Peptide Release after Simulated Infant In Vitro Digestion of Dry Heated Cow’s Milk Protein and Transport of Potentially Immunoreactive Peptides across the Caco-2 Cell Monolayer

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of Milk Powders and Heat Treatment
2.3. Infant In Vitro Digestion
2.4. Caco-2 Cell Culture
2.5. Transport across the Caco-2 Cell Monolayer
2.6. Peptide Analysis
2.7. Data Analysis
2.8. sIgE Binding Epitope Identification
2.9. T-Cell Epitope Prediction
3. Results
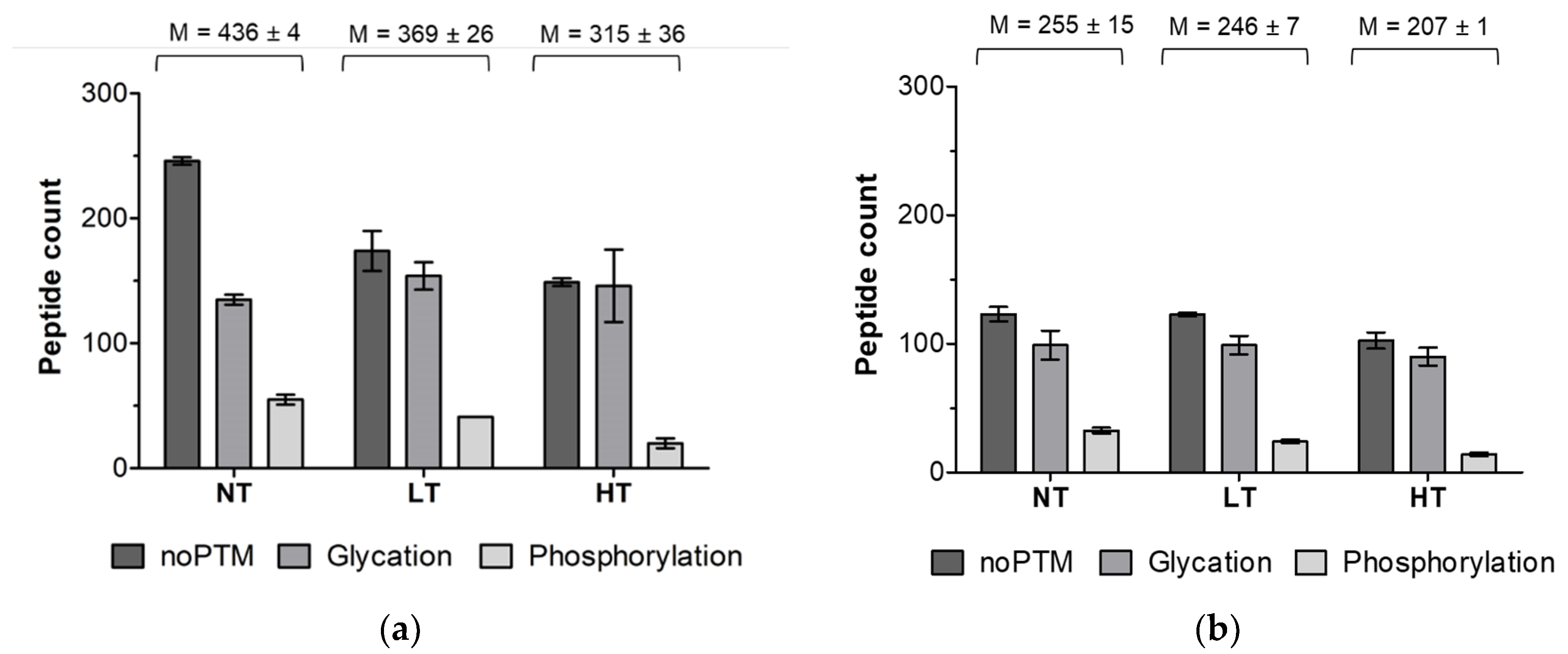
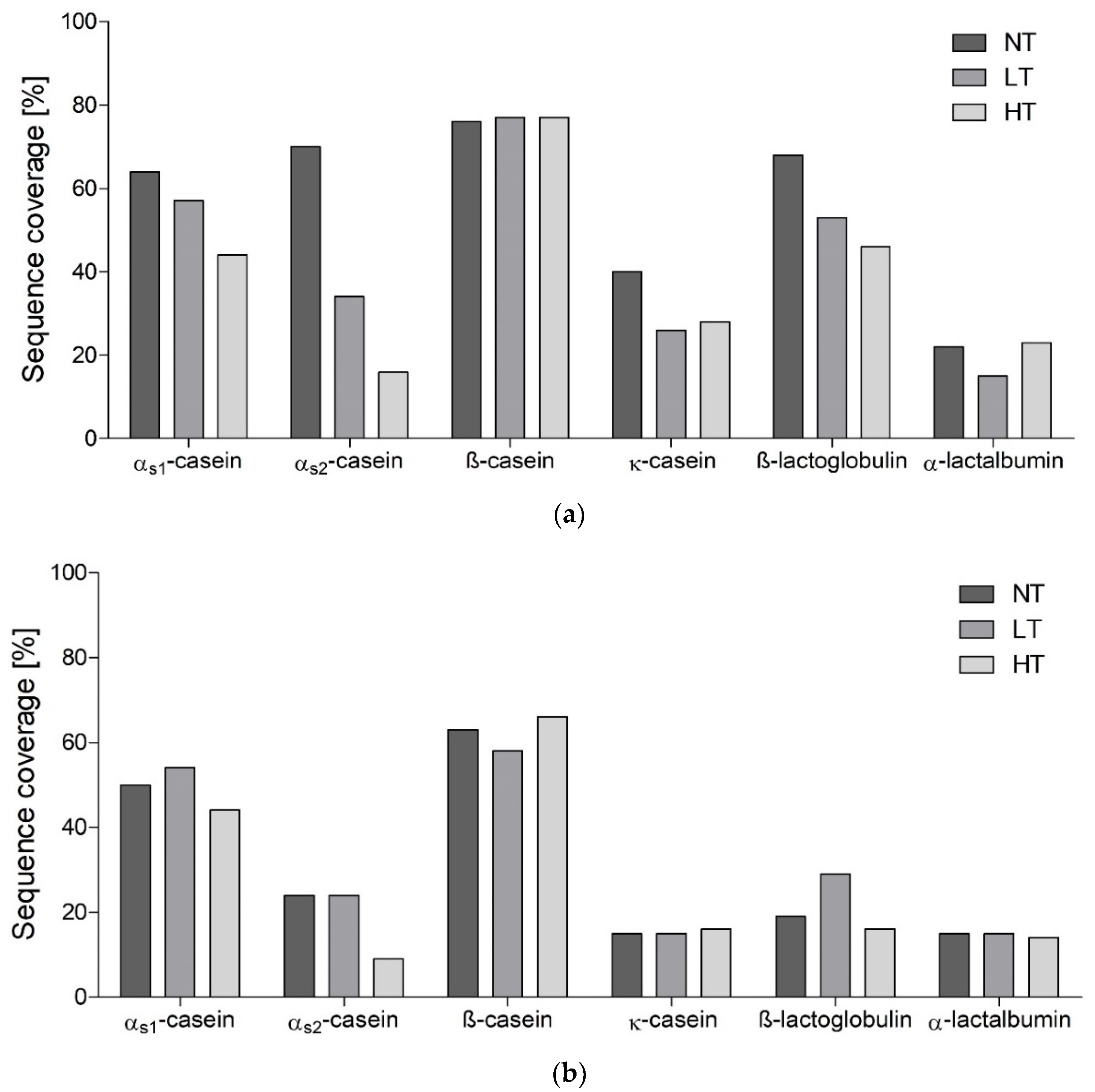
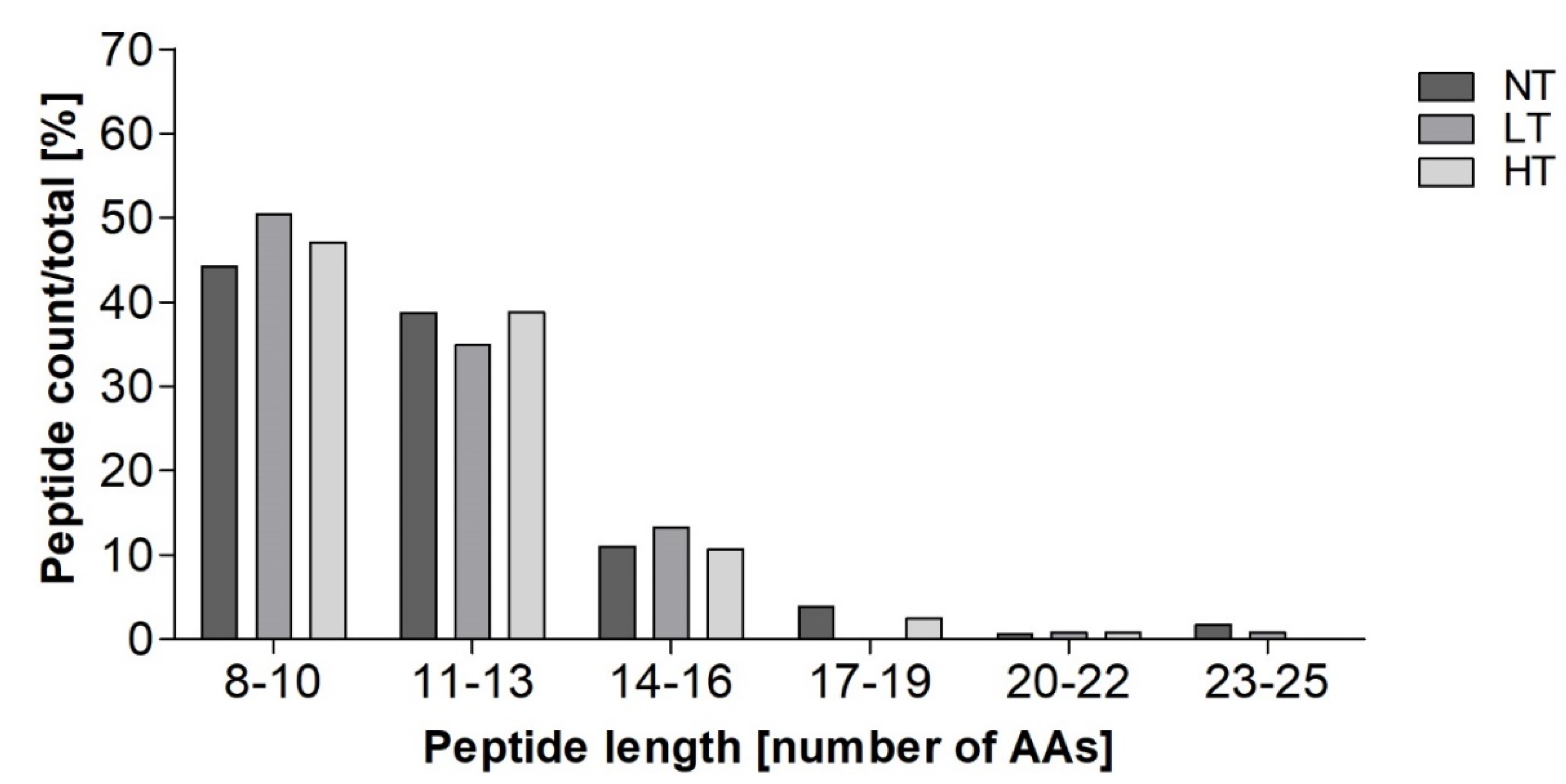
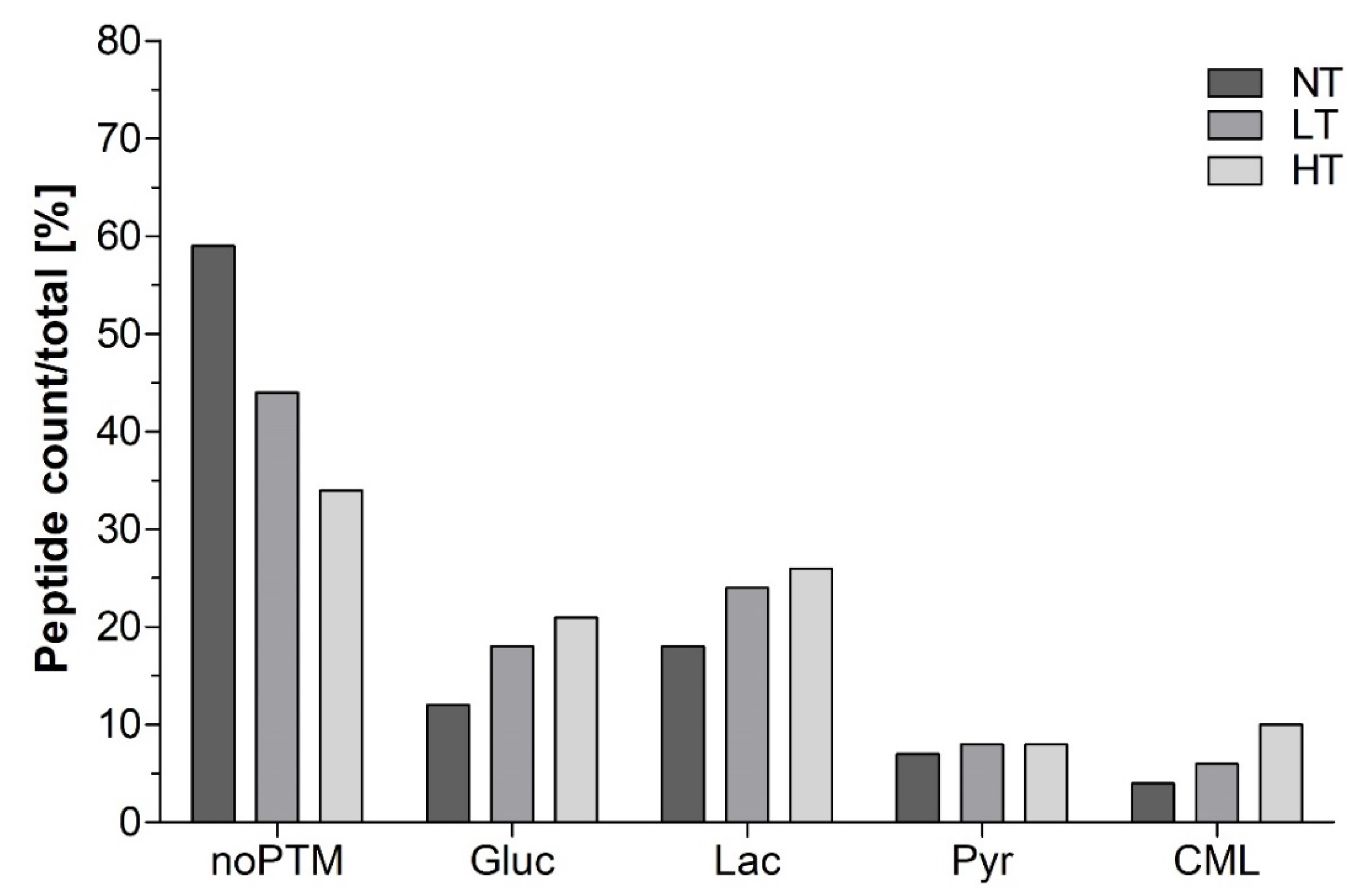
3.1. Identification of Peptides in In Vitro Digests
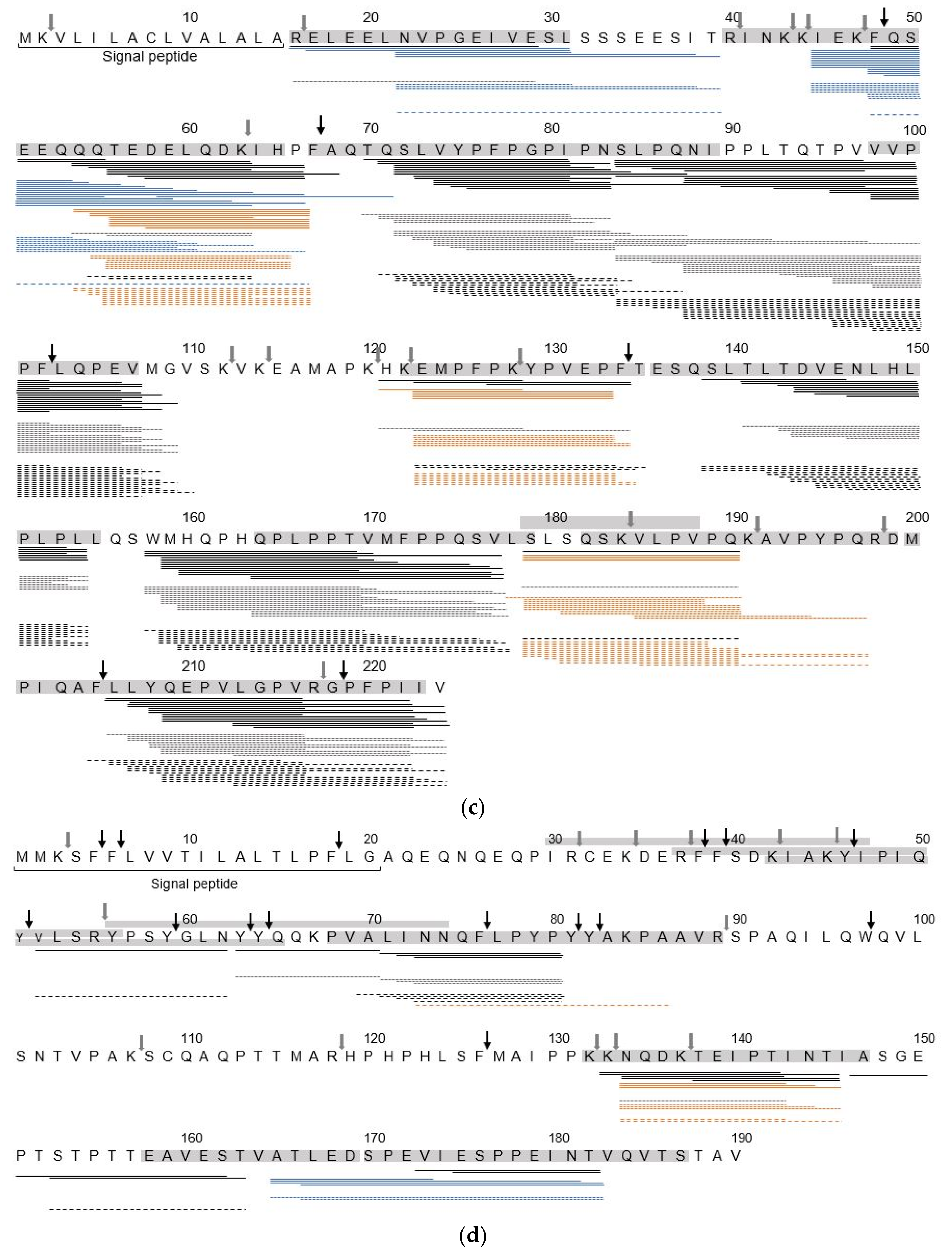
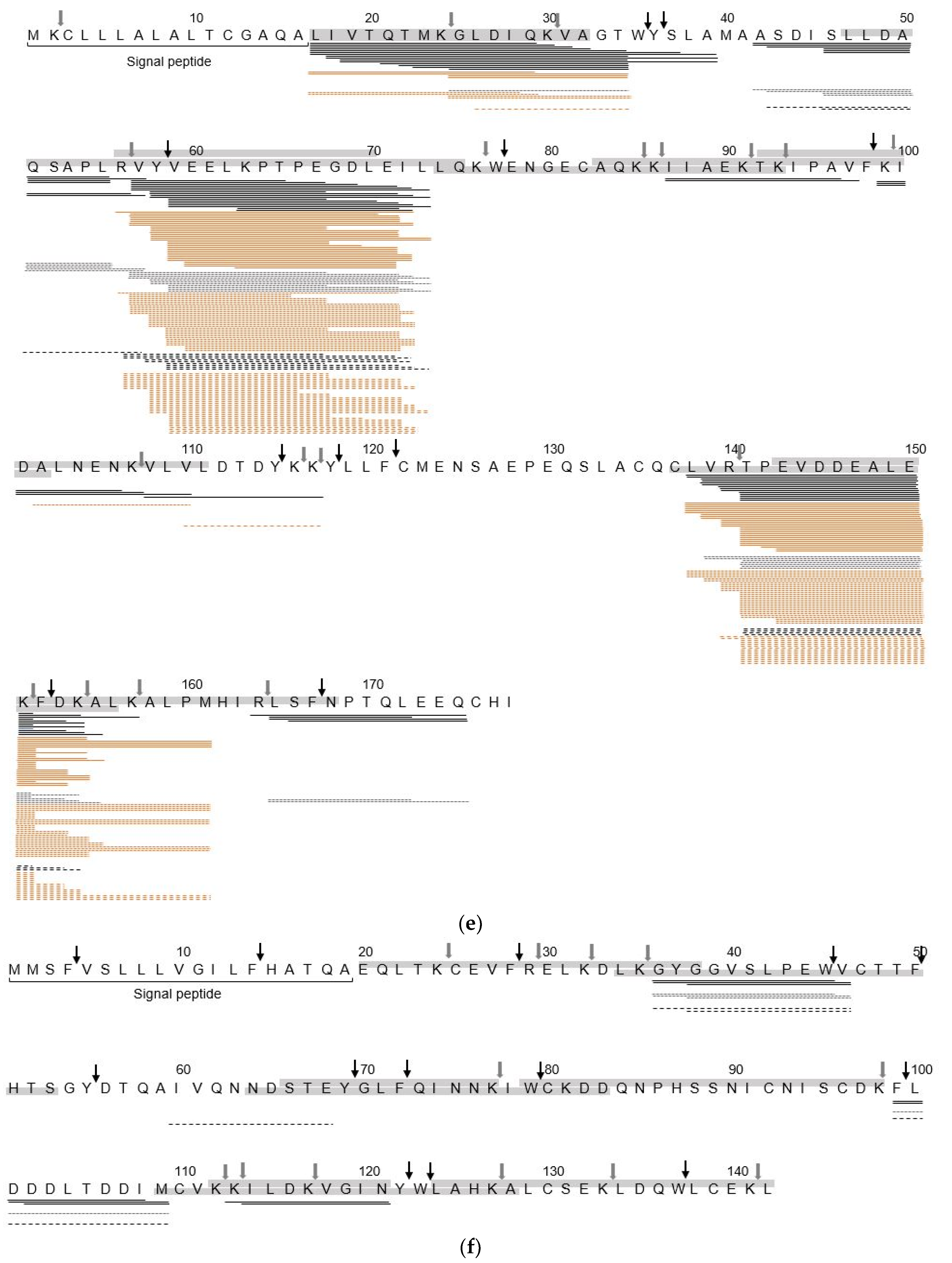
3.2. Identification of sIgE Epitopes and T-Cell Epitopes in the In Vitro Digest
3.3. Peptides Identified at the Basolateral Compartment of the Caco-2 Cell Monolayer
3.4. sIgE Binding Epitopes on the Basolateral Side of the Caco-2 Cell Monolayer
4. Discussion
4.1. Heat Treatment Dependent Differences in Peptide Profiles
4.2. Hydrolysis Resistant Areas
4.3. Effect of Heat Treatment on Identification of IgE Binding Epitopes
4.4. Identification of Peptides on the Basolateral Side of the Caco-2 Cell Monolayer
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, J.S.; Nowak-Wgrzyn, A.; Sicherer, S.H.; Noone, S.; Moshier, E.L.; Sampson, H.A. Dietary baked milk accelerates the resolution of cow’s milk allergy in children. J. Allergy Clin. Immunol. 2011, 128, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Boekel, M.A.J.S. Effect of heating on Maillard reactions in milk. Food Chem. 1998, 62, 403–414. [Google Scholar] [CrossRef]
- Arena, S.; Renzone, G.; D’Ambrosio, C.; Salzano, A.M.; Scaloni, A. Dairy products and the Maillard reaction: A promising future for extensive food characterization by integrated proteomics studies. Food Chem. 2017, 219, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Birlouez-Aragon, I.; Pischetsrieder, M.; Leclère, J.; Morales, F.J.; Hasenkopf, K.; Kientsch-Engel, R.; Ducauze, C.J.; Rutledge, D. Assessment of protein glycation markers in infant formulas. Food Chem. 2004, 87, 253–259. [Google Scholar] [CrossRef]
- Pischetsrieder, M.; Henle, T. Glycation products in infant formulas: Chemical, analytical and physiological aspects. Amino Acids 2012, 42, 1111–1118. [Google Scholar] [CrossRef]
- Dupont, D.; Mandalari, G.; Mollé, D.; Jardin, J.; Rolet-Répécaud, O.; Duboz, G.; Léonil, J.; Mills, C.E.N.; Mackie, A.R. Food processing increases casein resistance to simulated infant digestion. Mol. Nutr. Food Res. 2010, 54, 1677–1689. [Google Scholar] [CrossRef]
- Dupont, D.; Boutrou, R.; Menard, O.; Jardin, J.; Tanguy, G.; Schuck, P.; Haab, B.B.; Leonil, J. Heat treatment of milk during powder manufacture increases casein resistance to simulated infant digestion. Food Dig. 2010, 1, 28–39. [Google Scholar] [CrossRef]
- Sánchez-Rivera, L.; Ménard, O.; Recio, I.; Dupont, D. Peptide mapping during dynamic gastric digestion of heated and unheated skimmed milk powder. Food Res. Int. 2015, 77, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Kopf-Bolanz, K.A.; Schwander, F.; Gijs, M.; Vergères, G.; Portmann, R.; Egger, L. Impact of milk processing on the generation of peptides during digestion. Int. Dairy J. 2014, 35, 130–138. [Google Scholar] [CrossRef]
- Torcello-Gómez, A.; Dupont, D.; Jardin, J.; Briard-Bion, V.; Deglaire, A.; Risse, K.; Mechoulan, E.; Mackie, A. The pattern of peptides released from dairy and egg proteins is highly dependent on the simulated digestion scenario. Food Funct. 2020. [Google Scholar] [CrossRef]
- Wada, Y.; Phinney, B.S.; Weber, D.; Lönnerdal, B. In vivo digestomics of milk proteins in human milk and infant formula using a suckling rat pup model. Peptides 2017, 88, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Zenker, H.E.; Van Lieshout, G.A.A.; Van Gool, M.P.; Bragt, M.C.E.; Hettinga, K.A. Lysine blockage of milk proteins in infant formula impairs overall protein digestibility and peptide release. Food Funct. 2020, 11, 358–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Li, L.; Le, T.T.; Larsen, L.B.; Xu, D.; Jiao, W.; Sheng, B.; Li, B.; Zhang, X. Digestibility of glycated milk proteins and the peptidomics of their in vitro digests. J. Sci. Food Agric. 2019, 99, 3069–3077. [Google Scholar] [CrossRef] [PubMed]
- Teodorowicz, M.; Van Neerven, J.; Savelkoul, H. Food processing: The influence of the maillard reaction on immunogenicity and allergenicity of food proteins. Nutrients 2017, 9, 835. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Shi, J.; Yao, M.; Jiang, M.; Luo, Y. Effects of heat treatment on the antigenicity of four milk proteins in milk protein concentrates. Food Agric. Immunol. 2016, 27, 401–413. [Google Scholar] [CrossRef]
- Ehn, B.M.; Ekstrand, B.; Bengtsson, U.; Ahlstedt, S. Modification of IgE Binding during Heat Processing of the Cow’s Milk Allergen β-Lactoglobulin. J. Agric. Food Chem. 2004, 52, 1398–1403. [Google Scholar] [CrossRef]
- Taheri-Kafrani, A.; Gaudin, J.C.; Rabesona, H.; Nioi, C.; Agarwal, D.; Drouet, M.; Chobert, J.M.; Bordbar, A.K.; Haertle, T. Effects of heating and glycation of β-lactoglobulin on its recognition by ige of sera from cow milk allergy patients. J. Agric. Food Chem. 2009, 57, 4974–4982. [Google Scholar] [CrossRef]
- Nowak-Wegrzyn, A.; Fiocchi, A. Rare, medium, or well done? The effect of heating and food matrix on food protein allergenicity. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 234–237. [Google Scholar] [CrossRef]
- Corzo-Martínez, M.; Soria, A.C.; Belloque, J.; Villamiel, M.; Moreno, F.J. Effect of glycation on the gastrointestinal digestibility and immunoreactivity of bovine β-lactoglobulin. Int. Dairy J. 2010, 20, 742–752. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Rai, V.; Singer, D.; Chabierski, S.; Xie, J.; Reverdatto, S.; Burz, D.S.; Schmidt, A.M.; Hoffmann, R.; Shekhtman, A. Advanced glycation end product recognition by the receptor for AGEs. Structure 2011, 19, 722–732. [Google Scholar] [CrossRef] [Green Version]
- Heilmann, M.; Wellner, A.; Gadermaier, G.; Ilchmann, A.; Briza, P.; Krause, M.; Nagai, R.; Burgdorf, S.; Scheurer, S.; Vieths, S.; et al. Ovalbumin modified with pyrraline, a maillard reaction product, shows enhanced T-cell immunogenicity. J. Biol. Chem. 2014, 289, 7919–7928. [Google Scholar] [CrossRef] [Green Version]
- Zenker, H.E.; Teodorowicz, M.; Ewaz, A.; van Neerven, R.J.J.; Savelkoul, H.F.J.; De Jong, N.W.; Wichers, H.J.; Hettinga, K.A. Binding of CML-Modified as Well as Heat-Glycated β-lactoglobulin to Receptors for AGEs is Determined by Charge and Hydrophobicity. Int. J. Mol. Sci. 2020, 21, 4567. [Google Scholar] [CrossRef] [PubMed]
- Foerster, A.; Henle, T. Glycation in food and metabolic transit of dietary AGEs (advanced glycation end-products): Studies on the urinary excretion of pyrraline. Biochem. Soc. Trans. 2003, 31, 1383–1385. [Google Scholar] [CrossRef] [PubMed]
- Roncero-Ramos, I.; Delgado-Andrade, C.; Tessier, F.J.; Niquet-Léridon, C.; Strauch, C.; Monnier, V.M.; Navarro, M.P. Metabolic transit of Nε-carboxymethyl-lysine after consumption of AGEs from bread crust. Food Funct. 2013, 4, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.; Geissler, S.; Matthes, R.; Peto, A.; Silow, C.; Brandsch, M.; Henle, T. Transport of Free and Peptide-Bound Glycated Amino Acids: Synthesis, Transepithelial Flux at Caco-2 Cell Monolayers, and Interaction with Apical Membrane Transport Proteins. ChemBioChem 2011, 12, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Palin, K.J.; Davis, S.S. Intestinal absorption of proteins and macromolecules and the immunological response. Crit. Rev. Drug Carr. Syst. 1987, 4, 197–220. [Google Scholar]
- Picariello, G.; De Cicco, M.; Nocerino, R.; Paparo, L.; Mamone, G.; Addeo, F.; Berni Canani, R. Excretion of dietary cow’s milk derived peptides into breast milk. Front. Nutr. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Li, L.; Le, T.T.; Larsen, L.B.; Su, G.; Liang, Y.; Li, B. Digestibility of Glyoxal-Glycated β-Casein and β-Lactoglobulin and Distribution of Peptide-Bound Advanced Glycation End Products in Gastrointestinal Digests. J. Agric. Food Chem. 2017, 65, 5778–5788. [Google Scholar] [CrossRef]
- Zenker, H.E.; Raupbach, J.; Boeren, S.; Wichers, H.J.; Hettinga, K.A. The effect of low vs. high temperature dry heating on solubility and digestibility of cow’s milk protein. Food Hydrocoll. 2020, 109, 106098. [Google Scholar] [CrossRef]
- Ménard, O.; Bourlieu, C.; De Oliveira, S.C.; Dellarosa, N.; Laghi, L.; Carrière, F.; Capozzi, F.; Dupont, D.; Deglaire, A. A first step towards a consensus static in vitro model for simulating full-term infant digestion. Food Chem. 2018, 240, 338–345. [Google Scholar] [CrossRef]
- Dingess, K.A.; De Waard, M.; Boeren, S.; Vervoort, J.; Lambers, T.T.; Van Goudoever, J.B.; Hettinga, K. Human milk peptides differentiate between the preterm and term infant and across varying lactational stages. Food Funct. 2017, 8, 3769–3782. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Q.; Zeng, W.F.; Fang, P.; Cao, W.Q.; Liu, C.; Yan, G.Q.; Zhang, Y.; Peng, C.; Wu, J.Q.; Zhang, X.J.; et al. PGlyco 2.0 enables precision N-glycoproteomics with comprehensive quality control and one-step mass spectrometry for intact glycopeptide identification. Nat. Commun. 2017, 8, 438. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Boggs, I.; Hine, B.; Smolenski, G.; Hettinga, K.; Zhang, L.; Wheeler, T.T. Changes in the repertoire of bovine milk proteins during mammary involution. Eupa Open Proteom. 2015, 9, 65–75. [Google Scholar] [CrossRef]
- Matsuo, H.; Yokooji, T.; Taogoshi, T. Common food allergens and their IgE-binding epitopes. Allergol. Int. 2015, 64, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Knol, E.F.; de Jong, N.W.; Ulfman, L.H.; Tiemessen, M.M. Management of cow’s milk allergy from an immunological perspective: What are the options? Nutrients 2019, 11, 2734. [Google Scholar] [CrossRef] [Green Version]
- Milkovska-Stamenova, S.; Hoffmann, R. Identification and quantification of bovine protein lactosylation sites in different milk products. J. Proteom. 2016, 134, 112–126. [Google Scholar] [CrossRef]
- Donato, L.; Guyomarc’h, F. Formation and properties of the whey protein/κ-casein complexes in heated skim milk—A review. Dairy Sci. Technol. 2009, 89, 3–29. [Google Scholar] [CrossRef]
- Alexander, L.J.; Hayes, G.; Pearse, M.J.; Stewart, A.F.; Willis, I.M.; Mackinlay, A.G. Complete sequence of the bovine β-lactoglobulin cDNA. Nucleic Acids Res. 1989, 17, 6739. [Google Scholar] [CrossRef]
- Vetri, V.; Militello, V. Thermal induced conformational changes involved in the aggregation pathways of beta-lactoglobulin. Biophys. Chem. 2005, 113, 83–91. [Google Scholar] [CrossRef]
- Van Boekel, M.A.J.S. Heat-induced deamidation, dephosphorylation and breakdown of caseinate. Int. Dairy J. 1999, 9, 237–241. [Google Scholar] [CrossRef]
- Wada, Y.; Lönnerdal, B. Effects of Industrial Heating Processes of Milk-Based Enteral Formulas on Site-Specific Protein Modifications and Their Relationship to in Vitro and in Vivo Protein Digestibility. J. Agric. Food Chem. 2015, 63, 6787–6798. [Google Scholar] [CrossRef]
- Bernard, H.; Meisel, H.; Creminon, C.; Wal, J.M. Post-translational phosphorylation affects the IgE binding capacity of caseins. Febs Lett. 2000, 467, 239–244. [Google Scholar] [CrossRef]
- Egger, L.; Ménard, O.; Baumann, C.; Duerr, D.; Schlegel, P.; Stoll, P.; Vergères, G.; Dupont, D.; Portmann, R. Digestion of milk proteins: Comparing static and dynamic in vitro digestion systems with in vivo data. Food Res. Int. 2019, 118, 32–39. [Google Scholar] [CrossRef]
- Picariello, G.; Ferranti, P.; Fierro, O.; Mamone, G.; Caira, S.; Di Luccia, A.; Monica, S.; Addeo, F. Peptides surviving the simulated gastrointestinal digestion of milk proteins: Biological and toxicological implications. J. Chromatogr. B: Anal. Technol. Biomed. Life Sci. 2010, 878, 295–308. [Google Scholar] [CrossRef]
- Gasparini, A.; Buhler, S.; Faccini, A.; Sforza, S.; Tedeschi, T. Thermally-induced lactosylation of whey proteins: Identification and synthesis of lactosylated β-lactoglobulin epitope. Molecules 2020, 25, 1294. [Google Scholar] [CrossRef] [Green Version]
- Ruiter, B.; Trégoat, V.; M’Rabet, L.; Garssen, J.; Bruijnzeel-Koomen, C.A.F.M.; Knol, E.F.; Van Hoffen, E. Characterization of T cell epitopes in αs1-casein in cow’s milk allergic, atopic and non-atopic children. Clin. Exp. Allergy 2006, 36, 303–310. [Google Scholar] [CrossRef]
- Elsayed, S.; Eriksen, J.; Øysæd, L.K.; Idsøe, R.; Hill, D.J. T cell recognition pattern of bovine milk αS1-casein and its peptides. Mol. Immunol. 2004, 41, 1225–1234. [Google Scholar] [CrossRef]
- Nakajima-Adachi, H.; Hachimura, S.; Ise, W.; Honma, K.; Nishiwaki, S.; Hirota, M.; Shimojo, N.; Katsuki, T.; Ametani, A.; Kohno, Y.; et al. Determinant analysis of IgE and IgG4 antibodies and T cells specific for bovine alpha(s)1-casein from the same patients allergic to cow’s milk: Existence of alpha(s)1-casein-specific B cells and T cells characteristic in cow’s-milk allergy. J. Allergy Clin. Immunol. 1998, 101, 660–671. [Google Scholar] [CrossRef]
- Gouw, J.W.; Jo, J.; Meulenbroek, L.A.P.M.; Heijjer, T.S.; Kremer, E.; Sandalova, E.; Knulst, A.C.; Jeurink, P.V.; Garssen, J.; Rijnierse, A.; et al. Identification of peptides with tolerogenic potential in a hydrolysed whey-based infant formula. Clin. Exp. Allergy 2018, 48, 1345–1353. [Google Scholar] [CrossRef] [Green Version]
- Ilchmann, A.; Burgdorf, S.; Scheurer, S.; Waibler, Z.; Nagai, R.; Wellner, A.; Yamamoto, Y.; Yamamoto, H.; Henle, T.; Kurts, C.; et al. Glycation of a food allergen by the Maillard reaction enhances its T-cell immunogenicity: Role of macrophage scavenger receptor class A type I and II. J. Allergy Clin. Immunol. 2010, 125, 175–183.e111. [Google Scholar] [CrossRef]
- Xu, Q.; Hong, H.; Wu, J.; Yan, X. Bioavailability of bioactive peptides derived from food proteins across the intestinal epithelial membrane: A review. Trends Food Sci. Technol. 2019, 86, 399–411. [Google Scholar] [CrossRef]
- Reitsma, M.; Westerhout, J.; Wichers, H.J.; Wortelboer, H.M.; Verhoeckx, K.C.M. Protein transport across the small intestine in food allergy. Mol. Nutr. Food Res. 2014, 58, 194–205. [Google Scholar] [CrossRef]
- Moradi, S.V.; Hussein, W.M.; Varamini, P.; Simerska, P.; Toth, I. Glycosylation, an effective synthetic strategy to improve the bioavailability of therapeutic peptides. Chem. Sci. 2016, 7, 2492–2500. [Google Scholar] [CrossRef] [Green Version]
- Varamini, P.; Mansfeld, F.M.; Blanchfield, J.T.; Wyse, B.D.; Smith, M.T.; Toth, I. Synthesis and biological evaluation of an orally active glycosylated endomorphin-1. J. Med. Chem. 2012, 55, 5859–5867. [Google Scholar] [CrossRef]
- Polt, R.; Porreca, F.; Szabò, L.Z.; Bilsky, E.J.; Davis, P.; Abbruscato, T.J.; Davis, T.P.; Horvath, R.; Yamamura, H.I.; Hruby, V.J. Glycopeptide enkephalin analogues produce analgesia in mice: Evidence for penetration of the blood-brain barrier. Proc. Natl. Acad. Sci. USA 1994, 91, 7114–7118. [Google Scholar] [CrossRef] [Green Version]
- Zenker, H.E.; Ewaz, A.; Deng, Y.; Savelkoul, H.F.J.; Van Neerven, R.J.J.; De Jong, N.; Wichers, H.J.; Hettinga, K.A.; Teodorowicz, M. Differential effects of dry vs. Wet heating of β-lactoglobulin on formation of sRAGE binding ligands and sIgE epitope recognition. Nutrients 2019, 11, 1432. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.S.; Léonil, J.; Henry, G.; Cauty, C.; Carvalho, A.F.; Bouhallab, S. Heating and glycation of β-lactoglobulin and β-casein: Aggregation and in vitro digestion. Food Res. Int. 2014, 55, 70–76. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Sample | Peptide Sequence | AAs Position | sIgE Epitope AAs Position | PTM | |
|---|---|---|---|---|---|---|
| αs1-cn | NT, LT, HT | VNELSKDIGSESTEDQ | 52–67 | 54–63 | Phos | |
| NT, LT, HT | VNELSKDIGSESTEDQAMEDIK | 52–73 | 54–63 | Phos | ||
| NT, LT, HT | KVPQLEIVPNSAEE | 120–133 | 124–135 | Phos | ||
| NT, LT, HT | KVPQLEIVPNSAEER | 120–134 | 124–135 | Phos | ||
| NT, LT | VPQLEIVPNSAEER | 121–134 | 124–135 | Phos | ||
| NT, LT, HT | LEIVPNSAEE | 124–133 | 124–135 | Phos | ||
| NT, LT, HT | LEIVPNSAEER | 124–134 | 124–135 | Phos | ||
| NT | EIVPNSAEER | 125–134 | 124–135 | Phos | ||
| NT | KEGIHAQQKEPMIGV | 139–153 | 137–147 | N/A | ||
| NT | EGIHAQQKEPMIGV | 140–153 | 141–155 | N/A | ||
| NT, HT | GTQYTDAPSFSDIPNPI | 185–201 | 186–200 | N/A | ||
| NT, LT, HT | QYTDAPSFSDIPNPI | 187–201 | 186–200 | N/A | ||
| NT | QYTDAPSFSDIPNPIGSENSEK | 187–208 | 188–209 | N/A | ||
| NT, LT, HT | TDAPSFSDIPNPIGSENSEK | 189–208 | 188–209 | N/A | ||
| NT,LT,HT | TDAPSFSDIPNPIGSENSEK | 189–208 | 188–209 | Phos | ||
| NT | TDAPSFSDIPNPIGSENSEKT | 189–209 | 188–209 | N/A | ||
| αs2-cn | NT | KNTMEHVSSSEESIISQ | 16–32 | 16–35 | Phos | |
| NT | KNTMEHVSSSEESIISQET | 16–34 | 16–35 | Phos | ||
| HT | QGPIVLNPWDQVK | 116–128 | 120–129 | N/A | ||
| β-cn | NT, LT | RELEELNVPGEIVE | 16–29 | 16–31 | N/A | |
| NT | RELEELNVPGEIVESL | 16–31 | 16–31 | Phos | * | |
| NT | ELEELNVPGEIVESL | 17–31 | 16–31 | Phos | ||
| NT | TEDELQDKIHPFA | 56–68 | 60–69 | N/A | ||
| NT, LT, HT | SLVYPFPGPIPNS | 72–84 | 70–85 | N/A | ||
| NT, LT, HT | PVVVPPFLQPE | 96–106 | 98–107 | N/A | ||
| NT, LT, HT | PVVVPPFLQPE | 96–107 | 98–107 | N/A | ||
| NT, LT, HT | PVVVPPFLQPEVMG | 96–109 | 98–107 | N/A | ||
| LT, HT | PVVVPPFLQPEVMGV | 96–110 | 98–107 | N/A | ||
| NT, LT, HT | VVPPFLQPE | 98–106 | 98–107 | N/A | ||
| NT, LT, HT | VVPPFLQPEV | 98–107 | 98–107 | N/A | * | |
| NT, LT, HT | EMPFPKYPVEPF | 123–134 | 122–135 | N/A | ||
| NT, LT | QPLPPTVMFPPQS | 164–176 | 164–179 | N/A | ||
| NT, LT, HT | QPLPPTVMFPPQSV | 164–177 | 164–179 | N/A | ||
| κ-cn | NT | KNQDKTEIPTINT | 133–145 | 132–147 | N/A | |
| β-lg | NT | LIVTQTMKGLDIQ | 17–29 | 17–32 | N/A | |
| NT | LIVTQTMKGLDIQKV | 17–31 | 17–32 | N/A | ||
| NT | LIVTQTMKGLDIQKVA | 17–32 | 17–32 | N/A | ||
| NT | LIVTQTMKGLDIQKVAGT | 17–34 | 17–32 | N/A | ||
| NT | LIVTQTMKGLDIQKVAGTWYS | 17–37 | 17–32 | N/A | ||
| NT | LIVTQTMKGLDIQKVAGTWYSLA | 17–39 | 17–32 | N/A | ||
| NT | IVTQTMKGLDIQKVAGT | 18–34 | 17–32 | N/A | ||
| NT | IVTQTMKGLDIQKVAGTWYSLA | 18–39 | 17–32 | N/A | ||
| NT | VTQTMKGLDIQKVAGT | 19–34 | 17–32 | N/A | ||
| NT | VTQTMKGLDIQKVAGTWYSLA | 19–39 | 17–32 | N/A | ||
| NT, LT, HT | VYVEELKPTPEGDLE | 57–71 | 56–70 | N/A | ||
| NT, LT, HT | VYVEELKPTPEGDLEI | 57–72 | 56–70 | N/A | ||
| NT, LT | VYVEELKPTPEGDLEIL | 57–73 | 56–70 | N/A | ||
| NT, LT, HT | YVEELKPTPEGDLE | 58–71 | 56–70 | N/A | ||
| NT, LT, HT | YVEELKPTPEGDLEI | 58–72 | 56–70 | N/A | ||
| NT, LT | YVEELKPTPEGDLEIL | 58–73 | 56–70 | N/A | ||
| NT | LVRTPEVDDEALEK | 138–151 | 136–150 | N/A | ||
| NT | LVRTPEVDDEALEKFDK | 138–154 | 137–156 | N/A | ||
| α-lac | NT | KILDKVGIN | 113–121 | 112–121 | N/A |
| Protein | Sample | Peptide Sequence | AAs Position | sIgE Epitope AAs Position | PTM |
|---|---|---|---|---|---|
| αs1-cn | NT, LT | EGIHAQQKEPMIGV | 140–153 | 141–155 | Lac |
| HT | TDAPSFSDIPNPIGSENSEK | 189–208 | 188–209 | Lac | |
| HT | TDAPSFSDIPNPIGSENSEKTTMPL | 189–213 | 188–209 | Gluc | |
| HT | TDAPSFSDIPNPIGSENSEKTTMPL | 189–213 | 188–209 | Lac | |
| NT, LT | EGIHAQQKEPMIGV | 140–153 | 141–155 | Lac | |
| αs2-cn | LT, HT | LYQGPIVLNPWDQVK | 114–128 | 120–129 | Lac |
| LT, HT | LYQGPIVLNPWDQVK | 114–128 | 120–129 | Gluc | |
| LT, HT | LYQGPIVLNPWDQVK | 114–128 | 120–129 | CML | |
| LT, HT | LYQGPIVLNPWDQVK | 114–128 | 120–129 | Pyr | |
| LT, HT | YQGPIVLNPWDQVK | 115–128 | 120–129 | Lac | |
| LT, HT | YQGPIVLNPWDQVK | 115–128 | 120–129 | Gluc | |
| LT, HT | YQGPIVLNPWDQVK | 115–128 | 120–129 | CML | |
| HT | QGPIVLNPWDQVK | 116–128 | 120–129 | Lac | |
| HT | QGPIVLNPWDQVK | 116–128 | 120–129 | Gluc | |
| HT | QGPIVLNPWDQVK | 116–128 | 120–129 | CML | |
| HT | QGPIVLNPWDQVK | 116–128 | 120–129 | Pyr | |
| β-cn | LT, HT | EMPFPKYPVEPF | 123–134 | 122–135 | Lac |
| LT, HT | EMPFPKYPVEPF | 123–134 | 122–135 | Gluc | |
| HT | SLSQSK(1)VLPVPQK(1)AVPYPQ | 179–197 | 182–199 | Lac | |
| β-lg | LT, HT | LIVTQTMKGLDIQ | 17–29 | 17–32 | Lac |
| NT | LIVTQTMK(1)GLDIQK(1)VAGT | 17–34 | 17–32 | Lac | |
| LT | RVYVEELKPTPEGDLE | 56–71 | 56–70 | Lac | |
| NT | RVYVEELKPTPEGDLEI | 56–72 | 56–70 | Lac | |
| NT | VYVEELKPTPEGDLE | 57–70 | 56–70 | Lac | |
| NT, LT, HT | VYVEELKPTPEGDLE | 57–71 | 56–70 | Lac | |
| NT, LT, HT | VYVEELKPTPEGDLE | 57–71 | 56–70 | Gluc | |
| NT, LT, HT | VYVEELKPTPEGDLE | 57–71 | 56–70 | CML | |
| NT, LT, HT | VYVEELKPTPEGDLE | 57–71 | 56–70 | Pyr | |
| NT, LT, HT | VYVEELKPTPEGDLEI | 57–72 | 56–70 | Lac | |
| NT, LT, HT | VYVEELKPTPEGDLEI | 57–72 | 56–70 | Gluc | |
| NT, LT, HT | YVEELKPTPEGDLE | 58–71 | 56–70 | Lac | |
| NT, LT, HT | YVEELKPTPEGDLE | 58–71 | 56–70 | Gluc | |
| NT, LT, HT | YVEELKPTPEGDLE | 58–71 | 56–70 | CML | |
| NT, LT, HT | YVEELKPTPEGDLE | 58–71 | 56–70 | Pyr | |
| LT, HT | YVEELKPTPEGDLEI | 58–72 | 56–70 | Lac | |
| LT, HT | YVEELKPTPEGDLEI | 58–72 | 56–70 | Gluc | |
| LT, HT | YVEELKPTPEGDLEI | 58–72 | 56–70 | CML | |
| NT, HT | YVEELKPTPEGDLEIL | 58–73 | 56–70 | Lac | |
| NT | LVRTPEVDDEALEK(1)FDK(1) | 138–154 | 137–156 | Lac | |
| NT | LVRTPEVDDEALEK(1)FDK(1) | 138–154 | 137–156 | Pyr | |
| NT, LT | LVRTPEVDDEALEK(1)FDK(1)ALK(1)ALPM | 138–161 | 137–156 | Lac | |
| NT, LT | LVRTPEVDDEALEKFDK(0.8)ALK(0.2)ALPM | 138–161 | 137–156 | Gluc | |
| NT, LT | LVRTPEVDDEALEK(1)FDKALKALPM | 138–161 | 137–156 | CML | |
| NT, LT | LVRTPEVDDEALEK(1)FDK(1)ALKALPM | 138–161 | 137–156 | Pyr |
| Protein | Sample | Sequence | HLA-Allele | AAs Position | PTM | Perc. Rank |
|---|---|---|---|---|---|---|
| αs1-cn | NT, LT | EAESISSSEEIVPNSVEQ | HLA-DQA1*03:01/DQB1*03:02; | 76–93 | Phos | 2.5 |
| HLA-DQA1*04:01/DQB1*04:02 | 5.3 | |||||
| NT, LT | YKVPQLEIVPNSAEE | HLA-DRB1*04:05; | 119–133 | Phos | 1.9 | |
| HLA-DQA1*04:01/DQB1*04:02; | 4.1 | |||||
| HLA-DQA1*03:01/DQB1*03:02 | 5.8 | |||||
| NT, LT, HT | YKVPQLEIVPNSAEER | HLA-DRB1*04:05; | 119–134 | Phos | 2.2 | |
| HLA-DQA1*04:01/DQB1*04:02; | 5.9 | |||||
| HLA-DQA1*03:01/DQB1*03:02 | 6.5 | |||||
| NT, LT, HT | KVPQLEIVPNSAEER | HLA-DRB1*04:05; | 120–134 | Phos | 1.9 | |
| HLA-DQA1*04:01/DQB1*04:02; | 5.6 | |||||
| HLA-DQA1*03:01/DQB1*03:02 | 5.9 | |||||
| αs2-cn | NT, LT, HT | SIGSSSEESAEVATEEV | HLA-DQA1*04:01/DQB1*04:02; | 68–84 | n.a. | 0.14 |
| HLA-DQA1*03:01/DQB1*03:02 | 0.18 | |||||
| LT, HT | LYQGPIVLNPWDQVK | HLA-DRB1*13:02 | 114–128 | Gluc | 9.7 | |
| LT, HT | LYQGPIVLNPWDQVK | HLA-DRB1*13:02 | 114–128 | Lac | 9.7 | |
| LT, HT | LYQGPIVLNPWDQVK | HLA-DRB1*13:02 | 114–128 | CML | 9.7 | |
| LT, HT | LYQGPIVLNPWDQVK | HLA-DRB1*13:02 | 114–128 | Pyr | 9.7 | |
| β-cn | HT | SLTLTDVENLHLPLP | HLA-DPA1*03:01/DPB1*04:02 | 139–153 | N/A | 6.3 |
| β-lg | NT | VTQTMKGLDIQKVAGT | HLA-DRB4*01:01 | 19–34 | N/A | 7.9 |
| NT | ASDISLLDAQSAPLRV | HLA-DRB4*01:01; | 42–57 | N/A | 4.0; | |
| HLA-DRB1*01:01; | 6.6; | |||||
| HLA-DRB1*13:02; | 7.9; | |||||
| HLA-DRB1*12:01 | 8.2; | |||||
| HLA-DQA1*03:01/DQB1*03:02 | 9 | |||||
| HT | SDISLLDAQSAPLRV | HLA-DRB4*01:01; | 43–57 | N/A | 3.3 | |
| HLA-DRB1*01:01; | 4.4 | |||||
| HLA-DRB1*12:01; | 6.2 | |||||
| HLA-DRB1*13:02; | 6.3 | |||||
| HLA-DRB1*09:01 | 7.2 |
| Protein | Sample | Peptide Sequence | AAs Position | sIgE Epitope AAs Position | PTM | |
|---|---|---|---|---|---|---|
| αs1-cn | NT, LT | VNELSKDIGSESTEDQ | 52–67 | 54–63 | N/A | |
| NT, LT | KVPQLEIVPNSAEE | 120–133 | 124–135 | N/A | ||
| NT, LT | QLEIVPNSAEE | 123–133 | 124–135 | N/A | ||
| NT, LT, HT | LEIVPNSAEER | 124–134 | 124–135 | N/A | ||
| β-cn | NT, LT | PVVVPPFLQPEV | 96–107 | 98–107 | N/A | |
| NT, LT, HT | VVPPFLQPE | 98–106 | 98–107 | N/A | ||
| NT, LT, HT | VVPPFLQPEV | 98–107 | 98–107 | N/A | * | |
| β-lg | NT, LT, HT | VYVEELKPTPEGDLE | 57–71 | 56–70 | N/A | |
| NT | VYVEELKPTPEGDLE | 57–71 | 56–70 | CML | ||
| NT | VYVEELKPTPEGDLE | 57–71 | 56–70 | Lac | ||
| NT | VYVEELKPTPEGDLE | 57–71 | 56–70 | Pyr | ||
| NT, HT | YVEELKPTPEGDLE | 58–71 | 56–70 | N/A | ||
| NT, LT, HT | YVEELKPTPEGDLE | 58–71 | 56–70 | CML | ||
| NT, LT, HT | YVEELKPTPEGDLE | 58–71 | 56–70 | Lac | ||
| NT, LT, HT | YVEELKPTPEGDLE | 58–71 | 56–70 | Pyr |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zenker, H.E.; Wichers, H.J.; Tomassen, M.M.M.; Boeren, S.; De Jong, N.W.; Hettinga, K.A. Peptide Release after Simulated Infant In Vitro Digestion of Dry Heated Cow’s Milk Protein and Transport of Potentially Immunoreactive Peptides across the Caco-2 Cell Monolayer. Nutrients 2020, 12, 2483. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082483
Zenker HE, Wichers HJ, Tomassen MMM, Boeren S, De Jong NW, Hettinga KA. Peptide Release after Simulated Infant In Vitro Digestion of Dry Heated Cow’s Milk Protein and Transport of Potentially Immunoreactive Peptides across the Caco-2 Cell Monolayer. Nutrients. 2020; 12(8):2483. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082483
Chicago/Turabian StyleZenker, Hannah E., Harry J. Wichers, Monic M. M. Tomassen, Sjef Boeren, Nicolette W. De Jong, and Kasper A. Hettinga. 2020. "Peptide Release after Simulated Infant In Vitro Digestion of Dry Heated Cow’s Milk Protein and Transport of Potentially Immunoreactive Peptides across the Caco-2 Cell Monolayer" Nutrients 12, no. 8: 2483. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082483