
Coffeeberry Activates the CaMKII/CREB/BDNF Pathway, Normalizes Autophagy and Apoptosis Signaling in Nonalcoholic Fatty Liver Rodent Model

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals and Diets
2.2. Redox Status Analysis
2.3. Western Blotting
2.4. Statistics
3. Results
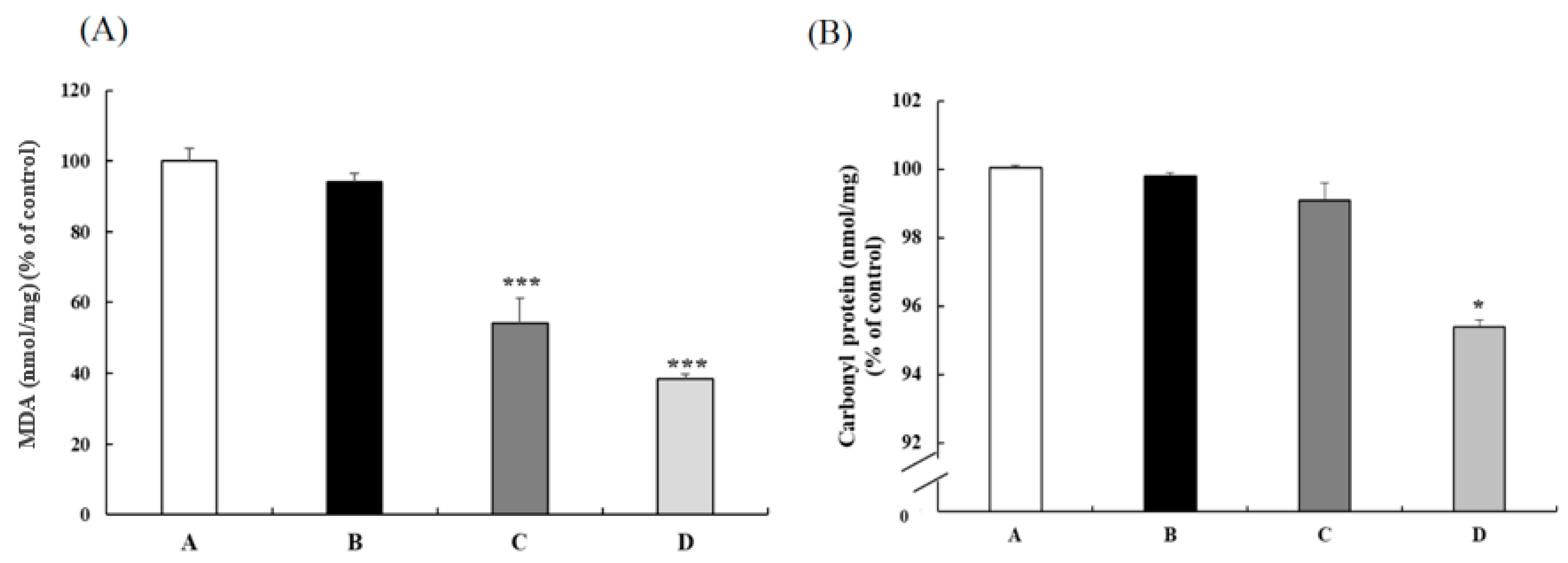
3.1. CB Reduced the Oxidative Status
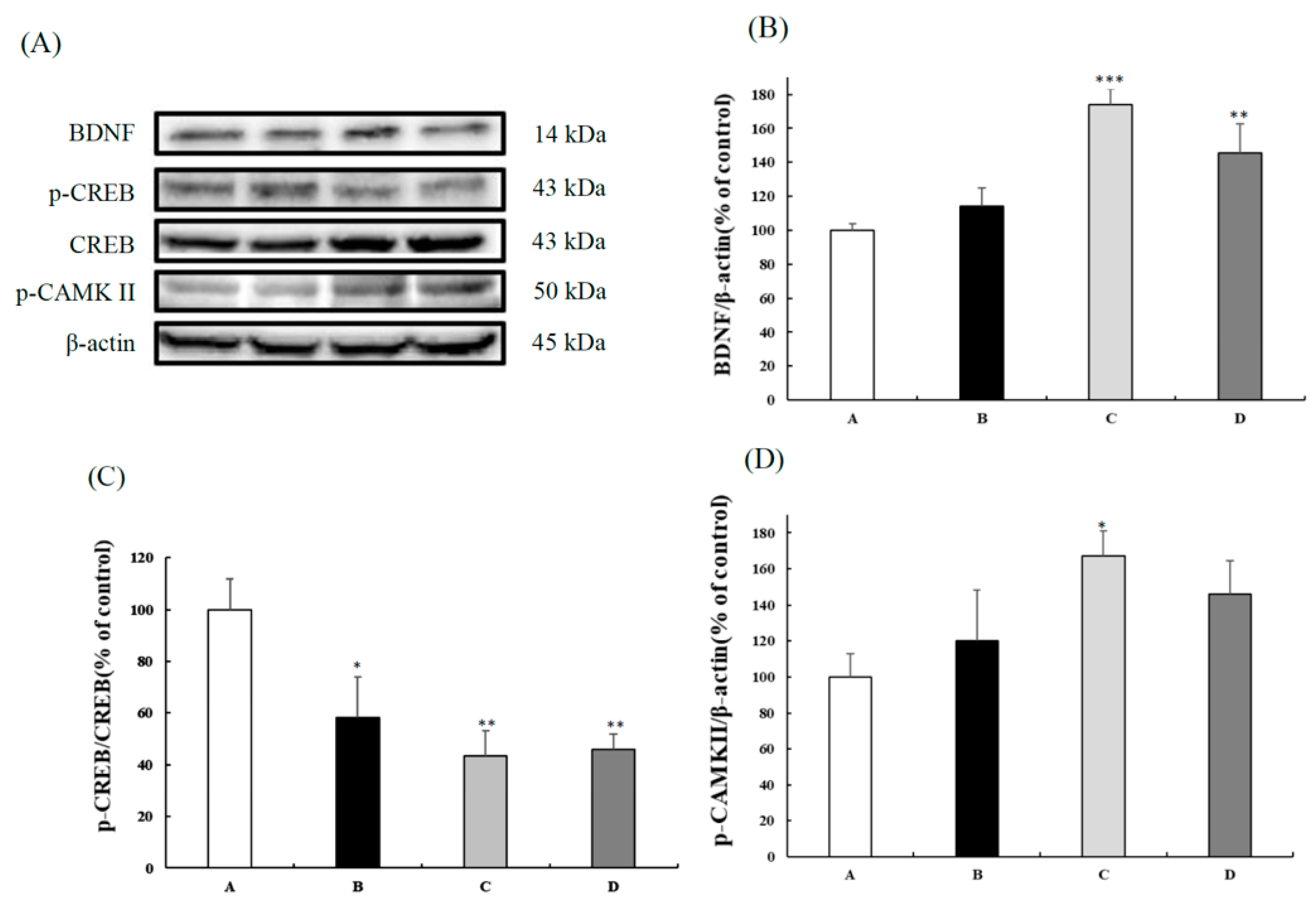
3.2. CB Improved the CaMKII/CREB/BDNF Signaling
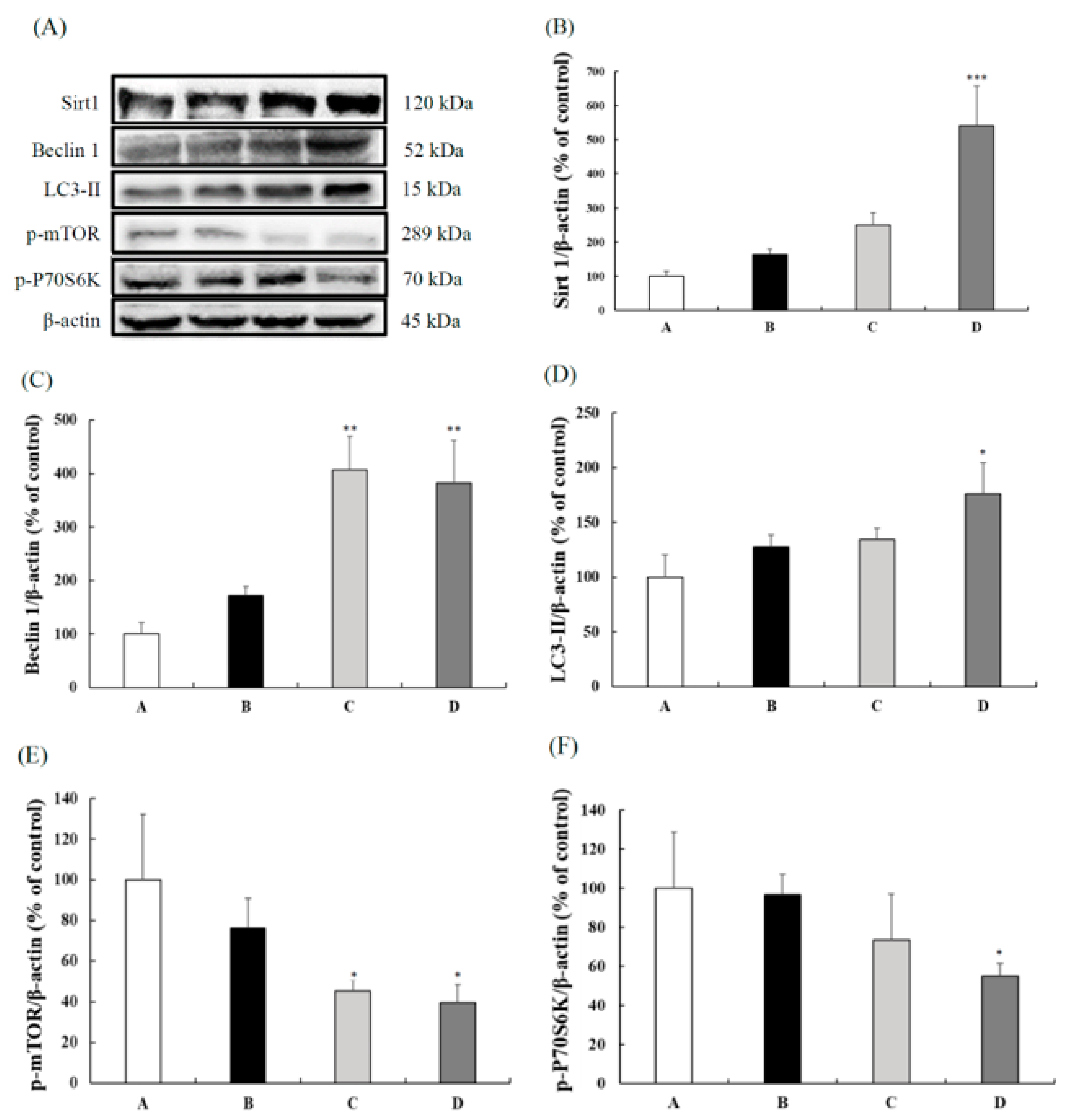
3.3. CB Enhanced the Autophagic Expressions
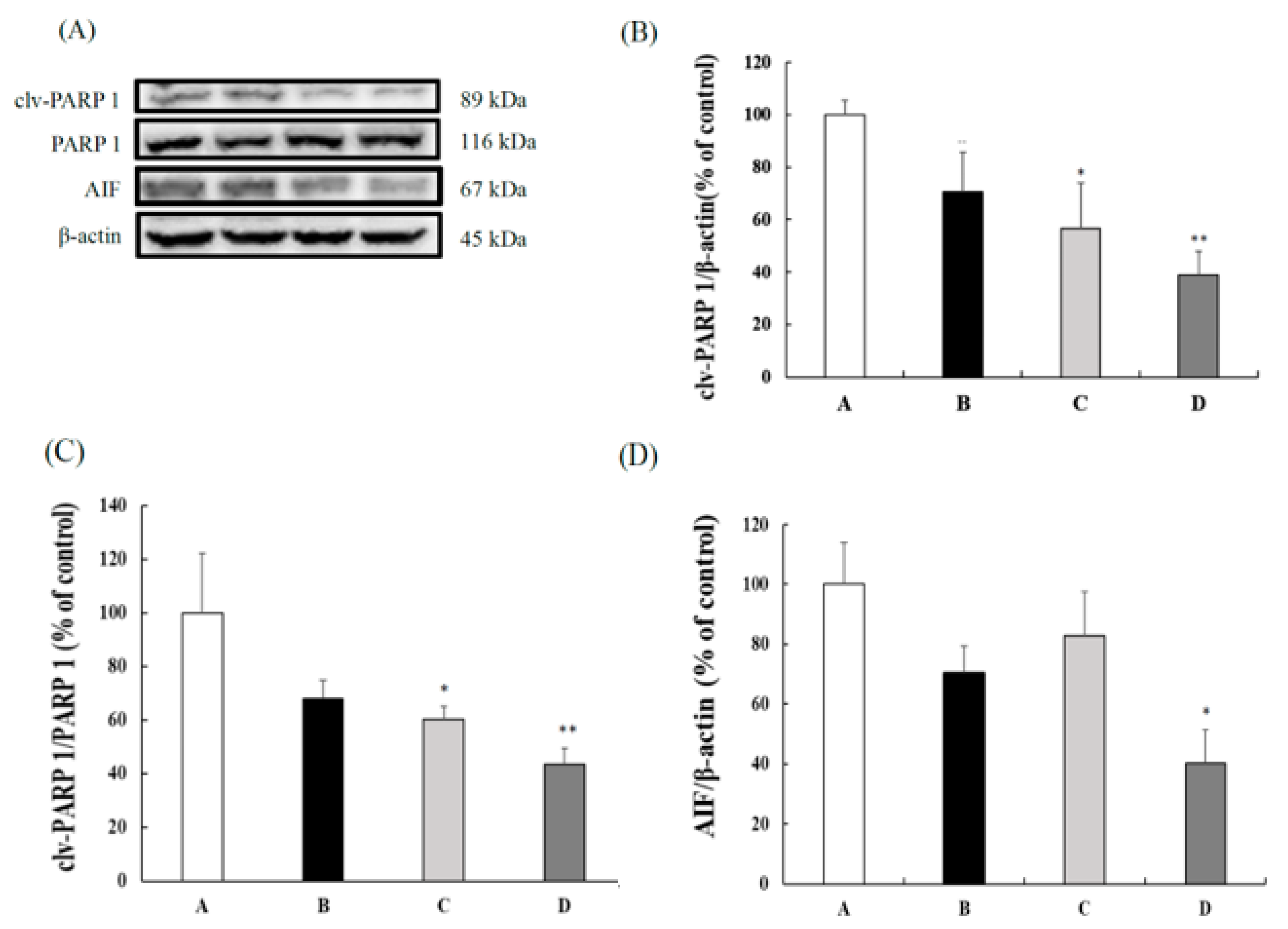
3.4. CB Inhibited the Caspase-Independent Apoptosis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ministry of Health and Welfare. 2020. Available online: https://www.mohw.gov.tw/mp-1.html (accessed on 12 October 2021).
- Hsu, C.S.; Kao, J.H. Non-alcoholic fatty liver disease: An emerging liver disease in Taiwan. J. Formos. Med. Assoc. 2012, 111, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Teoh, N.C.; Chitturi, S.; Farrell, G.C. Coffee and non-alcoholic fatty liver disease: Brewing evidence for hepatoprotection? J. Gastroenterol. Hepatol. 2014, 29, 435–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, M.; Itakura, Y.; Tsuchida, A.; Nakagawa, T.; Noguchi, H.; Taiji, M. Comparison of the antidiabetic effects of brain-derived neurotrophic factor and thiazolidinediones in obese diabetic mice. Diabetes Obes. Metab. 2007, 9, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, C.E.; Riener, R.; Reynolds, E.; Britton, G.B. NMDA receptor dysregulation in chronic state: A possible mechanism underlying depression with BDNF downregulation. Neurochem. Inter. 2014, 79, 88–97. [Google Scholar] [CrossRef]
- Zong, Y.; Sun, L.; Liu, B.; Deng, Y.S.; Zhan, D.; Chen, Y.L.; He, Y.; Liu, J.; Zhang, Z.J.; Sun, J.; et al. Resveratrol inhibits LPS-induced MAPKs activation via activation of the phosphatidylinositol 3-kinase pathway in murine RAW 264.7 macrophage cells. PLoS ONE 2012, 7, e44107. [Google Scholar] [CrossRef]
- Czaja, M.J. Function of autophagy in nonalcoholic fatty liver disease. Dig. Dis. Sci. 2016, 61, 1304–1313. [Google Scholar] [CrossRef]
- Komatsu, M.; Waguri, S.; Ueno, T.; Iwata, J.; Murata, S.; Tanida, I.; Ezaki, J.; Mizushima, N.; Ohsumi, Y.; Uchiyama, Y.; et al. Impairment of starvation-induced and constitutive autophagy in Atg7-deficient mice. J. Cell Biol. 2005, 169, 425–434. [Google Scholar] [CrossRef]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Czaja, M.J. Regulation of lipid stores and metabolism by lipophagy. Cell Death Differ. 2013, 20, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Alcendor, R.R.; Gao, S.; Zhai, P.; Zablocki, D.; Holle, E.; Yu, X.; Tian, B.; Wagner, T.; Vatner, S.F.; Sadoshima, J. Sirt1 regulates aging and resistance to oxidative stress in the heart. Circ. Res. 2007, 100, 1512–1521. [Google Scholar] [CrossRef]
- Rubinsztein, D.C.; Cuervo, A.M.; Ravikumar, B.; Sarkar, S.; Korolchuk, V.I.; Kaushik, S.; Klionsky, D.J. In search of an autophagomometer. Autophagy 2009, 5, 585–589. [Google Scholar] [CrossRef]
- Zhenhua, L.; Jian, W.; Xiao, Y. Functions of autophagy in pathological cardiac hypertrophy. Int. J. Biol. Sci. 2015, 11, 672–678. [Google Scholar]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abounit, K.; Scarabelli, T.M.; McCauley, R.B. Autophagy in mammalian cells. J. Biol. Chem. 2012, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dawson, V.L.; Dawson, T.M. Poly(ADP-ribose) signals to mitochondrial AIF: A key event in parthanatos. Exp. Neurol. 2009, 218, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostojic, S.M.; Stojanovic, M.D.; Djordjevic, B.; Jourkesh, M.; Vasiljevic, N. The effects of a 4-week coffeeberry supplementation on antioxidant status, endurance, and anaerobic performance in college athletes. Sports Med. 2008, 16, 281–294. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, D.H. Clinical safety and efficacy in photoaged skin with coffeeberry extract, a natural antioxidant. J. Cosmet. Dermatol. 2009, 22, 610–616. [Google Scholar]
- Reyes-Izquierdo, T.; Nemzer, B.; Shu, C.; Huynh, L.; Argumedo, R.; Keller, R.; Pietrzkowski, Z. Modulatory effect of coffee fruit extract on plasma levels of brain-derived neurotrophic factor in healthy subjects. Br. J. Nutr. 2013, 110, 420–425. [Google Scholar] [CrossRef]
- Suggs, A.; Oyetakin-White, P.; Baron, E.D. Effect of botanicals on inflammation and skin aging: Analyzing the evidence. Inflamm. Allergy Drug Targets 2014, 13, 168–176. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Stojanovic, M.D.; Djurovic, D. The effects of coffeeberry extract on plasma total phenolic content and antioxidant capacity in physically active men. Sci. Sports 2012, 27, 308–311. [Google Scholar] [CrossRef]
- Reed, R.A.; Mitchell, E.S.; Saunders, C.; O’Connor, P.J. Acute low and moderate doses of a caffeine-free polyphenol-rich coffeeberry extract improve feelings of alertness and fatigue resulting from the performance of fatiguing cognitive tasks. J. Cogn. Neurosci. 2019, 3, 193–206. [Google Scholar] [CrossRef]
- Ye, X.; Meeker, H.C.; Kozlowski, P.B.; Wegiel, J.; Wang, K.C.; Imaki, H.; Carp, R.I. Pathological changes in the liver of a senescence-accelerated mouse strain (SAMP8): A mouse model for the study of liver diseases. Histol. Histopathol. 2004, 19, 1141–1151. [Google Scholar] [PubMed]
- Lu, M.C.; Wu, C.S.; Yang, M.D.; Wang, M.F.; Chan, Y.C. Effects of coffeeberry on fatty liver and related inflammatory factors. Taiwan. J. Agric. Chem. Food Sci. 2020, 58, 1–9. [Google Scholar]
- Chan, Y.C.; Hwang, J.H. Effects of Spirulina on the functions and redox status of auditory system in senescence-accelerated prone-8 mice. PLoS ONE 2017, 12, e0178916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.C.; Wu, C.S.; Wu, T.C.; Lin, Y.H.; Chang, S.J. A Standardized extract of Asparagus officinalis Stem (ETAS®) ameliorates cognitive impairment, inhibits amyloid β deposition via BACE-1 and normalizes circadian rhythm signaling via MT1 and MT2. Nutrients 2019, 11, 1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, R.; Matsuzaki, K.; Sumiyoshi, E.; Hossain, M.E.; Hashimoto, M.; Katakura, M.; Sugimoto, N.; Shido, O. Theobromine improves working memory by activating the CaMKII/CREB/BDNF pathway in rats. Nutrients 2019, 11, 888. [Google Scholar] [CrossRef] [Green Version]
- Sheng, M.; Thompson, M.A.; Greenberg, M.E. CREB: A Ca(2+)-regulated transcription factor phosphorylated by calmodulin-dependent kinases. Science 1991, 252, 1427–1430. [Google Scholar] [CrossRef]
- Barco, A.; Bailey, C.H.; Kandel, E.R. Common molecular mechanisms in explicit and implicit memory. J. Neurochem. 2006, 97, 1520–1533. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Meng, X.; Zhai, Y.; Ye, T.; Zhou, P.; Nan, F.; Sun, G.; Sun, X. Antidepressant-like effects of the Guanxin Danshen formula via mediation of the CaMK II-CREB-BDNF signalling pathway in chronic unpredictable mild stress-induced depressive rats. Ann. Transl. Med. 2019, 7, 564. [Google Scholar] [CrossRef]
- Chao, M.V.; Rajagopal, R.; Lee, F.S. Neurotrophin signalling in health and disease. Clin. Sci. 2006, 110, 167–173. [Google Scholar] [CrossRef]
- Hashida, R.; Nakano, D.; Yamamura, S.; Kawaguchi, T.; Tsutsumi, T.; Matsuse, H.; Torimura, T. Association between activity and brain-derived neurotrophic factor in patients with non-alcoholic fatty liver disease: A data-mining analysis. Life 2021, 11, 799. [Google Scholar] [CrossRef] [PubMed]
- Shu, H.C.; Hu, J.; Jiang, X.B.; Deng, H.Q.; Zhang, K.H. BDNF gene polymorphism and serum level correlate with liver function in patients with hepatitis B-induced cirrhosis. Int. J. Clin. Exp. Pathol. 2019, 12, 2368–2380. [Google Scholar] [PubMed]
- Camer, D.; Yu, Y.; Szabo, A.; Dinh, C.H.; Wang, H.; Cheng, L.; Huang, X.F. Bardoxolone methyl prevents insulin resistance and the development of hepatic steatosis in mice fed a high-fat diet. Mol. Cell Endocrinol. 2015, 412, 36–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genzer, Y.; Chapnik, N.; Froy, O. Effect of brain-derived neurotrophic factor (BDNF) on hepatocyte metabolism. Int. J. Biochem. Cell Biol. 2017, 88, 69–74. [Google Scholar] [CrossRef]
- Herzig, S.; Long, F.; Jhala, U.S.; Hedrick, S.; Quinn, R.; Bauer, A.; Rudolph, D.; Schutz, G.; Yoon, C.; Puigserver, P.; et al. CREB regulates hepatic gluconeogenesis through the coactivator PGC-1. Nature 2001, 413, 179–183. [Google Scholar] [CrossRef]
- Herzig, S.; Hedrick, S.; Morantte, I.; Koo, S.H.; Galimi, F.; Montminy, M. CREB controls hepatic lipid metabolism through nuclear hormone receptor PPAR-γ. Nature 2003, 426, 190–193. [Google Scholar] [CrossRef]
- Houglum, K.; Lee, K.S.; Chojkier, M. Proliferation of hepatic stellate cells is inhibited by phosphorylation of CREB on serine 133. J. Clin. Investig. 1997, 99, 1322–1328. [Google Scholar] [CrossRef] [Green Version]
- Ozcan, L.; Wong, C.C.; Li, G.; Xu, T.; Pajvani, U.; Park, S.K.; Wronska, A.; Chen, B.X.; Marks, A.R.; Fukamizu, A.; et al. Calcium signaling through CaMKII regulates hepatic glucose production in fasting and obesity. Cell Metab. 2012, 15, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Khambu, B.; Yan, S.; Huda, N.; Liu, G.; Yin, X.M. Autophagy in non-alcoholic fatty liver disease and alcoholic liver disease. Liver 2018, 2, 112–119. [Google Scholar] [CrossRef]
- He, Y.; Ao, N.; Yang, J.; Wang, X.; Jin, S.; Du, J. The preventive effect of liraglutide on the lipotoxic liver injury via increasing autophagy. Ann. Hepatol. 2020, 19, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, P.; Fu, S.; Calay, E.S.; Hotamisligil, G.S. Defective hepatic autophagy in obesity promotes ER stress and causes insulin resistance. Cell Metab. 2010, 11, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yi, L.; Ye, X.; Zeng, X.; Qin, Y.; Zhang, Q.; Mi, M. Resveratrol ameliorates lipid droplet accumulation in liver through a SIRT1/ATF6-dependent mechanism. Cell. Physiol. Biochem. 2018, 51, 2397–2420. [Google Scholar] [CrossRef]
- Deng, X.Q.; Chen, L.L.; Li, N.X. The expression of SIRT1 in nonalcoholic fatty liver disease induced by high-fat diet in rats. Liver Int. 2007, 27, 708–715. [Google Scholar] [CrossRef]
- Nakadera, E.; Yamashina, S.; Izumi, K.; Inami, Y.; Sato, T.; Fukushima, H.; Kon, K.; Ikejima, K.; Ueno, T.; Watanabe, S. Inhibition of mTOR improves the impairment of acidification in autophagic vesicles caused by hepatic steatosis. Biochem. Biophys. Res. Commun. 2016, 469, 1104–1110. [Google Scholar] [CrossRef]
- Kang, J.W.; Choa, H.I.; Lee, S.M. Melatonin inhibits mTOR-dependent autophagy during liver ischemia/reperfusion. Cell. Physiol. Biochem. 2014, 33, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yanai, S.; Shimokado, K.; Ishigami, A. Coffee consumption in aged mice increases energy production and decreases hepatic mTOR levels. Nutrition 2017, 38, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Salomone, F.; Galvano, F.; Li Volti, G. Molecular bases underlying the hepatoprotective effects of coffee. Nutrients 2017, 9, 85. [Google Scholar] [CrossRef] [Green Version]
- Zadernowski, R.; Naczk, M.; Nesterowicz, J. Phenolic acid profiles in some small berries. J. Agric. Food Chem. 2005, 53, 2118–2124. [Google Scholar] [CrossRef] [PubMed]
- Saiki, S.; Sasazawa, Y.; Imamichi, Y.; Kawajiri, S.; Fujimaki, T.; Tanida, I.; Kobayashi, H.; Sato, F.; Sato, S.; Ishikawa, K.; et al. Caffeine induces apoptosis by enhancement of autophagy via PI3K/Akt/mTOR/p70S6K inhibition. Autophagy 2011, 7, 176–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Refolo, M.G.; Lippolis, C.; Carella, N.; Cavallini, A.; Messa, C.; D’Alessandro, R. Chlorogenic acid improves the Regorafenib effects in human hepatocellular carcinoma cells. Int. J. Mol. Sci. 2018, 19, 1518. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Gao, Y.Q.; Zhang, Y.; Wang, H.; Liu, G.S.; Lei, J.Y. Chlorogenic acid alleviates autophagy and insulin resistance by suppressing JNK pathway in a rat model of nonalcoholic fatty liver disease. J. Biosci. 2018, 43, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, P.; Horváth, B.; Rajesh, M.; Varga, Z.V.; Gariani, K.; Ryu, D.; Cao, Z.; Holovac, E.; Park, O.; Zhou, Z.; et al. PARP inhibition protects against alcoholic and non-alcoholic steatohepatitis. J. Hepatol. 2017, 66, 589–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gariani, K.; Ryu, D.; Menzies, K.J.; Yi, H.S.; Stein, S.; Zhang, H.; Perino, A.; Lemos, V.; Katsyuba, E.; Jha, P.; et al. Inhibiting poly ADP-ribosylation increases fatty acid oxidation and protects against fatty liver disease. J. Hepatol. 2017, 66, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salomone, F.; Barbagallo, I.; Godos, J.; Lembo, V.; Currenti, W.; Cinà, D.; Avola, R.; D’Orazio, N.; Morisco, F.; Galvano, F.; et al. Silibinin restores NAD⁺ levels and induces the SIRT1/AMPK pathway in non-alcoholic fatty liver. Nutrients 2017, 9, 1086. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, Y.; Huang, H.; Shi, M.; Yang, W.; Kuang, J.; Yan, J. Autophagy mediated by endoplasmic reticulum stress enhances the caffeine-induced apoptosis of hepatic stellate cells. Int. J. Mol. Med. 2017, 40, 1405–1414. [Google Scholar] [CrossRef]
- Wang, Z.; Gu, C.; Wang, X.; Lang, Y.; Wu, Y.; Wu, X.; Zhu, X.; Wang, K.; Yang, H. Caffeine enhances the anti-tumor effect of 5-fluorouracil via increasing the production of reactive oxygen species in hepatocellular carcinoma. Med. Oncol. 2019, 36, 97. [Google Scholar] [CrossRef]
- Chen, J.C.; Hwang, J.H.; Chiu, W.H.; Chan, Y.C. Tetrandrine and caffeine modulated cell cycle and increased glioma cell death via caspase-dependent and caspase-independent apoptosis pathways. Nutr. Cancer 2014, 66, 700–706. [Google Scholar] [CrossRef]
- Zhang, Y.; Miao, L.; Zhang, H.; Wu, G.; Zhang, Z.; Lv, J. Chlorogenic acid against palmitic acid in endoplasmic reticulum stress-mediated apoptosis resulting in protective effect of primary rat hepatocytes. Lipids Health Dis. 2018, 17, 270. [Google Scholar] [CrossRef] [Green Version]
- Heimbach, J.T.; Marone, P.A.; Hunter, J.M.; Nemzer, B.V.; Stanley, S.M.; Kennepohl, E. Safety studies on products from whole coffee fruit. Food Chem. Toxicol. 2010, 48, 2517–2525. [Google Scholar] [CrossRef]
- Halvorsen, B.L.; Carlsen, M.H.; Phillips, K.M.; Bøhn, S.K.; Holte, K.; Jacobs, D.R.; Blomhoff, R. Content of redox-active compounds in foods consumed in the United States. Am. J. Clin. Nutr. 2006, 84, 95–135. [Google Scholar] [CrossRef] [Green Version]
- Duangjai, A.; Nuengchamnong, N.; Suphrom, N.; Trisat, K.; Limpeanchob, N.; Saokaew, S. Potential of coffee fruit extract and quinic acid on adipogenesis and lipolysis in 3T3-L1 adipocytes. Kobe J. Med. Sci. 2018, 64, E84–E92. [Google Scholar]
- Vitaglione, P.; Morisco, F.; Mazzone, G.; Amoruso, D.C.; Ribecco, M.T.; Romano, A.; Fogliano, V.; Caporaso, N.; D’Argenio, G. Coffee reduces liver damage in a rat model of steatohepatitis: The underlying mechanisms and the role of polyphenols and melanoidins. Hepatology 2010, 52, 1652–1661. [Google Scholar] [CrossRef]
- Vistisen, K.; Poulsen, H.E.; Loft, S. Foreign compound metabolism capacity in man measured from metabolites of dietary caffeine. Carcinogenesis 1992, 13, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Devasagayam, T.P.; Kamat, J.P.; Mohan, H.; Kesavan, P.C. Caffeine as an antioxidant: Inhibition of lipid peroxidation induced by reactive oxygen species. Biochim. Biophys. Acta. 1996, 1282, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Sharp, D.S.; Benowitz, N.L. Re: “Alcohol, smoking, coffee, and cirrhosis” and “coffee and serum gamma-glutamyltransferase: A study of self-defense officials in Japan”. Am. J. Epidemiol. 1995, 141, 480–481. [Google Scholar] [CrossRef]
- Sinha, R.A.; Farah, B.L.; Singh, B.K.; Siddique, M.M.; Li, Y.; Wu, Y.; Ilkayeva, O.R.; Gooding, J.; Ching, J.; Zhou, J.; et al. Caffeine stimulates hepatic lipid metabolism by the autophagy-lysosomal pathway in mice. Hepatology 2014, 59, 1366–1380. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Weight Gain (gm) | Food Intakes (gm/day) | Liver (g/100g Body Weight) |
|---|---|---|---|---|
| A | 6 | 3.08 ± 0.43 | 7.83 ± 0.16 | 4.36 ± 0.15 |
| B | 6 | 3.41 ± 0.32 | 7.51 ± 0.17 | 4.21 ± 0.20 |
| C | 6 | 3.42 ± 0.35 | 7.73 ± 0.49 | 4.20 ± 0.19 |
| D | 6 | 3.33 ± 0.38 | 7.69 ± 0.28 | 4.25 ± 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, M.-C.; Lee, I.-T.; Hong, L.-Z.; Ben-Arie, E.; Lin, Y.-H.; Lin, W.-T.; Kao, P.-Y.; Yang, M.-D.; Chan, Y.-C. Coffeeberry Activates the CaMKII/CREB/BDNF Pathway, Normalizes Autophagy and Apoptosis Signaling in Nonalcoholic Fatty Liver Rodent Model. Nutrients 2021, 13, 3652. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13103652
Lu M-C, Lee I-T, Hong L-Z, Ben-Arie E, Lin Y-H, Lin W-T, Kao P-Y, Yang M-D, Chan Y-C. Coffeeberry Activates the CaMKII/CREB/BDNF Pathway, Normalizes Autophagy and Apoptosis Signaling in Nonalcoholic Fatty Liver Rodent Model. Nutrients. 2021; 13(10):3652. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13103652
Chicago/Turabian StyleLu, Meng-Chun, I-Te Lee, Ling-Zong Hong, Eyal Ben-Arie, Yu-Hsuan Lin, Wei-Ting Lin, Pei-Yu Kao, Mei-Due Yang, and Yin-Ching Chan. 2021. "Coffeeberry Activates the CaMKII/CREB/BDNF Pathway, Normalizes Autophagy and Apoptosis Signaling in Nonalcoholic Fatty Liver Rodent Model" Nutrients 13, no. 10: 3652. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13103652