Effects of Late-Life Caloric Restriction on Age-Related Alterations in the Rat Cortex and Hippocampus

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Caloric Restriction
2.3. Cortex Cholesterol Content
2.4. Total lysate and Membrane Preparation for Western Blot Analysis
2.5. Immunoblotting
2.6. Statistical Analyses
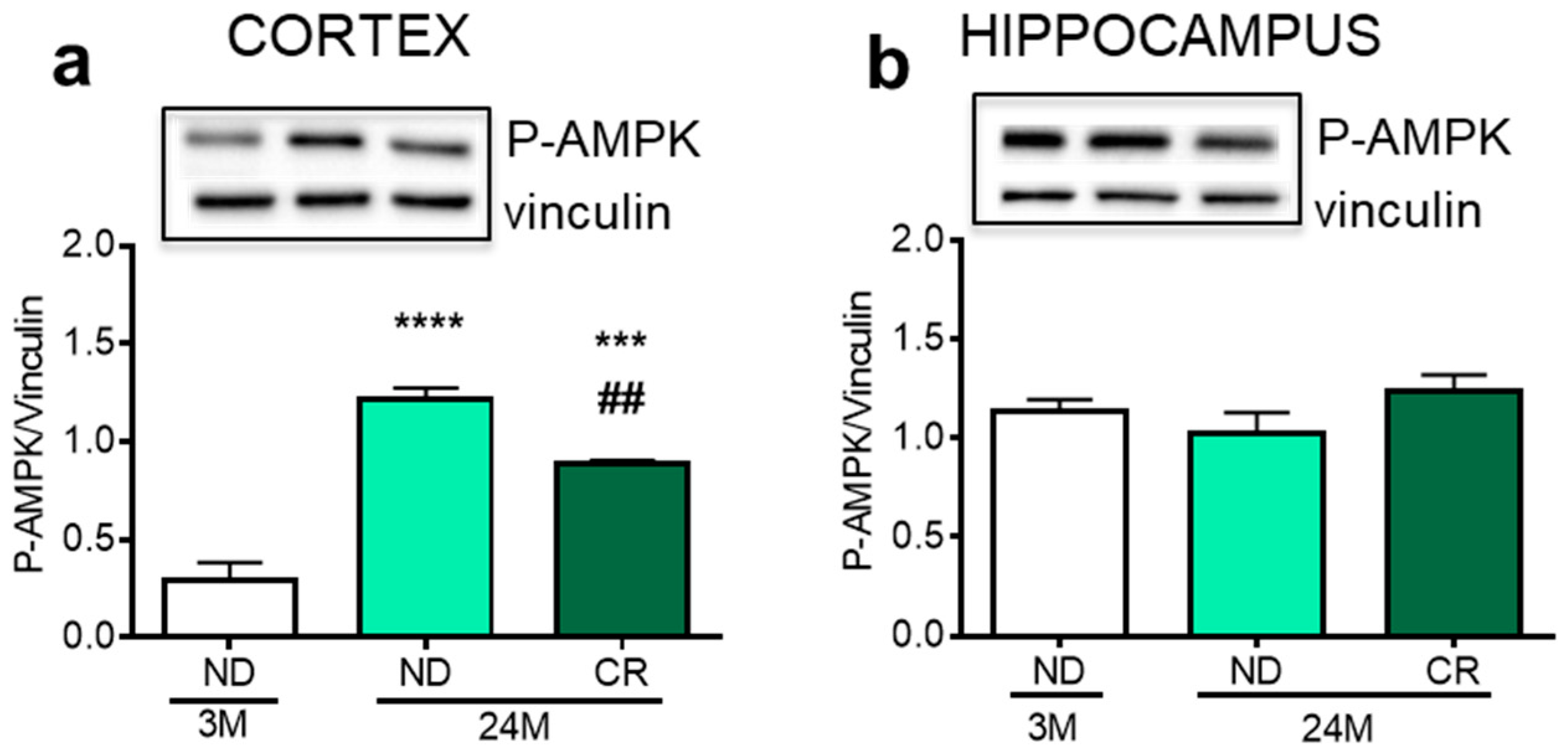
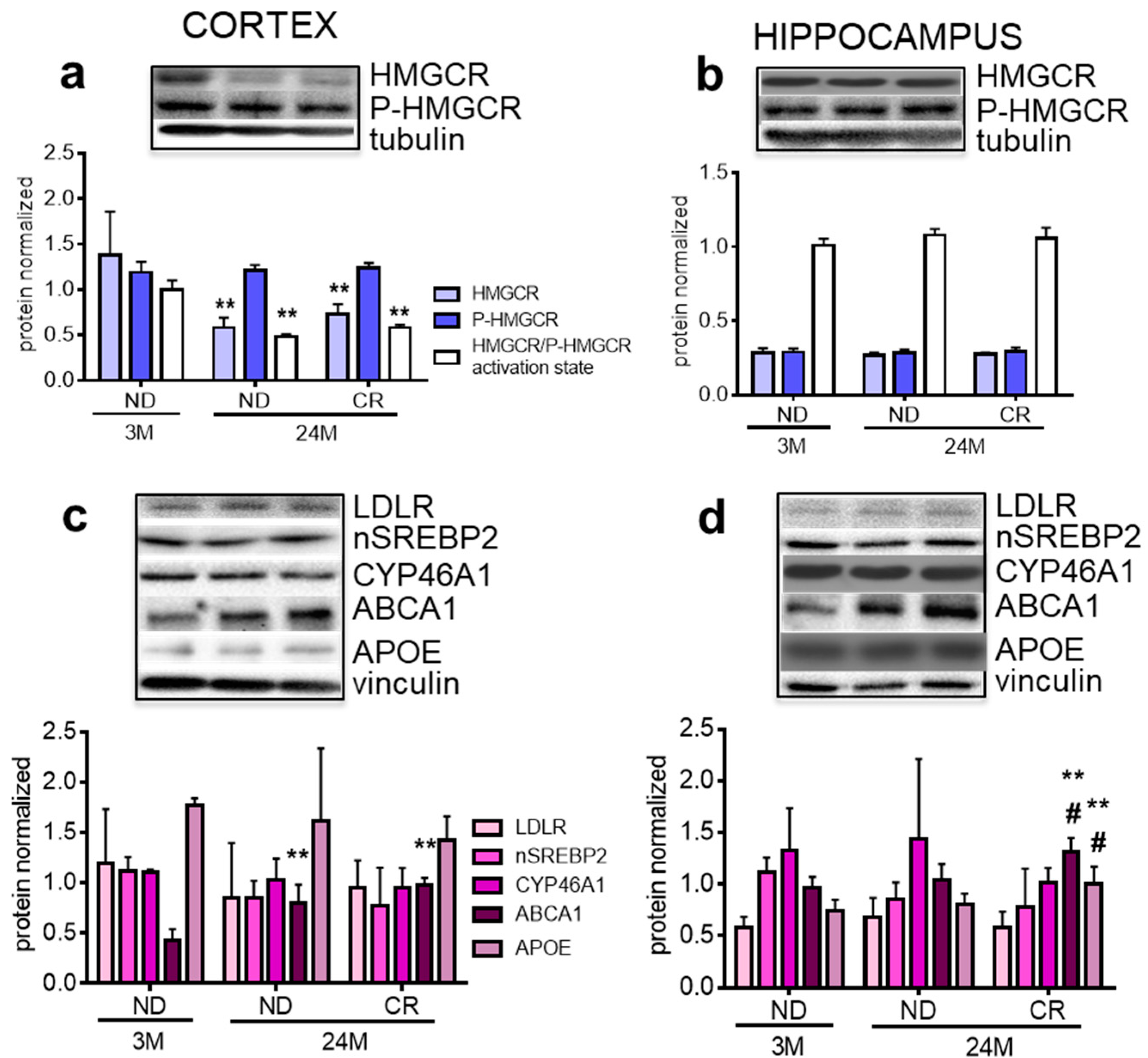
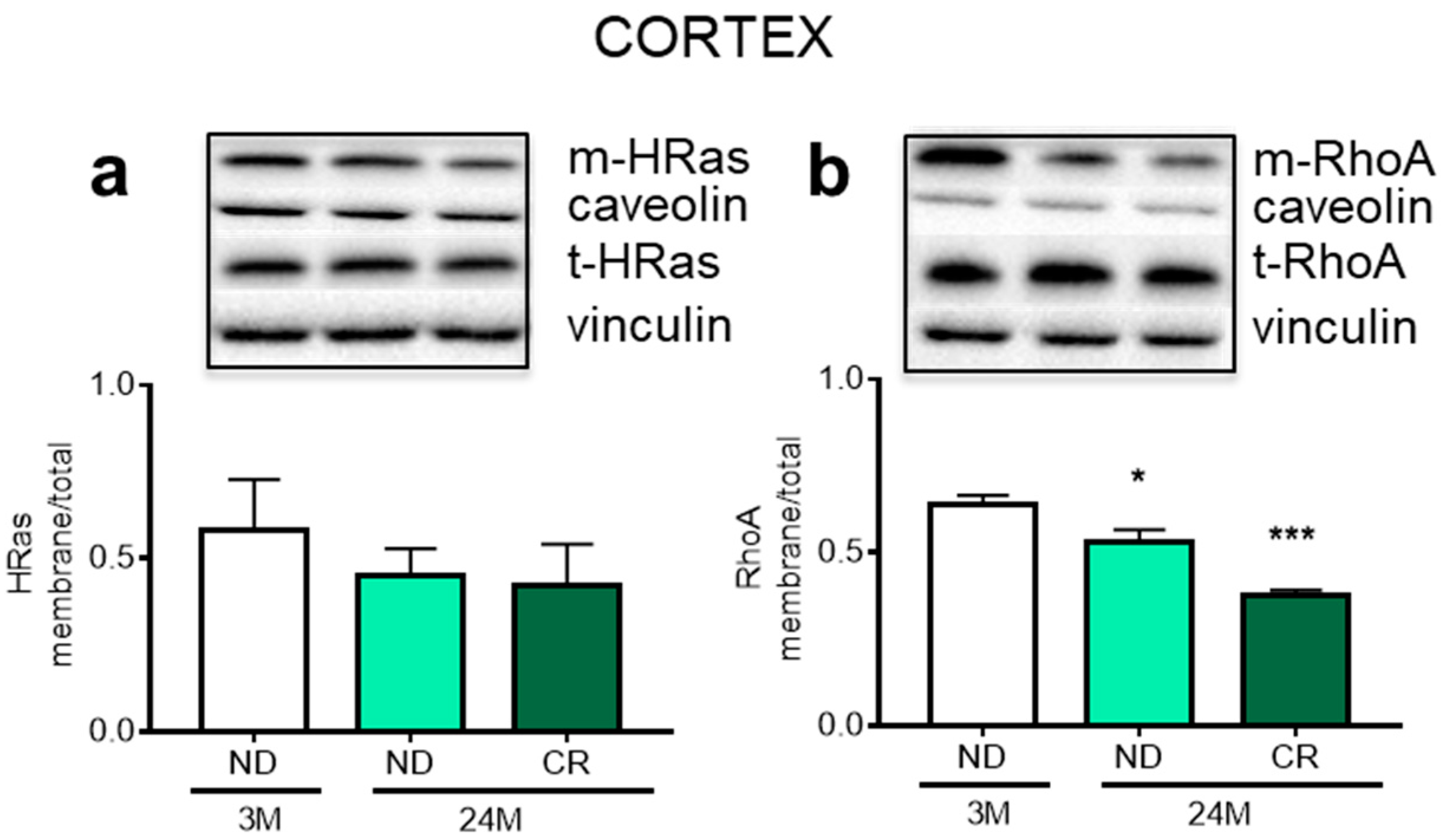
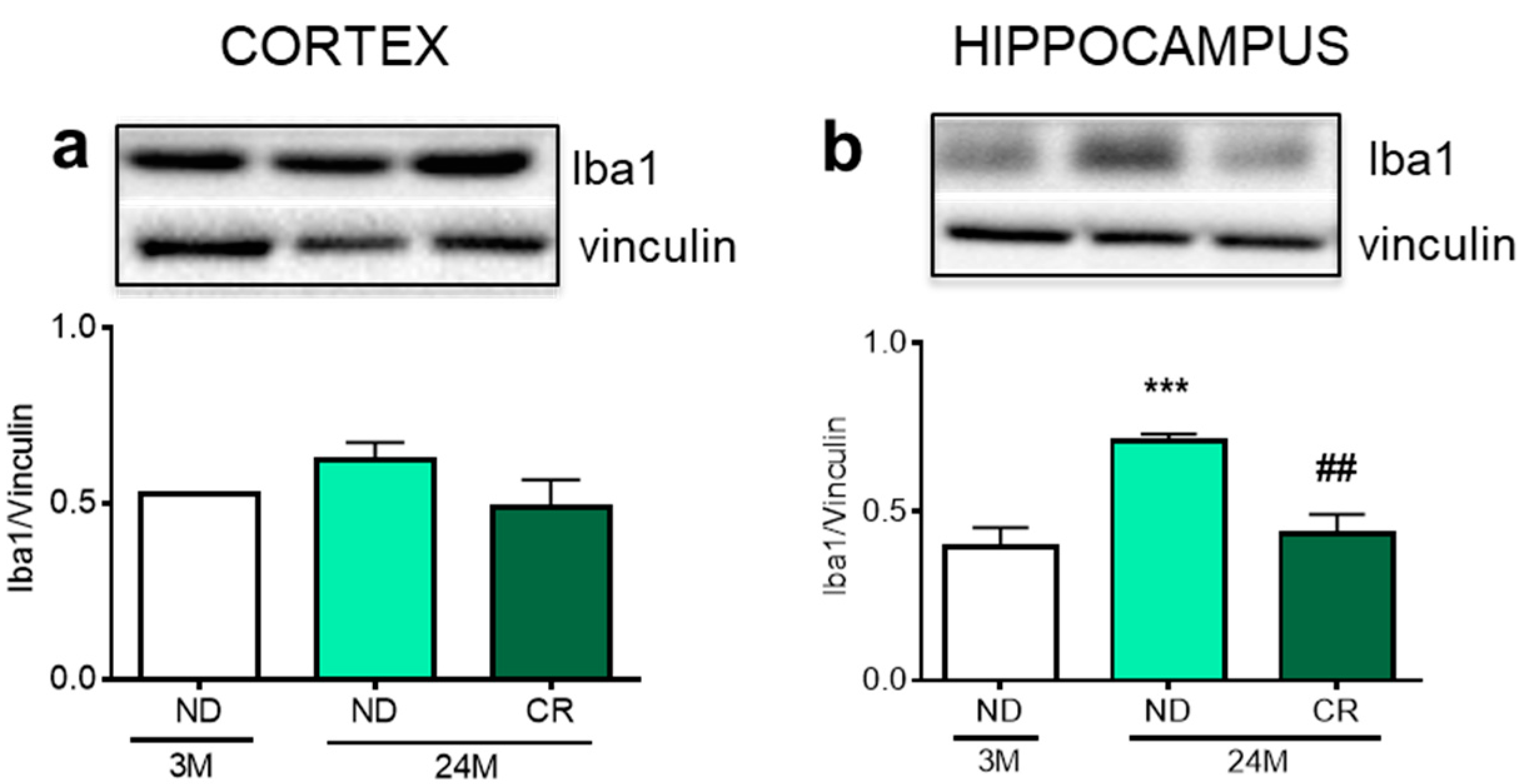
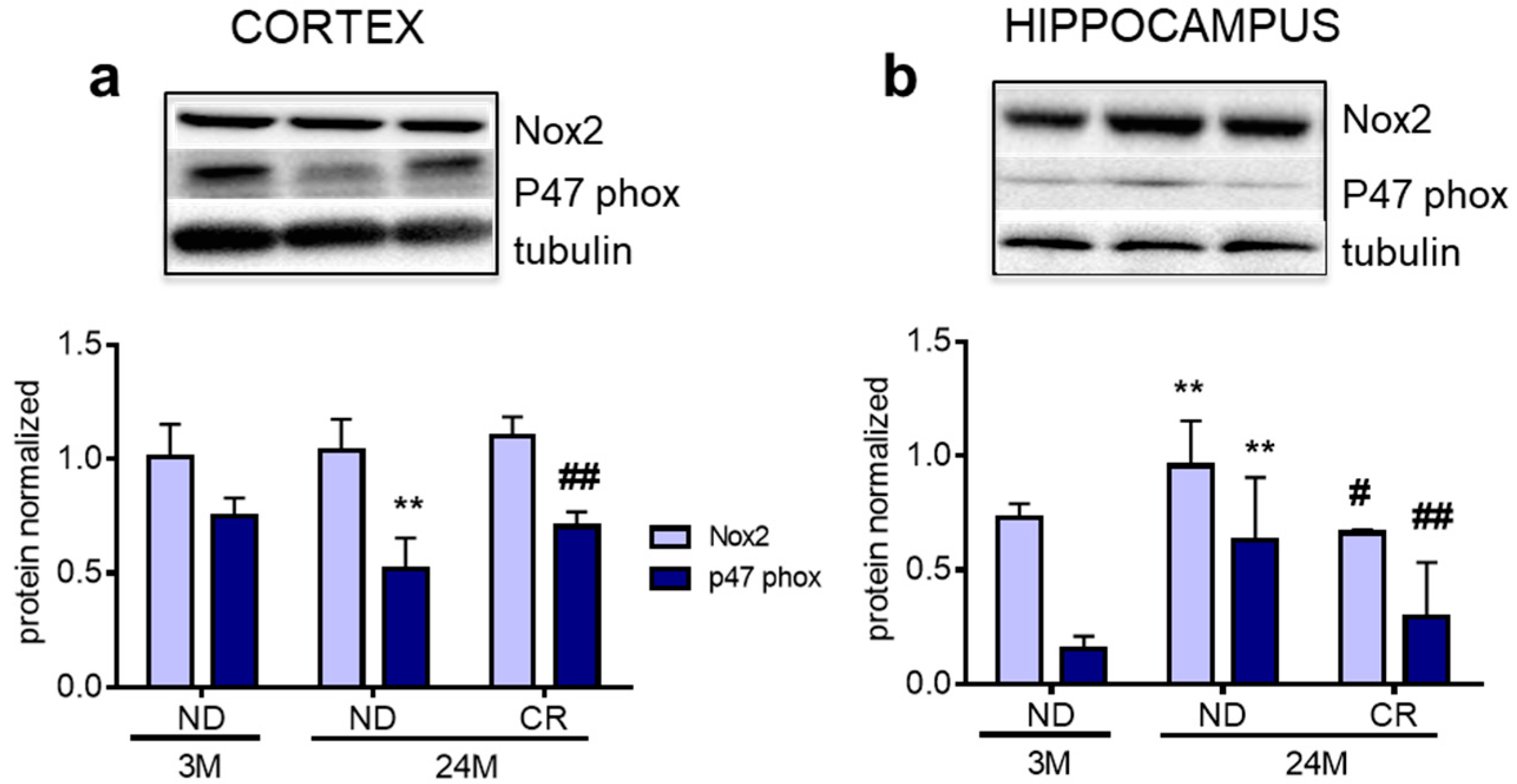
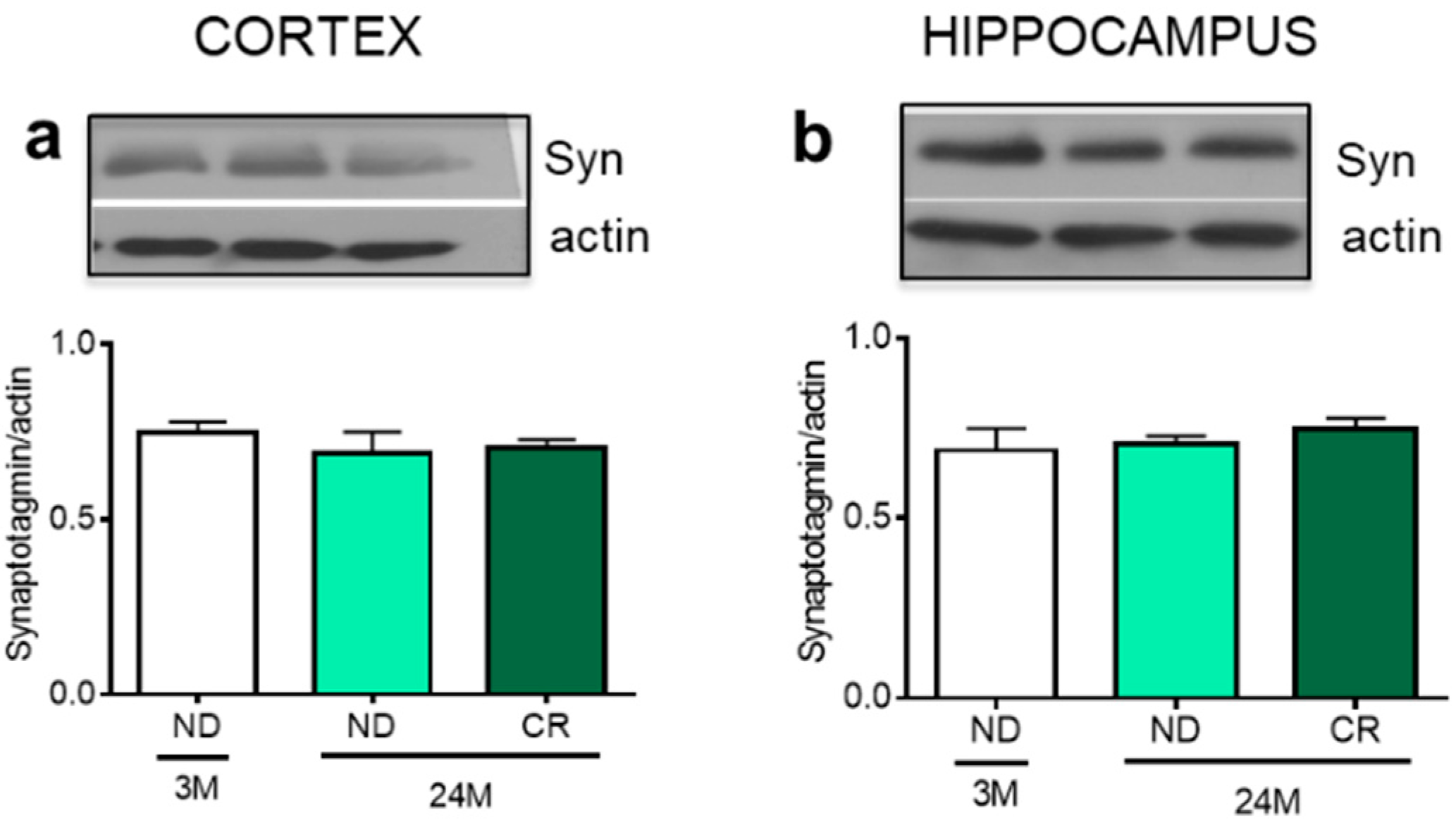
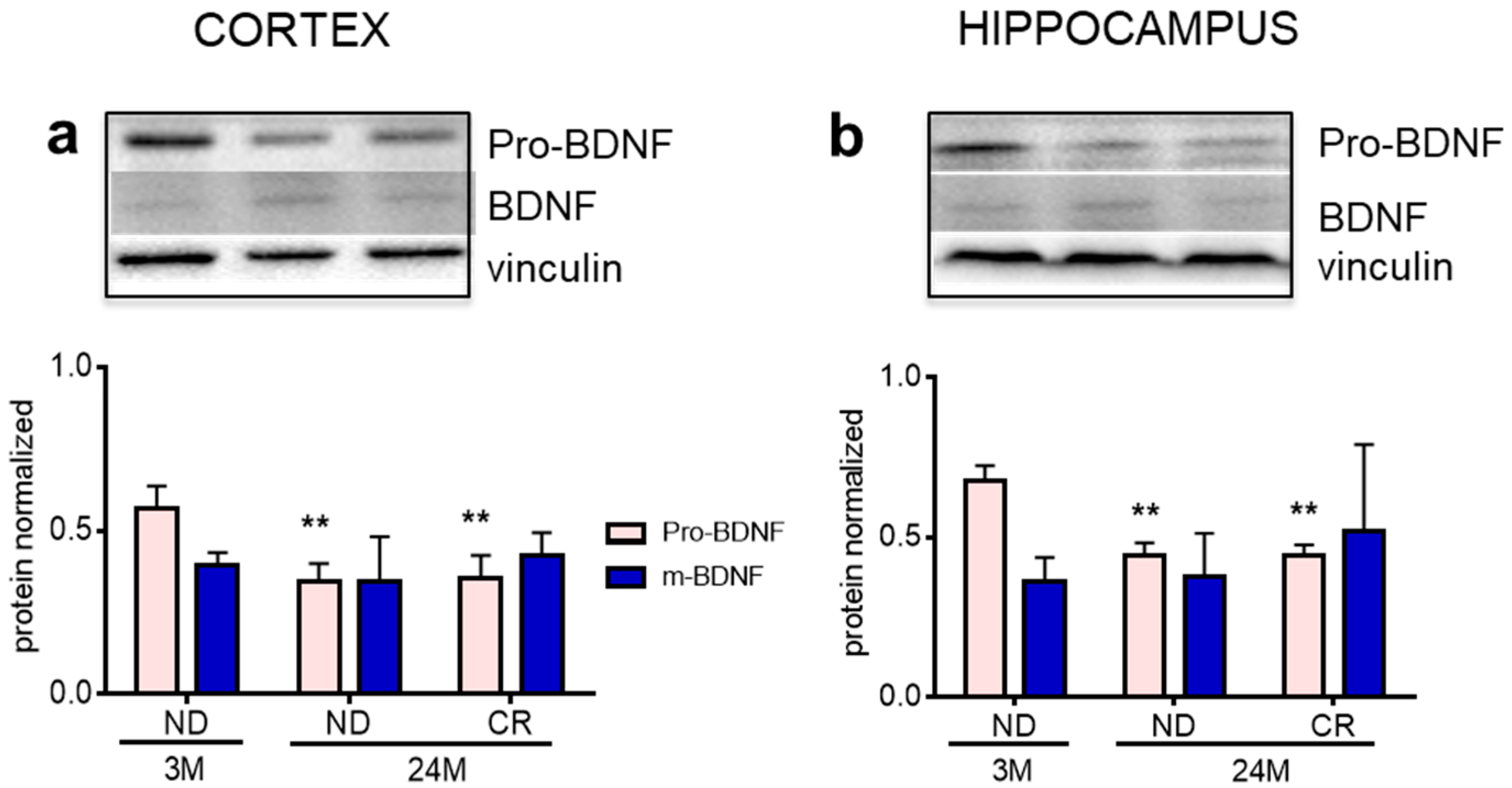
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Masoro, E.J.; Bertrand, H.; Liepa, G.; Yu, B.P. Analysis and Exploration of Age-Related Changes in Mammalian Structure and Function. Fed. Proc. 1979, 38, 1956–1961. [Google Scholar]
- Segatto, M.; Trapani, L.; Marino, M.; Pallottini, V. Age- and Sex-Related Differences in Extra-Hepatic Low-Density Lipoprotein Receptor. J. Cell. Physiol. 2011, 226, 2610–2616. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smiljanic, K.; Pesic, V.; Mladenovic Djordjevic, A.; Pavkovic, Z.; Brkic, M.; Ruzdijic, S.; Kanazir, S. Long-Term Dietary Restriction Differentially Affects the Expression of BDNF and Its Receptors in the Cortex and Hippocampus of Middle-Aged and Aged Male Rats. Biogerontology 2015, 16, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Trapani, L.; Pallottini, V. Age-Related Hypercholesterolemia and HMG-CoA Reductase Dysregulation: Sex Does Matter (A Gender Perspective). Curr. Gerontol. Geriatr. Res. 2010, 420139. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, D.; Magliozzi, R.; Mitsikostas, D.D.; Gorgoulis, V.G.; Nicholas, R.S. Aging, Cellular Senescence, and Progressive Multiple Sclerosis. Front. Cell. Neurosci. 2020, 14, 178. [Google Scholar] [CrossRef]
- Bruunsgaard, H.; Pedersen, M.; Pedersen, B.K. Aging and Proinflammatory Cytokines. Curr. Opin. Hematol. 2001, 8, 131–136. [Google Scholar] [CrossRef]
- Balietti, M.; Fattorini, G.; Pugliese, A.; Marcotulli, D.; Bragina, L.; Conti, F. Two Behavioral Tests Allow a Better Correlation Between Cognitive Function and Expression of Synaptic Proteins. Front. Aging Neurosci. 2018, 10, 91. [Google Scholar] [CrossRef]
- Erickson, K.I.; Prakash, R.S.; Voss, M.W.; Chaddock, L.; Heo, S.; McLaren, M.; Pence, B.D.; Martin, S.A.; Vieira, V.J.; Woods, J.A.; et al. Brain-Derived Neurotrophic Factor Is Associated with Age-Related Decline in Hippocampal Volume. J. Neurosci. 2010, 30, 5368–5375. [Google Scholar] [CrossRef] [Green Version]
- McCay, C.M.; Crowell, M.F.; Maynard, L.A. The Effect of Retarded Growth Upon the Length of Life Span and Upon the Ultimate Body Size. J. Nutr. 1935, 10, 63–79. [Google Scholar] [CrossRef]
- Masoro, E.J. Overview of Caloric Restriction and Ageing. Mech. Ageing Dev. 2005, 126, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Sun, S.; Geng, L.; Song, M.; Wang, W.; Ye, Y.; Ji, Q.; Zou, Z.; Wang, S.; He, X.; et al. Caloric Restriction Reprograms the Single-Cell Transcriptional Landscape of Rattus Norvegicus Aging. Cell 2020, 180, 984–1001. [Google Scholar] [CrossRef] [PubMed]
- Katewa, S.D.; Kapahi, P. Dietary Restriction and Aging, 2009. Aging Cell 2010, 9, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Martini, C.; Pallottini, V.; Cavallini, G.; Donati, A.; Bergamini, E.; Trentalance, A. Caloric Restrictions Affect Some Factors Involved in Age-Related Hypercholesterolemia. J. Cell. Biochem. 2007, 101, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J.; Kapahi, P.; Lithgow, G.J.; Melov, S.; Newman, J.C.; Verdin, E. From Discoveries in Ageing Research to Therapeutics for Healthy Ageing. Nature 2019, 571, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Hardie, D.G. Minireview: The AMP-Activated Protein Kinase Cascade: The Key Sensor of Cellular Energy Status. Endocrinology 2003, 144, 5179–5183. [Google Scholar] [CrossRef]
- Trapani, L.; Pallottini, V. Hypercholesterolemia and 3-Hydroxy 3-Methylglutaryl Coenzyme A Reductase Regulation during Ageing. Sci. World J. 2009, 9, 564–574. [Google Scholar] [CrossRef] [Green Version]
- Pallottini, V.; Marino, M.; Cavallini, G.; Bergamini, E.; Trentalance, A. Age-Related Changes of Isoprenoid Biosynthesis in Rat Liver and Brain. Biogerontology 2003, 4, 371–378. [Google Scholar] [CrossRef]
- Segatto, M.; Di Giovanni, A.; Marino, M.; Pallottini, V. Analysis of the Protein Network of Cholesterol Homeostasis in Different Brain Regions: An Age and Sex Dependent Perspective. J. Cell. Physiol. 2013, 228, 1561–1567. [Google Scholar] [CrossRef]
- Martini, C.; Pallottini, V. Cholesterol: From Feeding to Gene Regulation. Genes Nutr. 2007, 2, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallottini, V.; Martini, C.; Cavallini, G.; Donati, A.; Bergamini, E.; Notarnicola, M.; Caruso, M.G.; Trentalance, A. Modified HMG-CoA Reductase and LDLr Regulation Is Deeply Involved in Age-Related Hypercholesterolemia. J. Cell. Biochem. 2006, 98, 1044–1053. [Google Scholar] [CrossRef] [PubMed]
- Espenshade, P.J.; Hughes, A.L. Regulation of Sterol Synthesis in Eukaryotes. Annu. Rev. Genet. 2007, 41, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Boussicault, L.; Alves, S.; Lamazière, A.; Planques, A.; Heck, N.; Moumné, L.; Despres, G.; Bolte, S.; Hu, A.; Pagès, C.; et al. CYP46A1, the Rate-Limiting Enzyme for Cholesterol Degradation, Is Neuroprotective in Huntington’s Disease. Brain J. Neurol. 2016, 139 Pt 3, 953–970. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Smith, A.; Gelissen, I.C.; Ammit, A.J. The Effect of Statins and the Synthetic LXR Agonist T0901317 on Expression of ABCA1 Transporter Protein in Human Lung Epithelial Cell Lines in Vitro. Pharmacol. Rep. 2019, 71, 1219–1226. [Google Scholar] [CrossRef]
- Lanfranco, M.F.; Ng, C.A.; Rebeck, G.W. ApoE Lipidation as a Therapeutic Target in Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 6336. [Google Scholar] [CrossRef]
- Spagnuolo, M.S.; Pallottini, V.; Mazzoli, A.; Iannotta, L.; Tonini, C.; Morone, B.; Ståhlman, M.; Crescenzo, R.; Strazzullo, M.; Iossa, S.; et al. A Short-Term Western Diet Impairs Cholesterol Homeostasis and Key Players of Beta Amyloid Metabolism in Brain of Middle Aged Rats. Mol. Nutr. Food Res. 2020, e2000541. [Google Scholar] [CrossRef]
- Segatto, M.; Manduca, A.; Lecis, C.; Rosso, P.; Jozwiak, A.; Swiezewska, E.; Moreno, S.; Trezza, V.; Pallottini, V. Simvastatin Treatment Highlights a New Role for the Isoprenoid/Cholesterol Biosynthetic Pathway in the Modulation of Emotional Reactivity and Cognitive Performance in Rats. Neuropsychopharmacology 2014, 39, 841–854. [Google Scholar] [CrossRef] [Green Version]
- Drummond, G.R.; Selemidis, S.; Griendling, K.K.; Sobey, C.G. Combating Oxidative Stress in Vascular Disease: NADPH Oxidases as Therapeutic Targets. Nat. Rev. Drug Discov. 2011, 10, 453–471. [Google Scholar] [CrossRef] [Green Version]
- Shiose, A.; Sumimoto, H. Arachidonic Acid and Phosphorylation Synergistically Induce a Conformational Change of P47phox to Activate the Phagocyte NADPH Oxidase. J. Biol. Chem. 2000, 275, 13793–13801. [Google Scholar] [CrossRef] [Green Version]
- Dean, C.; Dunning, F.M.; Liu, H.; Bomba-Warczak, E.; Martens, H.; Bharat, V.; Ahmed, S.; Chapman, E.R. Axonal and Dendritic Synaptotagmin Isoforms Revealed by a PHluorin-Syt Functional Screen. Mol. Biol. Cell 2012, 23, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and Synaptic Plasticity, Cognitive Function, and Dysfunction. Handb. Exp. Pharmacol. 2014, 220, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Peters, R. Ageing and the Brain. Postgrad. Med. J. 2006, 82, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Duan, W.; Guo, Z. Meal Size and Frequency Affect Neuronal Plasticity and Vulnerability to Disease: Cellular and Molecular Mechanisms. J. Neurochem. 2003, 84, 417–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamrick, M.W.; Stranahan, A.M. Metabolic Regulation of Aging and Age-Related Disease. Ageing Res. Rev. 2020, 101175. [Google Scholar] [CrossRef]
- Vilchez, D.; Ros, S.; Cifuentes, D.; Pujadas, L.; Vallès, J.; García-Fojeda, B.; Criado-García, O.; Fernández-Sánchez, E.; Medraño-Fernández, I.; Domínguez, J.; et al. Mechanism Suppressing Glycogen Synthesis in Neurons and Its Demise in Progressive Myoclonus Epilepsy. Nat. Neurosci. 2007, 10, 1407–1413. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A Nutrient and Energy Sensor That Maintains Energy Homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Mair, W.; Morantte, I.; Rodrigues, A.P.C.; Manning, G.; Montminy, M.; Shaw, R.J.; Dillin, A. Lifespan Extension Induced by AMPK and Calcineurin Is Mediated by CRTC-1 and CREB. Nature 2011, 470, 404–408. [Google Scholar] [CrossRef]
- Martin-Montalvo, A.; Mercken, E.M.; Mitchell, S.J.; Palacios, H.H.; Mote, P.L.; Scheibye-Knudsen, M.; Gomes, A.P.; Ward, T.M.; Minor, R.K.; Blouin, M.-J.; et al. Metformin Improves Healthspan and Lifespan in Mice. Nat. Commun. 2013, 4, 2192. [Google Scholar] [CrossRef]
- Price, N.L.; Gomes, A.P.; Ling, A.J.Y.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. SIRT1 Is Required for AMPK Activation and the Beneficial Effects of Resveratrol on Mitochondrial Function. Cell Metab. 2012, 15, 675–690. [Google Scholar] [CrossRef] [Green Version]
- Martín, M.G.; Pfrieger, F.; Dotti, C.G. Cholesterol in Brain Disease: Sometimes Determinant and Frequently Implicated. EMBO Rep. 2014, 15, 1036–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietschy, J.M.; Turley, S.D. Thematic Review Series: Brain Lipids. Cholesterol Metabolism in the Central Nervous System during Early Development and in the Mature Animal. J. Lipid Res. 2004, 45, 1375–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perovic, M.; Mladenovic Djordjevic, A.; Smiljanic, K.; Tanic, N.; Rakic, L.; Ruzdijic, S.; Kanazir, S. Expression of Cholesterol Homeostasis Genes in the Brain of the Male Rat Is Affected by Age and Dietary Restriction. Biogerontology 2009, 10, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J. Rho GTPase Signalling in Cell Migration. Curr. Opin. Cell Biol. 2015, 36, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Cartocci, V.; Segatto, M.; Di Tunno, I.; Leone, S.; Pfrieger, F.W.; Pallottini, V. Modulation of the Isoprenoid/Cholesterol Biosynthetic Pathway During Neuronal Differentiation In Vitro. J. Cell. Biochem. 2016, 117, 2036–2044. [Google Scholar] [CrossRef]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, Oxidants, and Aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Hadem, I.K.H.; Sharma, R. Differential Regulation of Hippocampal IGF-1-Associated Signaling Proteins by Dietary Restriction in Aging Mouse. Cell. Mol. Neurobiol. 2017, 37, 985–993. [Google Scholar] [CrossRef]
- Alshamrani, A.A.; Bheemanapally, K.; Alhamyani, A.; Ali, M.H.; Napit, P.R.; Uddin, M.M.; Mahmood, A.S.M.H.; Ibrahim, M.M.H.; Briski, K.P. Sex-dimorphic Rostro-caudal Patterns of 5′-AMP-activated Protein Kinase Activation and Glucoregulatory Transmitter Marker Protein Expression in the Ventrolateral Ventromedial Hypothalamic Nucleus (VMNvl) in Hypoglycemic Male and Female Rats: Impact of Estradiol. J. Mol. Neurosci. 2020, 24. [Google Scholar] [CrossRef]
- Rivera, P.; Guerra-Cantera, S.; Vargas, A.; Díaz, F.; García-Úbedam, R.; Tovar, R.; Ramírez-López, M.T.; Argente, J.; de Fonseca, F.R.; Suárez, J.; et al. Maternal hypercaloric diet affects factors involved in lipid metabolism and the endogenous cannabinoid systems in the hypothalamus of adult offspring: Sex-specific response of astrocytes to palmitic acid and anandamide. Nutr. Neurosci. 2020, 21, 1–14. [Google Scholar] [CrossRef]
- Lejri, I.; Grimm, A.; Eckert, A. Mitochondria, Estrogen and Female Brain Aging. Front. Aging Neurosci. 2018, 27, 124. [Google Scholar] [CrossRef] [Green Version]
- Pierce, J.P.; Kievits, J.; Graustein, B.; Speth, R.C.; Iadecola, C.; Milner, T.A. Sex differences in the subcellular distribution of angiotensin type 1 receptors and NADPH oxidase subunits in the dendrites of C1 neurons in the rat rostral ventrolateral medulla. Neuroscience 2009, 29, 163, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 3M ND | 24M ND | 24M CR | |
|---|---|---|---|
| Animal weight g | 250 ± 21 | 444 ± 42 *** | 370 ± 23 ***,## |
| 3M ND | 24M ND | 24M CR | |
|---|---|---|---|
| Cholesterol µg/mg tissue | 4.77± 0.05 | 5.43 ± 0.8 | 4.65 ± 0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonini, C.; Segatto, M.; Martino, F.; Cigliano, L.; Nazzaro, M.; Barberio, L.; Mandalà, M.; Pallottini, V. Effects of Late-Life Caloric Restriction on Age-Related Alterations in the Rat Cortex and Hippocampus. Nutrients 2021, 13, 232. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13010232
Tonini C, Segatto M, Martino F, Cigliano L, Nazzaro M, Barberio L, Mandalà M, Pallottini V. Effects of Late-Life Caloric Restriction on Age-Related Alterations in the Rat Cortex and Hippocampus. Nutrients. 2021; 13(1):232. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13010232
Chicago/Turabian StyleTonini, Claudia, Marco Segatto, Francesca Martino, Luisa Cigliano, Martina Nazzaro, Laura Barberio, Maurizio Mandalà, and Valentina Pallottini. 2021. "Effects of Late-Life Caloric Restriction on Age-Related Alterations in the Rat Cortex and Hippocampus" Nutrients 13, no. 1: 232. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13010232