Almonds (Prunus Dulcis Mill. D. A. Webb): A Source of Nutrients and Health-Promoting Compounds

 ,
, 
 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Food Composition of the Different Varieties
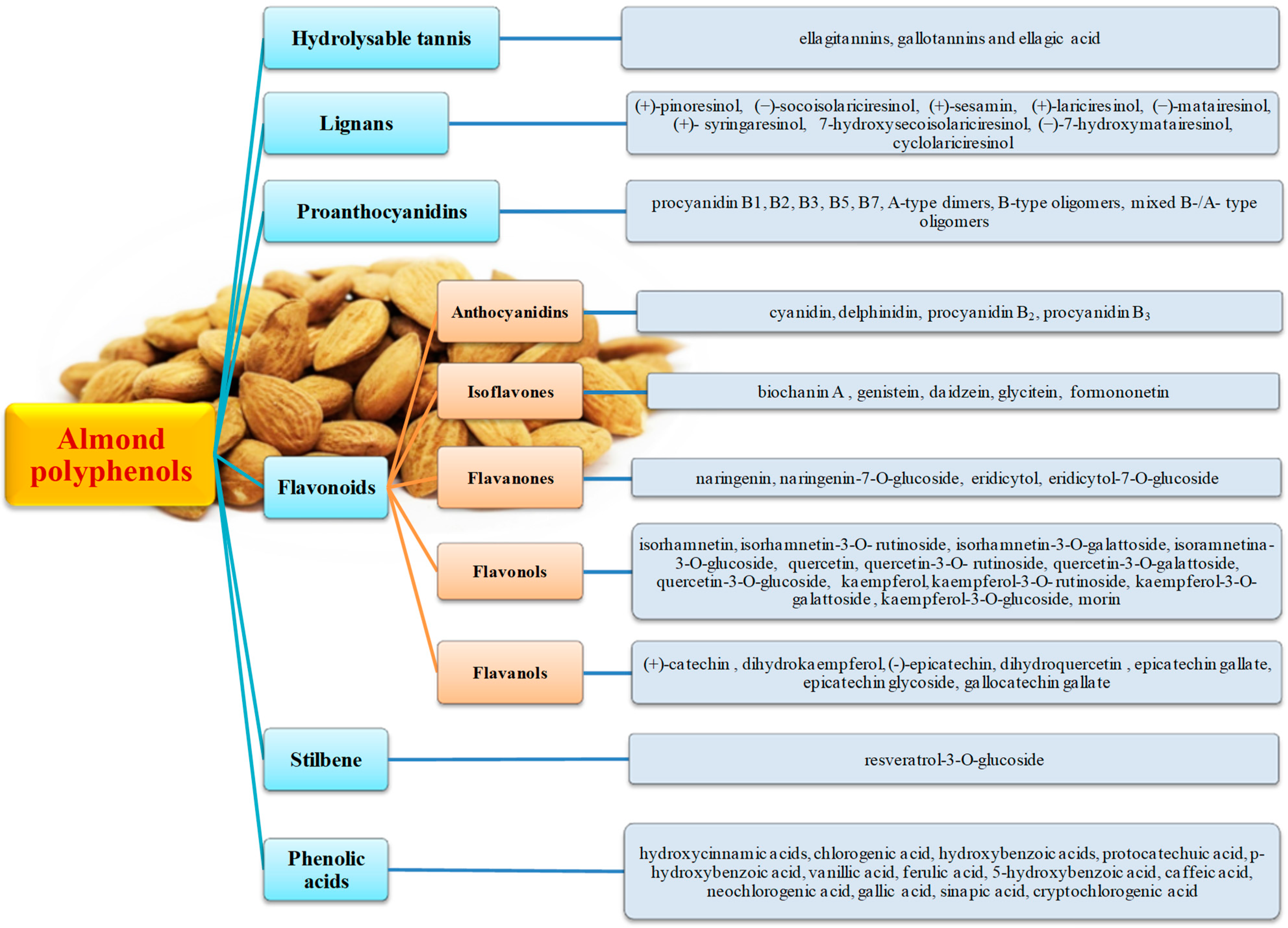
3. Main Almond Polyphenols
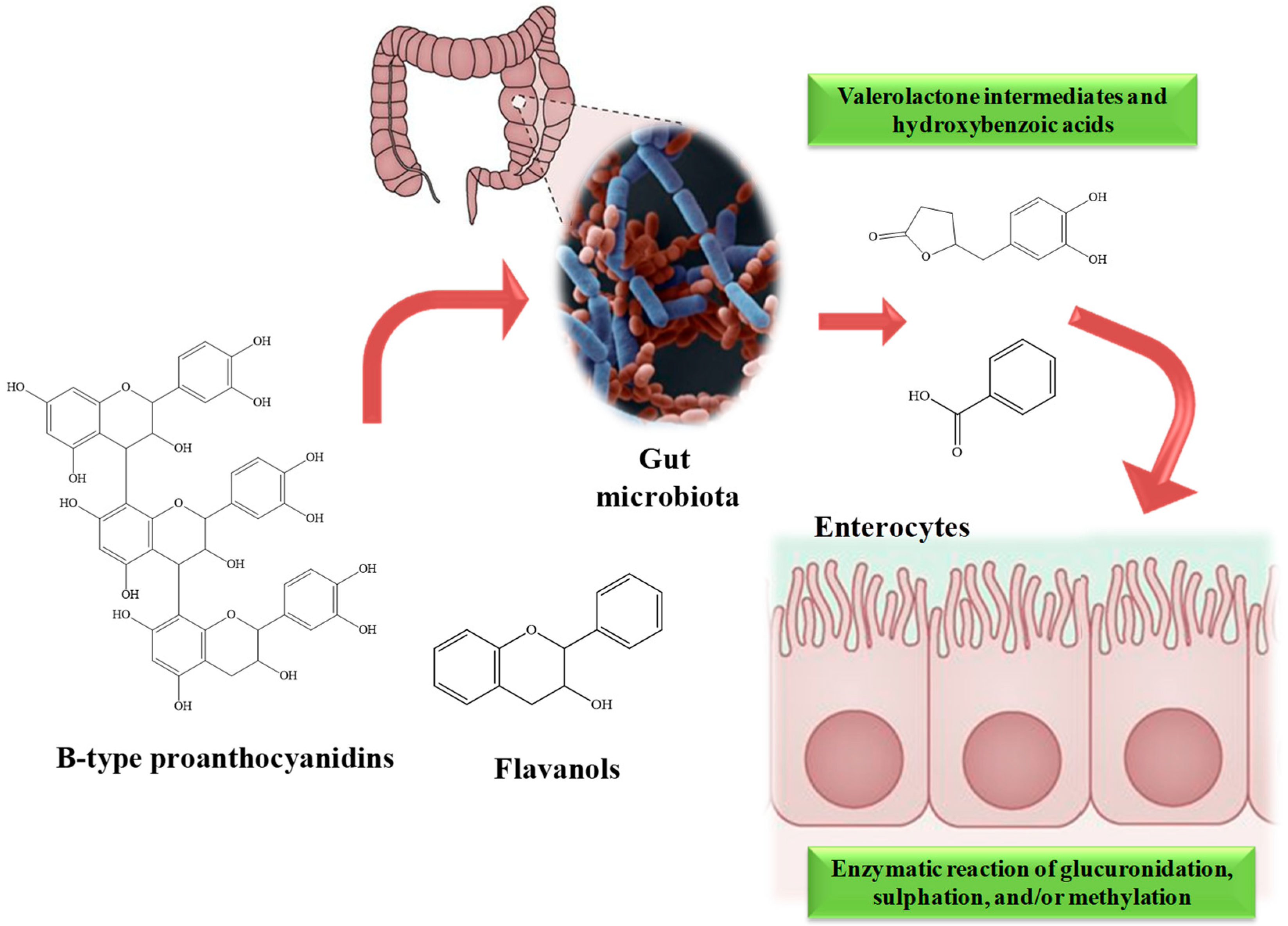
4. Bioavailability and Metabolism of Almond Components
5. Health-Promoting Properties: Evidence from Intervention Clinical Trials
5.1. Lipidemic Control
5.2. Glycemic Control
5.3. Obesity
5.4. Cardiovascular Risk
5.5. Inflammation and Oxidative Stress
5.6. Serum Uric Acid
5.7. Secondary Metabolites of Almonds as a Functional Food and Prebiotics
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AED | almond-enriched hypocaloric diet; |
| BMI | body mass index; |
| CHD | coronary heart disease; |
| CRP | C reactive protein; |
| GLP-1 | glucagon-like peptide-1; |
| HbA1c | glycosylated hemoglobin; |
| GSH-Px | glutathione peroxidase; |
| HDL-c | high-density lipoprotein cholesterol; |
| HOMA-IR | homeostatic model analysis for insulin resistance; |
| HOMA-β | homeostatic model analysis for beta-cell function; |
| LDL-c | low-density lipoprotein cholesterol; |
| MDA | malondialdehyde; |
| MUFA | monounsaturated fatty acids; |
| NFD | nut-free hypocaloric diet; |
| PUFA | polyunsaturated fatty acids; |
| SOD | superoxide dismutase; |
| TC | total cholesterol; |
| TG | triglycerides; |
| TNF | tumor-necrosis factor; |
| T2DM | type 2 diabetes mellitus; |
| 8-OH-dG | 8-hydroxy-2′-deoxyguanosine. |
References
- Casas-Agustench, P.; Salas-Huetos, A.; Salas-Salvadó, J. Mediterranean nuts: Origins, ancient medicinal benefits and symbolism. Public Health Nutr. 2011, 14, 2296–2301. [Google Scholar] [CrossRef]
- Becerra-Tomás, N.; Paz-Graniel, I.W.C.; Kendall, C.; Kahleova, H.; Rahelić, D.; Sievenpiper, J.L.; Salas-Salvadó, J. Nut consumption and incidence of cardiovascular diseases and cardiovascular disease mortality: A meta-analysis of prospective cohort studies. Nutr. Rev. 2019, 77, 691–709. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.G.M.; Schincaglia, R.M.; Pimentel, G.D.; Mota, J.F. Nuts and Human Health Outcomes: A Systematic Review. Nutrients 2017, 9, 1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechthold, A.; Boeing, H.; Schwedhelm, C.; Hoffmann, G.; Knüppel, S.; Iqbal, K.; Schwingshackl, L. Food groups and risk of coronary heart disease, stroke and heart failure: A systematic review and dose response meta-analysis of prospective studies. Crit. Rev. Food Sci. Nutr. 2017, 59, 1071–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Norat, T. Nut consumption and risk of cardiovascular disease, total cancer, all-cause and cause-specific mortality: A systematic review and dose-response meta-analysis of prospective studies. BMC Med. 2016, 14, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco Mejia, S.; Kendall, C.W.; Viguiliouk, E.; Augustin, L.S.; Ha, V.; Cozma, A.I.; Mirrahimi, A.; Maroleanu, A.; Chiavaroli, L.; Leiter, L.A.; et al. Effect of Tree Nuts on Metabolic Syndrome Criteria: A Systematic Review and Meta-analysis of Randomised Controlled Trials. BMJ Open 2014, 4, e004660. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, S.; Sabaté, J. Nuts, body weight and insulin resistance. Br. J. Nutr. 2006, 96, S79–S86. [Google Scholar] [CrossRef] [Green Version]
- Gradziel, T.M. Almond (Prunus dulcis) breeding. In Breeding Plantation Tree Crops: Temperate Species; Jain, S.M., Priyadarshan, P.M., Eds.; Springer: New York, NY, USA, 2009; pp. 1–31. ISBN 9780387712031. [Google Scholar]
- Oliveira, I.; Meyer, A.S.; Afonso, S.; Aires, A.; Goufo, P.; Trindade, H.; Gonçalves, B. Phenolic and fatty acid profiles, α-tocopherol and sucrose contents, and antioxidant capacities of understudied Portuguese almond cultivars. J. Food Biochem. 2019, 43, e12887. [Google Scholar] [CrossRef]
- Barreira, J.C.; Casal, S.; Ferreira, I.C.; Peres, A.M.; Pereira, J.A.; Oliveira, M.B. Supervised chemical pattern recognition in almond (Prunus dulcis) Portuguese PDO cultivars: PCA- and LDA-based triennial study. J. Agric. Food Chem. 2012, 60, 9697–9704. [Google Scholar] [CrossRef]
- Yada, S.; Lapsley, K.; Huang, G. A review of composition studies of cultivated almonds: Macronutrients and micronutrients. J. Food Compos. Anal. 2011, 24, 469–480. [Google Scholar] [CrossRef]
- Alamprese, C.; Ratti, S.; Rossi, M. Effects of roasting conditions on hazelnut characteristics in a two-step process. J. Food Eng. 2009, 95, 272–279. [Google Scholar] [CrossRef]
- Amaral, J.S.; Casal, S.; Seabra, R.M.; Oliveira, B.P. Effects of roasting on hazelnut lipids. J. Agric. Food Chem. 2006, 54, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, W.; Huang, G.; Zhang, W.; Ni, L. In vitro and in vivo evaluation of the prebiotic effect of raw and roasted almonds (Prunus amygdalus). J. Sci. Food Agric. 2016, 96, 1836–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandalari, G.; Nueno-Palop, C.; Bisignano, G.; Wickham, M.S.; Narbad, A. Potential prebiotic properties of almond (Amygdalus communis L.) seeds. Appl. Environ. Microbiol. 2008, 74, 4264–4270. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Lapsley, K.; Blumberg, J. A nutrition and health perspective on almonds. J. Sci. Agric. Food Sci. 2006, 86, 2245–2250. [Google Scholar] [CrossRef]
- Yada, S.; Huang, G.; Lapsley, K. Natural variability in the nutrient composition of california-grown almonds. J. Food Compos. Anal. 2013, 30, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Gama, T.; Wallace, H.M.; Trueman, S.J.; Bai, S.H. Variability in crude protein and mineral nutrient concentrations of almonds. Acta Hortic. 2018, 1219, 259–264. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture, A.R.S. U.S. Department of Agriculture, Agricultural Research Service. FoodData Central. 2019. Available online: https://fdc.nal.usda.gov/ (accessed on 1 March 2020).
- Tabela de Composição de Alimentos (TCA). Instituto Nacional de Saúde Doutor Ricardo Jorge, Lisboa, Version 3.2–2018. Available online: http://portfir.insa.pt/foodcomp/search (accessed on 4 October 2019).
- Pennington, J.A.T. Applications of food composition data: Data sources and considerations for use. J. Food Compos. Anal. 2008, 21, S3–S12. [Google Scholar] [CrossRef]
- Piscopo, A.; Romeo, F.V.; Petrovicova, B.; Poiana, M. Effect of the harvest time on kernel quality of several almond varieties (Prunus dulcis (Mill.) D.A. Webb). Sci. Hortic. 2010, 125, 41–46. [Google Scholar] [CrossRef]
- Kazantzis, I.; Nanos, G.D.; Stavroulakis, G.G. Effect of harvest time and storage conditions on almond kernel oil and sugar composition. J. Sci. Food Agric. 2003, 83, 354–359. [Google Scholar] [CrossRef]
- Summo, C.; Palasciano, M.; Angelis, D.D.; Paradiso, V.M.; Caponio, F.; Pasqualone, A. Evaluation of the chemical and nutritional characteristics of almonds (Prunus dulcis (Mill). D.A. Webb) as influenced by harvest time and cultivar. J. Sci. Food Agric. 2018, 98, 5647–5655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahrens, S.; Venkatachalam, M.; Mistry, A.M.; Lapsley, K.; Sathe, S.K. Almond (Prunis dulcis L.) protein quality. Plant Foods Hum. Nutr. 2005, 60, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, S.; Sanden, B.L.; Lampinen, B.D.; Saa, S.; Siddiqui, M.I.; Smart, D.R.; Olivos, A.; Shackel, K.A.; DeJong, T.; Brown, P.H. Seasonal changes in nutrient content and concentrations in a mature deciduous tree species: Studies in almond (Prunus dulcis (Mill.) D. A. Webb). Eur. J. Agron. 2015, 65, 52–68. [Google Scholar] [CrossRef]
- Barreira, J.C.M.; Pereira, J.A.; Oliveira, M.B.P.P.; Ferreira, I.C.F.R. Sugars profiles of different chestnut (Castanea sativa Mill.) and almond (Prunus dulcis) cultivars by HPLC-RI. Plant Foods Hum. Nutr. 2010, 65, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Balta, F.; Battal, P.; Fikret Balta, M.; Yoruk, H.I. Free sugar compositions based on kernel taste in almond genotypes Prunus dulcis from Eastern Turkey. Chem. Nat. Compd. 2009, 45, 221–224. [Google Scholar] [CrossRef]
- Gu, L.; Kelm, M.A.; Hammerstone, J.F.; Beecher, G.; Holden, J.; Haytowitz, D.; Prior, R.L. Screening of foods containing proanthocyanidins and their structural characterization using LC-MS/MS and thiolytic degradation. J. Agric. Food Chem. 2003, 51, 7513–7521. [Google Scholar] [CrossRef]
- Bolling, B.W.; Dolnikowski, G.; Blumberg, J.B.; Chena, C.-Y.O. Polyphenol content and antioxidant activity of California almonds depend on cultivar and harvest year. Food Chem. 2010, 122, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Bolling, B.W. Almond Polyphenols: Methods of Analysis, Contribution to Food Quality, and Health Promotion. Compr. Rev. Food Sci. Food Saf. 2017, 16, 346–368. [Google Scholar] [CrossRef] [Green Version]
- Bolling, B.W.; Blumberg, J.B.; Chen, C.O. The influence of roasting, pasteurisation, and storage on the polyphenol content and antioxidant capacity of California almond skins. Food Chem. 2010, 123, 1040–1047. [Google Scholar] [CrossRef] [Green Version]
- Monagas, M.; Garrido, I.; Lebrón-Aguilar, R.; Bartolome, B.; Gómez-Cordovés, C. Almond (Prunus dulcis(Mill.) D.A. Webb) skins as a potential source of bioactive polyphenols. J. Agric. Food Chem. 2007, 55, 8498–8507. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Torres, J.L. Analysis of proanthocyanidins in almond blanch water by HPLC–ESI–QqQ–MS/MS and MALDI–TOF/TOF MS. Food Res. Int. 2012, 49, 798–806. [Google Scholar] [CrossRef]
- De Pascual-Teresa, S.; Gutierrez-Fernandez, Y.; Rivas-Gonzalo, J.C.; Santos Buelga, C. Characterization of monomeric and oligomeric flavan-3-ols from unripe almond fruits. Phytochem. Anal. 1998, 9, 21–27. [Google Scholar] [CrossRef]
- Monagas, M.; Garrido, I.; Lebrón-Aguilar, R.; Gómez-Cordovés, C.; Rybarczyk, A.; Amarowicz, R.; Bartolomé, B. Comparative flavan-3-ol profile and antioxidant capacity of roasted peanut, hazelnut, and almond skins. J. Agric. Food Chem. 2009, 57, 10590–10599. [Google Scholar] [CrossRef]
- Tsujita, T.; Shintani, T.; Sato, H. Alpha-amylase inhibitory activity from nut seed skin polyphenols. 1. Purification and characterization of almond seed skin polyphenols. J. Agric. Food Chem. 2013, 61, 4570–4576. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Roto, A.V.; Bolling, B.W. Characterization of ellagitannins, gallotannins, and bound proanthocyanidins from California almond (Prunus dulcis) varieties. J. Agric. Food Chem. 2012, 60, 12151–12156. [Google Scholar] [CrossRef] [PubMed]
- Urpi-Sarda, M.; Garrido, I.; Monagas, M.; Gómez-Cordovés, C.; Medina-Remón, A.; Andres-Lacueva, C.; Bartolomé, B. Profile of plasma and urine metabolites after the intake of almond [Prunus dulcis (Mill.) D.A. Webb] polyphenols in humans. J. Agric. Food Chem. 2009, 57, 10134–10142. [Google Scholar] [CrossRef] [PubMed]
- Harnly, J.M.; Doherty, R.F.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Bhagwat, S.; Gebhardt, S. Flavonoid content of U.S. fruits, vegetables, and nuts. J. Agric. Food Chem. 2006, 54, 9966–9977. [Google Scholar] [CrossRef]
- Wijeratne, S.S.K.; Abou-Zaid, M.M.; Shahidi, F. Antioxidant polyphenols in almond and its coproducts. J. Agric. Food Chem. 2006, 54, 312–318. [Google Scholar] [CrossRef]
- Arráez-Román, D.; Fu, S.; Sawalha, S.M.S.; Segura-Carretero, A.; Fernández-Gutiérrez, A. HPLC/CE-ESI-TOF-MS methods for the characterization of polyphenols in almond-skin extracts. Electrophoresis 2010, 31, 2289–2296. [Google Scholar] [CrossRef]
- Takeoka, G.R.; Dao, L.T. Antioxidant constituents of almond [Prunus dulcis (Mill.) D.A. Webb] Hulls. J. Agric. Food Chem. 2003, 51, 496–501. [Google Scholar] [CrossRef]
- Mandalari, G.; Tomaino, A.; Arcoraci, T.; Martorana, M.; LoTurco, V.; Cacciola, F.; Rich, G.T.; Bisignano, C.; Saija, A.; Dugo, P.; et al. Characterization of polyphenols, lipids and dietary fibre from almond skins (Amygdalus communis L.). J. Food Comp. Anal. 2010, 23, 166–174. [Google Scholar] [CrossRef]
- Thompson, L.; Boucher, B.; Liu, Z.; Cotterchio, M.; Kreiger, N. Phytoestrogen content of foods consumed in Canada, including isoflavones, lignans, and coumestan. Nutr. Cancer 2006, 54, 184–201. [Google Scholar] [CrossRef] [PubMed]
- Kuhnle, G.G.C.; Dell’Aquila, C.; Aspinall, S.M.; Runswick, S.A.; Mulligan, A.A.; Bingham, S.A. Phytoestrogen content of beverages, nuts, seeds, and oils. J. Agric. Food Chem. 2008, 56, 7311–7315. [Google Scholar] [CrossRef]
- Bornhorst, G.M.; Roman, M.J.; Dreschler, K.C.; Singh, R.P. Physical property changes in raw and roasted almonds during gastric digestion in vivo and in vitro. Food Biophys. 2013, 9, 39–48. [Google Scholar] [CrossRef]
- Bornhorst, G.M.; Roman, M.J.; Rutherfurd, S.M.; Burri, B.J.; Moughan, P.J.; Singh, R.P. Gastric digestion of raw and roasted almonds in vivo. J. Food Sci. 2013, 78, H1807–H1813. [Google Scholar] [CrossRef] [PubMed]
- Grundy, M.M.; Grassby, T.; Mandalari, G.; Waldron, K.W.; Butterworth, P.J.; Berry, S.E.; Ellis, P.R. Effect of mastication on lipid bioaccessibility of almonds in a randomized human study and its implications for digestion kinetics, metabolizable energy, and postprandial lipemia. Am. J. Clin. Nutr. 2015, 101, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Grundy, M.M.-L.; Wilde, P.J.; Butterworth, P.J.; Gray, R.; Ellis, P.R. Impact of cell wall encapsulation of almonds on in vitro duodenal lipolysis. Food Chem. 2015, 185, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Mandalari, G.; Grundy, M.M.L.; Grassby, T.; Parker, M.L.; Cross, K.L.; Chessa, S.; Butterworth, P.J. The effects of processing and mastication on almond lipid bioaccessibility using novel methods of in vitro digestion modelling and micro-structural analysis. Br. J. Nutr. 2014, 112, 1521–1529. [Google Scholar] [CrossRef] [Green Version]
- Bartolomé, B.; Monagas, M.; Garrido, I.; Gómez-Cordovés, C.; Martín-Alvarez, P.J.; Lebrón-Aguilar, R.; Urpí-Sardà, M.; Llorach, R.; Andrés-Lacueva, C. Almond (Prunus dulcis (Mill.) D.A. Webb) polyphenols: From chemical characterization to targeted analysis of phenolic metabolites in humans. Arch. Biochem. Biophys. 2010, 501, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Garrido, M.; Urpi-Sarda, M.; Monagas, C.; Gomez-Cordoves, P.J.; Martin-Alvarez, R.; Llorach, B.; Bartolome, C.; Andres-Lacueva, C. Targeted analysis of conjugated and microbial-derived phenolic metabolites in human urine after consumption of an almond skin phenolic extract. J. Nutr. 2010, 140, 1799–1807. [Google Scholar] [CrossRef] [Green Version]
- Albala, K. Almonds along the Silk Road: The exchange and adaptation of ideas from West to East. Petits Propos Culin. 2009, 88, 19–34. [Google Scholar]
- Ryan, N.T. World almond market. In Almonds: Botany, Production and Uses; Socias i Company, R., Gradizel, T.M., Eds.; CABI: Wallingford, UK, 2017; pp. 449–459. [Google Scholar]
- Kodad, O.; Socias, I.C.R.; Alonso, J.M. Genotypic and Environmental Effects on Tocopherol Content in Almond. Antioxidants 2018, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyson, D.A.; Schneeman, B.O.; Davis, P.A. Almonds and almond oil have similar effects on plasma lipids and LDL oxidation in healthy men and women. J. Nutr. 2002, 132, 703–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bento, A.P.; Cominetti, C.; Simoes Filho, A.; Naves, M.M. Baru almond improves lipid profile in mildly hypercholesterolemic subjects: A randomized, controlled, crossover study. Nutr. Metab. Cardiovasc. Dis. NMCD 2014, 24, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hwang, H.J.; Kim, H.S.; Park, H. Time and Intervention Effects of Daily Almond Intake on the Changes of Lipid Profile and Body Composition among Free-Living Healthy Adults. J. Med. Food 2018, 21, 340–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zibaeenezhad, M.J.; Ostovan, P.; Mosavat, S.H.; Zamirian, M.; Attar, A. Almond oil for patients with hyperlipidemia: A randomized open-label controlled clinical trial. Complement. Ther. Med. 2019, 42, 33–36. [Google Scholar] [CrossRef]
- Jenkins, D.J.; Kendall, C.W.; Josse, A.R.; Salvatore, S.; Brighenti, F.; Augustin, L.S.; Ellis, P.R.; Vidgen, E.; Rao, A.V. Almonds decrease postprandial glycemia, insulinemia, and oxidative damage in healthy individuals. J. Nutr. 2006, 136, 2987–2992. [Google Scholar] [CrossRef] [Green Version]
- Cohen, A.E.; Johnston, C.S. Almond ingestion at mealtime reduces postprandial glycemia and chronic ingestion reduces hemoglobin A(1c) in individuals with well-controlled type 2 diabetes mellitus. Metab. Clin. Exp. 2011, 60, 1312–1317. [Google Scholar] [CrossRef]
- Dhillon, J.; Thorwald, M.; De La Cruz, N.; Vu, E.; Asghar, S.A.; Kuse, Q.; Diaz Rios, L.K.; Ortiz, R.M. Glucoregulatory and Cardiometabolic Profiles of Almond vs. Cracker Snacking for 8 Weeks in Young Adults: A Randomized Controlled Trial. Nutrients 2018, 10, 960. [Google Scholar] [CrossRef] [Green Version]
- Gulati, S.; Misra, A.; Pandey, R.M. Effect of Almond Supplementation on Glycemia and Cardiovascular Risk Factors in Asian Indians in North India with Type 2 Diabetes Mellitus: A 24-Week Study. Metab. Syndr. Relat. Disord. 2017, 15, 98–105. [Google Scholar] [CrossRef]
- Li, S.C.; Liu, Y.H.; Liu, J.F.; Chang, W.H.; Chen, C.M.; Chen, C.Y. Almond consumption improved glycemic control and lipid profiles in patients with type 2 diabetes mellitus. Metab. Clin. Exp. 2011, 60, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Sabate, J.; Haddad, E.; Tanzman, J.S.; Jambazian, P.; Rajaram, S. Serum lipid response to the graduated enrichment of a Step I diet with almonds: A randomized feeding trial. Am. J. Clin. Nutr. 2003, 77, 1379–1384. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, J.C.; Most, M.M.; Lefevre, M.; Greenway, F.L.; Rood, J.C. Effect of diets enriched in almonds on insulin action and serum lipids in adults with normal glucose tolerance or type 2 diabetes. Am. J. Clin. Nutr. 2002, 76, 1000–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, L.W.; Balasubramanyam, A.; Kimball, K.T.; Aherns, A.K.; Fordis, C.M., Jr.; Ballantyne, C.M. Long-term, randomized clinical trial of two diets in the metabolic syndrome and type 2 diabetes. Diabetes Care 2003, 26, 2481–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Waldron, K.W.; Pacey, J.F.; Brain, A.; Ellis, P.R. Chemical and histochemical characterization of cell wall polysaccharides in almond seeds in relation to lipid bioavailability. In Biologically Active Phytochemicals in Food; Pfannhauser, W., Fenwick, G.R., Khokhar, S., Eds.; Royal Society of Chemistry: Cambridge, UK, 2001; pp. 448–452. [Google Scholar]
- Dhillon, J.; Tan, S.Y.; Mattes, R.D. Almond Consumption during Energy Restriction Lowers Truncal Fat and Blood Pressure in Compliant Overweight or Obese Adults. J. Nutr. 2016, 146, 2513–2519. [Google Scholar] [CrossRef] [Green Version]
- Foster, G.D.; Shantz, K.L.; Vander Veur, S.S.; Oliver, T.L.; Lent, M.R.; Virus, A.; Szapary, P.O.; Rader, D.J.; Zemel, B.S.; Gilden-Tsai, A. A randomized trial of the effects of an almond-enriched, hypocaloric diet in the treatment of obesity. Am. J. Clin. Nutr. 2012, 96, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Berryman, C.E.; West, S.G.; Fleming, J.A.; Bordi, P.L.; Kris-Etherton, P.M. Effects of daily almond consumption on cardiometabolic risk and abdominal adiposity in healthy adults with elevated LDL-cholesterol: A randomized controlled trial. J. Am. Heart Assoc. 2015, 4, e000993. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Holbrook, M.; Duess, M.A.; Dohadwala, M.M.; Hamburg, N.M.; Asztalos, B.F.; Milbury, P.E.; Blumberg, J.B.; Vita, J.A. Effect of almond consumption on vascular function in patients with coronary artery disease: A randomized, controlled, cross-over trial. Nutr. J. 2015, 14, 61. [Google Scholar] [CrossRef] [Green Version]
- Jalali-Khanabadi, B.A.; Mozaffari-Khosravi, H.; Parsaeyan, N. Effects of almond dietary supplementation on coronary heart disease lipid risk factors and serum lipid oxidation parameters in men with mild hyperlipidemia. J. Altern. Complement. Med. 2010, 16, 1279–1283. [Google Scholar] [CrossRef]
- Jenkins, D.J.; Kendall, C.W.; Marchie, A.; Parker, T.L.; Connelly, P.W.; Qian, W.; Haight, J.S.; Faulkner, D.; Vidgen, E.; Lapsley, K.G.; et al. Dose response of almonds on coronary heart disease risk factors: Blood lipids, oxidized low-density lipoproteins, lipoprotein(a), homocysteine, and pulmonary nitric oxide: A randomized, controlled, crossover trial. Circulation 2002, 106, 1327–1332. [Google Scholar] [CrossRef] [Green Version]
- Nishi, S.; Kendall, C.W.; Gascoyne, A.M.; Bazinet, R.P.; Bashyam, B.; Lapsley, K.G.; Augustin, L.S.; Sievenpiper, J.L.; Jenkins, D.J. Effect of almond consumption on the serum fatty acid profile: A dose-response study. Br. J. Nutr. 2014, 112, 1137–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wien, M.; Bleich, D.; Raghuwanshi, M.; Gould-Forgerite, S.; Gomes, J.; Monahan-Couch, L.; Oda, K. Almond consumption and cardiovascular risk factors in adults with prediabetes. J. Am. Coll. Nutr. 2010, 29, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, S.; Connell, K.M.; Sabate, J. Effect of almond-enriched high-monounsaturated fat diet on selected markers of inflammation: A randomised, controlled, crossover study. Br. J. Nutr. 2010, 103, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Li, N.; Zhang, W.; Zhang, X.; Lapsley, K.; Huang, G.; Blumberg, J.; Ma, G.; Chen, J. A pilot study on the effects of almond consumption on DNA damage and oxidative stress in smokers. Nutr. Cancer 2006, 54, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jia, X.; Chen, C.Y.; Blumberg, J.B.; Song, Y.; Zhang, W.; Zhang, X.; Ma, G.; Chen, J. Almond consumption reduces oxidative DNA damage and lipid peroxidation in male smokers. J. Nutr. 2007, 137, 2717–2722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamshed, H.; Gilani, A.U.; Sultan, F.A.; Amin, F.; Arslan, J.; Ghani, S.; Masroor, M. Almond supplementation reduces serum uric acid in coronary artery disease patients: A randomized controlled trial. Nutr. J. 2016, 15, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phung, O.J.; Makanji, S.S.; White, C.M.; Coleman, C.I. Almonds have a neutral effect on serum lipid profiles: A meta-analysis of randomized trials. J. Am. Diet. Assoc. 2009, 109, 865–873. [Google Scholar] [CrossRef]
- Musa-Veloso, K.; Paulionis, L.; Poon, T.; Lee, H.Y. The effects of almond consumption on fasting blood lipid levels: A systematic review and meta-analysis of randomised controlled trials. J. Nutr. Sci. 2016, 5, e34. [Google Scholar] [CrossRef] [Green Version]
- Farzaei, M.H.; Singh, A.K.; Kumar, R.; Croley, C.R.; Pandey, A.K.; Coy-Barrera, E.; Kumar Patra, J.; Das, G.; Kerry, R.G.; Annunziata, G.; et al. Targeting Inflammation by Flavonoids: Novel Therapeutic Strategy for Metabolic Disorders. Int. J. Mol. Sci. 2019, 20, 4957. [Google Scholar] [CrossRef] [Green Version]
- Lankisch, P.G.; Otto, J. Salivary isoamylase in duodenal aspirates. Dig. Dis. Sci. 1986, 31, 1299–1302. [Google Scholar] [CrossRef]
- Kawamori, R.; Katsura, M.; Ishida, S.; Yamasaki, Y.; Tujii, M.; Kawano, S.; Kamada, T. Subclinical exocrine pancreatic derangement in human diabetic patients evaluated from pure pancreatic juice. J. Diabetes Complicat. 1995, 9, 69–73. [Google Scholar] [CrossRef]
- Garrido, I.; Monagas, M.; Gómez-Cordovés, C.; Bartolomé, B. Polyphenols and antioxidant properties of almond skins: Influence of industrial processing. J. Food Sci. 2008, 73, C106–C115. [Google Scholar] [CrossRef] [PubMed]
- Milbury, P.E.; Chen, C.Y.; Dolnikowski, G.G.; Blumberg, J.B. Determination of flavonoids and phenolics and their distribution in almonds. J. Agric. Food Chem. 2006, 54, 5027–5033. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.O.; Milbury, P.E.; Blumberg, J.B. Polyphenols in Almond Skins after Blanching Modulate Plasma Biomarkers of Oxidative Stress in Healthy Humans. Antioxidants 2019, 8, 95. [Google Scholar] [CrossRef] [Green Version]
- Mandalari, G.; Bisignano, C.; D’Arrigo, M.; Ginestra, G.; Arena, A.; Tomaino, A.; Wickham, M.S. Antimicrobial potential of polyphenols extracted from almond skins. Lett. Appl. Microbiol. 2010, 51, 83–89. [Google Scholar] [CrossRef]
- Musarra-Pizzo, M.; Ginestra, G.; Smeriglio, A.; Pennisi, R.; Sciortino, M.T.; Mandalari, G. The Antimicrobial and Antiviral Activity of Polyphenols from Almond (Prunus dulcis L.) Skin. Nutrients 2019, 11, 2355. [Google Scholar] [CrossRef] [Green Version]
- Faustino, M.; Veiga, M.; Sousa, P.; Costa, E.M.; Silva, S.; Pintado, M. Agro-Food Byproducts as a New Source of Natural Food Additives. Molecules 2019, 24, 1056. [Google Scholar] [CrossRef] [Green Version]
- Teets, A.S.; Were, L.M. Inhibition of lipid oxidation in refrigerated and frozen salted raw minced chicken breasts with electron beam irradiated almond skin powder. Meat Sci. 2008, 80, 1326–1332. [Google Scholar] [CrossRef]
- Prasetyo, M.; Chia, M.; Hughey, C.; Were, L.M. Utilization of electron beam irradiated almond skin powder as a natural antioxidant in ground top round beef. J. Food Sci. 2007, 73, T1–T6. [Google Scholar] [CrossRef]
- Wijeratne, S.S.K.; Amarowicz, R.; Shahidi, F. Antioxidant activity of almonds and their by-products in food model systems. J. Am. Oil Chem. Soc. 2006, 83, 223–230. [Google Scholar] [CrossRef]
- Liu, Z.; Lin, X.; Huang, G.; Zhang, W.; Rao, P.; Ni, L. Prebiotic effects of almonds and almond skins on intestinal microbiota in healthy adult humans. Anaerobe 2014, 26, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ukhanova, M.; Wang, X.; Baer, D.J.; Novotny, J.A.; Fredborg, M.; Mai, V. Effects of almond and pistachio consumption on gut microbiota composition in a randomised cross-over human feeding study. Br. J. Nutr. 2014, 111, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Faulks, R.M.; Bisignano, C.; Waldron, K.W.; Narbad, A.; Wickham, M.S. In vitro evaluation of the prebiotic properties of almond skins (Amygdalus communis L.). FEMS Microbiol. Lett. 2010, 304, 116–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Variety | Unknown | Unknown | Butte | Carmel | Desmajo Largueta | Ferragnés | Filippo Ceo | Francoli | Fritz | Genco | Johnston Prolific | Marcona | Mission | Monterey | Nonpareil | Sonora | Texas | Thompson | Tuono | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nutrients | |||||||||||||||||||||||
| Water | 4.4 | 4.9 | 4.7 | 3.1 | 4.1 | 5.7 | 6.5 | 5.4 | 5.7 | 4.6 | 6.5 | 5.4 | 6.1 | 4.3 | 4.6 | 3.9 | 4.0 | 3.9 | 4.1 | 6.5 | 5.3 | 5.3 | |
| Protein | 21.2 | 21.6 | 20.5 | 20.6 | 20.2 | 19.6 | 18.1 | 14.1 | 20.5 | 22.5 | 21.5 | 20.2 | 22.1 | 23.3 | 20.9 | 21.3 | 21.0 | 20.2 | 22.4 | 20.1 | 21.2 | 20.5 | |
| Total lipids | 49.9 | 56.0 | 50.0 | 47.5 | 50.1 | 50.6 | 50.1 | 56.2 | 44.3 | 48.4 | 42.4 | 47.0 | 52.7 | 44.1 | 49.6 | 49.4 | 43.3 | 49.6 | 50.2 | 48.8 | 48.0 | 47.3 | |
| SFA | 4.1 | 3.9 | 3.4 | 3.7 | 3.7 | 3.8 | 3.9 | ||||||||||||||||
| MUFA | 29.4 | 29.7 | 30.5 | 31.6 | 32.3 | 31.3 | 31.4 | ||||||||||||||||
| PUFA | 13.9 | 13.8 | 12.0 | 11.6 | 11.2 | 11.7 | 12.4 | ||||||||||||||||
| Ash | 3.0 | 3.0 | 2.8 | 3.7 | 2.9 | 3.2 | 2.9 | 2.5 | 3.5 | 2.9 | 3.0 | 2.6 | 3.4 | 4.6 | 3.0 | 3.0 | 4.5 | 2.9 | 3.0 | 2.9 | 3.1 | 3.0 | |
| Carbohydrate | 21.6 | 19.2 | 25.0 | 20.9 | 22.4 | 21.8 | 26.1 | 26.6 | 24.8 | 15.7 | 23.6 | 27.0 | 21.7 | 22.4 | 23.9 | ||||||||
| Fiber | 12.5 | 12.0 | 12.2 | 12.5 | 11.0 | 13.5 | 11.8 | 12.9 | 11.8 | ||||||||||||||
| Sugars | 4.4 | 5.4 | 6.2 | 7.5 | |||||||||||||||||||
| References | [19] | [20] | [17] | [25] | [17] | [24] | [24] | [24] | [24] | [17] | [24] | [24] | [24] | [25] | [17] | [17] | [25] | [17] | [17] | [24] | [24] | [24] | |
| Variety | Unknown | Unknown | Mixture | Butte | Carmel | Falsa Barese | Ferragnés | Fritz | Genco | Glorieta | Lauranne | Mas Bovera | Mission | Monterey | Nonpareil/Monterey | Nonpareil | Pepparuda | Sonora | Stelliette | Supernova | Trianella | Tuono | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nutrients | |||||||||||||||||||||||
| Aluminum (Al) | 0.39 | ||||||||||||||||||||||
| Boron (B) | 1.85 | ||||||||||||||||||||||
| Calcium (Ca) | 269 | 270 | 200 | 288 | 279 | 137–142 | 128–138 | 290 | 138 | 90–102 | 166–176.5 | 103–129 | 330 | 252 | 172–252 | 261 | 118–131 | 234 | 155–176 | 113–127 | 106–125 | 106–127 | |
| Iron (Fe) | 3.7 | 4 | 3.22 | 3.27 | 3.27 | 3.13–4.45 | 3.42–4.48 | 3.63 | 1.82–4.29 | 4.36–6.10 | 4.45–4.97 | 1.56–4.73 | 3.34 | 3.58 | 3.47 | 2.84–3.45 | 3.84 | 4.09–4.27 | 5.54–6.49 | 2.80–5.38 | 2.71–5.11 | ||
| Magnesium (Mg) | 270 | 260 | 300 | 263 | 262 | 235–254 | 226–233 | 260 | 254–235 | 245–249 | 154–246 | 256–276 | 272 | 278 | 121–145 | 275 | 223–246 | 256 | 241–249 | 218–234 | 226–241 | 222.5–225.5 | |
| Phosphorus (P) | 481 | 410 | 500 | 463 | 462 | 466 | 512 | 524 | 191–258 | 455 | 526 | ||||||||||||
| Potassium (K) | 733 | 860 | 600 | 664 | 679 | 690–726 | 661–695 | 664 | 572–674 | 694–727 | 652–737 | 733–786 | 724 | 766 | 1550–2370 | 762 | 729–745 | 773 | 699–794 | 659–672 | 525–727 | 676–741 | |
| Sodium (Na) | 1 | 6 | 3.54 | ||||||||||||||||||||
| Sulphur (S) | 200 | ||||||||||||||||||||||
| Zinc (Zn) | 3.1 | 3.1 | 21.3 | 2.98 | 2.77 | 2.83–5.51 | 3.71–4.89 | 2.82 | 3.95–4.69 | 4.66–5.55 | 4.45–5.93 | 3.88–5.46 | 2.76 | 2.79 | 3.23 | 1.08–4.99 | 3.8 | 4.37–5.53 | 3.25–5.30 | 4.38–5.37 | 4.3–5.1 | ||
| Copper (Cu) | 1 | 0.89 | 0.92 | 1.09 | 1.98–3.38 | 2.14–3.80 | 0.85 | 1.83–3.90 | 1.93–3.85 | 2.18–3.76 | 1.68–3.82 | 0.72 | 0.94 | 1.05 | 2.27–8.61 | 0.9 | 1.76–3.72 | 2.16–2.89 | 2.06–3.74 | 2.04–3.58 | |||
| Manganese (Mn) | 2.2 | 2.56 | 2 | 2.14 | 1.01–1.27 | 1.04–1.46 | 2.08 | 1.16–1.29 | 1.11–1.12 | 1.22–1.44 | 1.39 | 2.2 | 2.12 | 2.21 | 0.89–0.95 | 3.04 | 1.21–1.32 | 0.93–1.26 | 1.16–1.20 | 1.21–1.38 | |||
| Selenium (Se) | 4.1 × 10-3 | ||||||||||||||||||||||
| Units | mg/ 100 g FW | mg/ 100 g FW | mg/ 100 g | mg/ 100 g FW | mg/ 100 g FW | mg/ 100 g DW | mg/ 100 g DW | mg/ 100 g FW | mg/ 100 g DW | mg/ 100 g DW | mg/ 100 g DW | mg/ 100 g DW | mg/ 100 g FW | mg/ 100 g FW | mg/ 100 g | mg/ 100 g FW | mg/ 100 g DW | mg/ 100 g FW | mg/ 100 g DW | mg/ 100 g DW | mg/ 100 g DW | mg/ 100 g DW | |
| Reference | [19] | [20] | [18] | [17] | [17] | [22] | [22] | [17] | [22] | [22] | [22] | [22] | [17] | [17] | [26] | [17] | [22] | [17] | [22] | [22] | [22] | [22] | |
| Variety | Unknown | Unknown | Butte | Carmel | Fritz | Mission | Monterey | Nonpareil | Sonora | Units | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nutrients | |||||||||||
| Thiamin | 0.205 | 0.21 | mg | ||||||||
| Riboflavin | 1.138 | 0.75 | 1.68 | 1.17 | 1.01 | 1.11 | 1 | 1.32 | 1.25 | mg | |
| Niacin | 3.618 | 2.2 | 2.71 | 2.29 | 2.52 | 3.72 | 3.35 | 3.49 | 2.73 | mg | |
| Pantothenic acid | 0.471 | mg | |||||||||
| Vitamin B6 | 0.137 | 0.15 | mg | ||||||||
| Total folate | 44 | 49 | μg | ||||||||
| Vitamin E | 25.63 | 24 | 27.6 | 29.9 | 26.3 | 28.3 | 21.9 | 26 | 31 | mg | |
| References | [19] | [20] | [17] | ||||||||
| Units | Sucrose | Raffinose | Glucose | Fructose | Maltose | Inositol | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Almond Varieties | g/100 g FW | g/100 g DW | % DW | g/100 g FW | g/100 g DW | % DW | g/100 g DW | g/100 g DW | g/100 g DW | g/100 g DW | g/100 g DW | % DW | |
| Butte | 3.1 | ||||||||||||
| Carmel | 3.4 | ||||||||||||
| Casanova | 13.9 | 1.93 | 0.96 | 0.24 | |||||||||
| Duro italiano | 13.2 | 2.11 | 1.11 | 0.27 | |||||||||
| Ferraduel | 16.3 | 0.71 | 0.95 | 0.38 | |||||||||
| Ferragnes | 22.2 | 70.6–85.3 | 0.75 | 1.23–4.84 | 1.47 | 0.37 | 2.35–7.96 | ||||||
| Ferrastar | 22.0 | 1.73 | 1.04 | 0.17 | |||||||||
| Fritz | 3 | ||||||||||||
| Gloriette | 16.9 | 0.89 | 1.3 | 0.59 | |||||||||
| Marcona | 11.5 | 1.67 | 0.77 | 0.29 | |||||||||
| Mission | 2.9 | ||||||||||||
| Monterey | 3.7 | ||||||||||||
| Nonpareil | 4.1 | ||||||||||||
| Pegarinhos (one seed) | 12.0 | 1.43 | 0.71 | 0.19 | |||||||||
| Pegarinhos (two seeds) | 15.9 | 1.29 | 0.42 | 0.11 | |||||||||
| Refego | 14.7 | 1.4 | 0.68 | 0.25 | |||||||||
| Sonora | 3.1 | ||||||||||||
| Sweet kernelled almond selections | 2.53 (1.27–3.70) | 1.88 (1.00–4.30) | 4.08 (1.42–6.50) | 0.92 (0.29–1.50) | |||||||||
| Bitter kernelled almond selections | 2.52 (0.99–4.35) | 2.24 (1.18–4.40) | 2.98 (1.60–4.46) | 0.79 (0.18–1.30) | |||||||||
| Reference | [27] | [28] | [23] | [17] | [27] | [23] | [27] | [28] | [27] | [28] | [28] | [23] | |
| Varieties | Almond | Carmel | Mission | Nonpareil | |
|---|---|---|---|---|---|
| Amino Acids | (g/100 g protein) | ||||
| Histidine | 0.539 | 1.93 | 1.93 | 2.14 | |
| Isoleucine | 0.751 | 2.88 | 2.83 | 2.71 | |
| Leucine | 1.473 | 6.19 | 5.77 | 5.81 | |
| Lysine | 0.568 | 2.14 | 1.99 | 2.36 | |
| Methionine | 0.157 | 0.4 | 0.37 | 0.42 | |
| Cystine | 0.215 | 0.26 | 0.28 | 0.24 | |
| Threonine | 0.601 | 1.95 | 1.91 | 1.93 | |
| Tryptophan | 0.211 | 1.08 | 0.89 | 0.89 | |
| Valine | 0.855 | 3.39 | 3.3 | 3.17 | |
| Phenylalanine | 1.132 | 4.38 | 4.1 | 4.25 | |
| Tyrosine | 0.45 | 1.51 | 1.39 | 1.19 | |
| Arginine | 2.465 | 9.4 | 9.68 | 9.33 | |
| Alanine | 0.999 | 4.25 | 4.12 | 4.11 | |
| Reference | [19] | [25] | [25] | [25] | |
| Reference | Trial Type | Subjects | Number of Study Participants | Almond Intervention | Duration Intervention | Main Results |
|---|---|---|---|---|---|---|
| Lipidemic control | ||||||
| [57] | R, CO | Normolipemic subjects | 22 | Replacement of 50% of usual daily intake of dietary fat with whole almonds or almond oil | 6 weeks | Significantly reduced serum levels of triglycerides, TC and LDL-c and increased HDL-c |
| [58] | R, CO, C | Subjects with mild hypercholesterolemia | 20 | 20 g almonds daily | 6 weeks | Significantly reduced serum levels of TC, LDL-c and non-HDL-c |
| [59] | R, C | Healthy adults | 85 | 56 g almonds daily | 20 weeks | Significantly reduced TC, LDL-c, non-HDL-c, triglycerides, FM and WHR |
| [60] | R, OL, C | Subjects with hyperlipidemia | 97 | 10 mL almond oil twice daily | 4 weeks | Significantly reduce serum levels of TC and LDL-c |
| Glycemic control | ||||||
| [61] | R, C | Healthy subjects | 15 | Test meal with 60 g almonds | - | Significantly reduced postprandial glycaemia, insulinemia and increased the concentration of serum protein thiol, indicating less oxidative protein damage |
| [62] | R, CO, C | Healthy subjects/T2DM patients | 13/7 | Test meal with 28 g almonds | - | Significantly reduced postprandial glycaemia in the T2DM group |
| T2DM patients | 13 | 1 serving (28 g) of almonds five days a week | 12 weeks | Significantly reduced HbA1c in the T2DM group | ||
| [63] | R, C, PG | Young healthy adults | 73 | 56.7 g almonds daily | 8 weeks | A smaller decline in HDL-c; lower 2-h glucose AUC, IRI and higher Matsuda index during the OGTT, compared to control. Reduced fasting glucose and LDL-c |
| [65] | R, CO, C | T2DM patients with mild hyperlipidemia | 20 | Diet with almonds (20% of energy intake) | 4 weeks | Significantly decreased levels of TC, LDL-c, LDL/HDL ratio, ApoB, ApoB/ApoA1 ratio, non-esterified fatty acid, fasting insulin, fasting glucose and HOMA-IR |
| Obesity | ||||||
| [70] | R, C, PG | Overweight /obese subjects (BMI: 25‒40 kg/m2) | 86 | Hypocaloric, almond-enriched diet (15% energy from almond) | 12 weeks | Greater reduction in truncal, visceral and total fat, diastolic blood pressure |
| [71] | R, CO | Obese adults | 123 | Hypocaloric, almond-enriched diet (28 g daily) | 18 months | Less bodyweight reduction and no significant changes in body composition |
| Cardiovascular risk | ||||||
| [64] | Non-C | Patients with T2DM | 50 | Diet with raw almond (20% of energy intake) | 24 weeks | Significantly improved WC, waist-to-height ratio, TC, triglycerides, LDL-c, HbA1c, hs-CRP |
| [72] | R, CO, C | Subjects with elevated LDL-c | 48 | A cholesterol-lowering diet with 1.5 oz. almond/day | 6 weeks | Significantly reduced non-HDL-c and LDL-c levels accompanied by reduced abdominal and leg fat |
| [73] | R, CO, C | Patients with CAD | 45 | Diet with 85 g almonds daily | 6 weeks | Unchanged vascular outcomes (vascular function, peripheral arterial tonometry, pulse wave velocity), serum parameters (lipid, CRP, TNFα, E-selectin) and blood pressure. Reduced VCAM1 and increased urinary NO. |
| [74] | Non-C | Healthy men mild hyperlipidemia | 30 | 60 g of almonds daily | 4 weeks | Significantly decreased LDL-c, TC and ApoB100 levels |
| [75] | R, CO, C | Hyperlipidemic subjects | 27 | Isoenergetic supplements: full-dose almonds (73 g daily), half-dose almonds+half-dose muffins | 4 weeks | Significantly reduced levels of LDL-c, LDL-c/HDL-c ratio, lipoprotein A and ox-LDL |
| [76] | R, CO, C | Hyperlipidemic subjects | 27 | Isoenergetic supplements: full-dose almonds (73 g daily), half-dose almonds+half-dose muffins | 4 weeks | Increased content of OA and MUFA in TAG and NEFA fractions, which are inversely associated with both Framingham 10-year CHD risk score and CHD lipid risk |
| [77] | R, PG | Adults with prediabetes | 65 | ADA diet containing 20% of energy from almonds | 16 weeks | Significantly reduced levels of insulin, HOMA-IR, HOMA-β, LDL-c |
| Inflammation | ||||||
| [78] | R, CO, C | Healthy adults | 25 | Low-almond diet (10% isoenergetic replacement with almond) and high-almond diet (20% isoenergetic replacement with almonds) | 4 weeks | Significantly reduced levels of E-selectin in high-almond diet and significantly reduced levels of CRP in both diets |
| Oxidative stress | ||||||
| [79] | R, C | Healthy subjects, regular smokers | 30 | 86 g and 164 g almonds daily | 4 weeks | Significantly reduced levels of 8-OH-dG, MDA and single-strand DNA breaks. No significant effects on SOD and GSH-Px. |
| [80] | R, CO, C | Healthy subjects, regular smokers | 60 | 84 g almonds daily | 4 weeks | Significantly increased levels of SOD and GSH-Px and reduced levels of 8-OH-dG, MDA and DNA strand breaks |
| Serum uric acid | ||||||
| [81] | R, C | CAD patients | 150 | 10 g almonds | 12 weeks | Significant reduced uric acid serum levels |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barreca, D.; Nabavi, S.M.; Sureda, A.; Rasekhian, M.; Raciti, R.; Silva, A.S.; Annunziata, G.; Arnone, A.; Tenore, G.C.; Süntar, İ.; et al. Almonds (Prunus Dulcis Mill. D. A. Webb): A Source of Nutrients and Health-Promoting Compounds. Nutrients 2020, 12, 672. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030672
Barreca D, Nabavi SM, Sureda A, Rasekhian M, Raciti R, Silva AS, Annunziata G, Arnone A, Tenore GC, Süntar İ, et al. Almonds (Prunus Dulcis Mill. D. A. Webb): A Source of Nutrients and Health-Promoting Compounds. Nutrients. 2020; 12(3):672. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030672
Chicago/Turabian StyleBarreca, Davide, Seyed Mohammad Nabavi, Antoni Sureda, Mahsa Rasekhian, Roberto Raciti, Ana Sanches Silva, Giuseppe Annunziata, Angela Arnone, Gian Carlo Tenore, İpek Süntar, and et al. 2020. "Almonds (Prunus Dulcis Mill. D. A. Webb): A Source of Nutrients and Health-Promoting Compounds" Nutrients 12, no. 3: 672. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030672