
Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women


1
Department of Microbiology, College of Medicine, Gachon University, Incheon 21999, Korea
2
Lee Gil Ya Cancer and Diabetes Institute, Gachon University, Incheon 21999, Korea
*
Author to whom correspondence should be addressed.
Nutrients 2021, 13(12), 4556; https://0-doi-org.brum.beds.ac.uk/10.3390/nu13124556
Submission received: 6 December 2021
/
Revised: 15 December 2021
/
Accepted: 16 December 2021
/
Published: 20 December 2021
(This article belongs to the Special Issue Adipocyte Metabolism and Health in Postmenopausal Women)
Abstract
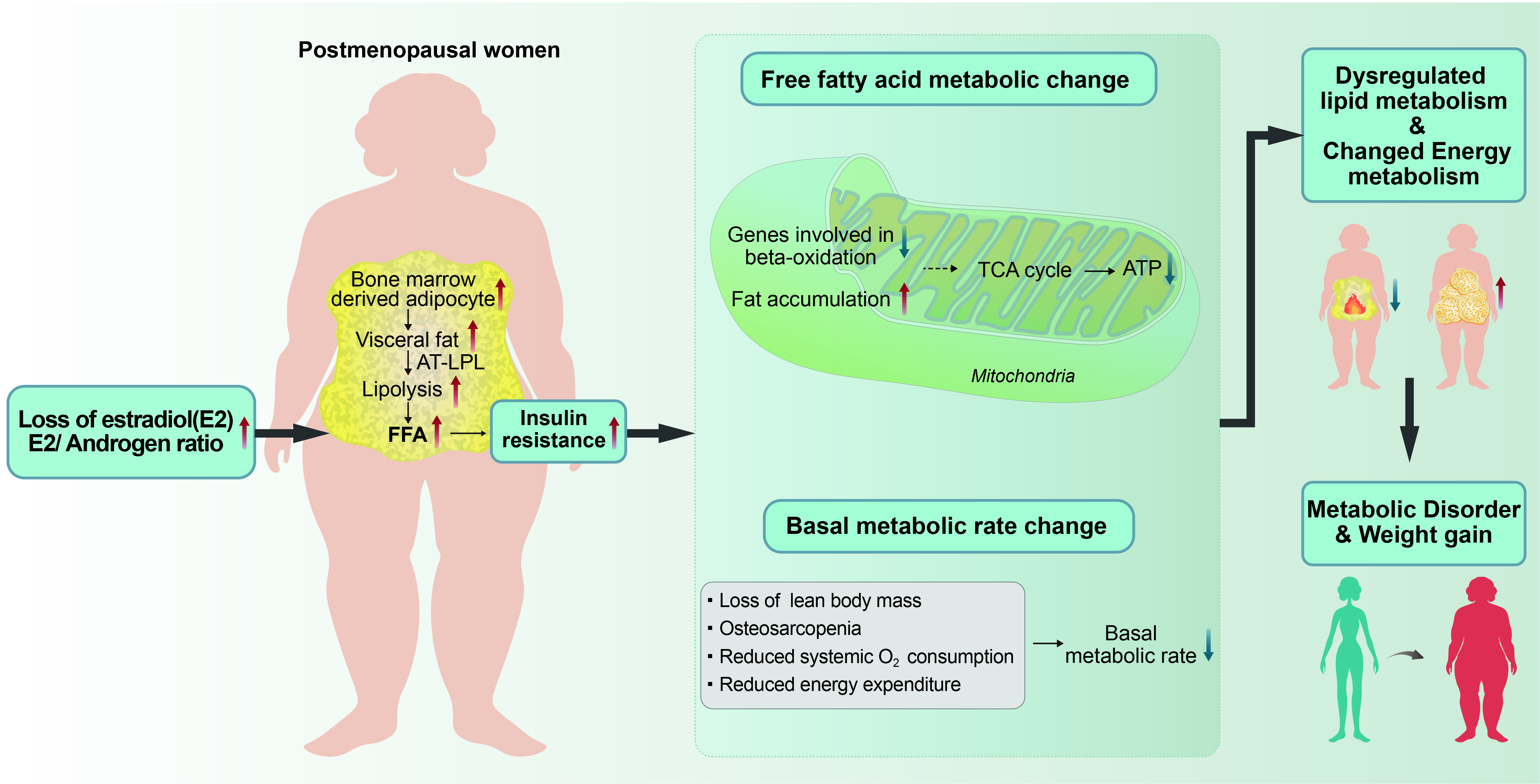
:Aging women experience hormonal changes, such as decreased estrogen and increased circulating androgen, due to natural or surgical menopause. These hormonal changes make postmenopausal women vulnerable to body composition changes, muscle loss, and abdominal obesity; with a sedentary lifestyle, these changes affect overall energy expenditure and basal metabolic rate. In addition, fat redistribution due to hormonal changes leads to changes in body shape. In particular, increased bone marrow-derived adipocytes due to estrogen loss contribute to increased visceral fat in postmenopausal women. Enhanced visceral fat lipolysis by adipose tissue lipoprotein lipase triggers the production of excessive free fatty acids, causing insulin resistance and metabolic diseases. Because genes involved in β-oxidation are downregulated by estradiol loss, excess free fatty acids produced by lipolysis of visceral fat cannot be used appropriately as an energy source through β-oxidation. Moreover, aged women show increased adipogenesis due to upregulated expression of genes related to fat accumulation. As a result, the catabolism of ATP production associated with β-oxidation decreases, and metabolism associated with lipid synthesis increases. This review describes the changes in energy metabolism and lipid metabolic abnormalities that are the background of weight gain in postmenopausal women.

1. Introduction
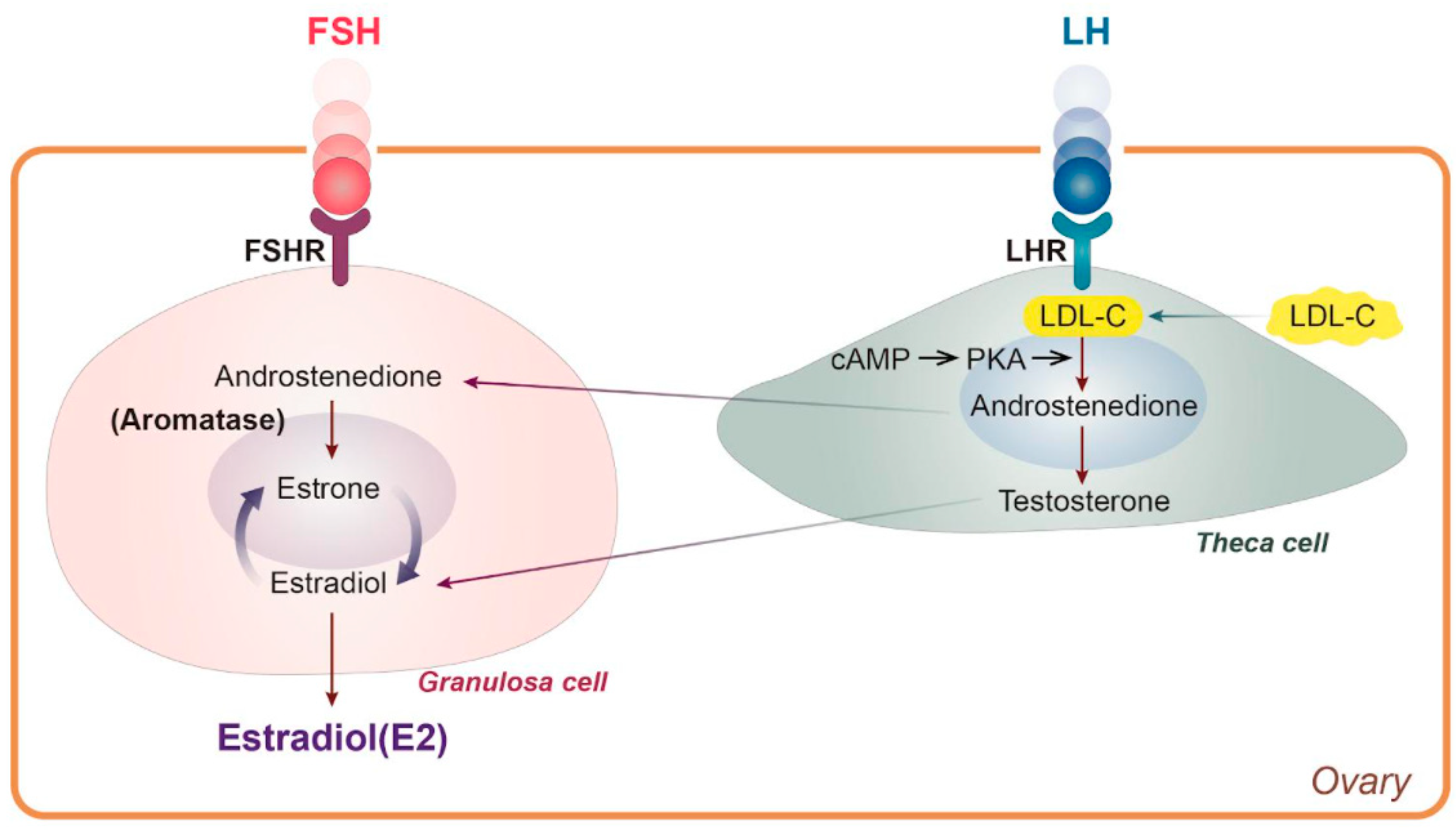
Women generally experience natural menopause due to loss of ovarian follicle activity between the ages of 45 and 55, and surgical menopause is also accompanied by loss of estradiol (E2) [1,2]. In general, women live longer than men, and life expectancy is increasing worldwide. The median age of women will reach 82 in developed countries by 2025 [3]. Thus, about half of a woman’s life is after menopause. It is well known that hormonal changes are one of the significant physiological effects of menopause. There are three types of estrogen—estrone (E1), E2, and estriol (E3)—all of which are C18 steroids and aromatic molecules [4]. Each form has a unique function that matches a woman’s life cycle characteristics, such as reproductive age, pregnancy, and menopause [5]. Before menopause and throughout the menstrual cycle, meningeal cells produce androstenedione, which acts as a metabolic precursor to E1 and testosterone in the ovaries and peripheral tissues [6]. In granulosa cells, androstenedione is converted to E1 by aromatase activity of CYP 19, after which E3 is converted to E2, with an average level of total estrogen of 100-250 pg/mL [6] (Figure 1). By contrast, circulating E2 levels drop sharply to 10 pg/mL in postmenopausal women [7], meaning that women spend half of their lives in estrogen deficiency.
With these hormonal changes, menopausal women are more likely to experience various metabolic disorders such as dysregulated lipid metabolism, fat redistribution, visceral fat accumulation, and altered fatty acid metabolism [8,9,10]. Furthermore, they are readily accompanied by changes in body composition and energy metabolism, loss of muscle volume and strength, and weight gain [11,12,13].
To better understand metabolic alterations, it is important to understand recommended body weight, body composition, and the basic concepts of energy metabolism, including basal metabolism, resting energy expenditure (REE), and metabolic rate. This review describes the concepts needed to understand energy metabolism, including the composition of the human body and the components of energy expenditure, and discusses possible changes in energy metabolism in postmenopausal women. By summarizing overall energy metabolism, this review provides information for preventing muscle and bone loss. In addition, it also addresses lipid metabolic changes related to visceral fat accumulation, lipolysis, fatty acid oxidation, fat redistribution, and weight gain.
2. Menopause Associated Changes in Energy Metabolism
2.1. Composition of the Human Body
The chemical compositions of the male and female human bodies are shown in Table 1, based on average physical dimensions from the measurements of thousands of subjects who participated in various anthropometric and nutrition surveys [14]. As seen in Table 1, the muscles of the reference man account for 44.8% of body weight, compared to 36% for women. The reference man has 15% total body fat versus the female’s 27%. The reference woman’s storage fat accounts for 15% of her weight, and essential fat accounts for 12%. The total amount of our body fat is composed of storage and essential fats. The latter is present in lipid-rich tissues throughout the central nervous system, bone marrow, heart, lungs, liver, kidneys, spleen, intestines, and muscles [15]. In females, essential fat also includes fat in mammary glands and the pelvic region. Fat that accumulates in adipose tissue is called storage fat, and triglyceride (triacylglycerol) is the typical component, which accounts for ~86% of body fat [16]. Triglyceride can be broken down into fatty acids and glycerol by lipase and used to produce ATP for energy metabolism [13]. In other words, triglyceride can be used as an energy source for heat generation in the human body. Fat mass is, therefore, the most changeable constituent of the body. In terms of muscle and body fat mass, the body compositions of men and women are distinctly different, with women having more fat and less muscle mass [17].
Changes in Body Composition in Postmenopausal Women
As women age, they tend to have an increased amount of fat tissue in the abdomen and relatively reduced fat in the hip-thigh area. Changes in body composition are also involved, including reductions of fat-free mass (FFM) and lean body mass (LBM) [18]. Women who experience natural menopause have changes in body fat mass, such as a decrease in total leg fat and an increase in abdominal fat [19]. It is likely that these changes are at least in part due to hormonal changes that occur when women have high levels of androgens versus E2 after menopause [20]. Several researchers have shown that menopause itself influences upper body fat distribution independent of aging [21,22,23]. However, many cross-sectional studies using dual-energy X-ray absorptiometry showed that postmenopausal women have lower FFM or LBM in the whole body, trunk, and lower extremities than premenopausal women [24]. In a 6-year follow-up longitudinal study, natural postmenopausal women lost more FFM than age-matched premenopausal women while showing increased central adiposity and reduced energy expenditure during rest and physical activity [25]. Interestingly, one out of five relatively healthy Korean postmenopausal women aged over 65 years exhibited a decline in muscle mass, and 7.6% of subjects showed declines in both muscle mass and strength [26]. The study also reported the intensified loss of skeletal muscle with aging [27].
2.2. Component of Energy Expenditure
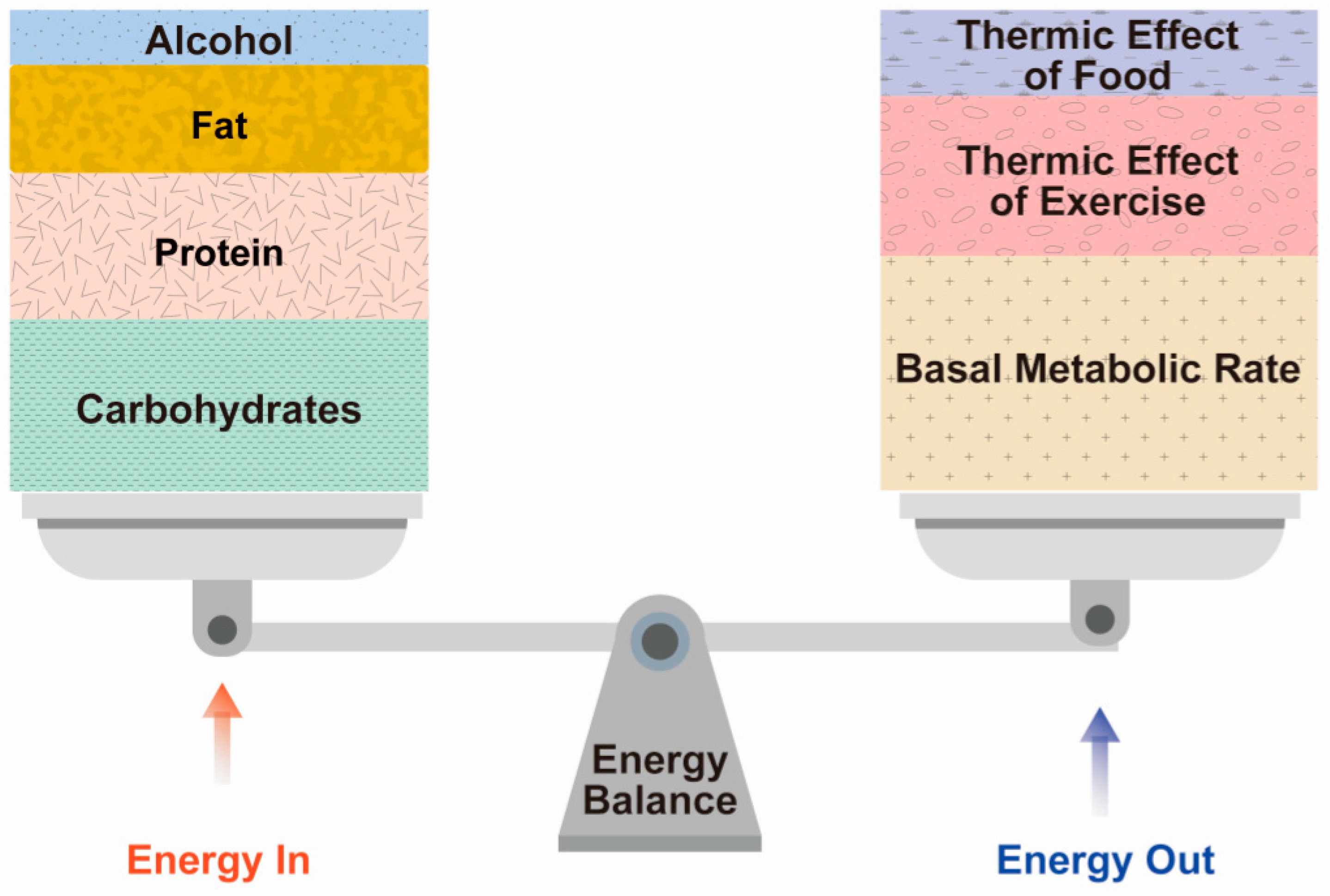
Menopausal women are likely to experience changes in lipid metabolism along with weight gain [8]. To understand why this occurs, it is necessary to know the basic concepts of energy expenditure that influence body weight and composition, the function of the basal metabolic rate (BMR), the thermic effect of food, and physical activity. Total energy expenditure consists of (1) the BMR or REE, (2) the thermic effect of food (TEF), and (3) the thermic effect of activity (TEA) (Figure 2).
2.2.1. BMR
Basal metabolism is the energy expended by internal processes during a period of complete rest in a climate-controlled environment at least 10–12 h after consumption of the most recent meal; that is, the minimum amount of energy needed to sustain life processes [28]. Typically, 50–65% of total energy expenditure is attributable to the BMR that is commonly used with REE, which is simply the basal metabolism during a non-active state in a climate-controlled environment at least 10 to 12 h after the consumption of the most recent meal (Figure 2) [29]. However, the most important difference between the BMR and REE is that a subject does not need to fast to measure REE.
2.2.2. TEF
The TEF refers to the metabolic reaction of food (i.e., heat generation) due to its digestion, absorption, processing, and storage [30]. TEF can increase metabolism by more than 5–15% compared to the BMR when consumed in large portions rather than small frequent meals, when carbohydrates and protein rather than dietary fat are consumed, and when a low-fat plant-based diet is followed [31]. Generally, TEF is estimated as 10% of total energy intake during a particular period (Figure 2) [32]. For example, TEF can be estimated at 160 kilocalories for an individual who consumes a mixed diet containing 1600 kilocalories over a 24-h period.
2.2.3. TEA
The TEA refers to skeletal muscle activity associated with the maintenance of position and posture, as well as skeletal activity during obvious movements such as walking, running, swimming, climbing stairs, or vacuuming [28]. Thus, TEA is a highly changeable component. While the contribution of skeletal muscle activity may seem trivial, simply sitting on a chair without back support augments heat generation by 3–5% [28]. This increase in metabolism is much more significant when standing [33]. Average physical activity accounts for 20–40% of total energy expenditure, but this depends on the individual’s physical activity (Figure 2) [34]. Therefore, sedentary people who are less active consume much less energy and may have lower energy than more physically active people [35]. These results indicate that a sedentary lifestyle before menopause may contribute to a decrease in total energy expenditure [36].
2.3. Metabolic Rate Difference between Skeletal Muscle and Adipose Tissue
The human body consists of protein, water, minerals, and fat, which can be largely divided into fat mass and FFM [37]. FFM is often conflated with LBM, which is calculated by subtracting fat mass weight from total body weight. LBM is the weight of internal organs, skin, bones, body water, tendons, and muscle mass [38]. The human body is composed of various tissues and organs, each of which has a specific function and mass, resulting in a different contribution to the BMR. FFM or skeletal muscle accounts for approximately 60–85% of the body mass; it is regarded as an energy consumer and a major determinant of the BMR or REE. Therefore, the metabolic energy rate differs depending on the body composition ratio [34]. In this regard, it is necessary to consider the difference between the energy consumptions of skeletal muscle and adipose tissue, which are 13 kcal/kg/day and 4.5 kcal/kg/day, respectively (Table 2) [39]. The energy expenditure of skeletal muscle is three times higher than that of adipose tissue, so the BMR is higher for individuals with high muscle mass, even if they have the same weight as a person with low muscle mass. During aging, fat increases at the expense of FFM; moreover, the loss of LBM due to sarcopenia (a decrease in skeletal muscle) and an increase in adipose tissue will result in a lower BMR [40].
2.3.1. Changes in Body Composition and Energy Expenditure in Postmenopausal Women
As aging progresses, FFM or skeletal muscle loss occurs, which leads to a decrease in the BMR [41,42]. Considering that the metabolic rate of skeletal muscle is more than three times higher than that of adipose tissue, loss of skeletal muscle or FFM may lead to a decrease in BMR among menopausal period women [43,44]. The decrease in E2, along with changes in body composition in postmenopausal women, plays an important role in regulating adipocyte differentiation and distribution. Gavin et al. reported that ovarian hormone depletion after ovariectomy (OVX) increased the production of bone marrow-derived adipocytes (BMDAs) in mice visceral fat depots that are gonad adipose depots [45]. In addition, E2 replacement continued to dampen the accelerated production of BMDAs. Furthermore, estrogen receptor (ER)α genetic knockdown enhanced BMDA production in both the gonadal and inguinal depots, which demonstrated that E2 regulates BMDA production [45]. If this is translatable across species, it suggests that the production of BMDAs occurs through a mechanism where visceral fat increases in estrogen-deficient postmenopausal women.
Estrogen plays a pivotal role in systemic energy homeostasis. OVX mice have been shown to reduce systemic O2 consumption and energy expenditure, leading to the weight gain associated with increased body fat. On the other hand, exogenous E2 supplementation increased systemic O2 consumption and energy expenditure, resulting in increased systemic insulin sensitivity in OVX mice [45]. This provides a mechanism by which E2 supplementation may ameliorate insulin resistance [45]. With regard to energy homeostasis, it was also reported that E2 directly regulates mitochondria membrane biophysical properties and bioenergetic functions, providing a direct pathway by which E2 states broadly influence energy homeostasis [46,47]. OVX mice exhibit decreased mitochondrial respiratory function, cellular redox state, and insulin sensitivity in skeletal muscle [46]. E2 may be a mitochondrial membrane component in many tissues that locally affects bioenergetic activity and energy homeostasis. Additionally, Gavin et al. showed that reducing endogenous E2 in humans reduced energy expenditure and increased visceral fat [45]. It was also reported that estrogen deficiency increased BMDA accumulation in the white adipose tissue of mice and was associated with decreased physical activity in women [48].
Postmenopausal women may experience body shape changes due to increased loss of FFM or LBM with high energy metabolisms, while fat tissue increases with low energy metabolism and increased BDMAs [45]. Because the E2 state has a widespread impact on energy homeostasis, a decrease in levels of this hormone reduces systemic O2 consumption and energy consumption. Postmenopausal women’s bodies also consume less energy to maintain their basic life processes [49]. Collectively, these changes may affect the BMR and lead to insulin resistance along with weight gain.
2.3.2. Sarcopenia in Postmenopausal Women
In general, skeletal muscle mass and muscle strength peak in the mid-20s and 30s and then gradually decrease [50]. Sarcopenia refers to the degenerative loss of skeletal muscle that occurs at a rate of 3–8% every 10 years after the age of 30 years and accelerates with age [8]. This condition is associated with increased risks of functional disability, falls, fractures, and overall mortality among the elderly [51]. Women develop sarcopenia earlier than men, and the decline of skeletal muscle mass and strength accelerates with the onset of menopause [52,53]. When sarcopenia coexists with osteoporosis, it results in a geriatric syndrome called “osteosarcopenia,” which increases the risk of weakness, hospitalization, and death [54]. In a prospective cross-sectional study by Buliana et al., the prevalence of osteosarcopenia was reported to be high among postmenopausal women with an increased risk of fracture [55]. Patients in the osteosarcopenia group had a greater risk of frailty than patients in the osteoporosis-alone group (odds ratio, 2.33; 95% confidence interval, 1.13–4.80; and p = 0.028). According to a retrospective observational study, one-year mortality of osteosarcopenia (15.1%) was higher than that of other groups (normal, 7.8%; osteoporosis only, 5.1%; and sarcopenia only, 10.3%) [56]. In postmenopausal women, insufficient protein and calcium intake and low levels of physical activity appear to be the most common risk factors for osteosarcopenia [55]. Generally, it is accepted that menopause is associated with accelerated loss of FFM or skeletal muscle, which further decreases energy expenditure during rest and physical activity [57]. Thus, body function impairment originates from muscle loss in menopausal women, leading to difficulties in carrying out voluntary activities and reducing the quality of life. The most promising strategy for increasing muscle and bone mass is resistance training, as well as sufficient amounts of protein, vitamin D, calcium, and creatine to help preserve these tissues during menopause [53].
3. Background of Weight Gain in Postmenopausal Women
3.1. Lipid Metabolic Abnormality Due to E2 Hormonal Change
Typically, women spontaneously experience menopause between the ages of 45 and 55 because of decreased ovarian follicular activity [1]. Menopause does not happen all at once; rather, it progresses through a transition period to the postmenopausal stage. With regard to surgical menopause, bilateral oophorectomy has been shown to cause dyslipidemia and significant loss of bone density within one year [2]. Around this time, the female hormones undergo drastic alterations. One of the major physiological changes associated with menopause is a sharp decrease in E2 that contributes to lipid metabolic disorders. Notably, women tend to develop more cardiovascular disease after menopause due to estrogen deficiency and alterations in lipid metabolism [58]. The reason is due to the unique role of E2 that is synthesized using low-density lipoprotein cholesterol (LDL-C) in the ovary (Figure 1). Therefore, a decrease in E2 synthesis due to menopause means that LDL-C is no longer used for synthesizing E2, so it remains in the systemic circulation (Figure 1). Since postmenopausal women have high LDL-C levels, there is an increased risk of metabolic syndrome symptoms, including central obesity, insulin resistance, dyslipidemia, hypertension, and cardiovascular disease [59].
3.2. Fat Redistribution in Postmenopausal Women
Most women tend to experience changes in the composition of the body as they get older, and this period almost perfectly coincides with menopause [60]. In premenopausal women, adipose tissue is predominantly distributed in the gluteal femoral subcutaneous compartment, whereas postmenopausal women tend to exhibit higher total body fat mass, fat percentage, and accumulation of central fat [61]. This can be partly explained by the changes in circulating endogenous sex hormone levels, because estrogen androgen receptors are expressed in both visceral and subcutaneous adipocytes [62,63]. Estrogens bind to ERα and ERβ, and androgens bind to the androgen receptor, enabling sex hormones act on their target cells [64]. Therefore, the reduction of the circulating sex hormones will change their action in target cells.
Ovarian estrogens induce peripheral fat storage mainly in the gluteal and femoral subcutaneous regions that express ERα, which mediates lipoprotein lipase activity and triacylglycerol accumulation in adipocytes [65]. On the other hand, androgens—Primarily bioavailable testosterones—Augment visceral abdominal fat accumulation [63]. In postmenopausal women, the concentration of E2 in circulation decreases, so the androgen to estrogen ratio increases [8]. Therefore, relative androgen excess (a higher baseline testosterone/E2 ratio) causes weight gain and body fat redistribution in postmenopausal women [63,66]. According to the longitudinal, community-based, 5-year follow-up Study of Women’s Health Across the Nation, postmenopausal women showed twice as much visceral abdominal fat and subcutaneous adipose tissue than premenopausal women [67]. However, testosterone levels were similar among pre-and postmenopausal women [68], which suggested that fat redistribution may be affected by a marked decrease in estrogen levels as opposed to testosterone levels. Moreover, it was also observed that the androgen to estrogen ratio was also elevated in premenopausal women with polycystic ovarian syndrome [69,70].
3.3. Excessive Visceral Abdominal Fat and Metabolic Alterations
The basic role of adipocytes in lipid metabolism is storing energy in adipose tissue in the form of triacylglycerol and releasing it as free fatty acids (FFAs) as needed to provide fuel for working muscles [71]. Adipocytes also control glucose homeostasis by secreting glycerol and fatty acids, which play critical roles in hepatic and peripheral glucose homeostasis by mediating the breakdown of triacylglycerol [72,73]. However, as women experience menopause, dysregulated adipocyte metabolism occurs due to estrogen reduction, and various metabolic diseases appear [8].
Visceral adipose tissue in both humans and rats is composed of mesenteric, retroperitoneal, osteoporosis, and reproductive gland deposits, so rodent models have been widely used to simulate fat metabolism in the human body [74]. In animal models such as OVX and ERα knockout mice, loss of ovarian hormones has been demonstrated to increase BMDA production in the visceral fat depot and gonadal adipose depot [45]. ERα plays an important role in regulating the de novo synthesis of BMDA, which supports the observation of increased visceral fat in estrogen-deficient postmenopausal women [45]. In this setting, visceral fat increases, and abdominal obesity intensifies in obese menopause women. Since the rate of lipolysis differs throughout the body, abdominal fat has a higher rate of lipolysis than gluteal fat due to the higher rate of catecholamine-mediated lipolysis in the abdomen [66,75].
3.4. Alterations in Fatty Acid Metabolism
It is widely known that FFAs are produced by the excessive decomposition of visceral fat, which promotes hepatic insulin resistance in connection with the increased flow of FFAs to the liver [76]. Interestingly, in a metabolomic study comparing the evaluation of fatty acid metabolism between pre-and postmenopausal women, fatty acid metabolites such as heptanoate, octanoate, and pelargonate were significantly higher in the visceral fat but not in the subcutaneous fat of postmenopausal women. This means that increased lipolysis of visceral fat may trigger an accumulation of fatty acid metabolites [77].
Extensive studies have been conducted on the role of estrogen in various metabolisms, immunity, and inflammatory processes in rodents and humans [78]. Thanks to more sensitive and accurate proteomic techniques, a number of proteins and pathways in visceral fat were found in OVX rodent models [79]. Interestingly, Boldarine et al. reported that OVX induced the upregulation of genes related to lipogenesis and downregulation of genes related to fatty acid oxidation [10]. It was reported that expression of the adipogenesis-associated gene all-trans-retinol 13,14-reductase (RETSAT) was increased in OVX mice. RETSAT is induced during adipocyte differentiation and is positively regulated by the transcription factor, peroxisome proliferator-activated receptor γ, which means that upregulated RETSAT indicates active fat accumulation in OVX mice [80]. Moreover, adipose tissue lipoprotein lipase (AT-LPL) showed increased expression, indicating a high ability of retroperitoneal adipose tissue to absorb lipoprotein-derived FFAs. AT-LPL is an enzyme that decomposes triglyceride into FFAs for absorption and storage by adipocytes, and plays an important role in fat accumulation and fat storage distribution. Therefore, it can be expected that increased uptake of lipoprotein-derived FFA in retroperitoneal adipose tissue due to enhanced expression of AT-LPL could be expected to affect the TG synthesis pool in OVX mice [72]. Estrogens are also associated with inflammatory responses in women. Although estrogens are known to enhance autoimmune diseases [81], decreased estrogen increases susceptibility to infectious diseases, as demonstrated by the defective innate immune responses against viral infection in OVX mice [82]. In addition, OVX enhanced the susceptibility of female rats to dyslipidemia with a decrease in innate cytokines, suggesting impaired metabolic and immune homeostatic responses with the loss of estrogens [83].
A pathway analysis study reported that OVX affected the fatty acid metabolism/mitochondrial fatty-acid-oxidation pathway and fatty acyl coenzyme A (CoA) biosynthesis pathway in visceral adipose tissue [10]. Fatty acid catabolism mainly occurs in the mitochondria [84]. Long-chain fatty acids with more than 14 carbons are converted into fatty acyl-CoA and pass through the mitochondrial membrane [85]. Fatty acid catabolism begins with acyl-CoA synthetase that adds CoA to fatty acids using ATP in the cytoplasm [85]. Once it has passed through the mitochondrial membrane, acyl-CoA begins the beta-oxidation process [85]. FFA activation occurs by bonding with CoA and is promoted by ligase enzymes, which are important steps in the oxidation and synthesis of triacylglycerol and other lipids [86]. Fatty acid oxidation is the largest contributor to ATP production, accounting for 40–60% [87]. Therefore, a decrease in fatty acid oxidation leads to the accumulation of lipids in a state that the body cannot efficiently burn as a fuel source, resulting in insulin resistance [10,88].
Taken together, the loss of estrogen and increase in circulating androgens in postmenopausal women induce changes in body fat distribution, leading to abdominal obesity. In addition, loss of estrogen increases BMDA production in mice visceral fat depots, which are gonad adipose depots [45]. As such, obese menopausal women have increased visceral fat and aggravated abdominal obesity. The rate of lipolysis is higher in abdominal fat than in gluteal fat [77], which triggers excess FFA production from excessive visceral fat breakdown, promotes insulin resistance, and leads to metabolic diseases [76]. In the OVX model, the expression of genes related to β-oxidation and lipogenesis is decreased and increased, respectively. Therefore, excessive fatty acids produced by the breakdown of visceral fat cannot be efficiently oxidized as a fuel source in the body through β-oxidation, which leads to fat accumulation. This can cause unfavorable changes in both fat metabolism and energy metabolism [10].
4. Conclusions
Both natural and surgical menopause is accompanied by changes in body composition due to loss of E2 secretion, and various changes can occur in energy and lipid metabolism. Postmenopausal sarcopenia and increased fat mass change the energy metabolic rate and affect the BMR. In addition, postmenopausal women are susceptible to obesity, and weight loss becomes more difficult. Moreover, fat redistribution caused by hormonal changes leads to changes in body shape. In particular, as the amount of visceral fat increases, FFAs also increase due to excessive fat decomposition, which can lead to insulin resistance and cause metabolic diseases. In addition, excessive FFAs produced by the lipolysis of visceral fat after menopause are not properly used as energy sources through β-oxidation, which is because genes related to β-oxidation are downregulated following the loss of E2. In this review, energy metabolism and lipid metabolic disorders related to menopause have been comprehensively summarized in relation to the basic concepts of body composition and energy expenditure. An understanding of overall energy metabolism, osteosarcopenia, and a recognition of the importance of preventing muscle loss will help to address individual health needs. This review also systematically organized the background of weight gain due to E2 loss and lipid metabolic abnormalities. However, we did not discuss physical activity and nutritional therapy, which are prominent factors influencing metabolic changes. The relation between menopausal status and overall energy and lipid metabolism, particularly fatty acid oxidation, must be clarified to prevent the growing problem of obesity-related disorders, including insulin resistance and metabolic syndrome.
Author Contributions
Conceptualization, S.-H.K. and Y.J.; investigation, S.-H.K. and Y.J.; writing—original draft preparation, S.-H.K.; writing—review and editing, S.-H.K. and Y.J.; supervision, Y.J. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (No. NRF-2021R1A5A2030333).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Landgren, B.M.; Collins, A.; Csemiczky, G.; Burger, H.G.; Baksheev, L.; Robertson, D.M. Menopause transition: Annual changes in serum hormonal patterns over the menstrual cycle in women during a nine-year period prior to menopause. J. Clin. Endocrinol. Metab. 2004, 89, 2763–2769. [Google Scholar] [CrossRef]
- Yoshida, T.; Takahashi, K.; Yamatani, H.; Takata, K.; Kurachi, H. Impact of surgical menopause on lipid and bone metabolism. Climacteric 2011, 14, 445–452. [Google Scholar] [CrossRef]
- Takahashi, T.A.; Johnson, K.M. Menopause. Med. Clin. North Am. 2015, 99, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. Disruption of aromatase homeostasis as the cause of a multiplicity of ailments: A comprehensive review. J. Steroid Biochem. Mol. Biol. 2017, 168, 19–25. [Google Scholar] [CrossRef]
- Cui, J.; Shen, Y.; Li, R. Estrogen synthesis and signaling pathways during aging: From periphery to brain. Trends Mol. Med. 2013, 19, 197–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewailly, D.; Robin, G.; Peigne, M.; Decanter, C.; Pigny, P.; Catteau-Jonard, S. Interactions between androgens, FSH, anti-Müllerian hormone and estradiol during folliculogenesis in the human normal and polycystic ovary. Hum. Reprod. Update 2016, 22, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Cervellati, C.; Bergamini, C.M. Oxidative damage and the pathogenesis of menopause related disturbances and diseases. Clin. Chem. Lab. Med. 2016, 54, 739–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, S.H.; Kim, H.S. Menopause-Associated Lipid Metabolic Disorders and Foods Beneficial for Postmenopausal Women. Nutrients 2020, 12, 202. [Google Scholar] [CrossRef] [Green Version]
- Ozbey, N.; Sencer, E.; Molvalilar, S.; Orhan, Y. Body fat distribution and cardiovascular disease risk factors in pre-and postmenopausal obese women with similar BMI. Endocr. J. 2002, 49, 503–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldarine, V.T.; Pedroso, A.P.; Brandão-Teles, C.; LoTurco, E.G.; Nascimento, C.M.O.; Oyama, L.M.; Bueno, A.A.; Martins-de-Souza, D.; Ribeiro, E.B. Ovariectomy modifies lipid metabolism of retroperitoneal white fat in rats: A proteomic approach. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E427–E437. [Google Scholar] [CrossRef] [PubMed]
- Maltais, M.L.; Desroches, J.; Dionne, I.J. Changes in muscle mass and strength after menopause. J. Musculoskelet. Neuronal Interact. 2009, 9, 186–197. [Google Scholar]
- Kapoor, E.; Collazo-Clavell, M.L.; Faubion, S.S. Weight Gain in Women at Midlife: A Concise Review of the Pathophysiology and Strategies for Management. Mayo Clin. Proc. 2017, 92, 1552–1558. [Google Scholar] [CrossRef]
- Pannemans, D.L.; Westerterp, K.R. Energy expenditure, physical activity and basal metabolic rate of elderly subjects. Br. J. Nutr. 1995, 73, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Gropper, S.S.; Smith, J.L. Advanced Nutrition and Human Metabolism, 4th ed.; Howe, E., Ed.; Cengage Learning: Boston, MA, USA, 2005. [Google Scholar]
- Lemos, T.; Gallagher, D. Current body composition measurement techniques. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 310–314. [Google Scholar] [CrossRef]
- Luke, A.; Schoeller, D.A. Basal metabolic rate, fat-free mass, and body cell mass during energy restriction. Metabolism 1992, 41, 450–456. [Google Scholar] [CrossRef]
- Choi, B.; Steiss, D.; Garcia-Rivas, J.; Kojaku, S.; Schnall, P.; Dobson, M.; Baker, D. Comparison of body mass index with waist circumference and skinfold-based percent body fat in firefighters: Adiposity classification and associations with cardiovascular disease risk factors. Int. Arch. Occup. Environ. Health 2016, 89, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Greendale, G.A.; Sternfeld, B.; Huang, M.; Han, W.; Karvonen-Gutierrez, C.; Ruppert, K.; Cauley, J.A.; Finkelstein, J.S.; Jiang, S.F.; Karlamangla, A.S. Changes in body composition and weight during the menopause transition. JCI Insight 2019, 4, e124865. [Google Scholar] [CrossRef]
- Ambikairajah, A.; Walsh, E.; Tabatabaei-Jafari, H.; Cherbuin, N. Fat mass changes during menopause: A metaanalysis. Am. J. Obstet. Gynecol. 2019, 221, 393–409.e350. [Google Scholar] [CrossRef]
- Janssen, I.; Powell, L.H.; Jasielec, M.S.; Kazlauskaite, R. Covariation of change in bioavailable testosterone and adiposity in midlife women. Obesity 2015, 23, 488–494. [Google Scholar] [CrossRef]
- Kaye, S.A.; Folsom, A.R.; Prineas, R.J.; Potter, J.D.; Gapstur, S.M. The association of body fat distribution with lifestyle and reproductive factors in a population study of postmenopausal women. Int. J. Obes. 1990, 14, 583–591. [Google Scholar] [CrossRef]
- Pasquali, R.; Casimirri, F.; Labate, A.M.; Tortelli, O.; Pascal, G.; Anconetani, B.; Gatto, M.R.; Flamia, R.; Capelli, M.; Barbara, L. Body weight, fat distribution and the menopausal status in women. The VMH Collaborative Group. Int. J. Obes. Relat. Metab. Disord. 1994, 18, 614–621. [Google Scholar]
- Ijuin, H.; Douchi, T.; Oki, T.; Maruta, K.; Nagata, Y. The contribution of menopause to changes in body-fat distribution. J. Obstet. Gynaecol. Res. 1999, 25, 367–372. [Google Scholar] [CrossRef]
- Douchi, T.; Kuwahata, R.; Yamasaki, H.; Yamamoto, S.; Oki, T.; Nakae, M.; Nagata, Y. Inverse relationship between the changes in trunk lean and fat mass during gonadotropin-releasing hormone agonist therapy. Maturitas 2002, 42, 31–35. [Google Scholar] [CrossRef]
- Poehlman, E.T.; Toth, M.J.; Gardner, A.W. Changes in energy balance and body composition at menopause: A controlled longitudinal study. Ann. Intern. Med. 1995, 123, 673–675. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Lee, D.-C. Muscle strength and quality are associated with severity of menopausal symptoms in peri- and post-menopausal women. Maturitas 2013, 76, 88–94. [Google Scholar] [CrossRef]
- Lee, E.S.; Park, H.M. Prevalence of Sarcopenia in Healthy Korean Elderly Women. J. Bone Metab. 2015, 22, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Medeiros Denis, M.; Wildman, R.E.C. Advanced Human Nutrition, 4th ed.; Sheehan, S., Ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2019. [Google Scholar]
- Tur, J.A.; Bibiloni, M.D.M. Anthropometry, Body Composition and Resting Energy Expenditure in Human. Nutrients 2019, 11, 1891. [Google Scholar] [CrossRef] [Green Version]
- Denzer, C.M.; Young, J.C. The effect of resistance exercise on the thermic effect of food. Int. J. Sport Nutr. Exerc. Metab. 2003, 13, 396–402. [Google Scholar] [CrossRef]
- Calcagno, M.; Kahleova, H.; Alwarith, J.; Burgess, N.N.; Flores, R.A.; Busta, M.L.; Barnard, N.D. The Thermic Effect of Food: A Review. J. Am. Coll. Nutr. 2019, 38, 547–551. [Google Scholar] [CrossRef]
- Shaw, E.; Leung, G.K.W.; Jong, J.; Coates, A.M.; Davis, R.; Blair, M.; Huggins, C.E.; Dorrian, J.; Banks, S.; Kellow, N.J.; et al. The Impact of Time of Day on Energy Expenditure: Implications for Long-Term Energy Balance. Nutrients 2019, 11, 2383. [Google Scholar] [CrossRef] [Green Version]
- Welle, S.; Nair, K.S. Relationship of resting metabolic rate to body composition and protein turnover. Am. J. Physiol. 1990, 258, E990–E998. [Google Scholar] [CrossRef]
- Sareen, S.; Gropper, J.L.S.; Groff, J.L. Advanced Nutrition and Human Metabolism, 4th ed.; Howe, E., Ed.; Wsdsworth, Inc.: Belmont, CA, USA, 2005; pp. 159–161. [Google Scholar]
- Thivel, D.; Tremblay, A.; Genin, P.M.; Panahi, S.; Rivière, D.; Duclos, M. Physical Activity, Inactivity, and Sedentary Behaviors: Definitions and Implications in Occupational Health. Front. Public Health 2018, 6, 288. [Google Scholar] [CrossRef]
- Colpani, V.; Oppermann, K.; Spritzer, P.M. Association between habitual physical activity and lower cardiovascular risk in premenopausal, perimenopausal, and postmenopausal women: A population-based study. Menopause 2013, 20, 525–531. [Google Scholar] [CrossRef]
- Schubert, C.M.; Rogers, N.L.; Remsberg, K.E.; Sun, S.S.; Chumlea, W.C.; Demerath, E.W.; Czerwinski, S.A.; Towne, B.; Siervogel, R.M. Lipids, lipoproteins, lifestyle, adiposity and fat-free mass during middle age: The Fels Longitudinal Study. Int. J. Obes. 2006, 30, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.N.; Kauffman, T.G.; Cooney, P.T.; Ramseur, K.R.; Brown, L.M. Comparison of DEXA and QMR for assessing fat and lean body mass in adult rats. Physiol. Behav. 2011, 103, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ying, Z.; Bosy-Westphal, A.; Zhang, J.; Heller, M.; Later, W.; Heymsfield, S.B.; Müller, M.J. Evaluation of specific metabolic rates of major organs and tissues: Comparison between men and women. Am. J. Hum. Biol. 2011, 23, 333–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polotsky, H.N.; Polotsky, A.J. Metabolic implications of menopause. Semin. Reprod. Med. 2010, 28, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Lazzer, S.; Bedogni, G.; Lafortuna, C.L.; Marazzi, N.; Busti, C.; Galli, R.; De Col, A.; Agosti, F.; Sartorio, A. Relationship between basal metabolic rate, gender, age, and body composition in 8,780 white obese subjects. Obesity 2010, 18, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Sabounchi, N.S.; Rahmandad, H.; Ammerman, A. Best-fitting prediction equations for basal metabolic rate: Informing obesity interventions in diverse populations. Int. J. Obes. 2013, 37, 1364–1370. [Google Scholar] [CrossRef] [Green Version]
- Kalyani, R.R.; Corriere, M.; Ferrucci, L. Age-related and disease-related muscle loss: The effect of diabetes, obesity, and other diseases. Lancet Diabetes Endocrinol. 2014, 2, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Wildman, R.E.; Medeiros, D.M. Advanced Human Nutrition, 4th ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2019; p. 212. [Google Scholar]
- Gavin, K.M.; Sullivan, T.M.; Kohrt, W.M.; Majka, S.M.; Klemm, D.J. Ovarian Hormones Regulate the Production of Adipocytes From Bone Marrow-Derived Cells. Front. Endocrinol. 2018, 9, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.J.; Kew, K.A.; Ryan, T.E.; Pennington, E.R.; Lin, C.T.; Buddo, K.A.; Fix, A.M.; Smith, C.A.; Gilliam, L.A.; Karvinen, S.; et al. 17β-Estradiol Directly Lowers Mitochondrial Membrane Microviscosity and Improves Bioenergetic Function in Skeletal Muscle. Cell Metab. 2018, 27, 167–179.e167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [Green Version]
- Gavin, K.M.; Kohrt, W.M.; Klemm, D.J.; Melanson, E.L. Modulation of Energy Expenditure by Estrogens and Exercise in Women. Exerc. Sport Sci. Rev. 2018, 46, 232–239. [Google Scholar] [CrossRef]
- Dube, M.C.; Lemieux, S.; Piche, M.E.; Corneau, L.; Bergeron, J.; Riou, M.E.; Weisnagel, S.J. The contribution of visceral adiposity and mid-thigh fat-rich muscle to the metabolic profile in postmenopausal women. Obesity 2011, 19, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Leon, A.S. Attenuation of Adverse Effects of Aging on Skeletal Muscle by Regular Exercise and Nutritional Support. Am. J. Lifestyle Med. 2017, 11, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.C.; Laakkonen, E.K.; Lowe, D.A. Aging of the musculoskeletal system: How the loss of estrogen impacts muscle strength. Bone 2019, 123, 137–144. [Google Scholar] [CrossRef]
- Moreira, M.A.; Zunzunegui, M.V.; Vafaei, A.; da Camara, S.M.; Oliveira, T.S.; Maciel, A.C. Sarcopenic obesity and physical performance in middle aged women: A cross-sectional study in Northeast Brazil. BMC Public Health 2016, 16, 43. [Google Scholar] [CrossRef] [Green Version]
- Agostini, D.; Zeppa Donati, S.; Lucertini, F.; Annibalini, G.; Gervasi, M.; Ferri Marini, C.; Piccoli, G.; Stocchi, V.; Barbieri, E.; Sestili, P. Muscle and Bone Health in Postmenopausal Women: Role of Protein and Vitamin D Supplementation Combined with Exercise Training. Nutrients 2018, 10, 1103. [Google Scholar] [CrossRef] [Green Version]
- Kirk, B.; Al Saedi, A.; Duque, G. Osteosarcopenia: A case of geroscience. Aging Med. 2019, 2, 147–156. [Google Scholar] [CrossRef]
- Hamad, B.; Basaran, S.; Coskun Benlidayi, I. Osteosarcopenia among postmenopausal women and handgrip strength as a practical method for predicting the risk. Aging Clin. Exp. Res. 2020, 32, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.I.; Kim, H.; Ha, Y.C.; Kwon, H.B.; Koo, K.H. Osteosarcopenia in Patients with Hip Fracture Is Related with High Mortality. J. Korean Med Sci. 2018, 33, e27. [Google Scholar] [CrossRef]
- Greco, E.A.; Pietschmann, P.; Migliaccio, S. Osteoporosis and Sarcopenia Increase Frailty Syndrome in the Elderly. Front. Endocrinol. 2019, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic, V.; van der Mooren, M.J.; Stehouwer, C.D.; Netelenbos, J.C.; Kenemans, P. Postmenopausal hormone replacement, risk estimators for coronary artery disease and cardiovascular protection. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 1999, 13, 130–144. [Google Scholar] [CrossRef]
- Mumusoglu, S.; Yildiz, B.O. Metabolic Syndrome During Menopause. Curr. Vasc. Pharmacol. 2019, 17, 595–603. [Google Scholar] [CrossRef]
- Al-Safi, Z.A.; Polotsky, A.J. Obesity and menopause. Best Pract. Res. Clin. Obstet. Gynaecol. 2015, 29, 548–553. [Google Scholar] [CrossRef]
- Razmjou, S.; Abdulnour, J.; Bastard, J.P.; Fellahi, S.; Doucet, E.; Brochu, M.; Lavoie, J.M.; Rabasa-Lhoret, R.; Prud’homme, D. Body composition, cardiometabolic risk factors, physical activity, and inflammatory markers in premenopausal women after a 10-year follow-up: A MONET study. Menopause 2018, 25, 89–97. [Google Scholar] [CrossRef]
- Lovejoy, J.C.; Bray, G.A.; Bourgeois, M.O.; Macchiavelli, R.; Rood, J.C.; Greeson, C.; Partington, C. Exogenous androgens influence body composition and regional body fat distribution in obese postmenopausal women—A clinical research center study. J. Clin. Endocrinol. Metab. 1996, 81, 2198–2203. [Google Scholar] [CrossRef]
- Perry, A.; Wang, X.; Goldberg, R.; Ross, R.; Jackson, L. Androgenic sex steroids contribute to metabolic risk beyond intra-abdominal fat in overweight/obese black and white women. Obesity 2013, 21, 1618–1624. [Google Scholar] [CrossRef] [PubMed]
- Hammes, S.R.; Levin, E.R. Impact of estrogens in males and androgens in females. J. Clin. Investig. 2019, 129, 1818–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, A.P.; de Souza Santos, R.; Palmer, B.F.; Clegg, D.J. Determinants of body fat distribution in humans may provide insight about obesity-related health risks. J. Lipid Res. 2019, 60, 1710–1719. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.M. Lipid metabolism in women. Proc. Nutr. Soc. 2004, 63, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Sutton-Tyrrell, K.; Wildman, R.P.; Matthews, K.A.; Chae, C.; Lasley, B.L.; Brockwell, S.; Pasternak, R.C.; Lloyd-Jones, D.; Sowers, M.F.; Torrens, J.I.; et al. Sex-hormone-binding globulin and the free androgen index are related to cardiovascular risk factors in multiethnic premenopausal and perimenopausal women enrolled in the Study of Women Across the Nation (SWAN). Circulation 2005, 111, 1242–1249. [Google Scholar] [CrossRef] [Green Version]
- Torrens, J.I.; Sutton-Tyrrell, K.; Zhao, X.; Matthews, K.; Brockwell, S.; Sowers, M.; Santoro, N. Relative androgen excess during the menopausal transition predicts incident metabolic syndrome in midlife women: Study of Women’s Health Across the Nation. Menopause 2009, 16, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.H. Androgens, estrogens, and cardiovascular disease: Considerations for women with polycystic ovary syndrome. Fertil. Steril. 2019, 112, 478–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douchi, T.; Yoshimitsu, N.; Nagata, Y. Relationships among serum testosterone levels, body fat and muscle mass distribution in women with polycystic ovary syndrome. Endocr. J. 2001, 48, 685–689. [Google Scholar] [CrossRef] [Green Version]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of lipolysis in adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, A.; Mottillo, E.P. Adipocyte lipolysis: From molecular mechanisms of regulation to disease and therapeutics. Biochem. J. 2020, 477, 985–1008. [Google Scholar] [CrossRef]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Luong, Q.; Huang, J.; Lee, K.Y. Deciphering White Adipose Tissue Heterogeneity. Biology 2019, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Berman, D.M.; Nicklas, B.J.; Rogus, E.M.; Dennis, K.E.; Goldberg, A.P. Regional differences in adrenoceptor binding and fat cell lipolysis in obese, postmenopausal women. Metabolism 1998, 47, 467–473. [Google Scholar] [CrossRef]
- Stefanska, A.; Bergmann, K.; Sypniewska, G. Metabolic Syndrome and Menopause: Pathophysiology, Clinical and Diagnostic Significance. Adv. Clin. Chem. 2015, 72, 1–75. [Google Scholar] [CrossRef]
- Yamatani, H.; Takahashi, K.; Yoshida, T.; Soga, T.; Kurachi, H. Differences in the fatty acid metabolism of visceral adipose tissue in postmenopausal women. Menopause 2014, 21, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Goettems-Fiorin, P.B.; Costa-Beber, L.C.; Dos Santos, J.B.; Friske, P.T.; Sulzbacher, L.M.; Frizzo, M.N.; Ludwig, M.S.; Rhoden, C.R.; Heck, T.G. Ovariectomy predisposes female rats to fine particulate matter exposure’s effects by altering metabolic, oxidative, pro-inflammatory, and heat-shock protein levels. Environ. Sci. Pollut. Res. Int. 2019, 26, 20581–20594. [Google Scholar] [CrossRef]
- de Souza, C.F.; Stopa, L.R.S.; Santos, G.F.; Takasumi, L.C.N.; Martins, A.B.; Garnica-Siqueira, M.C.; Ferreira, R.N.; de Andrade, F.G.; Leite, C.M.; Zaia, D.A.M.; et al. Estradiol protects against ovariectomy-induced susceptibility to the anabolic effects of glucocorticoids in rats. Life Sci. 2019, 218, 185–196. [Google Scholar] [CrossRef]
- Moise, A.R.; Kuksa, V.; Imanishi, Y.; Palczewski, K. Identification of all-trans-retinol:all-trans-13,14-dihydroretinol saturase. J. Biol. Chem. 2004, 279, 50230–50242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soucy, G.; Boivin, G.; Labrie, F.; Rivest, S. Estradiol is required for a proper immune response to bacterial and viral pathogens in the female brain. J. Immunol. 2005, 174, 6391–6398. [Google Scholar] [CrossRef] [Green Version]
- Costa-Beber, L.C.; Goettems-Fiorin, P.B.; Dos Santos, J.B.; Friske, P.T.; Frizzo, M.N.; Heck, T.G.; Hirsch, G.E.; Ludwig, M.S. Ovariectomy enhances female rats’ susceptibility to metabolic, oxidative, and heat shock response effects induced by a high-fat diet and fine particulate matter. Exp. Gerontol. 2021, 145, 111215. [Google Scholar] [CrossRef]
- Kastaniotis, A.J.; Autio, K.J.; Kerätär, J.M.; Monteuuis, G.; Mäkelä, A.M.; Nair, R.R.; Pietikäinen, L.P.; Shvetsova, A.; Chen, Z.; Hiltunen, J.K. Mitochondrial fatty acid synthesis, fatty acids and mitochondrial physiology. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2017, 1862, 39–48. [Google Scholar] [CrossRef]
- Kushwaha, P.; Wolfgang, M.J.; Riddle, R.C. Fatty acid metabolism by the osteoblast. Bone 2018, 115, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; Carneiro-Freire, N.; Seco-Filgueira, M.; Fernández-Fernández, C.; Mouriño-Bayolo, D. Mitochondrial β-oxidation of saturated fatty acids in humans. Mitochondrion 2019, 46, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Karwi, Q.G.; Uddin, G.M.; Ho, K.L.; Lopaschuk, G.D. Loss of Metabolic Flexibility in the Failing Heart. Front. Cardiovasc. Med. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holloway, G.P.; Bonen, A.; Spriet, L.L. Regulation of skeletal muscle mitochondrial fatty acid metabolism in lean and obese individuals. Am. J. Clin. Nutr. 2009, 89, 455s–462s. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
LDL cholesterol is used for E2 synthesis. cAMP: cyclic adenosine monophosphate, E2: estradiol, FSH: follicle-stimulating hormone, FSHR: follicle-stimulating hormone receptor, LDL-C: low-density lipoprotein cholesterol, LH: luteinizing hormone, LHR: luteinizing hormone receptor, PKA, protein kinase A.
Figure 1.
LDL cholesterol is used for E2 synthesis. cAMP: cyclic adenosine monophosphate, E2: estradiol, FSH: follicle-stimulating hormone, FSHR: follicle-stimulating hormone receptor, LDL-C: low-density lipoprotein cholesterol, LH: luteinizing hormone, LHR: luteinizing hormone receptor, PKA, protein kinase A.

Figure 2.
Energy balance between energy in and energy out.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Body compositions of Reference Men and Women [14].
Table 1.
Body compositions of Reference Men and Women [14].
| Men | Women | |
|---|---|---|
| Age, years | 20–24 | 20–24 |
| Height, in | 68.5 | 64.5 |
| Weight, lb | 154 | 125 |
| Total fat, lb (% body weight) | 23.1 (15.0%) | 33.8 (27.0%) |
| Storage fat, lb (% body weight) | 18.5 (12.0%) | 18.8 (15.0%) |
| Essential fat, lb (% body weight) | 4.6 (3.0%) | 15.0 (12.0%) |
| Muscle, lb (% body weight) | 69 (44.8%) | 45 (36.0%) |
| Bone, lb (% body weight) | 21 (14.9%) | 15 (12.0%) |
| Remainder, lb (% body weight) | 38.9 (25.3%) | 31.2 (25.3%) |
| Average body density | 1.070 g/mL | 1.040 g/mL |
Table 2.
Estimated metabolic rates of tissues and percentage contribution to total metabolism [34].
Table 2.
Estimated metabolic rates of tissues and percentage contribution to total metabolism [34].
| REE | MEN | WOMEN | |
|---|---|---|---|
| kcal/kg/Day | % Total REE | % Total REE | |
| Liver | 200 | 17 | 18 |
| Brain | 240 | 19 | 21 |
| Heart | 440 | 9 | 8 |
| Kidneys | 440 | 8 | 8 |
| Skeletal muscle a | 13 | 24 | 20 |
| Adipose tissue | 4.5 | 4 | 7 |
| Other b | 12 | 19 | 18 |
| Total | 100 | 100 |
REE, resting energy expenditure. a Resting and nonexercised recovery rate; b skeleton, blood, skin, gastrointestinal tract, lungs, spleen, and other organs.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ko, S.-H.; Jung, Y. Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women. Nutrients 2021, 13, 4556. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13124556
AMA Style
Ko S-H, Jung Y. Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women. Nutrients. 2021; 13(12):4556. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13124556
Chicago/Turabian StyleKo, Seong-Hee, and YunJae Jung. 2021. "Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women" Nutrients 13, no. 12: 4556. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13124556
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.