Nuts and their Effect on Gut Microbiota, Gut Function and Symptoms in Adults: A Systematic Review and Meta-Analysis of Randomised Controlled Trials


Abstract
:1. Introduction
2. Materials and Methods
2.1. Eligibilty Criteria
2.2. Search Strategy
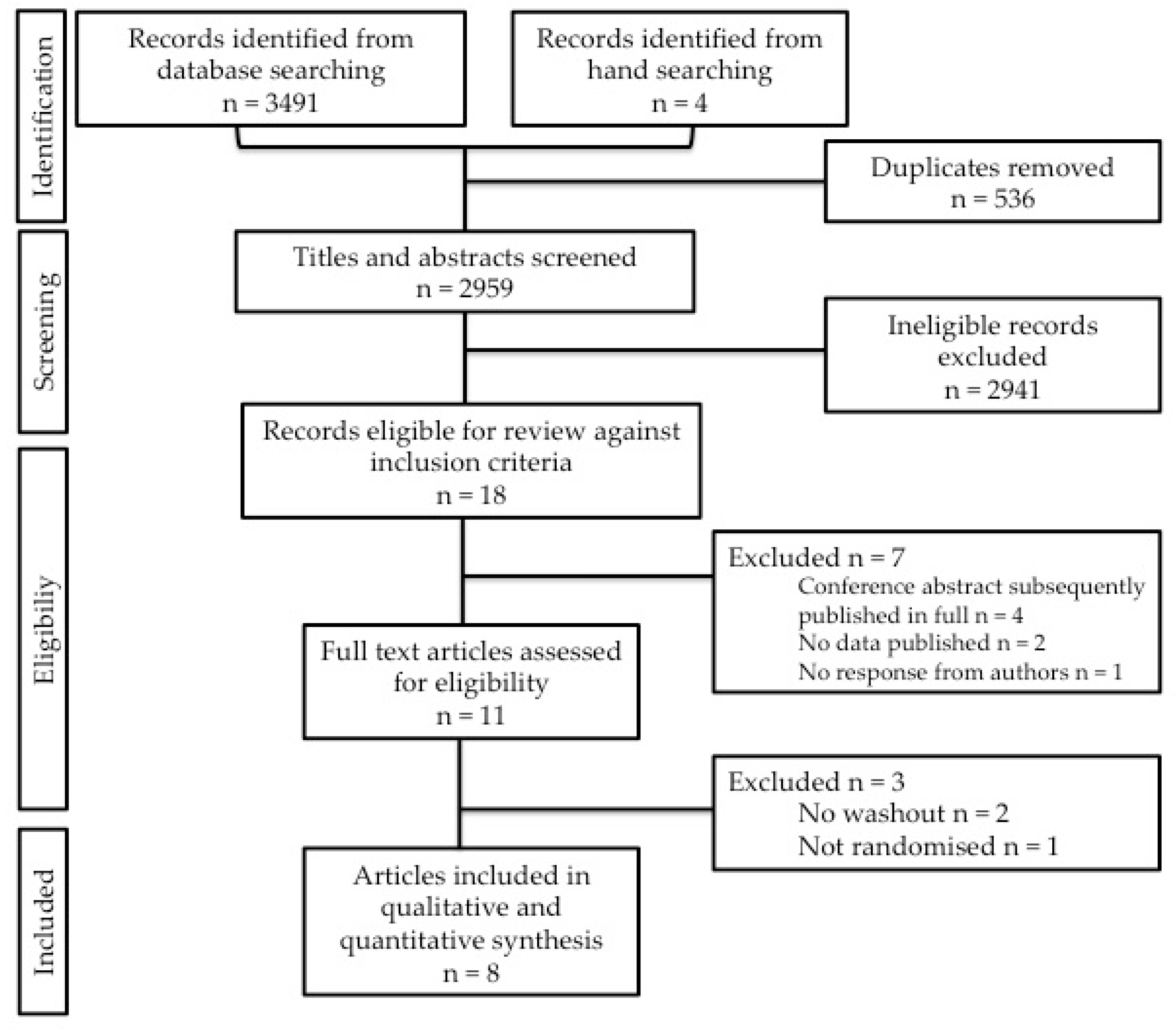
2.3. Screening
2.4. Data Extraction
2.5. Data Synthesis
3. Results
3.1. Study Characteristics
3.2. Outcomes
3.2.1. Summary of Meta-Analyses
3.2.2. Methods of Analysis and Reporting of Gut Microbiota
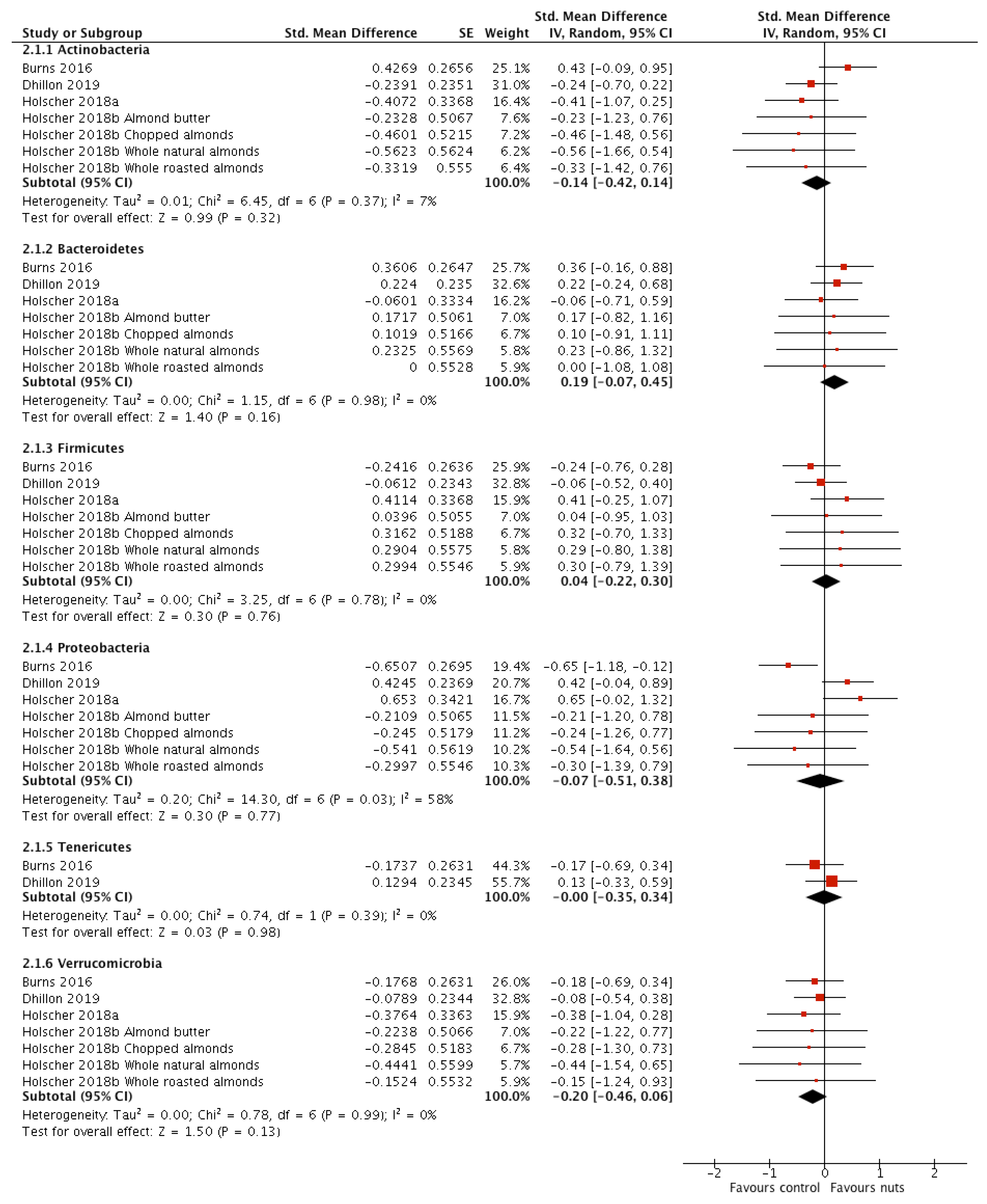
3.2.3. Relative Abundance of Bacterial Phyla
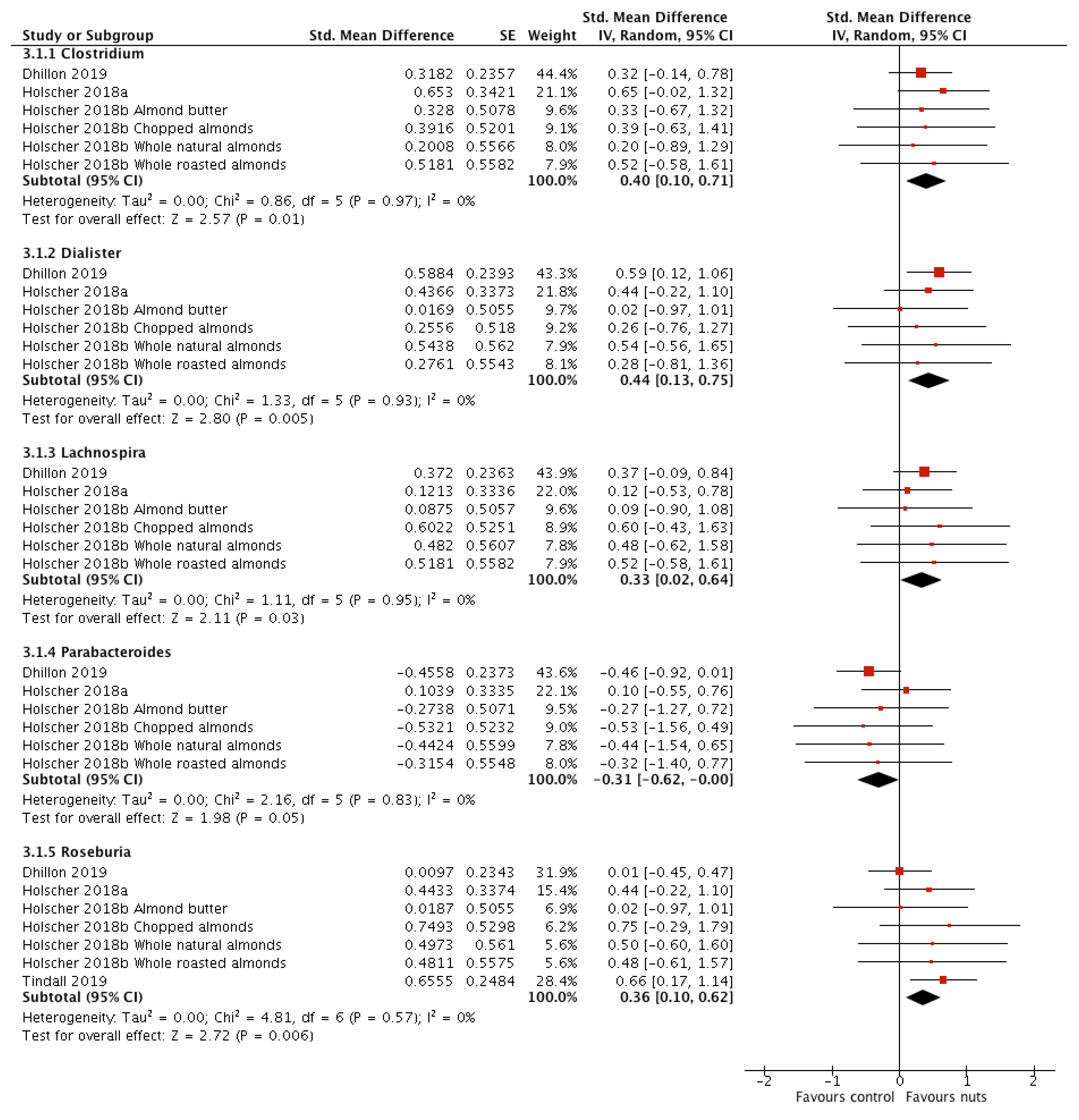
3.2.4. Relative Abundance of Bacterial Genera
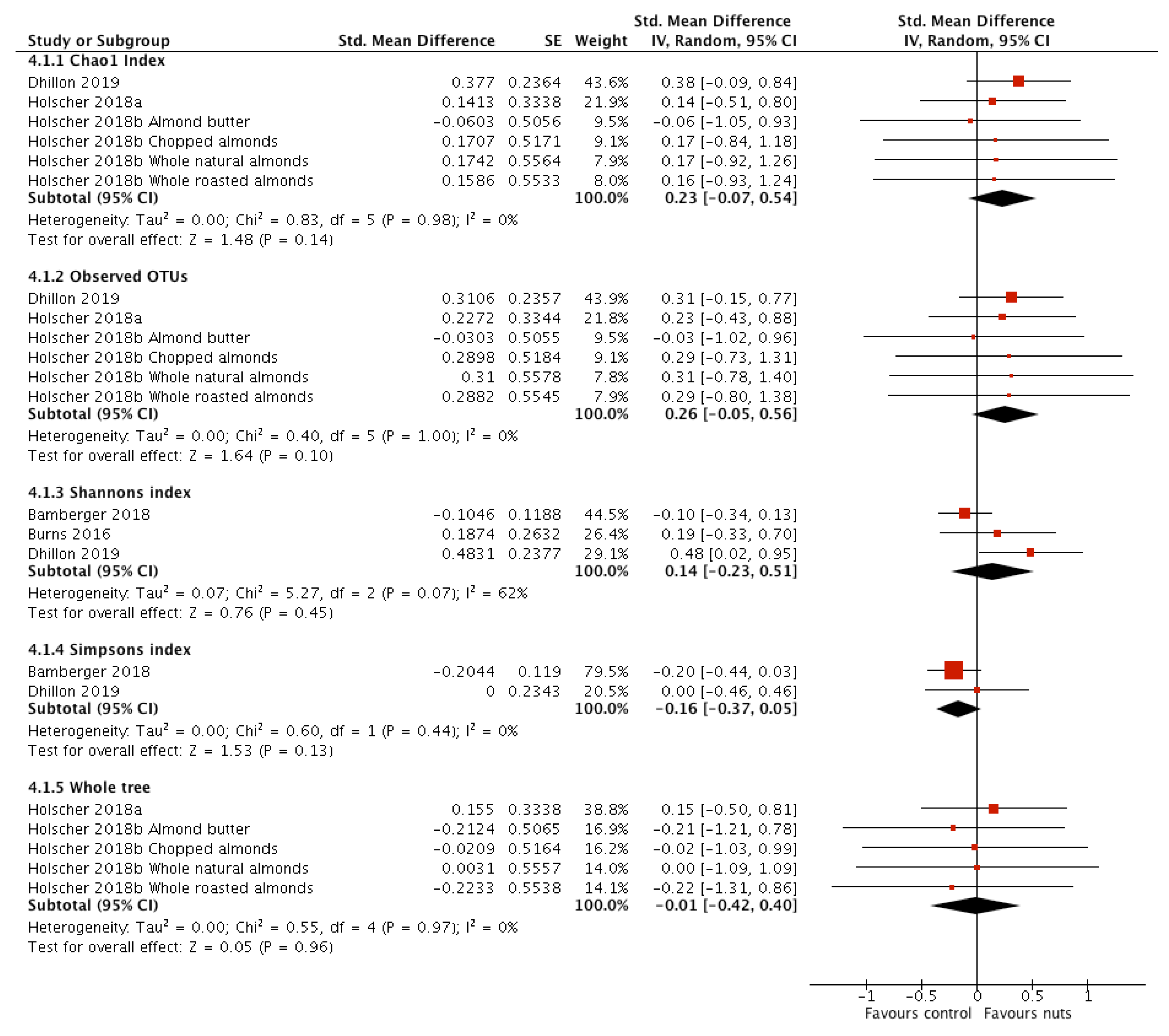
3.2.5. α-Diversity
3.2.6. β-Diversity
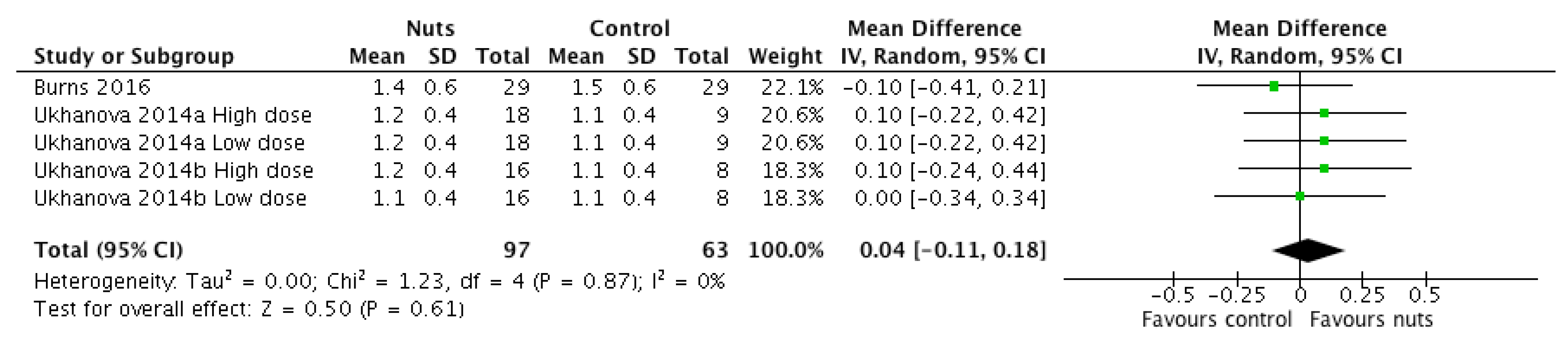
3.2.7. Stool Output
3.2.8. Gastrointestinal Symptoms
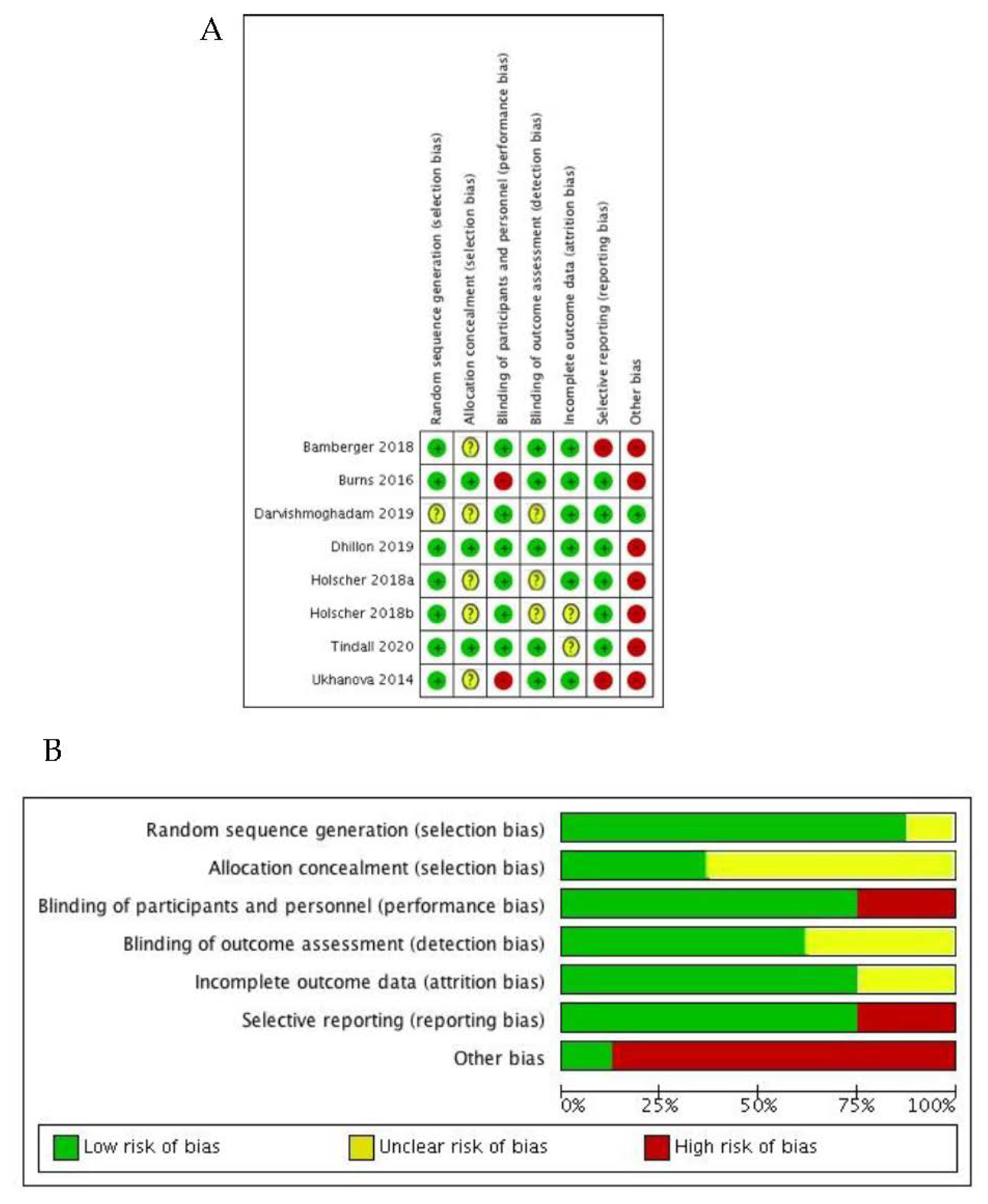
3.3. Risk of Bias
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mayhew, A.J.; de Souza, R.J.; Meyre, D.; Anand, S.S.; Mente, A. A systematic review and meta-analysis of nut consumption and incident risk of CVD and all-cause mortality. Br. J. Nutr. 2016, 115, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Neale, E.P.; Tapsell, L.C.; Guan, V.; Batterham, M.J. The effect of nut consumption on markers of inflammation and endothelial function: A systematic review and meta-analysis of randomised controlled trials. BMJ Open 2017, 7, e016863. [Google Scholar] [CrossRef] [PubMed]
- Tindall, A.M.; Johnston, E.A.; Kris-Etherton, P.M.; Petersen, K.S. The effect of nuts on markers of glycemic control: A systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2019, 109, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The Gut Microbiome in Health and in Disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-N.; Wu, H.; Chen, Y.-Z.; Chen, Y.-J.; Shen, X.-Z.; Liu, T.-T. Altered molecular signature of intestinal microbiota in irritable bowel syndrome patients compared with healthy controls: A systematic review and meta-analysis. Dig. Liver Dis. 2017, 49, 331–337. [Google Scholar] [CrossRef]
- Attaluri, A.; Jackson, M.; Valestin, J.; Rao, S.S.C. Methanogenic flora is associated with altered colonic transit but not stool characteristics in constipation without IBS. Am. J. Gastroenterol. 2010, 105, 1407–1411. [Google Scholar] [CrossRef] [Green Version]
- Jalanka-Tuovinen, J.; Salonen, A.; Nikkilä, J.; Immonen, O.; Kekkonen, R.; Lahti, L.; Palva, A.; de Vos, W.M. Intestinal microbiota in healthy adults: Temporal analysis reveals individual and common core and relation to intestinal symptoms. PLoS ONE 2011, 6, e23035. [Google Scholar] [CrossRef]
- Lamuel-Raventos, R.M.; Onge, M.P.S. Prebiotic nut compounds and human microbiota. Crit. Rev. Food Sci. Nutr. 2017, 57, 3154–3163. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Mandalari, G.; Nueno-Palop, C.; Bisignano, G.; Wickham, M.S.J.; Narbad, A. Potential prebiotic properties of almond (Amygdalus communis L.) seeds. Appl. Environ. Microbiol. 2008, 74, 4264–4270. [Google Scholar] [CrossRef] [Green Version]
- Blaiotta, G.; La Gatta, B.; Di Capua, M.; Di Luccia, A.; Coppola, R.; Aponte, M. Effect of chestnut extract and chestnut fiber on viability of potential probiotic Lactobacillus strains under gastrointestinal tract conditions. Food Microbiol. 2013, 36, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.W.; de Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Ellis, P.R.; Kendall, C.W.C.; Ren, Y.; Parker, C.; Pacy, J.F.; Waldron, K.W.; Jenkins, D.J.A. Role of cell walls in the bioaccessibility of lipids in almond seeds. Am. J. Clin. Nutr. 2004, 80, 604–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Lin, X.; Huang, G.; Zhang, W.; Rao, P.; Ni, L. Prebiotic effects of almonds and almond skins on intestinal microbiota in healthy adult humans. Anaerobe 2014, 26, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bamberger, C.; Rossmeier, A.; Lechner, K.; Wu, L.; Waldmann, E.; Fischer, S.; Stark, R.G.; Altenhofer, J.; Henze, K.; Parhofer, K.G. A Walnut-Enriched Diet Affects Gut Microbiome in Healthy Caucasian Subjects: A Randomized, Controlled Trial. Nutrients 2018, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- Holscher, H.D.; Guetterman, H.M.; Swanson, K.S.; An, R.; Matthan, N.R.; Lichtenstein, A.H.; Novotny, J.A.; Baer, D.J. Walnut Consumption Alters the Gastrointestinal Microbiota, Microbially Derived Secondary Bile Acids, and Health Markers in Healthy Adults: A Randomized Controlled Trial. J. Nutr. 2018, 148, 861–867. [Google Scholar] [CrossRef]
- Burns, A.M.; Zitt, M.A.; Rowe, C.C.; Langkamp-Henken, B.; Mai, V.; Nieves, C.; Ukhanova, M.; Christman, M.C.; Dahl, W.J. Diet quality improves for parents and children when almonds are incorporated into their daily diet: A randomized, crossover study. Nutr. Res. 2016, 36, 80–89. [Google Scholar] [CrossRef]
- Holscher, H.D.; Taylor, A.M.; Swanson, K.S.; Novotny, J.A.; Baer, D.J. Almond consumption and processing affects the composition of the gastrointestinal microbiota of healthy adult men and women: A randomized controlled trial. Nutrients 2018, 10, 126. [Google Scholar] [CrossRef] [Green Version]
- Ukhanova, M.; Wang, X.; Baer, D.J.; Novotny, J.A.; Fredborg, M.; Mai, V. Effects of almond and pistachio consumption on gut microbiota composition in a randomised cross-over human feeding study. Br. J. Nutr. 2014, 111, 2146–2152. [Google Scholar] [CrossRef]
- Dhillon, J.; Li, Z.; Ortiz, R.M. Almond Snacking for 8 wk Increases Alpha-Diversity of the Gastrointestinal Microbiome and Decreases Bacteroides fragilis Abundance Compared with an Isocaloric Snack in College Freshmen. Curr. Dev. Nutr. 2019, 3, nzz079. [Google Scholar] [CrossRef] [Green Version]
- Darvishmoghadam, S.; Ansari, M.; Ahmadipour, H.; Azimipour, M.; Esmaeilzade, M.; Zahedi, M.J.; Hayat Bakhsh Abbasi, M.; Ahmadi, B.; Asadi, A.; Sharififar, F. Assessment of the effect of sachet formulation of almond (Amygdalus dulcis L.) on diarrhea prominent irritable bowel syndrome (IBS-D) symptoms: A clinical trial. Complement. Ther. Med. 2019, 45, 242–247. [Google Scholar] [CrossRef]
- Higgins, J.; Thomas, J.; Chandler, J.; Cumpston, M.; Li, T.; Page, M.; Welch, V. Cochrane Handbook for Systematic Reviews of Interventions. Available online: http://www.training.cochrane.org/handbook (accessed on 31 March 2020).
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. PLoS Med. 2009, 6, 1–28. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, C.E.; Fulgoni, V.L.; Nicklas, T.A.; Nicklas, T.A. Tree Nut consumption is associated with better adiposity measures and cardiovascular and metabolic syndrome health risk factors in U.S. Adults: NHANES 2005–2010. Nutr. J. 2015, 14, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.P.T.; Altman, D.G.; Gøtzsche, P.C.; Jüni, P.; Moher, D.; Oxman, A.D.; Savović, J.; Schulz, K.F.; Weeks, L.; Sterne, J.A.C. The Cochrane Collaboration’s tool for assessing risk of bias in randomised trials. BMJ 2011, 343, d5928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tindall, A.M.; McLimans, C.J.; Petersen, K.S.; Kris-Etherton, P.M.; Lamendella, R. Walnuts and Vegetable Oils Containing Oleic Acid Differentially Affect the Gut Microbiota and Associations with Cardiovascular Risk Factors: Follow-up of a Randomized, Controlled, Feeding Trial in Adults at Risk for Cardiovascular Disease. J. Nutr. 2020, 150, 806–817. [Google Scholar] [CrossRef] [PubMed]
- Finotello, F.; Mastrorilli, E.; Di Camillo, B. Measuring the diversity of the human microbiota with targeted next-generation sequencing. Brief. Bioinform. 2016, bbw119, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.; Sue, A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science (80-) 2012, 334, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 1–19. [Google Scholar] [CrossRef] [Green Version]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic inulin-type fructans induce specific changes in the human gut microbiota. Gut 2017, 66, 1968–1974. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, W.; Huang, G.; Zhang, W.; Ni, L. In vitro and in vivo evaluation of the prebiotic effect of raw and roasted almonds (Prunus amygdalus). J. Sci. Food Agric. 2016, 96, 1836–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandalari, G.; Faulks, R.M.; Bisignano, C.; Waldron, K.W.; Narbad, A.; Wickham, M.S.J. In vitro evaluation of the prebiotic properties of almond skins (Amygdalus communis L.). FEMS Microbiol. Lett. 2010, 304, 116–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meehan, C.J.; Beiko, R.G. A phylogenomic view of ecological specialization in the Lachnospiraceae, a family of digestive tract-associated bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bisanz, J.E.; Upadhyay, V.; Turnbaugh, J.A.; Ly, K.; Turnbaugh, P.J. Meta-Analysis Reveals Reproducible Gut Microbiome Alterations in Response to a High-Fat Diet. Cell Host Microbe 2019, 26, 265–272.e4. [Google Scholar] [CrossRef]
- Staley, C.; Weingarden, A.R.; Khoruts, A.; Sadowsky, M.J. Interaction of gut microbiota with bile acid metabolism and its influence on disease states. Appl. Microbiol. Biotechnol. 2017, 101, 47–64. [Google Scholar] [CrossRef] [Green Version]
- Lambeth, S.M.; Carson, T.; Lowe, J.; Ramaraj, T.; Leff, J.W.; Luo, L.; Bell, C.J.; Shah, V.O. Composition, Diversity and Abundance of Gut Microbiome in Prediabetes and Type 2 Diabetes. J. Diabetes Obes. 2015, 2, 1–7. [Google Scholar]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Goffau, M.C.; Luopajärvi, K.; Knip, M.; Ilonen, J.; Ruohtula, T.; Härkönen, T.; Orivuori, L.; Hakala, S.; Welling, G.W.; Harmsen, H.J.; et al. Fecal microbiota composition differs between children with β-cell autoimmunity and those without. Diabetes 2013, 62, 1238–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manichanh, C.; Rigottier-Gois, L.; Bonnaud, E.; Gloux, K.; Pelletier, E.; Frangeul, L.; Nalin, R.; Jarrin, C.; Chardon, P.; Marteau, P.; et al. Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut 2006, 55, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Grassby, T.; Picout, D.R.; Mandalari, G.; Faulks, R.M.; Kendall, C.W.C.; Rich, G.T.; Wickham, M.S.J.; Lapsley, K.; Ellis, P.R. Modelling of nutrient bioaccessibility in almond seeds based on the fracture properties of their cell walls. Food Funct. 2014, 5, 3096–3106. [Google Scholar] [CrossRef]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra, A.; Latreille-Barbier, M.; Donazzolo, Y.; Pelletier, X.; Ouwehand, A.C. Effects of 28-day Bifidobacterium animalis subsp. lactis HN019 supplementation on colonic transit time and gastrointestinal symptoms in adults with functional constipation: A double-blind, randomized, placebo-controlled, and dose-ranging trial. Gut Microbes 2018, 9, 236–251. [Google Scholar] [CrossRef] [Green Version]
- Ringel, Y. The Gut Microbiome in Irritable Bowel Syndrome and Other Functional Bowel Disorders. Gastroenterol. Clin. N. Am. 2017, 46, 91–101. [Google Scholar] [CrossRef]
- Gebauer, S.K.; Novotny, J.A.; Bornhorst, G.M.; Baer, D.J. Food processing and structure impact the metabolizable energy of almonds. Food Funct. 2016, 7, 4231–4238. [Google Scholar] [CrossRef] [Green Version]
- Grundy, M.M.L.; Grassby, T.; Mandalari, G.; Waldron, K.W.; Butterworth, P.J.; Berry, S.E.E.; Ellis, P.R. Effect of mastication on lipid bioaccessibility of almonds in a randomized human study and its implications for digestion kinetics, metabolizable energy, and postprandial lipemia. Am. J. Clin. Nutr. 2015, 101, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Baer, D.J.; Novotny, J.A. Metabolizable Energy from Cashew Nuts is Less than that Predicted by Atwater Factors. Nutrients 2018, 11, 33. [Google Scholar] [CrossRef] [Green Version]
- Baer, D.J.; Gebauer, S.K.; Novotny, J.A. Measured energy value of pistachios in the human diet. Br. J. Nutr. 2012, 107, 120–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baer, D.J.; Gebauer, S.K.; Novotny, J.A. Walnuts Consumed by Healthy Adults Provide Less Available Energy than Predicted by the Atwater Factors. J. Nutr. 2016, 146, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Palermo, M.; Pellegrini, N.; Fogliano, V. The effect of cooking on the phytochemical content of vegetables. J. Sci. Food Agric. 2014, 94, 1057–1070. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PICOS | Inclusion and Exclusion Criteria | Data Extraction |
|---|---|---|
| Participants | Adults who were healthy/experiencing minor organ dysfunction not requiring inpatient care were included. Trials comparing clinical populations to healthy populations were excluded unless the impact of nut consumption on the healthy population alone could be extracted. Trials exclusively in children, animals, ex vivo or in vitro were excluded. There were no restrictions for sex or ethnicity. | Age, sex, inclusion and exclusion criteria, number of participants randomised to control and intervention groups. |
| Interventions | Interventions consisting of a minimum dose of 7 g of tree nuts and/or peanuts per day [25] for a minimum duration of 1 week were eligible. Trials administering nuts with or without skins were eligible as were trials consisting of processed nuts provided the nuts had not been mixed with additional ingredients. Trials administering individual nut components alone, e.g., skins/oils were excluded. Eligible trials were those in which the nuts were provided to participants; interventions consisting of dietary advice to increase nut consumption were excluded. Mixed interventions were excluded unless it was possible to extract the effect of nut consumption alone. Trials with multiple intervention arms were included. | Nut type, dose, frequency, duration of intervention, presentation, processing, instructions for consumption. |
| Comparators | Trials comparing consumption of nuts to a control involving consumption of no nuts were included. When trials were conducted in controlled feeding environments, only those in which control and intervention diets were matched in energy were included. | Type and dose of comparator, nutrient composition of intervention and control foods. |
| Outcomes | Trials reporting outcomes relating to faecal microbiota, such as composition or outcomes assessing bacterial activity, were included. Trials reporting clinical subjective or objective measures of gut function including gut transit time, stool form and frequency or gut symptoms were included. | Outcomes measured, method of measurement, baseline and endpoint values or change from baseline. Adverse events and compliance. |
| Study design | Only randomised controlled trials, utilising parallel or crossover designs, were eligible. If crossover design was used only trials with a washout period were eligible to limit carryover effects. Studies conducted in controlled feeding or free-living environments were eligible. | Study design, duration of washout period, intention to treat analysis, number of excluded participants, reasons for exclusion, randomisation and blinding methods. |
| Study | Participants | Sample Size (%female) | Nut Type | Dose (g/d) | Duration | Comparator | Outcomes Included in Meta-Analysis | Trial Design | Washout |
|---|---|---|---|---|---|---|---|---|---|
| Bamberger 2018 [16] | Healthy adults | 142 (64.7) | Walnuts | 43 | 8 wk | Western style control—dietary advice | Microbiota | Crossover Free-living | 4 wk |
| Burns 2016 [18] | Healthy adults | 29 (82.8) | Almonds | 42.5 | 3 wk | Usual diet (avoid nuts) | Microbiota, stool frequency | Crossover Free-living | 6 wk |
| Darvishmogh-adam 2019 [22] | IBS-D—Rome IV criteria | 50 (58.0) | Almonds | 40 | 20 d | Wheat | None | Parallel Free-living | NA |
| Dhillon 2019 [21] | Healthy adults | 73 (56.2) | Almonds | 56.7 | 8 wk | Graham crackers | Microbiota | Parallel Free-living | NA |
| Holscher 2018a [17] | Healthy adults | 18 (44.4) | Walnuts | 42 | 3 wk | Base diet (food provided) | Microbiota | Crossover Controlled feeding | 1 wk |
| Holscher 2018b [19] | Healthy adults | 18 (44.4) | Almonds (whole natural, whole roasted, chopped, butter) | 42 | 3 wk | Base diet (food provided) | Microbiota | Crossover Controlled feeding | 1 wk |
| Tindall 2020 [27] | Adults at risk of cardiovascular disease | 42 (45.2) | Walnuts | 57–99 g (18% daily energy intake) | 6 wk | Base diet with ALA from walnut matched for oleic acid | Microbiota | Crossover Controlled feeding | Mean: 23 d Range: 1–164 d |
| Ukhanova 2014 [20] | Healthy adults | Almond 18 (44.4) Pistachio 16 (50.0) | Almond OR Pistachio | 42 OR 84 | 18 d | Base diet (food provided) | Stool frequency | Crossover Controlled feeding | Almond: 1 wk Pistachio: 2 wk |
| Outcome | No. of Studies in the Meta-Analysis (Ref) | Results | Heterogeneity | ||||
|---|---|---|---|---|---|---|---|
| Participants n | Meta-Analysis Overall Estimate (95% CI) | P | Chi-Square Test | P | I2 (%) | ||
| Phyla | |||||||
| p_Actinobacteria | 4 [17,18,19,21] | 138 | −0.14 (−0.42, 0.14) | 0.32 | 6.45 | 0.37 | 7 |
| p_Bacteroidetes | 4 [17,18,19,21] | 138 | 0.19 (−0.07, 0.45) | 0.16 | 1.15 | 0.98 | 0 |
| p_Firmicutes | 4 [17,18,19,21] | 138 | 0.04 (−0.22, 0.30) | 0.76 | 3.25 | 0.78 | 0 |
| p_Proteobacteria | 4 [17,18,19,21] | 138 | −0.07 (−0.51, 0.38) | 0.77 | 14.3 | 0.03 | 58 |
| p_Tenericutes | 2 [18,21] | 102 | −0.00 (−0.35, 0.34) | 0.98 | 0.74 | 0.39 | 0 |
| p_Verrucomicrobia | 4 [17,18,19,21] | 138 | −0.20 (−0.46, 0.06) | 0.13 | 0.78 | 0.99 | 0 |
| Genus | |||||||
| g_Faecalibacterium | 4 [17,19,21,27] | 151 | 0.11 (−0.16, 0.38) | 0.43 | 6.45 | 0.37 | 7 |
| g_Roseburia | 4 [17,19,21,27] | 151 | 0.36 (0.10, 0.62) | 0.006 | 4.81 | 0.57 | 0 |
| g_Streptococcus | 4 [17,19,21,27] | 151 | −0.02 (−0.27, 0.24) | 0.91 | 0.08 | 1 | 0 |
| g_Blautia | 4 [16,17,19,21] | 251 | −0.15 (−0.34, 0.03) | 0.11 | 2.25 | 0.89 | 0 |
| g_Bifidobacteria | 4 [16,17,19,21] | 251 | −0.09 (−0.39, 0.21) | 0.56 | 9.31 | 0.16 | 36 |
| g_Coprococcus | 3 [17,19,21] | 109 | −0.10 (−0.41, 0.20) | 0.52 | 0.77 | 0.98 | 0 |
| g_Lachnospira | 3 [17,19,21] | 109 | 0.33 (0.02, 0.64) | 0.03 | 1.11 | 0.95 | 0 |
| g_Ruminococcus | 3 [17,19,21] | 109 | −0.10 (−0.40, 0.21) | 0.54 | 1.38 | 0.93 | 0 |
| g_Dorea | 3 [17,19,21] | 109 | −0.08 (−0.39, 0.22) | 0.59 | 1.24 | 0.94 | 0 |
| g_Clostridium | 3 [17,19,21] | 109 | 0.40 (0.10, 0.71) | 0.01 | 0.86 | 0.97 | 0 |
| g_Oscillospira | 3 [17,19,21] | 109 | −0.10 (−0.42, 0.22) | 0.55 | 5.25 | 0.39 | 5 |
| g_Dialister | 3 [17,19,21] | 109 | 0.44 (0.13, 0.75) | 0.005 | 1.33 | 0.93 | 0 |
| g_Bacteroides | 3 [17,19,21] | 109 | 0.08 (−0.23, 0.38) | 0.61 | 0.23 | 1 | 0 |
| g_Parabacteroides | 3 [17,19,21] | 109 | −0.31 (−0.62, −0.00) | 0.05 | 2.16 | 0.83 | 0 |
| g_Collinsella | 3 [17,19,21] | 109 | −0.16 (−0.46, 0.15) | 0.32 | 0.92 | 0.97 | 0 |
| g_Akkermansia | 3 [17,19,21] | 109 | −0.21 (−0.51, 0.10) | 0.18 | 0.77 | 0.98 | 0 |
| g_Anaerostipes | 3 [16,17,21] | 233 | 0.09 (−0.47, 0.64) | 0.75 | 9.79 | 0.007 | 80 |
| g_Phascolarctobacterium | 2 [19,21] | 91 | 0.16 (−0.19, 0.50) | 0.37 | 0.75 | 0.94 | 0 |
| g_Prevotella | 2 [19,21] | 91 | 0.14 (−0.21, 0.48) | 0.44 | 0.44 | 0.98 | 0 |
| Alpha diversity | |||||||
| Chao-1 index | 3 [17,19,21] | 109 | 0.23 (−0.07, 0.54) | 0.14 | 0.83 | 0.98 | 0 |
| Observed OTUs | 3 [17,19,21] | 109 | 0.26 (−0.05, 0.56) | 0.10 | 0.40 | 1 | 0 |
| Shannon index | 3 [16,18,21] | 244 | 0.14 (−0.23, 0.51) | 0.45 | 5.27 | 0.07 | 62 |
| Simpson index | 2 [16,21] | 215 | −0.16 (−0.37, 0.05) | 0.13 | 0.60 | 0.44 | 0 |
| Whole tree | 2 [17,19] | 36 | −0.01 (−0.42, 0.40) | 0.96 | 0.55 | 0.97 | 0 |
| Stool frequency 2 | 3 [18,20] | 63 | 0.04 (−0.11, 0.18) | 0.61 | 1.23 | 0.87 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Creedon, A.C.; Hung, E.S.; Berry, S.E.; Whelan, K. Nuts and their Effect on Gut Microbiota, Gut Function and Symptoms in Adults: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients 2020, 12, 2347. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082347
Creedon AC, Hung ES, Berry SE, Whelan K. Nuts and their Effect on Gut Microbiota, Gut Function and Symptoms in Adults: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients. 2020; 12(8):2347. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082347
Chicago/Turabian StyleCreedon, Alice C., Estella S. Hung, Sarah E. Berry, and Kevin Whelan. 2020. "Nuts and their Effect on Gut Microbiota, Gut Function and Symptoms in Adults: A Systematic Review and Meta-Analysis of Randomised Controlled Trials" Nutrients 12, no. 8: 2347. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082347