The Presence of Caffeic Acid in Cerebrospinal Fluid: Evidence That Dietary Polyphenols Can Cross the Blood-Brain Barrier in Humans


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Protocol and Patients
2.2. Cerebrospinal Fluid Collection
3. Matched Plasma Samples
3.1. Determination of Selected Phenolics in CSF and Plasma Samples
3.2. Solid Phase Extraction Technique
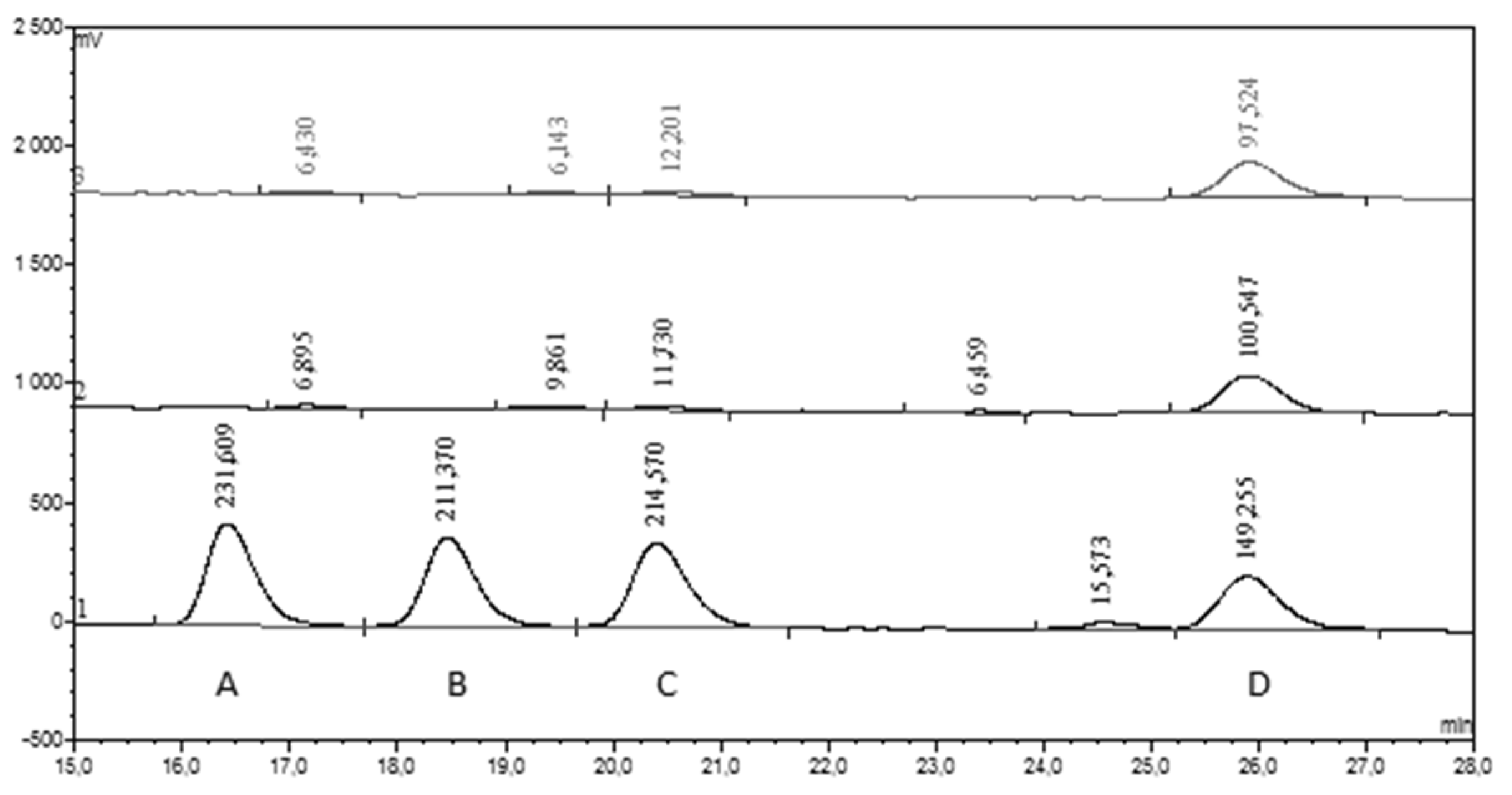
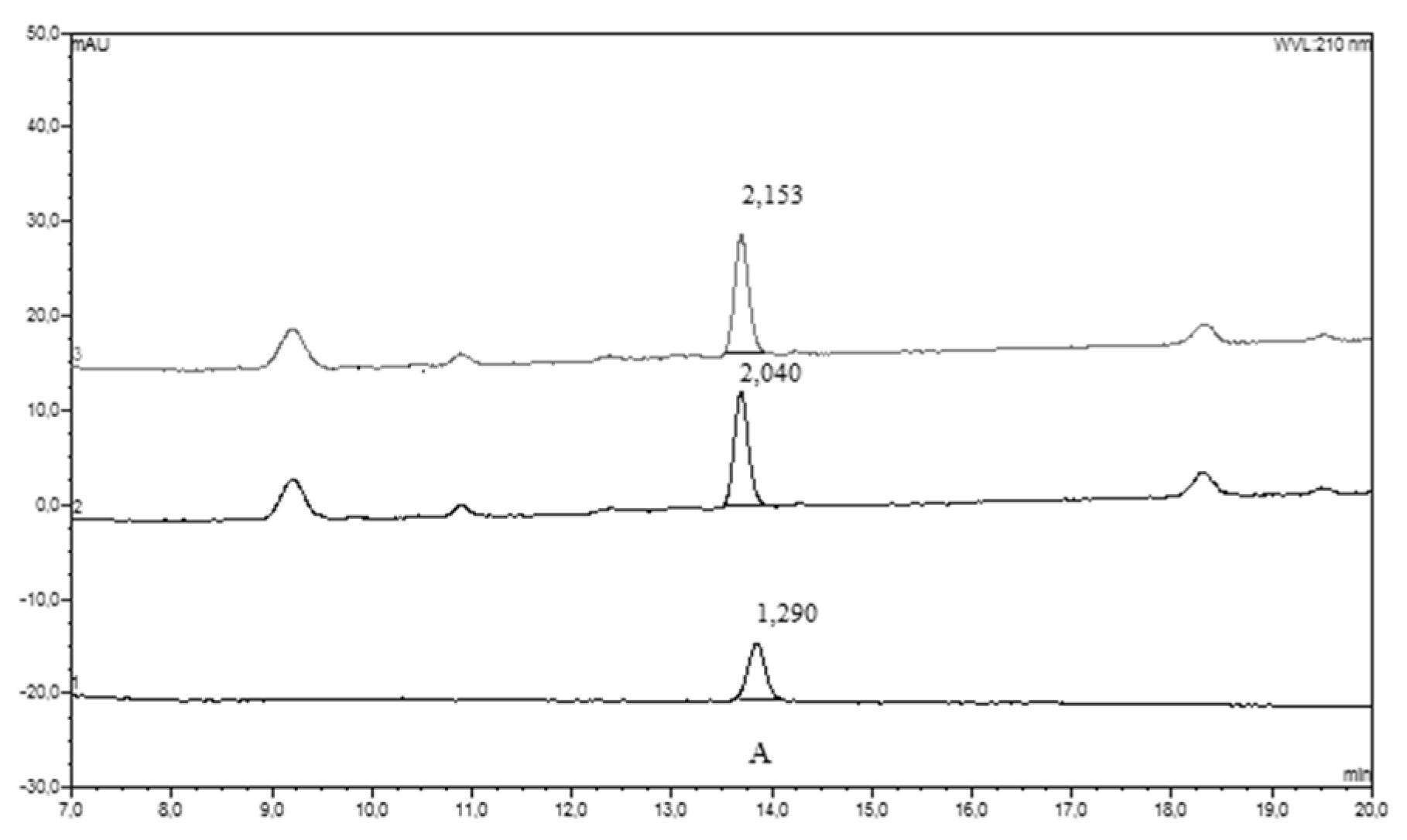
3.2.1. HPLC-UV-Vis Separation and Detection
3.2.2. HPLC-ECD Separation and Detection
3.2.3. HPLC Data Collection and Elaboration
3.3. Other Determinations
4. Statistical Analyses
5. Results
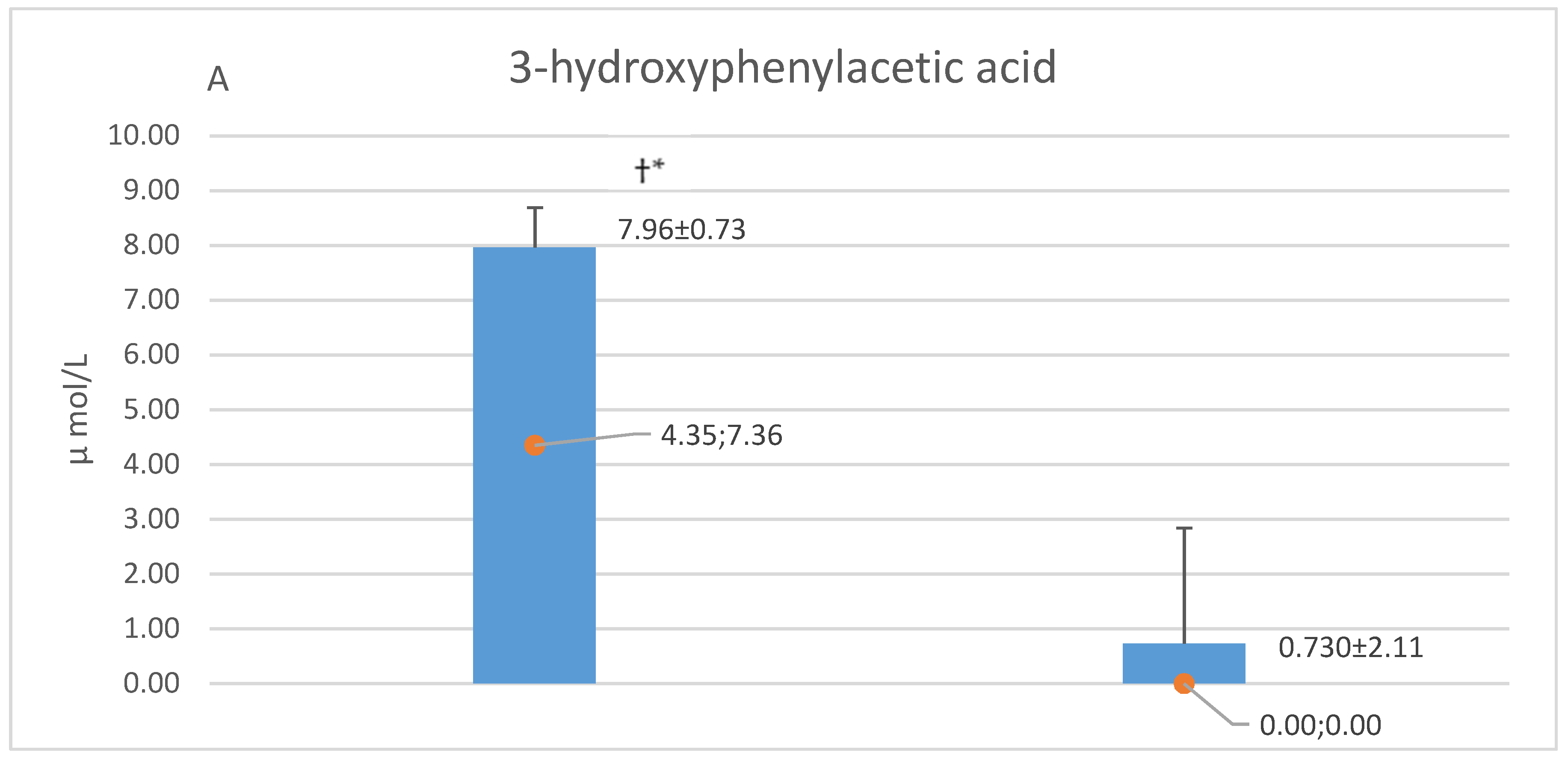
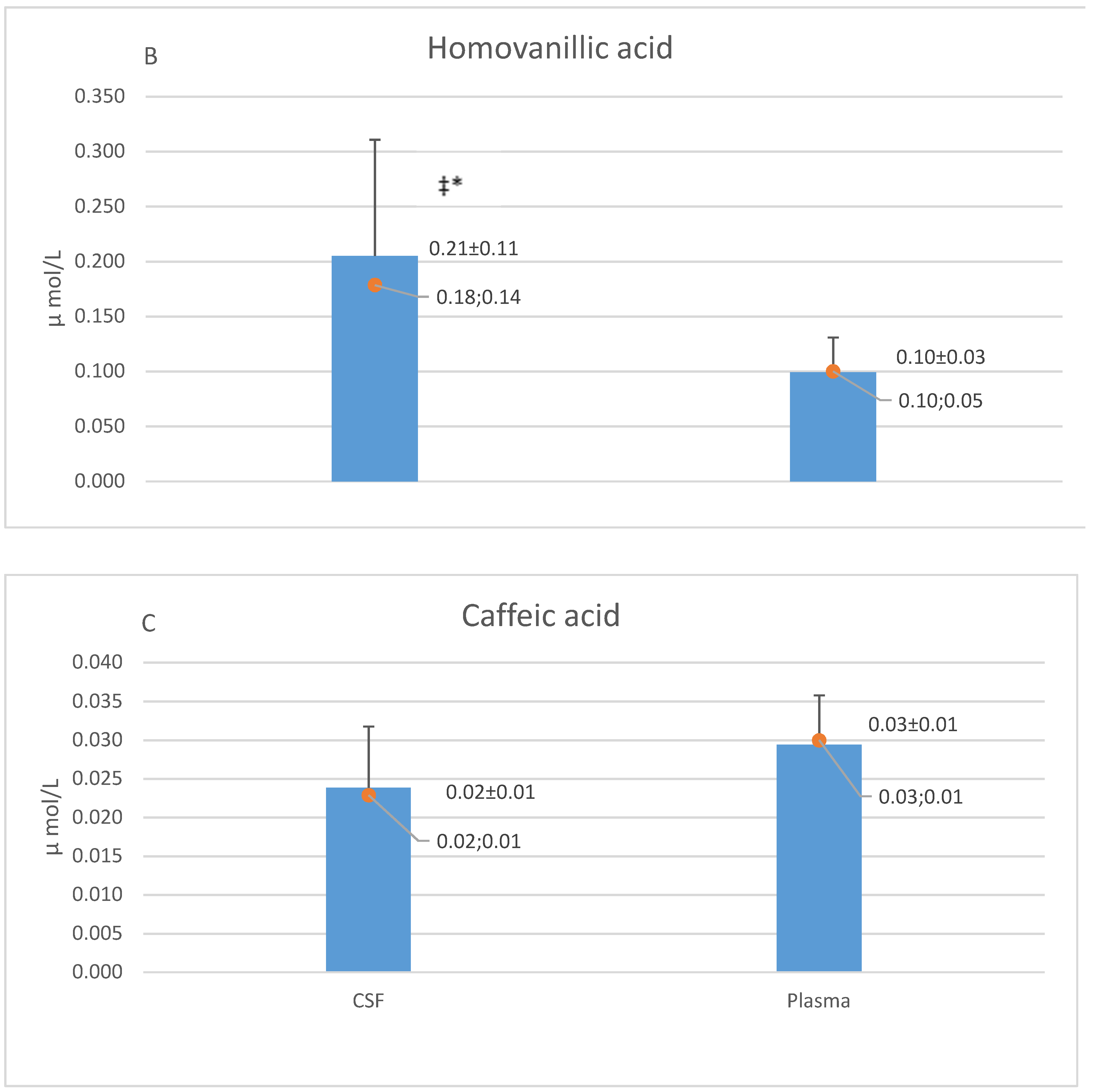
5.1. CSF and Plasma Concentrations of Caffeic Acid, Homovanillic Acid and 3-hydroxyphenyl Acetic Acid
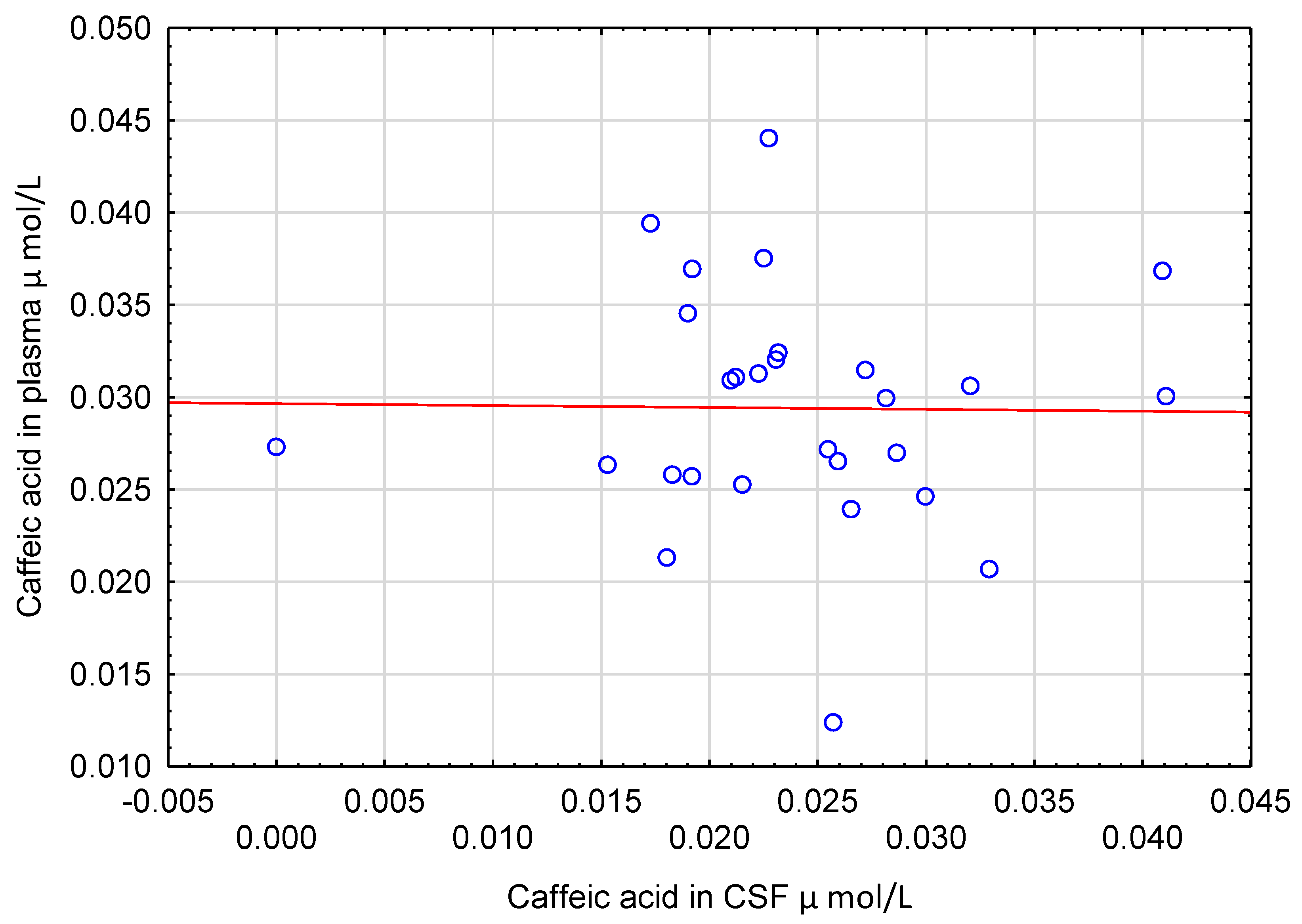
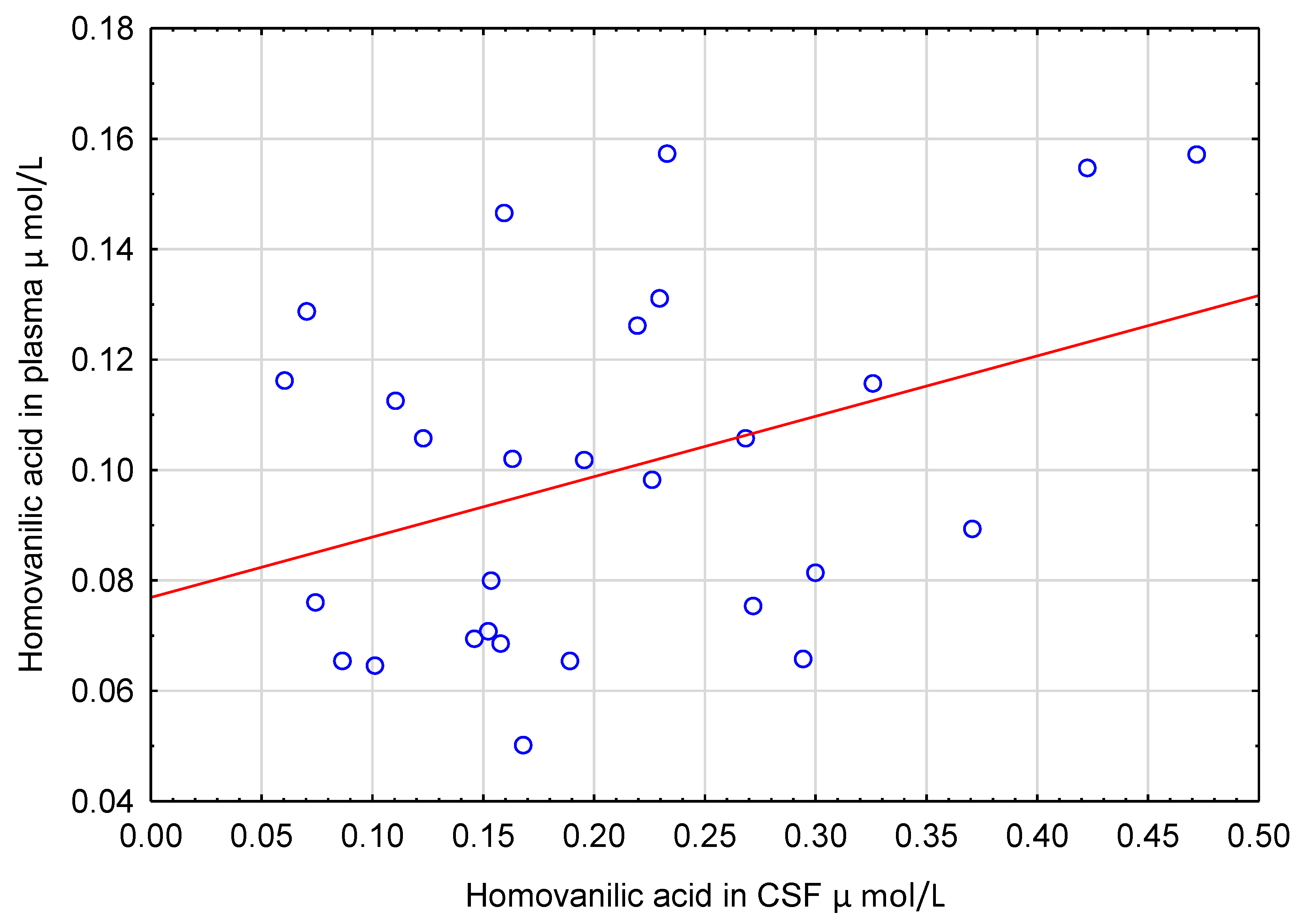
5.2. Correlations of Polyphenols in CSF with Other Measured Variables
6. Discussion
6.1. Homovanillic Acid in CSF
6.2. 3-hydroxyphenyl Acetic Acid in CSF
6.3. Caffeic Acid in CSF
7. Limitations of the Study
8. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martin, K.R.; Apple, C.L. Polyphenols as dietary supplements: A double-edged sword. Nutr. Diet. Suppl. 2009, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Nowak, D. Antioxidant Plant Polyphenols and Cognitive Disorders. In Studies on Psychiatric Disorders. Oxidative Stress in Applied Basic Research and Clinical Practice, 1st ed.; Dietrich-Muszalska, A., Chauhan, V., Grignon, S., Eds.; Humana Press: New York, NY, USA, 2015; pp. 521–552. [Google Scholar]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Nurk, E.; Refsum, H.; Drevon, C.A.; Tell, G.S.; Nygaard, H.A.; Engedal, K.; Smith, A.D. Intake of flavonoid-rich wine, tea, and chocolate by elderly men and women is associated with better cognitive test performance. J. Nutr. 2009, 139, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Letenneur, L.; Proust-Lima, C.; Le Gouge, A.; Dartigues, J.F.; Barberger-Gateau, P. Flavonoid intake and cognitive decline over a 10-year period. Am. J. Epidemiol. 2007, 165, 1364–1371. [Google Scholar] [CrossRef] [PubMed]
- Summers, W.K.; Martin, R.L.; Cunningham, M.; DeBoynton, V.L.; Marsh, G.M. Complex antioxidant blend improves memory in community-dwelling seniors. J. Alzheimers Dis. 2010, 19, 429–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, J.; Croft, K.; Mori, T.; Wesnes, K.; Spong, J.; Downey, L.; Kure, C.; Lloyd, J.; Stough, C. An examination of the effects of the antioxidant Pycnogenol on cognitive performance, serum lipid profile, endocrinological and oxidative stress biomarkers in an elderly population. J. Psychopharmacol. 2008, 22, 553–562. [Google Scholar] [CrossRef]
- Vauzour, D. Dietary polyphenols as modulators of brain functions: Biological actions and molecular mechanisms underpinning their beneficial effects. Oxid. Med. Cell Longev. 2012, 2012, 914273. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, B.; Sorokin, L. The blood-brain and the blood-cerebrospinal fluid barriers: Function and dysfunction. Semin. Immunopathol. 2009, 31, 497–511. [Google Scholar] [CrossRef] [Green Version]
- Figueira, I.; Menezes, R.; Macedo, D.; Costa, I.; Dos Santos, C.N. Polyphenols Beyond Barriers: A Glimpse into the Brain. Curr. Neuropharmacol. 2017, 15, 562–594. [Google Scholar] [CrossRef] [Green Version]
- Andres-Lacueva, C.; Shukitt-Hale, B.; Galli, R.L.; Jauregui, O.; Lamuela-Raventos, R.M.; Joseph, J.A. Anthocyanins in aged blueberry-fed rats are found centrally and may enhance memory. Nutr. Neurosci. 2005, 8, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Talavéra, S.; Felgines, C.; Texier, O.; Besson, C.; Gil-Izquierdo, A.; Lamaison, J.L.; Rémésy, C. Anthocyanin metabolism in rats and their distribution to digestive area, kidney, and brain. J. Agric. Food Chem. 2005, 53, 3902–3908. [Google Scholar] [CrossRef]
- Peng, H.W.; Cheng, F.C.; Huang, Y.T.; Chen, C.F.; Tsai, T.H. Determination of naringenin and its glucuronide conjugate in rat plasma and brain tissue by high-performance liquid chromatography. J. Chromatogr. B 1998, 714, 369–374. [Google Scholar] [CrossRef]
- Suganuma, M.; Okabe, S.; Oniyama, M.; Tada, Y.; Ito, H.; Fujiki, H. Wide distribution of (3H)(−)-epigallocatechin gallate, a cancer preventive tea polyphenol, in mouse tissue. Carcinogenesis 1998, 19, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Abd El Mohsen, M.M.; Kuhnle, G.; Rechner, A.R.; Schroeter, H.; Rose, S.; Jenner, P.; Rice-Evans, C.A. Uptake and metabolism of epicatechin and its access to the brain after oral ingestion. Free Radic. Biol. Med. 2002, 33, 1693–1702. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y.; Zhao, Z.; Xue, Y. Effects of green tea polyphenols on caveolin-1 of microvessel fragments in rats with cerebral ischemia. Neurol. Res. 2010, 32, 963–970. [Google Scholar] [CrossRef]
- Zafra-Gómez, A.; Luzón-Toro, B.; Jiménez-Diaz, I.; Ballesteros, O.; Navalón, A. Quantification of phenolic antioxidants in rat cerebrospinal fluid by GC-MS after oral administration of compounds. J. Pharm. Biomed. Anal. 2010, 53, 103–108. [Google Scholar] [CrossRef]
- Lü, K.; Ding, M.Y.; Li, H.X.; Liu, D.L. Determination of ferulic acid in Chuanxiong and in animal serum and cerebrospinal fluid by reversed-phase high performance liquid chromatography. Chin. J. Chromatogr. 2000, 18, 518–520. [Google Scholar]
- Ebinger, G.; Verheyden, R. On the occurrence of vanillic acid in human-brain and cerebrospinal-fluid. J. Neurol. 1976, 212, 133–138. [Google Scholar] [CrossRef]
- Watson, E.; Wilk, E.; Roboz, J. Derivatization and gas-chromatographic determination of some biologically important acids in cerebrospinal-fluid. Anal. Biochem. 1974, 59, 441–451. [Google Scholar] [CrossRef]
- Vogt, W.; Jacob, K.; Ohnesorge, A.B.; Schwertfeger, G. Highly sensitive method for the quantitation of homovanillic-acid in cerebrospinal-fluid. J. Chromatogr. 1980, 199, 191–197. [Google Scholar] [CrossRef]
- Pullar, I.A.; Weddell, J.M.; Ahmed, R.; Gillingham, F.J. Phenolic acid concentrations in lumbar cerebrospinal fluid of parkinsonian patients treated with L-Dopa. J. Neurol. Neurosurg. Psychiatry 1970, 33, 851–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jong, A.P.J.M.; Kok, R.M.; Cramers, C.A.; Wadman, S.K. Determination of acidic catecholamine metabolites in plasma and cerebrospinal fluid using gas chromatography negative ion mass spectrometry. J. Chromatogr. B 1986, 382, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Faria, A.; Pestana, D.; Teixeira, D.; Couraud, P.O.; Romero, I.; Weksler, B.; de Freitas, V.; Mateus, N.; Calhau, C. Insights into the putative catechin and epicatechin transport across blood-brain barrier. Food Funct. 2011, 2, 9–44. [Google Scholar] [CrossRef] [PubMed]
- Figueira, I.; Tavares, L.; Jardim, C.; Costa, I.; Terrasso, A.P.; Almeida, A.F.; Govers, C.; Mes, J.J.; Gardner, R.; Becker, J.D.; et al. Blood-brain barrier transport and neuroprotective potential of blackberry-digested polyphenols: An in vitro study. Eur. J. Nutr. 2019, 58, 113–130. [Google Scholar] [CrossRef]
- Teunissen, C.E.; Petzold, A.; Bennett, J.L.; Berven, F.S.; Brundin, L.; Comabella, M.; Franciotta, D.; Frederiksen, J.L.; Fleming, J.O.; Furlan, R.; et al. A consensus protocol for the standardization of cerebrospinal fluid collection and biobanking. Neurology 2009, 73, 1914–1922. [Google Scholar] [CrossRef] [Green Version]
- Prymont-Przyminska, A.; Zwolinska, A.; Sarniak, A.; Wlodarczyk, A.; Krol, M.; Nowak, M.; de Graft-Johnson, J.; Padula, G.; Bialasiewicz, P.; Markowski, J.; et al. Consumption of strawberries on a daily basis increases the non-urate 2,2-diphenyl-1-picryl-hydrazyl (DPPH) radical scavenging activity of fasting plasma in healthy subjects. J. Clin. Biochem. Nutr. 2014, 55, 48–55. [Google Scholar] [CrossRef]
- Bialasiewicz, P.; Prymont-Przyminska, A.; Zwolinska, A.; Sarniak, A.; Wlodarczyk, A.; Krol, M.; Glusac, J.; Nowak, P.; Markowski, J.; Rutkowski, K.P.; et al. Addition of strawberries to the usual diet decreases resting chemiluminescence of fasting blood in healthy subjects-possible health-promoting effect of these fruits consumption. J. Am. Coll. Nutr. 2014, 33, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Aura, A.M. Microbial metabolism of dietary phenolic compounds in the colon. Phytochem. Rev. 2008, 7, 407–429. [Google Scholar] [CrossRef]
- Eisenhofer, G.; Kopin, I.J.; Goldstein, D.S. Catecholamine metabolism: A contemporary view with implications for physiology and medicine. Pharmacol. Rev. 2004, 56, 331–349. [Google Scholar] [CrossRef]
- Peaston, R.T.; Weinkove, C. Measurement of catecholamines and their metabolites. Ann. Clin. Biochem. 2004, 41, 17–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Koide, Y.; Shohmori, T. Determination of p-hydroxyphenylacetic acid in cerebrospinal fluid by high-performance liquid chromatography with electrochemical detection. Clin. Chim. Acta 1982, 123, 161–168. [Google Scholar] [CrossRef]
- Phenol-Explorer Database. Available online: http://www.phenol-explorer.eu (accessed on 27 March 2020).
- The Human Metabolome Database. Available online: https://hmdb.ca (accessed on 27 March 2020).
- Lambert, G.W.; Eisenhofer, G.; Cox, H.S.; Horne, M.; Kalff, V.; Kelly, M.; Jennings, G.L.; Esler, M.D. Direct determination of homovanillic acid release from the human brain, an indicator of central dopaminergic activity. Life Sci. 1991, 49, 1061–1072. [Google Scholar] [CrossRef]
- Combet, E.; Lean, M.E.; Boyle, J.G.; Crozier, A.; Davidson, D.F. Dietary flavonols contribute to false-positive elevation of homovanillic acid, a marker of catecholamine-secreting tumors. Clin. Chim. Acta 2011, 412, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Weldin, J.; Jack, R.; Dugaw, K.; Kapur, R.P. Quercetin, an over-the-counter supplement, causes neuroblastoma-like elevation of plasma homovanillic acid. Pediatr. Dev. Pathol. 2003, 6, 547–551. [Google Scholar] [CrossRef]
- Jaganath, I.B.; Mullen, W.; Lean, M.E.; Edwards, C.A.; Crozier, A. In vitro catabolism of rutin by human fecal bacteria and the antioxidant capacity of its catabolites. Free Radic. Biol. Med. 2009, 47, 1180–1189. [Google Scholar] [CrossRef]
- Mansoorian, B.; Combet, E.; Alkhaldy, A.; Garcia, A.L.; Edwards, C.A. Impact of Fermentable Fibres on the Colonic Microbiota Metabolism of Dietary Polyphenols Rutin and Quercetin. Int. J. Environ. Res. Public Health 2019, 16, 292. [Google Scholar] [CrossRef] [Green Version]
- Vetrani, C.; Rivellese, A.A.; Annuzzi, G.; Adiels, M.; Borén, J.; Mattila, I.; Orešič, M.; Aura, A.M. Metabolic transformations of dietary polyphenols: Comparison between in vitro colonic and hepatic models and in vivo urinary metabolites. J. Nutr. Biochem. 2016, 33, 111–118. [Google Scholar] [CrossRef]
- Urpi-Sarda, M.; Monagas, M.; Khan, N.; Lamuela-Raventos, R.M.; Santos-Buelga, C.; Sacanella, E.; Castell, M.; Permanyer, J.; Andres-Lacueva, C. Epicatechin, procyanidins, and phenolic microbial metabolites after cocoa intake in humans and rats. Anal. Bioanal. Chem. 2009, 394, 1545–1556. [Google Scholar] [CrossRef] [PubMed]
- Rios, L.Y.; Gonthier, M.P.; Rémésy, C.; Mila, I.; Lapierre, C.; Lazarus, S.A.; Williamson, G.; Scalbert, A. Chocolate intake increases urinary excretion of polyphenol-derived phenolic acids in healthy human subjects. Am. J. Clin. Nutr. 2003, 77, 912–918. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Koide, Y.; Yoshino, K.; Shohmori, T. P-hydroxyphenylacetic acid concentrations in cerebrospinal fluid. No Shinkei Brain Nerve 1982, 34, 769–774. [Google Scholar] [PubMed]
- Berry, M.D. Mammalian central nervous system trace amines. Pharmacologic amphetamines, physiologic neuromodulators. J. Neurochem. 2004, 90, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Andersen, G.; Marcinek, P.; Sulzinger, N.; Schieberle, P.; Krautwurst, D. Food sources and biomolecular targets of tyramine. Nutr. Rev. 2019, 77, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Könczöl, Á.; Rendes, K.; Dékány, M.; Müller, J.; Riethmüller, E.; Balogh, G.T. Blood-brain barrier specific permeability assay reveals N-methylated tyramine derivatives in standardized leaf extracts and herbal products of Ginkgo biloba. J. Pharm. Biomed. Anal. 2016, 131, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Constantinou, M.A.; Papakonstantinou, E.; Spraul, M.; Sevastiadou, S.; Costalos, C.; Koupparis, M.A.; Shulpis, K.; Tsantili-Kakoulidou, A.; Mikros, E. 1H NMR-based metabonomics for the diagnosis of inborn errors of metabolism in urine. Anal. Chim. Acta 2005, 542, 169–177. [Google Scholar] [CrossRef]
- Xiong, X.; Liu, D.; Wang, Y.; Zeng, T.; Peng, Y. Urinary 3-(3-Hydroxyphenyl)-3-hydroxypropionic Acid, 3-Hydroxyphenylacetic Acid, and 3-Hydroxyhippuric Acid Are Elevated in Children with Autism Spectrum Disorders. Biomed Res. Int. 2016, 2016, 9485412. [Google Scholar] [CrossRef] [Green Version]
- Patabendige, A.; Youdim, K.; Abbott, J. Ability of phenolic compounds to protect the blood-brain barrier against oxidative stress. Conference: The Physiological Society at King’s College London. J. Physiol. 2005, 565P, PC157. [Google Scholar]
- Espíndola, K.M.M.; Ferreira, R.G.; Narvaez, L.E.M.; Silva Rosario, A.C.R.; da Silva, A.H.M.; Silva, A.G.B.; Vieira, A.P.O.; Monteiro, M.C. Chemical and Pharmacological Aspects of Caffeic Acid and Its Activity in Hepatocarcinoma. Front. Oncol. 2019, 9, 541. [Google Scholar] [CrossRef] [Green Version]
- Nałęcz, K.A. Solute Carriers in the Blood-Brain Barier: Safety in Abundance. Neurochem. Res. 2017, 42, 795–809. [Google Scholar] [CrossRef]
- Suominen, T.; Piepponen, T.P.; Kostiainen, R. Permeation of Dopamine Sulfate through the Blood-Brain Barrier. PLoS ONE 2015, 10, e0133904. [Google Scholar] [CrossRef]
- Faria, A.; Mateus, N.; Calhau, C. Flavonoid transport across blood-brain barrier: Implication for their direct neuroprotective actions. Nutr. Aging 2012, 1, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Moosavi, F.; Hosseini, R.; Saso, L.; Firuzi, O. Modulation of neurotrophic signaling pathways by polyphenols. Drug Des. Devel. Ther. 2015, 10, 23–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szwajgier, D.; Borowiec, K.; Pustelniak, K. The Neuroprotective Effects of Phenolic Acids: Molecular Mechanism of Action. Nutrients 2017, 9, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro Fernandes, F.D.; Fontenele Menezes, A.P.; de Sousa Neves, J.C.; Fonteles, A.A.; da Silva, A.T.; de Araújo, R.P.; Santos do Carmo, M.R.; de Souza, C.M.; de Andrade, G.M. Caffeic acid protects mice from memory deficits induced by focal cerebral ischemia. Behav. Pharmacol. 2014, 25, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Małkiewicz, M.A.; Szarmach, A.; Sabisz, A.; Cubała, W.J.; Szurowska, E.; Winklewski, P.J. Blood-brain barrier permeability and physical exercise. J. Neuroinflamm. 2019, 16, 15. [Google Scholar] [CrossRef]
- Atluri, V.S.; Hidalgo, M.; Samikkannu, T.; Kurapati, K.R.; Jayant, R.D.; Sagar, V.; Nair, M.P. Effect of human immunodeficiency virus on blood-brain barrier integrity and function: An update. Front. Cell Neurosci. 2015, 9, 212. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81 (Suppl. 1), 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Olthof, M.R.; Hollman, P.C.; Katan, M.B. Chlorogenic acid and caffeic acid are absorbed in humans. J. Nutr. 2001, 131, 66–71. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic/Clinical Variables | Whole Group † | Multiple Sclerosis | Other Neurological Disorders ‡ |
|---|---|---|---|
| Number of subjects | 28 | 15 | 13 |
| Sex F/M | 18/10 | 13/2 | 5/8 |
| Age [years] | 46 ± 16 (44; 29) | 40 ± 14 (23; 4) | 53 ± 15 (30; 9) * |
| plasma CRP [mg/L] | 4.1 ± 6.1 (1.5; 5.2) | 1.7 ± 1.7 (0.2; 0.2) | 7.5 ± 8.2 (0.5; 1.0) * |
| Cerebrospinal fluid samples | |||
| Cell count (cells/µL) | 22 ± 41 (6; 9) | 10 ± 11 (2; 1) | 31 ± 52 (1; 0) |
| Erythrocytes (cells/µL) | 118 ± 127 (3; 74) | 123 ± 199 (4; 82) | 39 ± 59 (1; 35) |
| Total protein (g/L) | 0.56 ± 0.36 (0.45; 0.19) | 0.44 ± 0.12 (0.29; 0.04) | 0.65 ± 0.38 (0.30; 0.12) |
| Glucose (mg/dL) | 74 ± 22 (68; 13) | 67 ± 6 (69; 7) | 81 ± 29 (68; 34) |
| Phenolic Acid | Whole Group n = 28 | |||
|---|---|---|---|---|
| Cerebrospinal Fluid Samples | Plasma Samples | |||
| Positive | Negative | Positive | Negative | |
| Homovanillic acid | 28 | 0 | 28 | 0 |
| Caffeic acid | 27 | 1 | 28 | 0 |
| 3-hydroxyphenyl acetic acid | 27 | 1 | 5 | 23 |
| Dihydrocaffeic acid | 0 | 28 | 17 | 11 |
| Vanillic acid | 0 | 28 | 10 | 18 |
| Hippuric acid | 0 | 28 | 17 | 11 |
| 3,4 dihydroxybenzoic acid | 0 | 28 | 1 | 27 |
| Phenolic Acid (µmol/L) | Multiple Sclerosis | Other Neurological Disorders |
|---|---|---|
| 3HPAA | 8.12 ± 10.08 (4.50; 5.39) | 7.78 ± 7.18 (4.20; 8.32) |
| HVA | 0.19 ± 0.13 (0.16; 0.15) | 0.22 ± 0.13 (0.19; 0.17) |
| CA | 0.02 ± 0.01 (0.03; 0.01) | 0.02 ± 0.01 (0.02; 0.00) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grabska-Kobylecka, I.; Kaczmarek-Bak, J.; Figlus, M.; Prymont-Przyminska, A.; Zwolinska, A.; Sarniak, A.; Wlodarczyk, A.; Glabinski, A.; Nowak, D. The Presence of Caffeic Acid in Cerebrospinal Fluid: Evidence That Dietary Polyphenols Can Cross the Blood-Brain Barrier in Humans. Nutrients 2020, 12, 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051531
Grabska-Kobylecka I, Kaczmarek-Bak J, Figlus M, Prymont-Przyminska A, Zwolinska A, Sarniak A, Wlodarczyk A, Glabinski A, Nowak D. The Presence of Caffeic Acid in Cerebrospinal Fluid: Evidence That Dietary Polyphenols Can Cross the Blood-Brain Barrier in Humans. Nutrients. 2020; 12(5):1531. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051531
Chicago/Turabian StyleGrabska-Kobylecka, Izabela, Justyna Kaczmarek-Bak, Malgorzata Figlus, Anna Prymont-Przyminska, Anna Zwolinska, Agata Sarniak, Anna Wlodarczyk, Andrzej Glabinski, and Dariusz Nowak. 2020. "The Presence of Caffeic Acid in Cerebrospinal Fluid: Evidence That Dietary Polyphenols Can Cross the Blood-Brain Barrier in Humans" Nutrients 12, no. 5: 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051531