dependent manner in both men and women. Appl. Physiol. Nutr. Metab. 2013, 38, 140–147. [Google Scholar] [CrossRef]
- Banks, N.F.; Tomko, P.M.; Colquhoun, R.J.; Muddle, T.W.D.; Emerson, S.R.; Jenkins, N.D.M. Genetic polymorphisms in ADORA2A and CYP1A2 influence caffeine’s effect on postprandial glycaemia. Sci. Rep. 2019, 9, 10532. [Google Scholar] [CrossRef]
- Jarrar, S.F.; Obeid, O.A. Timing of caffeine ingestion alters postprandial metabolism in rats. Nutrition 2014, 30, 107–111. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primers (5′→3′) | Reverse Primers (5′→3′) |
|---|---|---|
| CD36 | GCCAAGCTATTGCGACATGA | CAATGGTTGTCTGGATTCTGG |
| PPARα | AATGCAATTCGCTTTGGAAG | GGCCTTGACCTTGTTCATGT |
| MTP | CATCTCCACAGTGCAGTTCTCACA | GGAGTTCACATCCGGCCACTA |
| CPT 1A | GATCTACAATTCCCCTCTGCTCT | TAGAGCCAGACCTTGAAGTAACG |
| SREBP-1c | TGGACTACTAGTGTTGGCCTGCTT | ATCCAGGTCAGCTTGTTTGCGATG |
| FASN | TGGGTTCTAGCCAGCAGAGT | ACCACCAGAGACCGTTATGC |
| ACC | GGAGATGTACGCTGACCGAGAA | ACCCGACGCATGGTTTTCA |
| PEPCK | GGTGTTTACTGGGAAGGCATC | CAATAAGGGGCACTGGCTG |
| PC | GAGCTTATCCCGAACATCCC | TCCATACCATTCTCTTTGGCC |
| TBP | CTGGAATTGTACCGCAGCTT | ATGATGACTGCAGCAAATCG |
| Parameters | Experimental Groups | |||
|---|---|---|---|---|
| Control | CDAHFD | Caffeine | CGA | |
| Food consumption (g/day) | 3.0 ± 0.2 | 2.3 ± 0.3 a | 2.7 ± 0.4 a,b | 2.3 ± 0.3 a |
| Calorie intake (kcal/day) | 10.5 ± 0.5 | 12.1 ± 1.6 a | 14.0 ± 2.0 a,b | 12.1 ± 1.3 a |
| Body weight (g) | 25.3 ± 1.4 | 20.4 ± 0.8 a | 20.7 ± 1.2 a | 20.6 ± 0.5 a |
| Liver weight (g) | 1.0 ± 0.0 | 1.3 ± 0.2 a | 1.3 ± 0.2 a | 1.4 ± 0.0 a |
| Liver to body weight ratio (%) | 3.9 ± 0.3 | 6.5 ± 0.9 a | 6.6 ± 0.2 a | 6.9 ± 0.2 a |
| EAT weight (g) | 0.5 ± 0.1 | 0.3 ± 0.0 a | 0.3 ± 0.1 a | 0.3 ± 0.0 a |
| EAT to body weight ratio (%) | 2.0 ± 0.2 | 1.4 ± 0.3 a | 1.3 ± 0.3 a | 1.5 ± 0.1 a |
| Parameters | Experimental Groups | |||
|---|---|---|---|---|
| Control | CDAHFD | Caffeine | CGA | |
| AST (IU/L) | 175.3 ± 88.7 | 326.8 ± 25.1 a | 339.6 ± 27.6 a | 340.0 ± 61.4 a |
| ALT (IU/L) | 27.7 ± 4.3 | 272.3 ± 42.3 a | 342.4 ± 39.6 a,b | 349.7 ± 84.8 a,b |
| T-Cho (mg/dL) | 102.0 ± 3.2 | 47.3 ± 13.3 a | 42.3 ± 6.3 a | 55.2 ± 3.6 a |
| TG (mg/dL) | 75.7 ± 23.7 | 42.7 ± 16.9 a | 35.6 ± 2.6 a | 48.3 ± 14.1 a |
| Glucose (mg/dL) | 48.3 ± 9.2 | 65.0 ± 5.8 | 85.7 ± 40.9 a | 78.0 ± 19.9 |
| Insulin (pg/mL) | 226.8 ± 134.9 | 59.2 ± 62.0 a | 147.2 ± 125.2 | 48.2 ± 49.5 a |
| Adiponectin (µg/mL) | 15.4 ± 1.8 | 12.8 ± 0.2 a | 13.1 ± 0.9 a | 13.0 ± 1.2 a |
| Parameters | Experimental Groups | |||
|---|---|---|---|---|
| Control | CDAHFD | Caffeine | CGA | |
| Steatosis | 0 (0, 0) | 3 (3, 3) a | 3 (3, 3) a | 3 (3, 3) a |
| Intralobular inflammation | 0 (0, 0) | 3 (3, 3) a | 3 (3, 3) a | 3 (3, 3) a |
| Portal inflammation | 0 (0, 0) | 0.5 (0, 1) | 0.5 (0, 1) | 0 (0, 1) |
| Ballooning | 0 (0, 0) | 1 (1, 2) a | 1 (1, 2) a | 2 (1, 2) a |
| NAS | 0 (0, 0) | 7 (7, 8) a | 7 (7, 8) a | 8 (7, 8) a |
| Perisinusoidal fibrosis | 0 (0, 1) | 2 (2, 2) a | 2 (1, 2) a | 2 (2, 2) a |
| Fibrosis stage | 0 (0, 1) | 1 (1, 1) a | 1 (1, 1) a | 1 (1, 1) a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dungubat, E.; Watabe, S.; Togashi-Kumagai, A.; Watanabe, M.; Kobayashi, Y.; Harada, N.; Yamaji, R.; Fukusato, T.; Lodon, G.; Sevjid, B.; et al. Effects of Caffeine and Chlorogenic Acid on Nonalcoholic Steatohepatitis in Mice Induced by Choline-Deficient, L-Amino Acid-Defined, High-Fat Diet. Nutrients 2020, 12, 3886. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12123886
Dungubat E, Watabe S, Togashi-Kumagai A, Watanabe M, Kobayashi Y, Harada N, Yamaji R, Fukusato T, Lodon G, Sevjid B, et al. Effects of Caffeine and Chlorogenic Acid on Nonalcoholic Steatohepatitis in Mice Induced by Choline-Deficient, L-Amino Acid-Defined, High-Fat Diet. Nutrients. 2020; 12(12):3886. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12123886
Chicago/Turabian StyleDungubat, Erdenetsogt, Shiori Watabe, Arisa Togashi-Kumagai, Masato Watanabe, Yasuyuki Kobayashi, Naoki Harada, Ryoichi Yamaji, Toshio Fukusato, Galtsog Lodon, Badamjav Sevjid, and et al. 2020. "Effects of Caffeine and Chlorogenic Acid on Nonalcoholic Steatohepatitis in Mice Induced by Choline-Deficient, L-Amino Acid-Defined, High-Fat Diet" Nutrients 12, no. 12: 3886. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12123886