Maternal Folic Acid Deficiency Is Associated to Developing Nasal and Palate Malformations in Mice

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diet
2.2. Morphological Study
2.3. Head and Nasopalatine Region Measurements
2.4. Statistical Analysis
3. Results
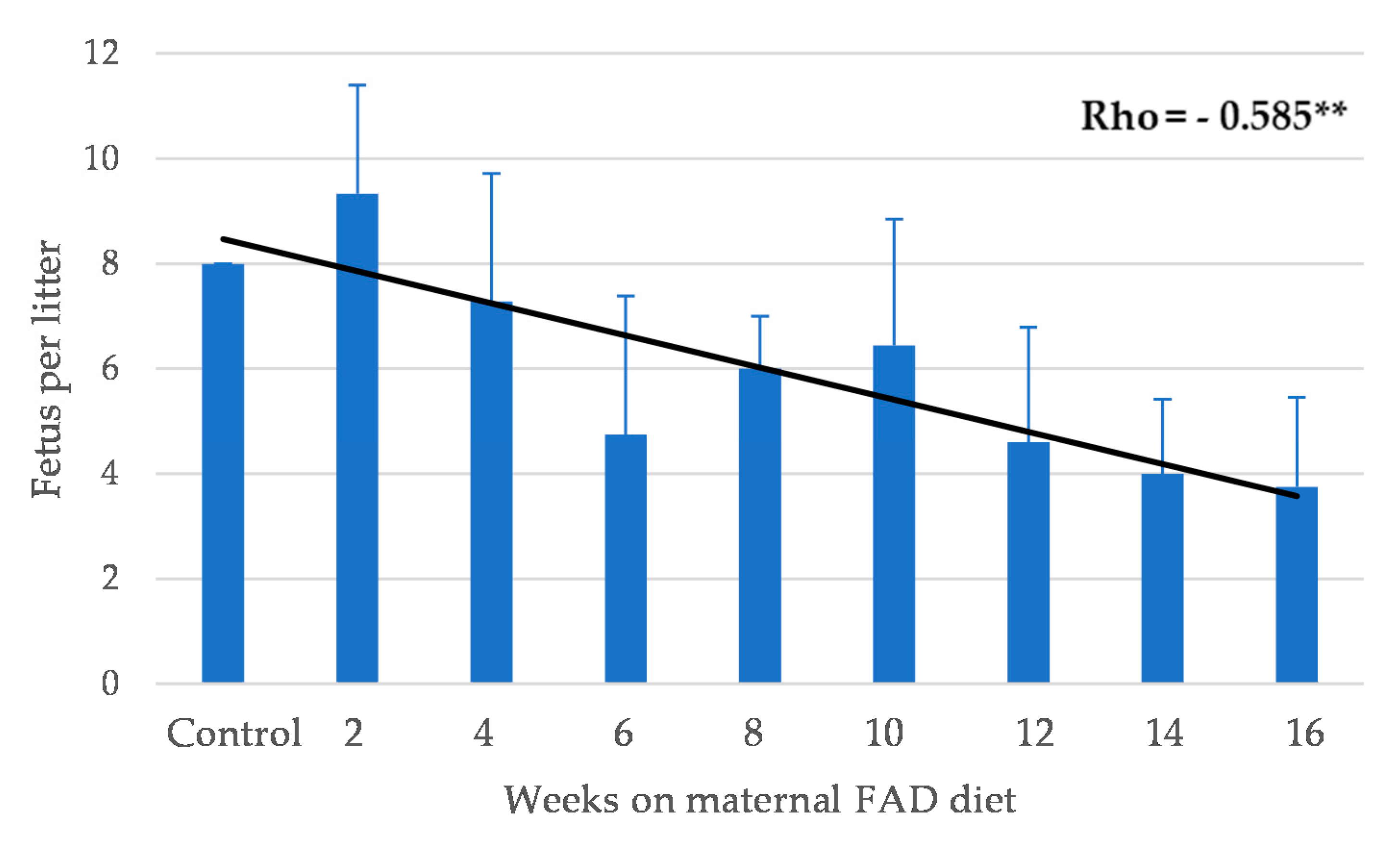
3.1. General Outcomes
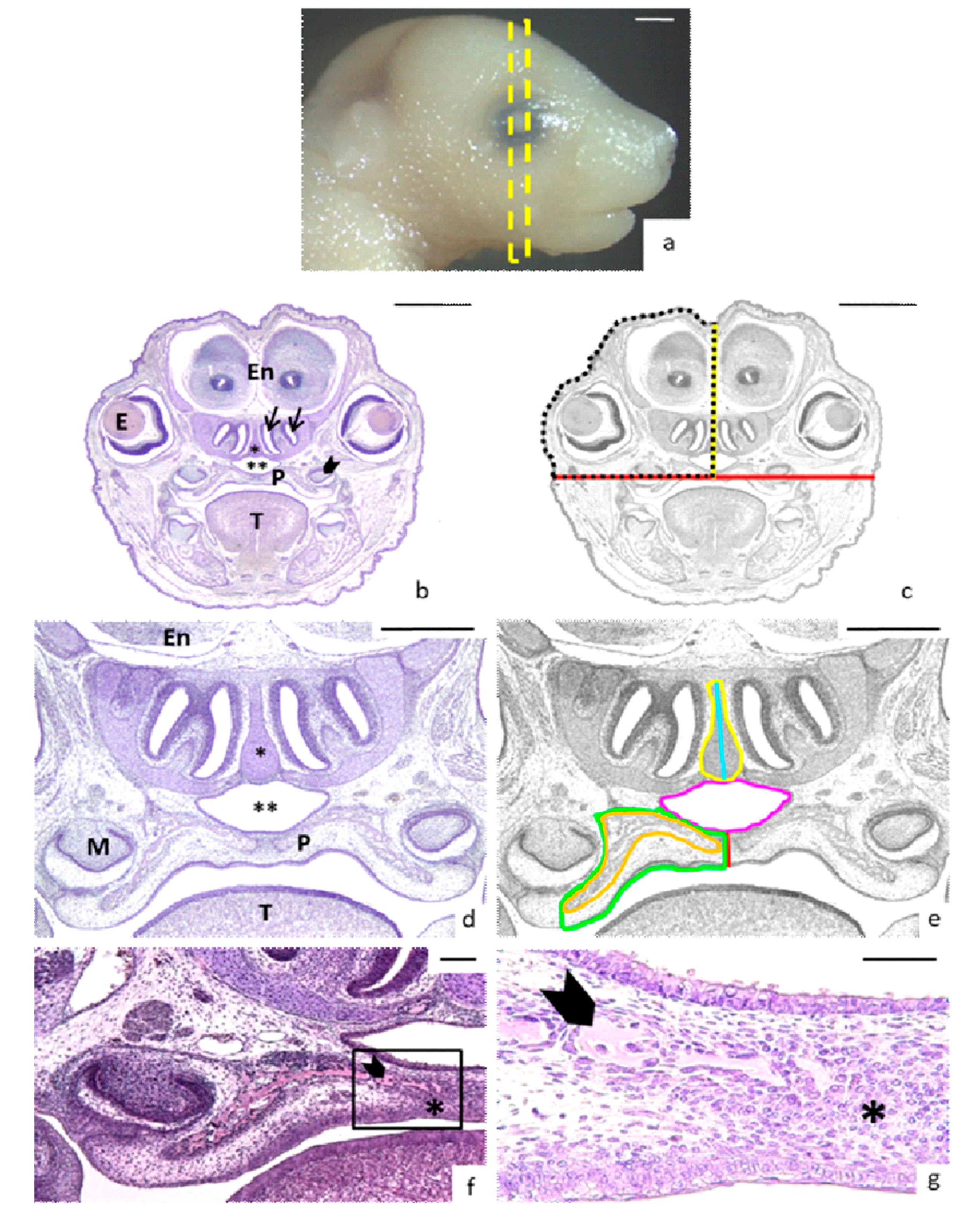
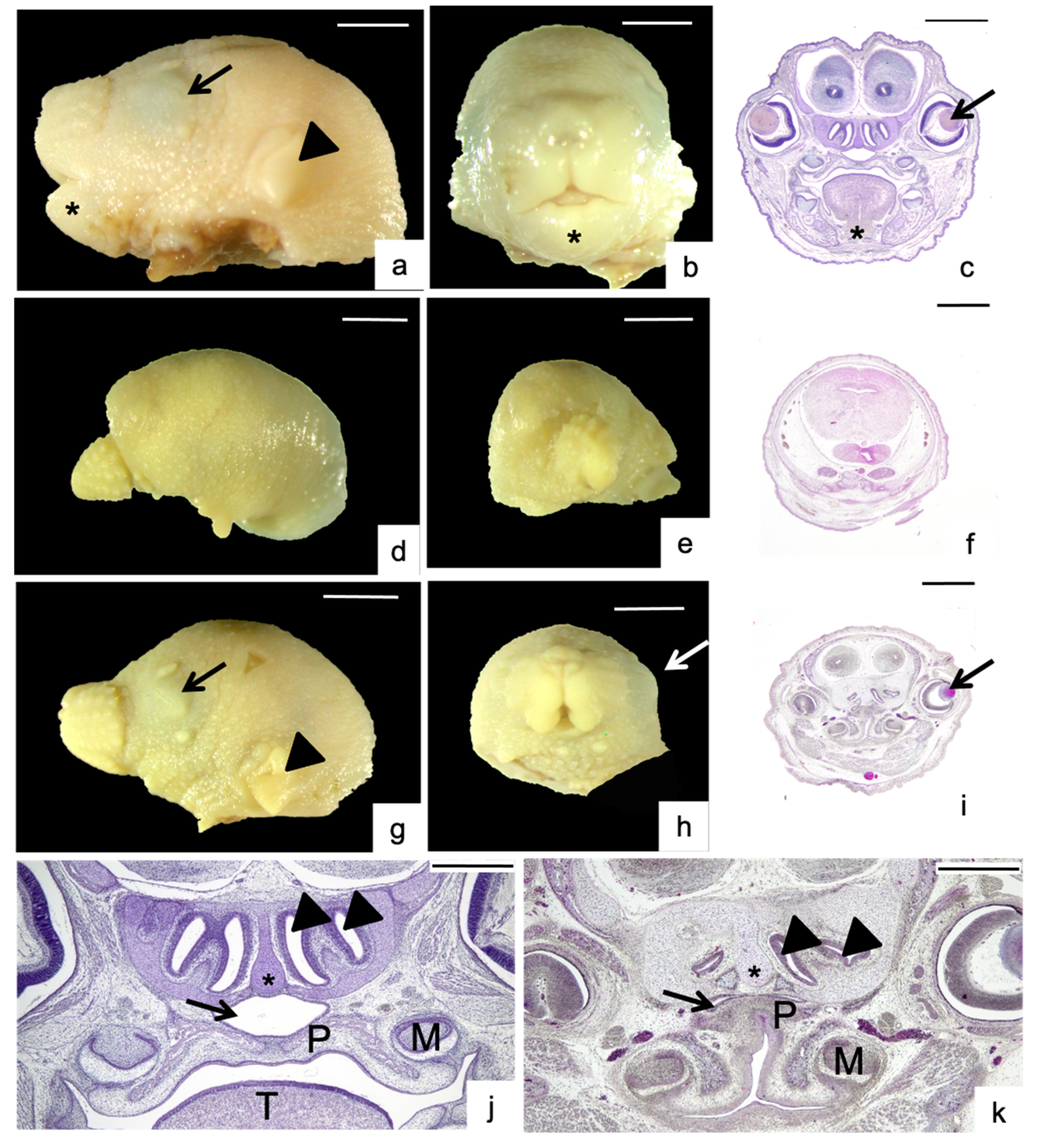
3.2. Morphological Analysis
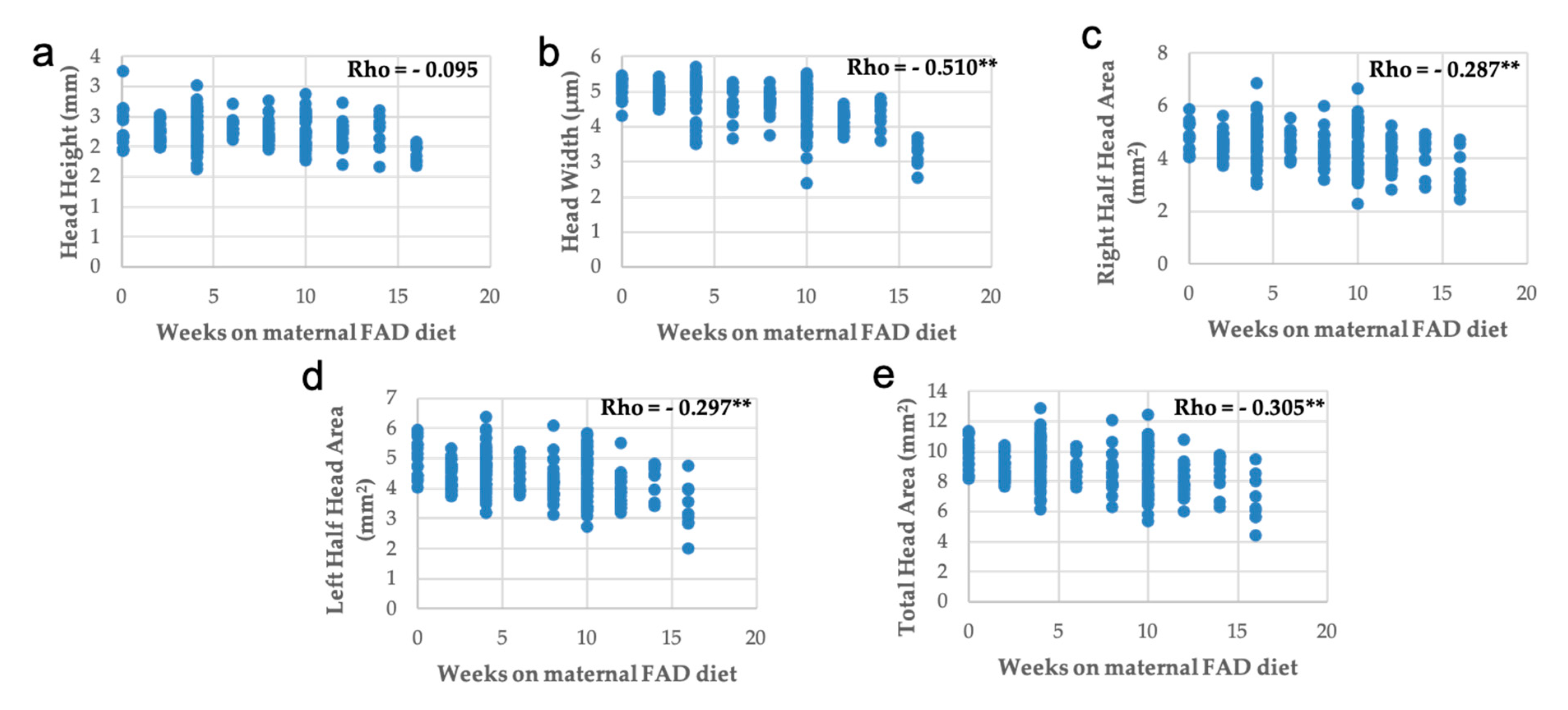
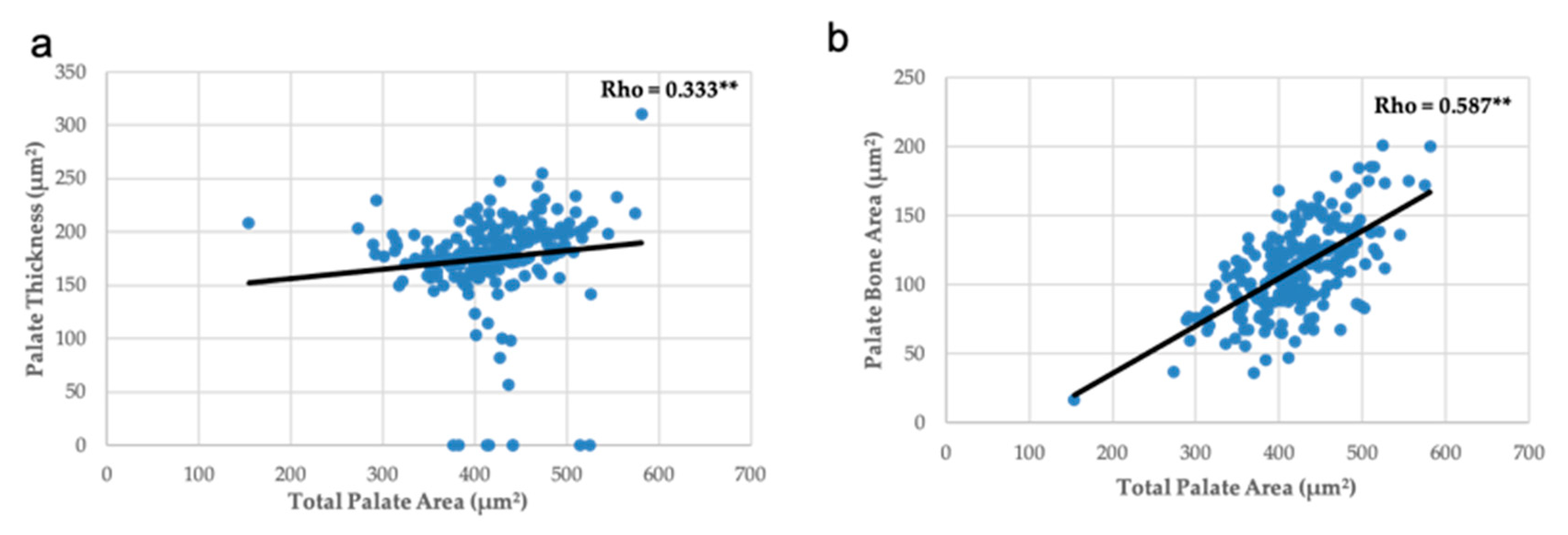
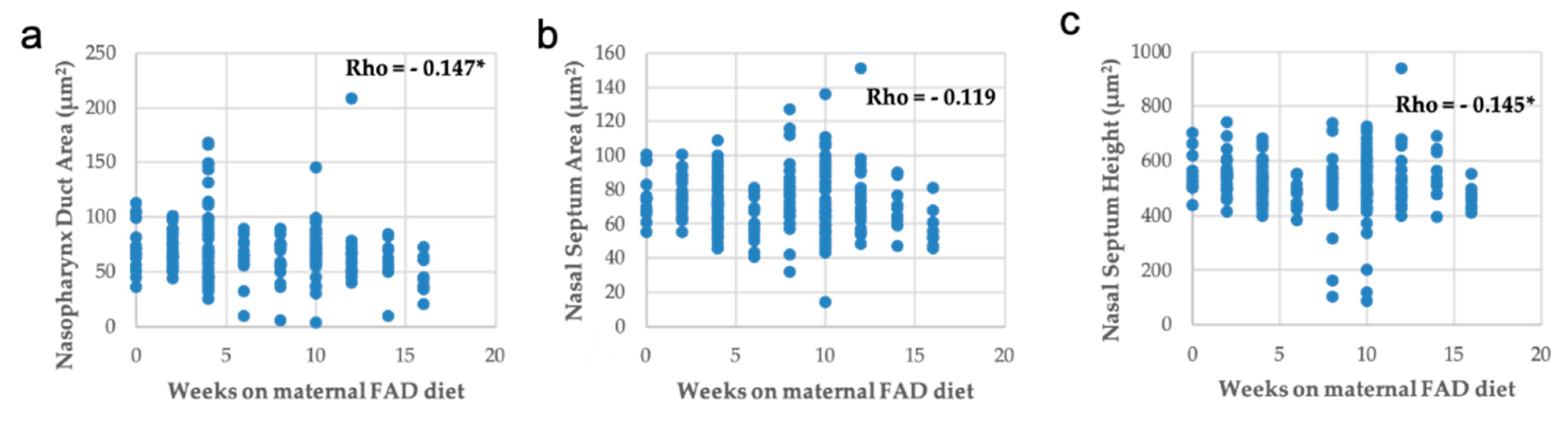
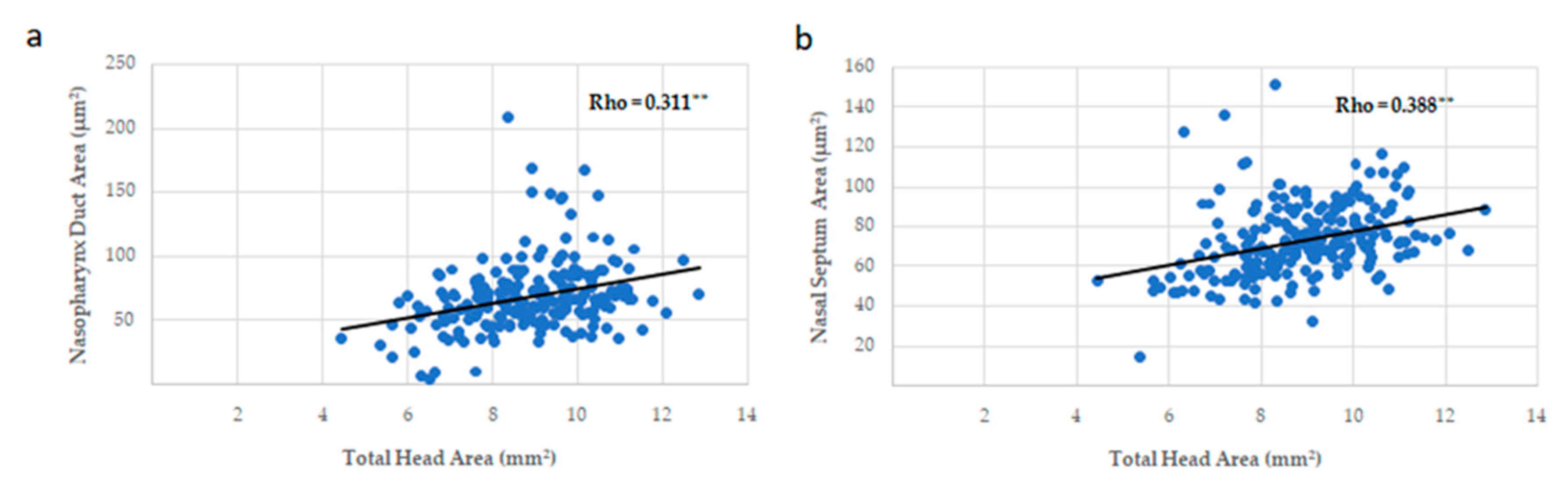
3.3. Head, Palate, and Nasal Measurements
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sperber, G.H. Craniofacial Development; BC Decker: Hamilton, ON, Canada, 2000. [Google Scholar]
- Ornoy, A. Craniofacial Malformations and Their Association with Brain Development: The Importance of a Multidisciplinary Approach for Treatment. Odontology 2020, 108, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mossey, P.; Castilla, E. Global Registry and Database on Craniofacial Anomalies. In Report of a WHO Registry Meeting on Craniofacial Anomalies; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- Mossey, P.A.; Little, J.; Munger, R.G.; Dixon, M.J.; Shaw, W.C. Cleft Lip and Palate. Lancet 2009, 374, 1773–1785. [Google Scholar] [CrossRef]
- Maldonado, E.; López-Gordillo, Y.; Partearroyo, T.; Varela-Moreiras, G.; Martínez-Álvarez, C.; Pérez-Miguelsanz, J. Tongue Abnormalities Are Associated to a Maternal Folic Acid Deficient Diet in Mice. Nutrients 2017, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado, E.; López, Y.; Herrera, M.; Martínez-Sanz, E.; Martínez-Álvarez, C.; Pérez-Miguelsanz, J. Craniofacial Structure Alterations of Foetuses from Folic Acid Deficient Pregnant Mice. Ann. Anat. 2018, 218, 59–68. [Google Scholar] [CrossRef]
- Wilcox, A.J.; Lie, R.T.; Solvoll, K.; Taylor, J.; McConnaughey, D.R.; Åbyholm, F.; Vindenes, H.; Vollset, S.E.; Drevon, C.A. Folic Acid Supplements and Risk of Facial Clefts: National Population Based Case-Control Study. BMJ 2007, 334, 464. [Google Scholar] [CrossRef] [Green Version]
- Wehby, G.L.; Murray, J.C. Folic Acid and Orofacial Clefts: A Review of the Evidence. Oral Dis. 2010, 16, 11–19. [Google Scholar] [CrossRef]
- López-Gordillo, Y.; Maldonado, E.; Nogales, L.; Del Río, A.; Barrio, M.C.; Murillo, J.; Martínez-Sanz, E.; Paradas-Lara, I.; Alonso, M.I.; Partearroyo, T.; et al. Maternal Folic Acid Supplementation Reduces the Severity of Cleft Palate in Tgf-β 3 Null Mutant Mice. Pediatr. Res. 2019, 85, 566–573. [Google Scholar] [CrossRef]
- Millacura, N.; Pardo, R.; Cifuentes, L.; Suazo, J. Effects of Folic Acid Fortification on Orofacial Clefts Prevalence: A Meta-Analysis. Public Health Nutr. 2017, 20, 2260–2268. [Google Scholar] [CrossRef]
- National Organization for Rare Disorders (NORD) Database; NIH/NIDCD: Danbury, CT, USA; Available online: https://rarediseases.org/for-patients-and-families/information-resources/rare-disease-information/ (accessed on 1 June 2020).
- Young, A.; Spinner, A. Hemifacial Microsomia. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Howe, A.M.; Hawkins, J.K.; Webster, W.S. The Growth of the Nasal Septum in the 6–9 Week Period of Foetal Development—Warfarin Embryopathy Offers a New Insight into Prenatal Facial Development. Aust. Dent. J. 2004, 49, 171–176. [Google Scholar] [CrossRef]
- Foster, A.; Holton, N. Variation in the Developmental and Morphological Interaction between the Nasal Septum and Facial Skeleton. Anat. Rec. 2016, 299, 730–740. [Google Scholar] [CrossRef] [Green Version]
- Katsube, M.; Yamada, S.; Yamaguchi, Y.; Takakuwa, T.; Yamamoto, A.; Imai, H.; Saito, A.; Shimizu, A.; Suzuki, S. Critical Growth Processes for the Midfacial Morphogenesis in the Early Prenatal Period. Cleft Palate Craniofac. J. 2019, 56, 1026–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado, E.; Murillo, J.; Barrio, C.; del Río, A.; Pérez-Miguelsanz, J.; López-Gordillo, Y.; Partearroyo, T.; Paradas, I.; Maestro, C.; Martínez-Sanz, E.; et al. Occurrence of Cleft-Palate and Alteration of Tgf-β(3) Expression and the Mechanisms Leading to Palatal Fusion in Mice Following Dietary Folic-Acid Deficiency. Cells Tissues Organs (Print) 2011, 194, 406–420. [Google Scholar] [CrossRef] [PubMed]
- Maestro-de-las-Casas, C.; Pérez-Miguelsanz, J.; López-Gordillo, Y.; Maldonado, E.; Partearroyo, T.; Varela-Moreiras, G.; Martínez-Álvarez, C. Maternal Folic Acid-Deficient Diet Causes Congenital Malformations in the Mouse Eye. Birth Defects Res. Part A Clin. Mol. Teratol. 2013, 97, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Ray, J.G.; Laskin, C.A. Folic Acid and Homocyst(e)Ine Metabolic Defects and the Risk of Placental Abruption, Pre-Eclampsia and Spontaneous Pregnancy Loss: A Systematic Review. Placenta 1999, 20, 519–529. [Google Scholar] [CrossRef]
- Sikora, J.; Magnucki, J.; Zietek, J.; Kobielska, L.; Partyka, R.; Kokocinska, D.; Białas, A. Homocysteine, Folic Acid and Vitamin B12 Concentration in Patients with Recurrent Miscarriages. Neuro Endocrinol. Lett. 2007, 28, 507–512. [Google Scholar]
- De la Calle, M.; Usandizaga, R.; Sancha, M.; Magdaleno, F.; Herranz, A.; Cabrillo, E. Homocysteine, Folic Acid and B-Group Vitamins in Obstetrics and Gynaecology. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 107, 125–134. [Google Scholar] [CrossRef]
- Pickell, L.; Li, D.; Brown, K.; Mikael, L.G.; Wang, X.-L.; Wu, Q.; Luo, L.; Jerome-Majewska, L.; Rozen, R. Methylenetetrahydrofolate Reductase Deficiency and Low Dietary Folate Increase Embryonic Delay and Placental Abnormalities in Mice. Birth Defects Res. Part A Clin. Mol. Teratol. 2009, 85, 531–541. [Google Scholar] [CrossRef]
- Heid, M.K.; Bills, N.D.; Hinrichs, S.H.; Clifford, A.J. Folate Deficiency Alone Does Not Produce Neural Tube Defects in Mice. J. Nutr. 1992, 122, 888–894. [Google Scholar] [CrossRef] [Green Version]
- Burgoon, J.M.; Selhub, J.; Nadeau, M.; Sadler, T.W. Investigation of the Effects of Folate Deficiency on Embryonic Development through the Establishment of a Folate Deficient Mouse Model. Teratology 2002, 65, 219–227. [Google Scholar] [CrossRef]
- Kalhan, S.C.; Marczewski, S.E. Methionine, Homocysteine, One Carbon Metabolism and Fetal Growth. Rev. Endocr. Metab. Disord. 2012, 13, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Jamilian, A.; Sarkarat, F.; Jafari, M.; Neshandar, M.; Amini, E.; Khosravi, S.; Ghassemi, A. Family History and Risk Factors for Cleft Lip and Palate Patients and Their Associated Anomalies. Stomatologija 2017, 19, 6. [Google Scholar]
- Wehby, G.L.; Félix, T.M.; Goco, N.; Richieri-Costa, A.; Chakraborty, H.; Souza, J.; Pereira, R.; Padovani, C.; Moretti-Ferreira, D.; Murray, J.C. High Dosage Folic Acid Supplementation, Oral Cleft Recurrence and Fetal Growth. Int. J. Environ. Res. Public Health 2013, 10, 590–605. [Google Scholar] [CrossRef] [PubMed]
- Tabler, J.M.; Barrell, W.B.; Szabo-Rogers, H.L.; Healy, C.; Yeung, Y.; Perdiguero, E.G.; Schulz, C.; Yannakoudakis, B.Z.; Mesbahi, A.; Wlodarczyk, B.; et al. Fuz Mutant Mice Reveal Shared Mechanisms between Ciliopathies and FGF-Related Syndromes. Dev. Cell 2013, 25, 623–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorlin, R.J.; Cohen, M.M.; Hennekam, R.C.M. Syndromes of the Head and Neck. Oxford Monographs on Medical Genetics, 4th ed.; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Hartman, C.; Holton, N.; Miller, S.; Yokley, T.; Marshall, S.; Srinivasan, S.; Southard, T. Nasal Septal Deviation and Facial Skeletal Asymmetries. Anat. Rec. 2016, 299, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Goergen, M.J.; Holton, N.E.; Grünheid, T. Morphological Interaction between the Nasal Septum and Nasofacial Skeleton during Human Ontogeny. J. Anat. 2017, 230, 689–700. [Google Scholar] [CrossRef]
- Xiao, S.; Hansen, D.K.; Horsley, E.T.M.; Tang, Y.-S.; Khan, R.A.; Stabler, S.P.; Jayaram, H.N.; Antony, A.C. Maternal Folate Deficiency Results in Selective Upregulation of Folate Receptors and Heterogeneous Nuclear Ribonucleoprotein-E1 Associated with Multiple Subtle Aberrations in Fetal Tissues. Birth Defects Res. Part A Clin. Mol. Teratol. 2005, 73, 6–28. [Google Scholar] [CrossRef]
- Gray, J.D.; Nakouzi, G.; Slowinska-Castaldo, B.; Dazard, J.-E.; Sunil Rao, J.; Nadeau, J.H.; Elizabeth Ross, M. Functional Interactions between the LRP6 WNT Co-Receptor and Folate Supplementation. Hum. Mol. Genet. 2010, 19, 4560–4572. [Google Scholar] [CrossRef] [Green Version]
- Copp, A.J.; Stanier, P.; Greene, N.D.E. Neural Tube Defects: Recent Advances, Unsolved Questions, and Controversies. Lancet Neurol. 2013, 12, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Canellos, G.P.; Mead, J.A.; Greenberg, N.H.; Schrecker, A.W. The Effect of Treatment with Cytotoxic Agents on Mouse Spleen Dihydrofolate Reductase Activity. Cancer Res. 1967, 27, 784–788. [Google Scholar]
- Ross, M.E. Gene-Environment Interactions, Folate Metabolism and the Embryonic Nervous System. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 471–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antony, A.C.; Hansen, D.K. Hypothesis: Folate-Responsive Neural Tube Defects and Neurocristopathies. Teratology 2000, 62, 42–50. [Google Scholar] [CrossRef]
- Dorsky, R.I.; Moon, R.T.; Raible, D.W. Environmental Signals and Cell Fate Specification in Premigratory Neural Crest. BioEssays 2000, 22, 708–716. [Google Scholar] [CrossRef]
- Santoso, M.I.E.; Rohman, M.S. Decreased TGF-Beta1 and IGF-1 Protein Expression in Rat Embryo Skull Bone in Folic Acid-Restricted Diet. J. Nutr. Biochem. 2006, 17, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Simpson, F.; Kerr, M.C.; Wicking, C. Trafficking, Development and Hedgehog. Mech. Dev. 2009, 126, 279–288. [Google Scholar] [CrossRef]
- Badano, J.L.; Mitsuma, N.; Beales, P.L.; Katsanis, N. The Ciliopathies: An Emerging Class of Human Genetic Disorders. Annu. Rev. Genom. Hum. Genet. 2006, 7, 125–148. [Google Scholar] [CrossRef] [Green Version]
- Fry, A.M.; Leaper, M.J.; Bayliss, R. The Primary Cilium. Organogenesis 2014, 10, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Cela, P.; Hampl, M.; Shylo, N.A.; Christopher, K.J.; Kavkova, M.; Landova, M.; Zikmund, T.; Weatherbee, S.D.; Kaiser, J.; Buchtova, M. Ciliopathy Protein Tmem107 Plays Multiple Roles in Craniofacial Development. J. Dent. Res. 2018, 97, 108–117. [Google Scholar] [CrossRef] [Green Version]
- Crider, K.S.; Yang, T.P.; Berry, R.J.; Bailey, L.B. Folate and DNA Methylation: A Review of Molecular Mechanisms and the Evidence for Folate’s Role2. Adv. Nutr. 2012, 3, 21–38. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weeks on Maternal FAD Diet | Mothers (n) | Number of Fetuses (n) | Nasal Malformations Only n (% Total) | Nasopalatine Malformations n (% Total) | Total Malformed n (% Group) |
|---|---|---|---|---|---|
| Control | 3 | 24 | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| 2 | 6 | 56 | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| 4 | 7 | 51 | 13 (5.0) | 0 (0.0) | 13 (5.0) |
| 6 | 4 | 19 | 0 (0.0) | 2 (0.8) | 2 (0.8) |
| 8 | 5 | 30 | 5 (1.9) | 5 (1.9) | 10 (3.8) |
| 10 | 9 | 58 | 4 (1.5) | 13 (5.0) | 17 (6.5) |
| 12 | 5 | 23 | 7 (2.7) | 1 (0.4) | 8 (3.1) |
| 14 | 3 | 12 | 1 (0.4) | 2 (0.8) | 3 (1.1) |
| 16 | 3 | 11 | 1 (0.4) | 2 (0.8) | 3 (1.1) |
| Total | 45 | 284 | 31 (11.9) | 25 (9.6) | 56 (21.5) |
| Asymmetry | Palatal Region Malformations | Nasal Region Malformations | |||||
|---|---|---|---|---|---|---|---|
| Weeks on Maternal FAD Diet | n (% Total; % Malformed) | HAP | CP | Nasal Septum | Nasopharynx Duct Shape | Cilia | Nasopharynx Epithelium |
| n (% Total; % Malformed) | n (% Total; % Malformed) | n (% Total; % Malformed) | n (% Total; % Malformed) | n (% Total; % Malformed) | n (% Total; % Malformed) | ||
| Control | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) |
| 2 | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) |
| 4 | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 9 (3.5; 16.1) | 12 (4.6; 21.4) | 5 (1.9; 8.9) |
| 6 | 0 (0.0; 0.0) | 1 (0.4; 1.8) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 1 (0.4; 1.8) | 0 (0.0; 0.0) | 0 (0.0; 0.0) |
| 8 | 5 (1.9; 8.9) | 1 (0.4; 1.8) | 3 (1.2; 5.4) | 6 (2.31; 10.71) | 7 (2.7; 12.5) | 4 (1.5; 7.1) | 1 (0.4; 1.8) |
| 10 | 5 (1.9; 8.9) | 5 (1.9; 8.9) | 5 (1.9; 8.9) | 10 (3.08; 14.29) | 14 (5.4; 25.0) | 5 (1.9; 8.9) | 4 (1.5; 7.1) |
| 12 | 2 (0.7; 3.6) | 0 (0.0; 0.0) | 1 (0.4; 1.8) | 2 (0.8; 3.6) | 5 (1.9; 8.9) | 5 (1.9; 8.9) | 3 (1.2; 5.4) |
| 14 | 0 (0.0; 0.0) | 1 (0.4; 1.8) | 0 (0.0; 0.0) | 0 (0.0; 0.0) | 1 (0.4; 1.8) | 1 (0.4; 1.8) | 1 (0.4; 1.8) |
| 16 | 0 (0.0; 0.0) | 1 (0.4; 1.8) | 1 (0.4; 1.8) | 1 (0.4; 1.8) | 2 (0.8; 3.6) | 2 (0.8; 3.6) | 1 (0.4; 1.8) |
| Total | 12 (4.6; 21.4) | 9 (3.5; 16.1) | 10 (3.8; 17.9) | 19 (7.3; 33.9) | 39 (15.0; 69.6) | 29 (11.2; 51.8) | 15 (5.8; 26.8) |
| Weeks on Maternal FAD Diet | |||||
|---|---|---|---|---|---|
| 0 | 2–6 | 8–16 | Total | ||
| Mothers without malformed fetuses | n (% group) | 3 a (100.0) | 11 a (64.7) | 4 b (16.0) | 18 (40.0) |
| Mothers with malformed fetuses | n (% group) | 0 (0.0) | 6 a (35.3) | 21 b (84.0) | 27 (60.0) |
| Total | n (% group) | 3 (100.0) | 17 (100.0) | 25 (100.0) | 45 (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maldonado, E.; Martínez-Sanz, E.; Partearroyo, T.; Varela-Moreiras, G.; Pérez-Miguelsanz, J. Maternal Folic Acid Deficiency Is Associated to Developing Nasal and Palate Malformations in Mice. Nutrients 2021, 13, 251. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13010251
Maldonado E, Martínez-Sanz E, Partearroyo T, Varela-Moreiras G, Pérez-Miguelsanz J. Maternal Folic Acid Deficiency Is Associated to Developing Nasal and Palate Malformations in Mice. Nutrients. 2021; 13(1):251. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13010251
Chicago/Turabian StyleMaldonado, Estela, Elena Martínez-Sanz, Teresa Partearroyo, Gregorio Varela-Moreiras, and Juliana Pérez-Miguelsanz. 2021. "Maternal Folic Acid Deficiency Is Associated to Developing Nasal and Palate Malformations in Mice" Nutrients 13, no. 1: 251. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13010251