
The Fatty Acid Species and Quantity Consumed by the Breastfed Infant Are Important for Growth and Development

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Recruitment, Data Collection and Human Milk Sampling
2.2. Human Milk Fatty Acid Analysis
2.3. Daily Milk, Total Lipid and Fatty Acid Intake
2.4. Statistical Analysis
3. Results
3.1. Study Cohort
3.2. Maternal Diet
3.3. Fatty Acid Composition of Human Milk
3.4. Maternal Diet and Human Milk Fatty Acid Composition
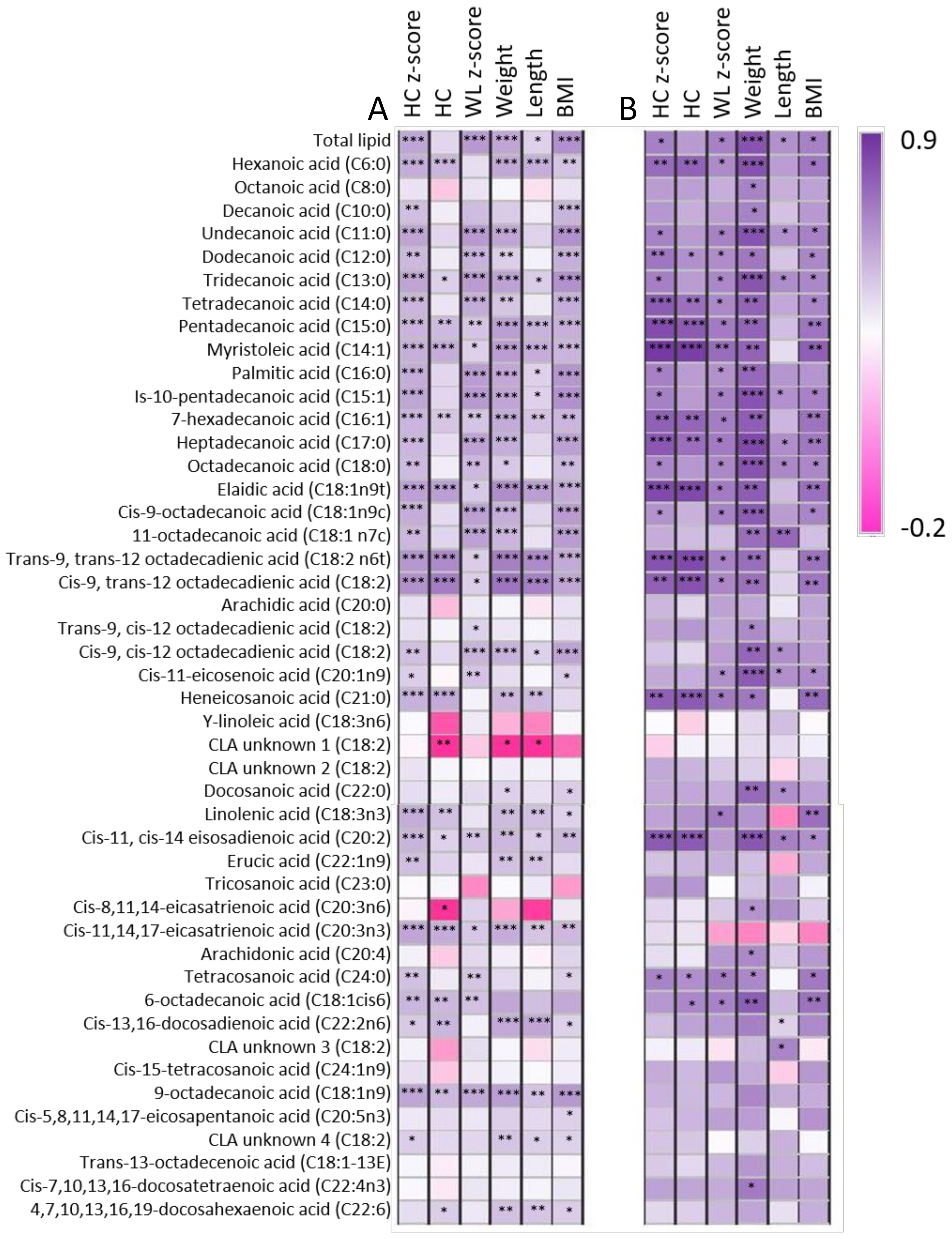
3.5. Relationships between Maternal Characteristics, Infant Intake, and Infant Growth Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koletzko, B. Human Milk Lipids. Ann. Nutr. Metab. 2016, 69, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Lopez, C.; Ménard, O. Human milk fat globules: Polar lipid composition and in situ structural investigations revealing the heterogeneous distribution of proteins and the lateral segregation of sphingomyelin in the biological membrane. Colloids Surf. B Biointerfaces 2011, 83, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Hachey, D.L.; Silber, G.H.; Wong, W.W.; Garza, C. Human lactation. II: Endogenous fatty acid synthesis by the mammary gland. Pediatric Res. 1989, 25, 63–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dingess, K.A.; Valentine, C.J.; Ollberding, N.J.; Davidson, B.S.; Woo, J.G.; Summer, S.; Peng, Y.M.; Guerrero, M.L.; Ruiz-Palacios, G.M.; Ran-Ressler, R.R. Branched-chain fatty acid composition of human milk and the impact of maternal diet: The Global Exploration of Human Milk (GEHM) Study. Am. J. Clin. Nutr. 2017, 105, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Fidler, N.; Sauerwald, T.; Pohl, A.; Demmelmair, H.; Koletzko, B. Docosahexaenoic acid transfer into human milk after dietary supplementation: A randomized clinical trial. J. Lipid Res. 2000, 41, 1376–1383. [Google Scholar] [CrossRef]
- Miliku, K.; Duan, Q.L.; Moraes, T.J.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; Subbarao, P.; Field, C.J.; et al. Human milk fatty acid composition is associated with dietary, genetic, sociodemographic, and environmental factors in the CHILD Cohort Study. Am. J. Clin. Nutr. 2019, 110, 1370–1383. [Google Scholar] [CrossRef]
- Gardner, A.S.; Rahman, I.A.; Lai, C.T.; Hepworth, A.; Trengove, N.; Hartmann, P.E.; Geddes, D.T. Changes in Fatty Acid Composition of Human Milk in Response to Cold-Like Symptoms in the Lactating Mother and Infant. Nutrients 2017, 9, 1034. [Google Scholar] [CrossRef]
- Koletzko, B.; Rodriguez-Palmero, M. Polyunsaturated fatty acids in human milk and their role in early infant development. J. Mammary Gland. Biol. Neoplasia 1999, 4, 269–284. [Google Scholar] [CrossRef]
- Much, D.; Brunner, S.; Vollhardt, C.; Schmid, D.; Sedlmeier, E.-M.; Brüderl, M.; Heimberg, E.; Bartke, N.; Boehm, G.; Bader, B.L.; et al. Breast milk fatty acid profile in relation to infant growth and body composition: Results from the INFAT study. Pediatric Res. 2013, 74, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Mitoulas, L.R.; Gurrin, L.C.; Doherty, D.A.; Sherriff, J.L.; Hartmann, P.E. Infant intake of fatty acids from human milk over the first year of lactation. Br. J. Nutr. 2003, 90, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Kent, J.C.; Mitoulas, L.R.; Cregan, M.D.; Ramsay, D.T.; Doherty, D.A.; Hartmann, P.E. Volume and frequency of breastfeedings and fat content of breast milk throughout the day. Pediatrics 2006, 117, e387–e395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, A.D.; Gay, M.C.L.; Murray, K.; Muhlhausler, B.S.; Wlodek, M.E.; Geddes, D.T. Human Milk Sampling Protocols Affect Estimation of Infant Lipid Intake. J. Nutr. 2020, 150, 2924–2930. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Hernandez, C.; Goeuriot, S.; Giuffrida, F.; Thakkar, S.K.; Destaillats, F. Direct quantification of fatty acids in human milk by gas chromatography. J. Chromatogr. A 2013, 1284, 174–179. [Google Scholar] [CrossRef]
- Mitoulas, L.R.; Kent, J.C.; Cox, D.B.; Owens, R.A.; Sherriff, J.L.; Hartmann, P.E. Variation in fat, lactose and protein in human milk over 24 h and throughout the first year of lactation. Br. J. Nutr. 2002, 88, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.; Gibbs, J.A.; Lyster, R.L.; Baum, J.D. Creamatocrit: Simple clinical technique for estimating fat concentration and energy value of human milk. Br. Med. J. 1978, 1, 1018–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Wu, K.; Yu, Z.; Ren, Y.; Zhao, Y.; Jiang, Y.; Xu, X.; Jin, Y.; Yuan, J.; Li, D. Changes in fatty acid composition of human milk over lactation stages and relationship with dietary intake in Chinese women. Food Funct. 2016, 7, 3154–3162. [Google Scholar] [CrossRef]
- Lattka, E.; Rzehak, P.; Szabó, É.; Jakobik, V.; Weck, M.; Weyermann, M.; Grallert, H.; Rothenbacher, D.; Heinrich, J.; Brenner, H.; et al. Genetic variants in the FADS gene cluster are associated with arachidonic acid concentrations of human breast milk at 1.5 and 6 mo postpartum and influence the course of milk dodecanoic, tetracosenoic, and trans-9-octadecenoic acid concentrations over the duration of lactation. Am. J. Clin. Nutr. 2011, 93, 382–391. [Google Scholar] [PubMed] [Green Version]
- Meyer, B.J. Australians are not Meeting the Recommended Intakes for Omega-3 Long Chain Polyunsaturated Fatty Acids: Results of an Analysis from the 2011-2012 National Nutrition and Physical Activity Survey. Nutrients 2016, 8, 111. [Google Scholar] [CrossRef] [Green Version]
- Innis, S.M. Impact of maternal diet on human milk composition and neurological development of infants. Am. J. Clin. Nutr. 2014, 99, 734s–741s. [Google Scholar] [CrossRef] [Green Version]
- Makrides, M.; Simmer, K.; Neumann, M.; Gibson, R. Changes in the polyunsaturated fatty acids of breast milk from mothers of full-term infants over 30 wk of lactation. Am. J. Clin. Nutr. 1995, 61, 1231–1233. [Google Scholar] [CrossRef]
- Jensen, C.L.; Maude, M.; Anderson, R.E.; Heird, W.C. Effect of docosahexaenoic acid supplementation of lactating women on the fatty acid composition of breast milk lipids and maternal and infant plasma phospholipids. Am. J. Clin. Nutr. 2000, 71, 292s–299s. [Google Scholar] [CrossRef] [Green Version]
- Smit, E.N.; Koopmann, M.; Boersma, E.R.; Muskiet, F.A. Effect of supplementation of arachidonic acid (AA) or a combination of AA plus docosahexaenoic acid on breastmilk fatty acid composition. Prostaglandins Leukot. Essent. Fat. Acids 2000, 62, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Boris, J.; Jensen, B.; Salvig, J.D.; Secher, N.J.; Olsen, S.F. A randomized controlled trial of the effect of fish oil supplementation in late pregnancy and early lactation on the n−3 fatty acid content in human breast milk. Lipids 2004, 39, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, J.A.; Mitoulas, L.R.; Dixon, G.; Doherty, D.A.; Hartmann, P.E.; Simmer, K.; Prescott, S.L. The effects of fish oil supplementation in pregnancy on breast milk fatty acid composition over the course of lactation: A randomized controlled trial. Pediatric Res. 2007, 62, 689–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, A.D.; Gay, M.C.L.; Wlodek, M.E.; Geddes, D.T. The importance of infants’ lipid intake in human milk research. Nutr. Rev. 2021, 79, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, M.; Agostoni, C.; Salari, P.C. The role of lipids in nutrition during the first months of life. J. Int. Med. Res. 1991, 19, 351–362. [Google Scholar] [CrossRef]
- Huang, L.; Lin, J.S.; Aris, I.M.; Yang, G.; Chen, W.Q.; Li, L.J. Circulating Saturated Fatty Acids and Incident Type 2 Diabetes: A Systematic Review and Meta-Analysis. Nutrients 2019, 11, 998. [Google Scholar] [CrossRef] [Green Version]
- Lagou, V.; Scott, R.A.; Manios, Y.; Chen, T.L.; Wang, G.; Grammatikaki, E.; Kortsalioudaki, C.; Liarigkovinos, T.; Moschonis, G.; Roma-Giannikou, E.; et al. Impact of peroxisome proliferator-activated receptors gamma and delta on adiposity in toddlers and preschoolers in the GENESIS Study. Obesity 2008, 16, 913–918. [Google Scholar] [CrossRef]
- Makrides, M.; Neumann, M.; Simmer, K.; Pater, J.; Gibson, R. Are long-chain polyunsaturated fatty acids essential nutrients in infancy? Lancet 1995, 345, 1463–1468. [Google Scholar] [CrossRef]
- Rudolph, M.C.; Young, B.E.; Lemas, D.J.; Palmer, C.E.; Hernandez, T.L.; Barbour, L.A.; Friedman, J.E.; Krebs, N.F.; MacLean, P.S. Early infant adipose deposition is positively associated with the n-6 to n-3 fatty acid ratio in human milk independent of maternal BMI. Int. J. Obes. 2017, 41, 510–517. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Cohort Values (Mean ± SD (Range)) | |
|---|---|
| Maternal age at delivery (years) | 32.0 ± 3.3 (26.0–37.1) |
| Parity (n) | 2.0 ± 1.7 (1.0–3.0) |
| Gestational age (weeks) | 40.1 ± 1.6 (37.7–41.7) |
| 24 h milk intake (mL/day) | 740.8 ± 150.2 (473.0–946.0) |
| Birth | 1 Month | 2 Months | 3 Months | 4 Months | 5 Months | 6 Months | p-Value | |
|---|---|---|---|---|---|---|---|---|
| Maternal weight (kg) | 78.9 ± 18.1 | 78.5 ± 18.6 | 77.7 ± 17.8 | 77.1 ± 17.5 | 76.2 ± 16.9 | 76.5 ± 17.2 | 0.0147 | |
| Maternal BMI (kg/m2) | 28.4 ± 6.2 | 28.2 ± 6.4 | 27.9 ± 6.3 | 27.7 ± 6.2 | 27.4 ± 6.0 | 27.5 ± 6.1 | 0.0108 | |
| Infant weight (kg) | 3.9 ± 0.4 | 4.8 ± 0.6 | 5.6 ± 0.7 | 6.2 ± 0.8 | 6.8 ± 0.9 | 7.4 ± 1.0 | 7.9 ± 0.9 | <0.0001 |
| Infant length (cm) | 52.3 ± 1.4 | 55.4 ± 2.1 | 58.7 ± 2.2 | 61.3 ± 2.2 | 63.7 ± 2.4 | 65.4 ± 2.4 | 67.0 ± 2.4 | <0.0001 |
| Infant w-l z score | 0.0 ± 0.9 | 0.4 ± 0.9 | 0.1 ± 1.1 | 0.0 ± 1.1 | 0.0 ± 1.1 | 0.3 ± 1.1 | 0.4 ± 1.1 | 0.107 |
| Infant HC (cm) | 36.1 ± 0.7 | 38.2 ± 1.0 | 40.0 ± 1.1 | 41.4 ± 0.9 | 42.5 ± 1.3 | 43.5 ± 1.2 | 44.5 ± 1.4 | <0.0001 |
| Infant HC z score | −0.2 ± 0.4 | −0.2 ± 0.6 | 0.0 ± 0.7 | 0.2 ± 0.6 | 0.5 ± 0.7 | 0.7 ± 0.7 | 0.9 ± 0.8 | <0.0001 |
| Infant BMI (kg/m2) | 14.1 ± 1.1 | 15.5 ± 1.1 | 16.2 ± 1.5 | 16.5 ± 1.5 | 16.8 ± 1.7 | 17.3 ± 1.9 | 17.5 ± 1.8 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, A.D.; Gay, M.C.L.; Wlodek, M.E.; Murray, K.; Geddes, D.T. The Fatty Acid Species and Quantity Consumed by the Breastfed Infant Are Important for Growth and Development. Nutrients 2021, 13, 4183. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13114183
George AD, Gay MCL, Wlodek ME, Murray K, Geddes DT. The Fatty Acid Species and Quantity Consumed by the Breastfed Infant Are Important for Growth and Development. Nutrients. 2021; 13(11):4183. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13114183
Chicago/Turabian StyleGeorge, Alexandra D., Melvin C. L. Gay, Mary E. Wlodek, Kevin Murray, and Donna T. Geddes. 2021. "The Fatty Acid Species and Quantity Consumed by the Breastfed Infant Are Important for Growth and Development" Nutrients 13, no. 11: 4183. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13114183