Modulation of the Systemic Immune Response in Suckling Rats by Breast Milk TGF-β2, EGF and FGF21 Supplementation

 , , ,
, , ,  , , and
, , and 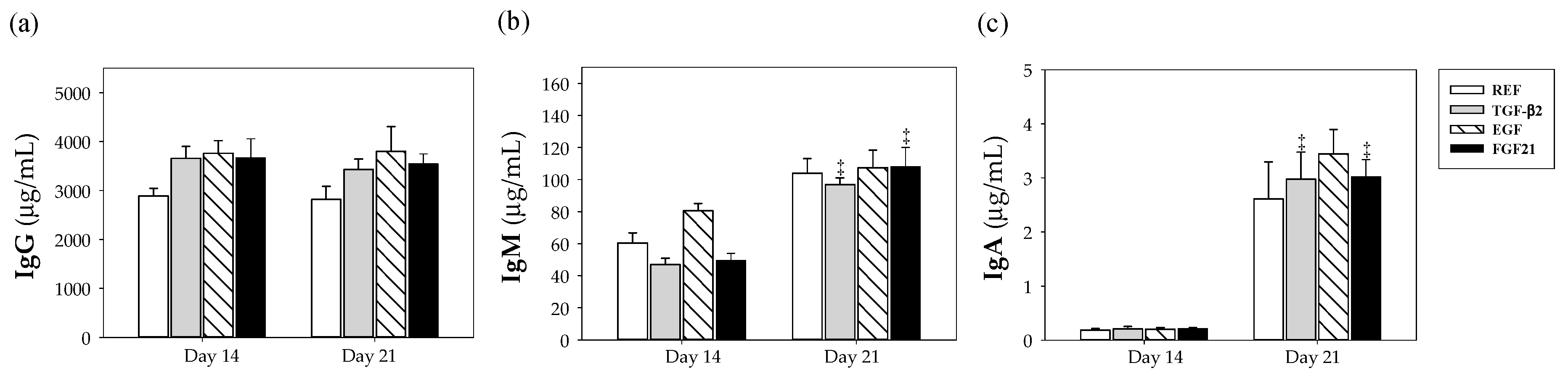{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Dietary Supplementation
2.4. Sample Collection and Processing
2.5. Immunoglobulin Quantification
2.6. Lymphocyte Isolation from Spleen
2.7. Spleen Cells Stimulation and Proliferation Assay
2.8. Quantification of Cytokine Secretion
2.9. Immunofluorescence Staining and Flow Cytometry Analysis
2.10. Statistical Analysis
3. Results
3.1. Animal and Organ Weights
3.2. Plasmatic IgG, IgM and IgA Concentrations
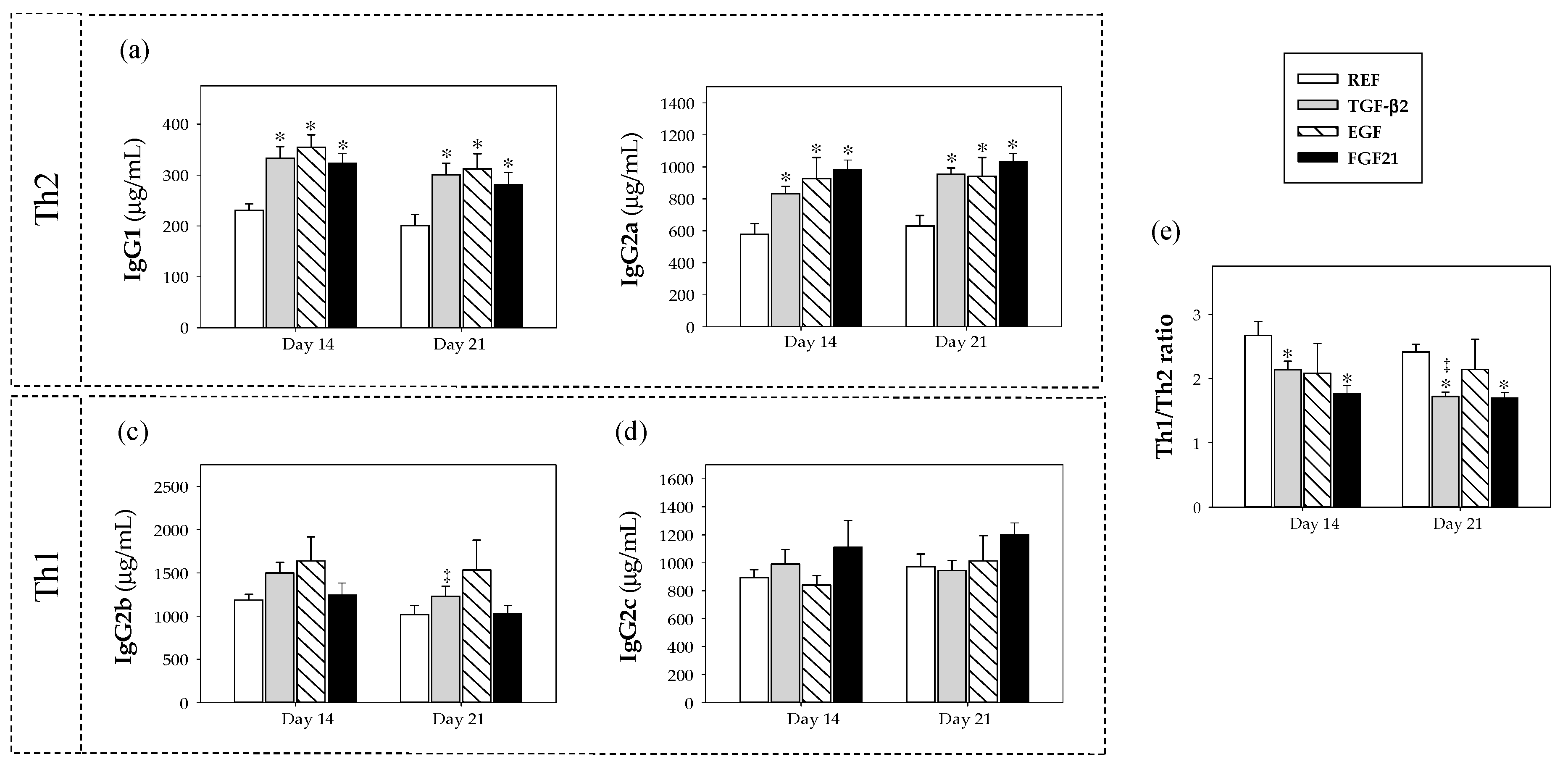
3.3. Plasmatic IgG Isotypes Concentration
3.4. Proliferation and Cytokine Production by Spleen Lymphocytes
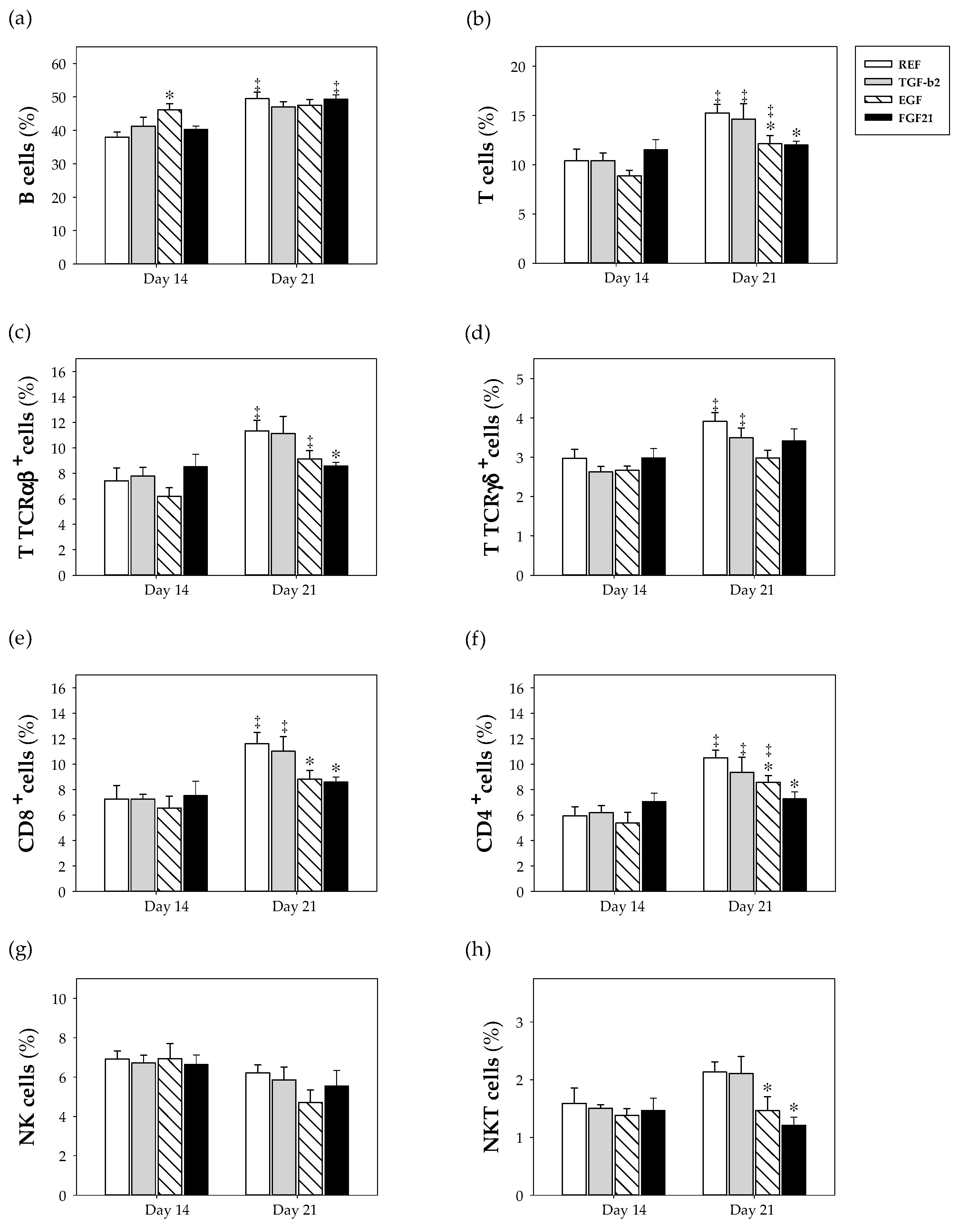
3.5. Spleen Lymphocyte Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO Recommendations on Postnatal Care of the Mother and Newborn; WQ 500; WHO: Geneva, Switzerland, 2013; p. 22. [Google Scholar]
- Cacho, N.T.; Lawrence, R.M. Innate immunity and breast milk. Front. Immunol. 2017, 8, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Santaella, M.; Mira-Pascual, L.; Martínez-Arias, E.; Khodayar-Pardo, P.; Ros, G.; Martínez-Costa, C. Longitudinal study of cytokine expression, lipid profile and neuronal growth factors in human breast milk from term and preterm deliveries. Nutrients 2015, 7, 8577–8591. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.D.; Richard, C.; Larsen, B.M.; Field, C.J. The importance of human milk for immunity in preterm infants. Clin. Perinatol. 2017, 44, 23–47. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, A. Breast milk as the gold standard for protective nutrients. J. Pediatr. 2010, 156, S3–S7. [Google Scholar] [CrossRef] [PubMed]
- le Doare, K.; Holder, B.; Bassett, A.; Pannaraj, P.S. Mother’s milk: A purposeful contribution to the development of the infant microbiota and immunity. Front. Immunol. 2018, 9, 361. [Google Scholar] [CrossRef] [Green Version]
- Field, C.J. The immunological components of human milk and their effect on immune development in infants. J. Nutr. 2005, 135, 1–4. [Google Scholar] [CrossRef]
- Järvinen, K.M.; Martin, H.; Oyoshi, M.K. Immunomodulatory effects of breast milk on food allergy. Ann. Allergy Asthma Immunol. 2019, 123, 133–143. [Google Scholar] [CrossRef]
- Victora, C.G.; Bahl, R.; Barros, A.J.; França, G.V.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C.; et al. Breastfeeding in the 21st century: Epidemiology, mechanisms, and lifelong effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Berdi, M.; de Lauzon-Guillain, B.; Forhan, A.; Castelli, F.A.; Fenaille, F.; Charles, M.A.; Heude, B.; Junot, C.; Adel-Patient, K.; EDEN Mother-Child Cohort Study Group. Immune components of early breastmilk: Association with maternal factors and with reported food allergy in childhood. Pediatr. Allergy Immunol. 2019, 30, 107–116. [Google Scholar] [CrossRef]
- Turfkruyer, M.; Verhasselt, V. Breast milk and its impact on maturation of the neonatal immune system. Curr. Opin. Infect. Dis. 2015, 28, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Penttila, I.A.; van Spriel, A.B.; Zhang, M.F.; Xian, C.J.; Steeb, C.B.; Cummins, A.G.; Zola, H.; Read, L.C. Transforming growth factor-beta levels in maternal milk and expression in postnatal rat duodenum and ileum. Pediatr. Res. 1998, 44, 524–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, A.P.; Braun, A.; Ritchie, S.C.; Carbone, F.R.; Mackay, L.K.; Gebhardt, T.; Inouye, M.Y. Comparative analysis reveals a role for TGF-β in shaping the residency-related transcriptional signature in tissue-resident memory CD8+ T cells. PLoS ONE 2019, 14, e0210495. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Yoshida, M.; Ichijo, M.; Ishizaka, S.; Tsujh, T. Transforming growth factor-beta (TGF-β) in human milk. Clin. Exp. Immunol. 1993, 94, 220–224. [Google Scholar] [CrossRef] [PubMed]
- David, C.J.; Massagué, J. Contextual determinants of TGFβ action in development, immunity and cancer. Nat. Rev. Mol. Cell Biol. 2018, 19, 419–435. [Google Scholar] [CrossRef]
- Castellote, C.; Casillas, R.; Ramírez-Santana, C.; Pérez-Cano, F.J.; Castell, M.; Moretones, M.G.; López-Sabater, M.C.; Franch, A. Premature delivery influences the immunological composition of colostrum and transitional and mature human milk. J. Nutr. 2011, 141, 1181–1187. [Google Scholar] [CrossRef]
- Kalliomäki, M.; Ouwehand, A.; Arvilommi, H.; Kero, P.; Isolauri, E. Transforming growth factor-beta in breast milk: A potential regulator of atopic disease at an early age. J. Allergy Clin. Immunol. 1999, 104, 1251–1257. [Google Scholar] [CrossRef]
- Ogawa, J.; Sasahara, A.; Yoshida, T.; Sira, M.M.; Futatani, T.; Kanegane, H.; Miyawaki, T.Y. Role of transforming growth factor-β in breast milk for initiation of IgA production in newborn infants. Early Hum. Dev. 2004, 77, 67–75. [Google Scholar] [CrossRef]
- Ando, T.; Hatsushika, K.; Wako, M.; Ohba, T.; Koyama, K.; Ohnuma, Y.; Katoh, R.; Ogawa, H.; Okumura, K.; Luo, J.; et al. Orally administered TGF-β is biologically active in the intestinal mucosa and enhances oral tolerance. J. Allergy Clin. Immunol. 2007, 120, 916–923. [Google Scholar] [CrossRef]
- Dvorak, B.; Halpern, M.D.; Holubec, H.; Williams, C.S.; McWilliam, D.L.; Dominguez, J.A.; Stepankova, R.; Payne, C.M.; McCuskey, R.S. Epidermal growth factor reduces the development of necrotizing enterocolitis in a neonatal rat model. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G156–G164. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Liu, H.; Yang, S.; Li, Z.; Zhong, J.; Fang, R. Epidermal growth factor and intestinal barrier function. Mediat. Inflamm. 2016, 2016, 1927348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavaldà-Navarro, A.; Hondares, E.; Giralt, M.; Mampel, T.; Iglesias, R.; Villarroya, F. Fibroblast growth factor 21 in breast milk controls neonatal intestine function. Sci. Rep. 2015, 5, 13717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hondares, E.; Rosell, M.; Gonzalez, F.J.; Giralt, M.; Iglesias, R.; Villarroya, F. Hepatic FGF21 expression is induced at birth via PPARalpha in response to milk intake and contributes to thermogenic activation of neonatal brown fat. Cell Metab. 2010, 11, 206–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Castro, P.; Abril-Gil, M.; Rodríguez-Lagunas, M.J.; Castell, M.; Pérez-Cano, F.J.; Franch, À. TGF-β2, EGF, and FGF21 growth factors present in breast milk promote mesenteric lymph node lymphocytes maturation in suckling rats. Nutrients 2018, 10, 1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 purified diets for laboratory rodents: Final report of the American institute of nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Grases-Pintó, B.; Abril-Gil, M.; Rodríguez-Lagunas, M.J.; Castell, M.; Pérez-Cano, F.J.; Franch, À. Leptin and adiponectin supplementation modifies mesenteric lymph node lymphocyte composition and functionality in suckling rats. Br. J. Nutr. 2018, 119, 486–495. [Google Scholar] [CrossRef] [Green Version]
- Rigo-Adrover, M.D.M.; van Limpt, K.; Knipping, K.; Garssen, J.; Knol, J.; Costabile, A.; Franch, À.; Castell, M.; Pérez-Cano, F.J. Preventive effect of a synbiotic combination of galacto- and fructooligosaccharides mixture with Bifidobacterium breve M-16V in a model of multiple rotavirus infections. Front. Immunol. 2018, 9, 1318. [Google Scholar] [CrossRef] [Green Version]
- Maynard, A.A.; Dvorak, K.; Khailova, L.; Dobrenen, H.; Arganbright, K.M.; Halpern, M.D.; Kurundkar, A.R.; Maheshwari, A.; Dvorak, B. Epidermal growth factor reduces autophagy in intestinal epithelium and in the rat model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G614–G622. [Google Scholar] [CrossRef] [Green Version]
- Penttila, I.A.; Flesch, I.E.; McCue, A.L.; Powell, B.C.; Zhou, F.H.; Read, L.C.; Zola, H. Maternal milk regulation of cell infiltration and interleukin 18 in the intestine of suckling rat pups. Gut 2003, 52, 1579–1586. [Google Scholar] [CrossRef] [Green Version]
- Azagra-Boronat, I.; Massot-Cladera, M.; Knipping, K.; van’t Land, B.; Stahl, B.; Garssen, J.; Rodríguez-Lagunas, M.J.; Franch, À.; Castell, M.; Pérez-Cano, F.J. Supplementation with 2’-FL and scGOS/lcFOS ameliorates rotavirus-induced diarrhea in suckling rats. Front. Cell Infect. Microbiol. 2018, 8, 372. [Google Scholar] [CrossRef]
- Ramiro-Puig, E.; Pérez-Cano, F.J.; Ramírez-Santana, C.; Castellote, C.; Izquierdo-Pulido, M.; Permanyer, J.; Franch, A.; Castell, M. Spleen lymphocyte function modulated by a cocoa-enriched diet. Clin. Exp. Immunol. 2007, 149, 535–542. [Google Scholar] [CrossRef]
- Camps-Bossacoma, M.; Pérez-Cano, F.J.; Franch, À.; Untersmayr, E.; Castell, M. Effect of a cocoa diet on the small intestine and gut-associated lymphoid tissue composition in an oral sensitization model in rats. J. Nutr. Biochem. 2017, 42, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Gracie, J.A.; Bradley, J.A. Interleukin-12 induces interferon-gamma-dependent switching of IgG alloantibody subclass. Eur. J. Immunol. 1996, 26, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Cuturi, M.C.; Josien, R.; Cantarovich, D.; Bugeon, L.; Anegon, I.; Menoret, S.; Smit, H.; Douillard, P.; Soulillou, J.-P. Decreased anti-donor major histocompatibility complex class I and increased class II alloantibody response in allograft tolerance in adult rats. Eur. J. Immunol. 1994, 24, 1627–1631. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cano, F.J.; Castellote, C.; Marín-Gallén, S.; González-Castro, A.; Franch, À.; Castell, M. Phenotypic and functional characteristics of rat spleen lymphocytes during suckling. Dev. Comp. Immunol. 2007, 31, 1264–1277. [Google Scholar] [CrossRef]
- Schumacher, M.A.; Danopoulos, S.; Al Alam, D.; Frey, M.R. Growth factors in the intestinal tract. In Physiology of the Gastrointestinal Tract, 6th ed.; Said, H., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 71–101. [Google Scholar]
- Friedman, M.I. Some determinants of milk ingestion in suckling rats. J. Comp. Physiol. 1975, 89, 636–647. [Google Scholar] [CrossRef]
- Corsini, I.; Polvani, S.; Tarocchi, M.; Tempesti, S.; Marroncini, G.; Generoso, M.; Bresci, C.; Gozzini, E.; Bianconi, T.; Galli, A.; et al. Peroxisome proliferator-activated receptor-γ agonist pioglitazone reduces the development of necrotizing enterocolitis in a neonatal preterm rat model. Pediatr. Res. 2017, 81, 364–368. [Google Scholar] [CrossRef]
- Satoh, T.; Izumi, H.; Iwabuchi, N.; Odamaki, T.; Namba, K.; Abe, F.; Xiao, J.Z. Bifidobacterium breve prevents necrotising enterocolitis by suppressing inflammatory responses in a preterm rat model. Benef. Microbes 2016, 7, 75–82. [Google Scholar] [CrossRef]
- Kojima, T.; Nishimura, M.; Yajima, T.; Kuwata, T.; Suzuki, Y.-I.; Goda, T.; Takase, S.; Harada, E. Effect of intermittent feeding on the development of disaccharidase activities in artificially reared rat pups. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1998, 121, 289–297. [Google Scholar] [CrossRef]
- Toriz, C.G.; Melo, A.I.; Solano-Agama, C.; Gómez-Domínguez, E.G.; Martínez-Muñoz, M.L.A.; Castañeda-Obeso, J.; Vera-Aguilar, E.; Aguirre-Benítez, E.L.; Romero-Aguilar, L.; González-del Pliego, M.; et al. Physiological changes of growth hormone during lactation in pup rats artificially reared. PLoS ONE 2019, 14, e0220853. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, G. Epidermal growth factor is a major growth-promoting agent in human milk. Science 1980, 210, 198–199. [Google Scholar] [CrossRef] [PubMed]
- Opleta, K.; O’Loughlin, E.V.; Shaffer, E.A.; Hayden, J.; Hollenberg, M.; Gall, D.G. Effect of epidermal growth factor on growth and postnatal development of the rabbit liver. Am. J. Physiol. 1987, 253, G622–G626. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Masuda, Y.; Ohta, H.; Tanaka, T.; Washida, M.; Nabeshima, Y.I.; Miyake, A.; Itoh, N.; Konishi, M. FgF21 regulates T-cell development in the neonatal and juvenile thymus. Sci. Rep. 2017, 7, 330. [Google Scholar] [CrossRef] [Green Version]
- Garly, M.L.; Trautner, S.L.; Marx, C.; Danebod, K.; Nielsen, J.; Ravn, H.; Martins, C.L.; Balé, C.; Aaby, P.; Lisse, I.M. Thymus size at 6 months of age and subsequent child mortality. J. Pediatr. 2008, 153, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Yajima, M.; Hoshiba, J.; Terahara, M.; Yajima, T. Reduced thymic size and numbers of splenic CD4+ and CD8+ cells in artificially reared mouse pups. Biosci. Biotechnol. Biochem. 2007, 71, 2420–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, K.; Koyanagi, A.; Kamachi, F.; Harauma, A.; Chiba, A.; Hisata, K.; Moriguchi, T.; Shimizu, T.; Miyake, S. Breast-feeding regulates immune system development via transforming growth factor-β in mice pups. Pediatr. Int. 2018, 60, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Adkins, B.; Leclerc, C.; Marshall-Clarke, S. Neonatal adaptive immunity comes of age. Nat. Rev. Immunol. 2004, 4, 553–564. [Google Scholar] [CrossRef]
- Adkins, B. Development of neonatal Th1/Th2 function. Int. Rev. Immunol. 2000, 19, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cano, F.J.; Franch, À.; Castellote, C.; Castell, M. The suckling rat as a model for immunonutrition studies in early life. Clin. Dev. Immunol. 2012, 2012, 537310. [Google Scholar] [CrossRef] [Green Version]
- Härtel, C.; Adam, N.; Strunk, T.; Temming, P.; Müller-Steinhardt, M.; Schultz, C. Cytokine responses correlate differentially with age in infancy and early childhood. Clin. Exp. Immunol. 2005, 142, 446–453. [Google Scholar] [CrossRef]
- Penttila, I. Effects of transforming growth factor-beta and formula feeding on systemic immune responses to dietary beta-lactoglobulin in allergy-prone rats. Pediatr. Res. 2006, 59, 650–655. [Google Scholar] [CrossRef] [Green Version]
- Kotiranta-Ainamo, A.; Rautonen, J.; Rautonen, N. Imbalanced cytokine secretion in newborns. Biol. Neonate 2004, 85, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, H.N.; HayGlass, K.T.; Gangur, V.; Allardice, J.G.; Embree, J.E.; Plummer, F.A. Analysis of neonatal T cell and antigen presenting cell functions. Hum. Immunol. 1997, 57, 69–79. [Google Scholar] [CrossRef]
- Itoh, N. FGF21 as a hepatokine, adipokine, and myokine in metabolism and diseases. Front. Endocrinol. (Lausanne) 2014, 5, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.M.; Yu, Y.H.; Li, L.; Wang, W.F.; Li, D.S. Treatment of CIA mice with FGF21 down-regulates TH17-IL-17 axis. Inflammation 2016, 39, 309–319. [Google Scholar] [CrossRef]
- Li, J.Y.; Wang, N.; Khoso, M.H.; Shen, C.B.; Guo, M.Z.; Pang, X.X.; Li, D.S.; Wang, W.F. FGF-21 elevated IL-10 production to correct LPS-induced inflammation. Inflammation 2018, 41, 751–759. [Google Scholar] [CrossRef]
- Wong, W.M.; Wright, N.A. Epidermal growth factor, epidermal growth factor receptors, intestinal growth, and adaptation. JPEN J. Parenter. Enteral Nutr. 1999, 23, S83–S88. [Google Scholar] [CrossRef]
- Sanjabi, S.; Oh, S.A.; Li, M.O. Regulation of the immune response by TGF-β: From conception to autoimmunity and infection. Cold Spring Harb. Perspect. Biol. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M. The Role of Milk Transforming Growth Factor-β (TGF-β) in the Development of the Infant Gut and Gut Mucosal Immune System; University of Adelaide: Adelaide, Australia, 2000. [Google Scholar]
- Grases-Pintó, B.; Torres-Castro, P.; Marín-Morote, L.; Abril-Gil, M.; Castell, M.; Rodríguez-Lagunas, M.J.; Pérez-Cano, F.J.; Franch, À. Leptin and EGF supplementation enhance the immune system maturation in preterm suckling rats. Nutrients 2019, 11, 2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín-Gallén, S.; Pérez-Cano, F.J.; Castell, M.; Castellote, C.; Franch, A. Intestinal intraepithelial NK and NKT cell ontogeny in Lewis rats. Dev. Comp. Immunol. 2008, 32, 1405–1408. [Google Scholar] [CrossRef] [PubMed]
- Kalliomäki, M.A.; Isolauri, E. Probiotics and down-regulation of the allergic response. Immunol. Allergy Clin. N. Am. 2004, 24, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Balato, A.; Unutmaz, D.; Gaspari, A.A. Natural killer T cells: An unconventional T-cell subset with diverse effector and regulatory functions. J. Investig. Dermatol. 2009, 129, 1628–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamura, C.; Nakayama, T. Role of NKT cells in allergic asthma. Curr. Opin. Immunol. 2010, 22, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Burbank, A.J.; Sood, A.K.; Kesic, M.J.; Peden, D.B.; Hernandez, M.L. Environmental determinants of allergy and asthma in early life. J. Allergy Clin. Immunol. 2017, 140, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Caminati, M.; Durić-Filipović, I.; Arasi, S.; Peroni, D.G.; Živković, Z.; Senna, G. Respiratory allergies in childhood: Recent advances and future challenges. Pediatr. Allergy Immunol. 2015, 26, 702–710. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Castro, P.; Grases-Pintó, B.; Abril-Gil, M.; Castell, M.; Rodríguez-Lagunas, M.J.; Pérez-Cano, F.J.; Franch, À. Modulation of the Systemic Immune Response in Suckling Rats by Breast Milk TGF-β2, EGF and FGF21 Supplementation. Nutrients 2020, 12, 1888. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12061888
Torres-Castro P, Grases-Pintó B, Abril-Gil M, Castell M, Rodríguez-Lagunas MJ, Pérez-Cano FJ, Franch À. Modulation of the Systemic Immune Response in Suckling Rats by Breast Milk TGF-β2, EGF and FGF21 Supplementation. Nutrients. 2020; 12(6):1888. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12061888
Chicago/Turabian StyleTorres-Castro, Paulina, Blanca Grases-Pintó, Mar Abril-Gil, Margarida Castell, María J. Rodríguez-Lagunas, Francisco J. Pérez-Cano, and Àngels Franch. 2020. "Modulation of the Systemic Immune Response in Suckling Rats by Breast Milk TGF-β2, EGF and FGF21 Supplementation" Nutrients 12, no. 6: 1888. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12061888