
Lesser Investigated Natural Ingredients for the Management of Obesity
by
 , ,
, ,
Muhammed Majeed
1,2, 
Shaheen Majeed
2 ,
,
Kalyanam Nagabhushanam
2,
Muthuraman Gnanamani
1 and
Lakshmi Mundkur
1,* 1
Sami-Sabinsa Group Limited, Peenya Industrial Area, Bangalore 560058, Karnataka, India
2
Sabinsa Corporation, 20 Lake Drive, East Windsor, NJ 08520, USA
*
Author to whom correspondence should be addressed.
Nutrients 2021, 13(2), 510; https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020510
Submission received: 31 December 2020
/
Revised: 30 January 2021
/
Accepted: 31 January 2021
/
Published: 4 February 2021
(This article belongs to the Special Issue Nutrients, Metabolism and Obesity Prevention)
Abstract
:Obesity, an epidemiological disorder, is related to various complications in both the developed and developing world. It epitomizes a crucial risk factor for health, decreasing productivity and life expectancy while increasing health care costs worldwide. Conventional therapies with synthetic drugs or bariatric surgery, associated with numerous side effects, recurrence, and surgical complexity, have been restricted in their use. Lifestyle changes and dietary restrictions are the proven methods for successful weight loss, although maintaining a strict lifestyle is a challenge. Multiple natural products have been explored for weight management with varied efficacy. The current review explores less explored natural herbs, their active constituents, and their mechanisms of action against obesity.
Keywords:
anti-obesity; phytochemicals; energy balance; gut microbiota; leptin resistance; satiety; mechanism1. Introduction
Obesity is a challenging condition of excess body fat, caused by an imbalance in energy consumption and expenditure [1]. It is the result of a complex interaction between environment, diet, genetics, lifestyle, endocrine disorders, medication, and psychological factors [2]. As per the World Health Organization (WHO), individuals with body mass index (BMI) of more than 30.0 kg/m2 are considered obese, and those with BMI between 25.0 and 30.0 kg/m2 are categorized as overweight [3].
Obesity is associated with comorbidities like diabetes, hypertension, hyperlipidemia, cancer, and sleep apnea, and is considered as an independent risk factor for cardiovascular diseases (CVD) [4]. An increase by one unit BMI causes a 4% rise in ischemic risk and a 6% rise in hemorrhagic strokes [5]. Higher systolic and diastolic blood pressure of 3.0 mm and 2.3 mm Hg, respectively, in individuals with 10 kg excess body weight can induce a 12% increased risk of CVD and 24% increased risk of stroke [6]. Besides, obesity is also associated with increased risk for several cancers including colon, endometrial, kidney, esophageal, liver, pancreatic, breast, Hodgkin’s lymphoma, and myeloma [7,8]. According to the WHO, incidence of obesity has tripled since 1975. In a survey conducted in 2016, 1.9 billion adults and 340 million children and adolescents were reported to be obese or overweight [9]. The rapid increase in obesity prevalence and its associated devastating health effects and comorbidities highlight the immediate need for early recognition, control, and treatment of this problem. Although diet control, exercise and lifestyle changes are the fundamental therapeutic options, few drugs have been approved for pharmacotherapy. Several natural products have been widely studied and numerous review articles reference the use of herbs for weight management [10,11,12]. In the present review, we focus on a few of these less explored natural extracts, their phytochemicals, and their mechanism of action in controlling obesity.
2. Pharmaceutical Drugs for Obesity
Several anti-obesity drugs have been evaluated since the beginning of the 20th century. The earliest drugs were the thyroid hormones due to their thermogenic effect, followed by chemicals such as dinitrophenol, mitochondrial uncouplers, amphetamines, and serotonergics as appetite suppressors, which were subsequently withdrawn due to safety concerns [13]. Following this, poly diet pills using the combination of amphetamines and thyroid hormones along with several other ingredients were distributed as rainbow pills for diet control. Though they were highly popular, fatalities associated with their indiscriminate use led to their withdrawal mandated by the US FDA [13]. A list of drugs and their present status is presented in Table 1. The search for novel drugs to reduce appetite and improve glucose metabolism to control obesity and diabetes is being pursued with renewed vigor and attention as the economic burden associated with obesity exceeds US$200 billion in the United States alone and is increasing steadily [14].
3. Pathophysiology of Obesity
In the last few decades, significant advances have been made in understanding the pathophysiological mechanisms involved in obesity. The neuroendocrinal feedback associated with pathological overeating coupled with physical inactivity seem to be the major factors governing obesity. Apart from this, genetic predisposition, hormonal imbalance, and gut microbial dysbiosis also contribute to accumulation of fat stores [18]. Figure 1 describes the pathophysiological parameters causing obesity.
3.1. Energy Intake vs. Expenditure
Obesity can be viewed as an imbalance in energy intake versus energy expenditure. Three components involved in energy expenditure are (i) resting metabolic rate, which is the energy necessary to fuel the body at rest, (ii) activity-related energy expenditure and (iii) diet-induced thermogenesis, which is the energy spent in absorbing and metabolizing food consumed [19]. When an individual ingests more energy than their expenditure, a positive energy balance develops, and this excess energy is converted into triglycerides and stored. When energy intake exceeds energy expenditure by more than 20 kcal/day, 1 kg of fat per year gets accumulated [20]. Thus, a proper balance of energy intake and expenditure is necessary to manage obesity. A complex physiological control system involving signals from the periphery about the status of stored energy, and those that affect energy intake and expenditure, are responsible for maintaining the energy balance [21].
Energy Expenditure and Thermogenesis
The adipose tissue is generally considered as a passive depot for the storage of excess calories. Recent studies have shown that it is an active endocrine organ, which takes part in energy balance by releasing free fatty acids, proinflammatory cytokines and adipokines such as leptin and adiponectin, regulating food intake and insulin sensitivity [22]. Adipose tissues are heterogeneous and are classified into white adipose tissue (WAT), which are storage organs, and brown adipose tissue (BAT), which burns energy for thermogenesis. Intermediary beige adipocytes, arising from WAT, have also been described in mammals [23]. Exposure to cold, adrenergic stimulation, and long-term treatment with peroxisome proliferator-activated receptor (PPAR)γ agonists are some of the external cues that induce these beige adipocytes [24]. The brown and the beige adipocytes contain numerous mitochondria and express the uncoupling protein 1 (UCP-1), which regulates energy expenditure, reduces adiposity, and protects experimental animals from diet-induced obesity [25]. Besides UCP-1, brown adipocytes express type 2 iodothyronine deiodinase (DIO2), the transcription coregulators PR domain containing 16, (PRDM16) and Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), and Cell death activator (CIDE-A), which regulates UCP1 transcription in BAT [26]. With the understanding that BAT can help in energy dissipation, pharmacological interventions targeting browning of WAT are being actively investigated to induce energy balance. Few natural ingredients have been reported to convert WAT to BAT, but Forskolin, Capsaicin, Resveratrol, and Berberine are examples of molecules that could induce browning of WAT [27,28].
3.2. Hormonal Imbalance
Neurobiological mechanisms are reported to contribute to eating in the absence of energy demand and hunger. The cortico-limbic system, hypothalamus and hindbrain are three heavily interconnected brain regions that are involved in controlling eating behavior [29]. These are affected by various visual food stimuli under conditions of fasting, weight loss, overfeeding, exercise, hormone infusion, leanness, obesity, and voluntary cognitive control [30]. The important neurostimulators such as serotonin and dopamine play a significant role in food intake. Increased serotonergic signaling is associated with decreased food intake, whereas its decrease induces hyperphagia and weight gain [31]. Likewise, lesser dopaminergic signaling promotes overconsumption of food beyond homeostatic needs [32].
The gut hormones play the most critical role in obesity. The cells of the gastrointestinal tract sense ingested food and release the gut hormones that regulate energy and glucose homeostasis through autocrine, paracrine, and endocrine pathways. Glucagon-like peptide 1 (GLP-1), peptide (PYY) and oxyntomodulin signal the availability of nutrients to the brain and suppress appetite [33]. Additionally, leptin secreted by the adipose tissue acts as a signal for energy availability and promotes satiety. In the absence of food, ghrelin is secreted, which sensitizes the brain to the intake of food [34].
Leptin Resistance
Leptin is an adipokine that regulates food intake, energy expenditure, immune function, and numerous other physiological activities [35]. Circulating leptin concentrations directly reflect the adipose tissue energy stores and it generally enables energy expenditure while reducing food craving [36]. Leptin binds to its receptor in the brain and mediates its action through the neuroendocrine axes. It also attenuates the hyperglycemia caused by insulin deficiency [37]. Since leptin acts as a messenger for peripheral energy stores, increasing its circulating levels was thought to be a potential treatment for obesity [38]. During the progression of obesity, leptin signaling is affected, leading to leptin resistance. In these cases, the leptin levels are high in serum, but it is unable to bind to its receptor and mediate the physiological activity [39]. Obesity is also associated with leptin resistance, which affects leptin signaling and its downstream physiological effects. Obese patients develop leptin resistance despite high circulating levels of the adipokine, rendering the leptin therapy ineffective. Alleviating leptin resistance is an exciting research area as potential anti-obesity therapy as no drugs are known for this function (Figure 2).
3.3. Gut Microbiota
Various bacteria, viruses, fungi, and protozoa colonize the gastrointestinal tract, which is interactively involved in immune, metabolic, and neurological health [40,41]. The gut microbes play a major role in metabolism by fermenting the non-digestible dietary fibers to short-chain fatty acids (SCFA) such as butyrate, propionate, and acetate [42]. These SCFAs are involved in cholesterol metabolism and lipogenesis and are reported to play a central role in appetite regulation [43]. In the last two decades, several studies have shown that probiotic supplements can reduce body weight and improve glucose metabolism in rodents by changing the composition of gut microbiota [44]. Overweight and obese people show a dysbiosis characterized by lower microbial diversity associated with impaired glucose homeostasis and low-grade inflammation [45,46]. A meta-analysis of human clinical trials concluded that obesity was associated with higher counts of Firmicutes, Fusobacteria, Proteobacteria, and Lactobacillus reuteri, and lower counts of Bacteroidetes, Akkermansia muciniphila, Faecalibacterium prausnitzii, Lactobacillus plantarum, and Lactobacillus paracasei. An increase in the Firmicutes/Bacteroidetes ratio was observed in association with obesity [47], while a reduction in Firmicutes’ proportion with a rise in Bacteroidetes was associated with weight loss [48]. Further, the gut microbiome composition was found to be directly altered by the diet. In animal models, a high-fat diet favored Firmicutes and lowered the Bacteroidetes [49]. The metabolites derived from the fermentation of food by microbiome play a vital role in regulating host metabolism. The gut bacteria convert bile acid in the intestine to deoxycholic acid and lithocholic acid, which stimulate the secretion of incretin hormone GLP-1 and insulin, thereby promoting energy expenditure [50]. Dietary choline metabolism is also linked to microbiome composition. Conversion of choline into trimethylamine-N-oxide (TMAO) by microbiome has been associated with atherosclerosis and metabolic disorders [51]. The conversion of choline to the intermediate trimethylamine is mediated by several intestinal-resident bacteria. The SCFAs produced by gut bacteria are involved in insulin signaling associated with fat accumulation, modulate the secretion of GLP-1 and suppress the inflammatory immune response in the gut [45,52,53]. Inflammation and gut permeability are other markers associated with adiposity [54,55]. These two factors are interlinked as an increased permeability allows bacterial metabolites to leak into the circulation causing low-grade inflammation, a characteristic feature of obesity and insulin resistance [56]. The proinflammatory cytokines, in turn, can cause intestinal barrier disruption [57].
3.4. Genetic Predisposition
Mutation in the leptin-melanocortin pathway [58,59], polygenic obesity [60] and epigenetic disorders like Prader-Willi and Temple syndrome [61] play major roles in the pathogenesis of obesity. Some single gene mutations which disrupt the regulatory system of appetite and weight are described in Table 2. The genetic predisposition to obesity has been reviewed by several authors and is beyond the scope of this manuscript.
4. Adipogenesis and Growth of Adipose Tissue
A variety of cell types, including endothelial cells, fibroblasts, pericytes, preadipocytes, macrophages, and other immune cells are present in the adipose tissue. The expansion of adipose tissues occurs by both increase in their numbers (hyperplasia) and size (hypertrophy). The adipocytes differentiate from the mesenchymal stem cells (MSC), by a multistep process, involving several genes, signaling and transcriptional factors. Three well-defined phases are involved in the process of adipogenesis: the commitment of MSC to adipocyte lineage, mitotic clonal expansion, followed by terminal differentiation. The final stages of differentiation involve the expression of genes and transcriptional factors such as PPARγ, CCAAT/enhancer-binding proteins (C/EBPs), increase in lipogenesis, and the induction of lipogenic genes such as acetyl CoA carboxylase (ACC), fatty acid synthase (FAS) and adipocyte fatty acid binding protein (aP2) [69]. The canonical Wnt/catenin, Hedgehog and transforming growth factor beta (TGF-β) 1 and 2, Sirtuin (Sirt) 1, microRNA (MiR)-27a and MiR-93, signaling pathways inhibit PPARγ, and C/EBPα expression. The expression of these genes is promoted by the glucocorticoid, cAMP, and bone morphogenic proteins (BMPs) signaling. The microRNA signals by MiR-210 and MiR-146, bromodomain-containing protein 4 (BRD4), Sirt 7 mediated signals also promote PPARγ and C/EBPα expression [70]. The white and brown adipose cells have a distinct origin and morphological characters. The WAT is differentiated from the mesenchymal precursor cells while the origin of BAT is the Myf5 expressing precursors [71]. Further, the brown and white adipocytes store and metabolize lipids in different manners. White adipocytes accumulate nutrient-derived triglycerides and release them by lipolysis during periods of fasting, whereas brown adipocytes oxidize their lipid stores in an elegant heat-producing pathway mediated essentially by UCP1. Recent studies have shown physiological plasticity in WAT. Cold exposure, or treatment with β-adrenergic receptor (β3) agonists that enhance lipolysis, induces a subset of UCP1 positive in WAT, which share additional characteristics with brown adipocytes [72]. Unlike classical BAT these brown-like cells in WAT are not derived from Myf5 positive precursors.
5. Herbal and Dietary Supplementation for Weight Management
Traditional and complementary medicine has become an indispensable part of the healthcare system all over the world. In recent decades natural products have come into focus, for their potent pharmacological activities with minimal adverse effects. A variety of natural plants and other natural dietary products have been reported to have anti-obesity activity. There are reportedly more than 54 plant families whose species have shown an anti-obesity potential, the major phytoconstituents being the flavonoids and polyphenols followed by terpenoids, alkaloids and organic acids. These phytochemicals have been reported to regulate fatty acid oxidation, reduce plasma lipid levels, and inhibit pancreatic lipase activity. Few of these phytochemicals have been extensively studied for their molecular mechanism of action [12]. They have been shown to reduce the oxidative stress and systemic inflammation induced by obesity and reduce adipogenesis by inhibiting cell cycle and AMPKα signaling [73,74]. In the next sections we describe the studies related to the mechanism of action of lesser explored extracts and their phytochemicals in relation to the pathophysiology of obesity.
5.1. Coleus forskohlii
Forskolin is a diterpenoid from the root of Coleus forskohlii, a native Indian plant of the Lamiaceae family. It stimulates adenylate cyclase enzyme, producing cellular cyclic adenosine monophosphate (cAMP), the secondary messenger having a broad range of activities [75]. cAMP induces biochemical events that trigger the metabolic processes and diet-induced thermogenesis, increase lean body mass, and stimulate the loss of body fat [76]. The leaves, roots and structure of Forskolin are shown in Figure 3.
Several clinical and pre-clinical studies report the effect of forskolin in promoting lean body mass and decreasing body fat. In one of the earlier clinical studies, fourteen overweight volunteers, were administered forskolin at a dose of 125 mg twice a day for 12 weeks. The total body weight and body fat decreased significantly, and BMI improved at the end of the study [77]. In an 8 week open labelled study, 6 overweight, women receiving 500 mg of the C. forskohlii extract, equivalent to 50 mg forskolin per day, showed significant reduction of body weight and fat content and an increase in lean body mass [78]. In 41 obese patients administered with forskolin along with 250 mg of C. forskohlii extract for 12 weeks, body weight differences were not significant, but changes in hip and waist circumference were significant, suggesting a decrease in fat mass and an increase in bone mass [79]. Similar results were observed in other randomized trials in obese men and women [80,81,82].
Mechanism of action: Forskolin increases the production of Hormone-sensitive lipase (HSL) by the action of cAMP. HSL releases stored triglycerides for metabolic consumption, thus reducing fat storage [83]. The increased cAMP activates the downstream protein kinase A. This cAMP/PKA dependent pathway leads to the overexpression of UCP1, the protein involved in adipocyte browning. PKA activates the p38 mitogen-activated protein kinase (MAPK), which stimulates the expression of the PPARγ coactivator-1α (PGC-1α) and the activating transcription factor 2 increasing UCP1 expression. Alternately, cAMP response element binding protein directly binds on UCP1 promoter in a p38 MAPK-independent manner to induce UCP1 expression [84]. Forskolin also activates AMPK by phosphorylating the tyrosine-172 residue, required for lipolysis activity [85].
5.2. Garcinia cambogia and Garcinia indica
Garcinia, commonly known as kokum, belongs to the Clusiaceae family and is traditionally used as a flavoring agent (Figure 3). For centuries it has been used for culinary purposes in place of tamarind or lemon and as a pharmaceutical and nutraceutical in many regions of South India [86]. The major metabolites in G. indica are hydroxy citric acid (HCA), malic acid, citric acid, and tartaric acid while benzophenones, bioflavonoids, xanthones and anthocyanin pigments are the secondary metabolites in the fruit rind [87,88]. Garcinol, Isogarcinol, and Isoxanthochymol are some of the major derivatives of poly-isoprenylated-benzophenones isolated from fruits, dry rinds and leaves [89,90]. As an anti-obesity agent, HCA is reported to reduce food intake, increase energy expenditure, suppress fatty acid synthesis, and enhance glycogen synthesis in the liver [91]. The habitat of the plant and the structures of its phytochemicals are presented in Figure 4.
Garcinol is a yellow crystalline poly-isoprenylated benzophenone derivative extracted from fruit rind of Garcinia indica [92]. The extract of G. indica standardized for 20% w/w Garcinol inhibited the differentiation of 3T3-L1 preadipocytes in vitro by downregulating the adipogenic genes and by increasing the transcripts associated with energy metabolism and adipocyte browning [93]. In the animal model, Garcinol induced a dose-dependent reduction of total body weight and visceral adipose tissue in high-fat diet- (HFD)-induced obese mice. Garcinol mediated the anti-obesity activity by reducing endoplasmic stress in preadipocytes and adipose tissues. The results suggested a novel molecular mechanism of Garcinol’s action, acting on the AMPK-ER stress axis [93]. Lee et al. reported a similar result, wherein Garcinol administration to HFD-fed mice showed a reduction in glutamate pyruvate transaminase, total cholesterol, and triacylglycerol. In this study, Garcinol reversed gut dysbiosis by decreasing the Firmicutes-to-Bacteroidetes ratio and controlled inflammation by increasing intestinal commensal bacteria, Akkermansia muciniphila, which has been strongly and negatively correlated with age and HFD feeding in mice [94]. In obese mice, a reduction in the A. muciniphila could be correlated with expression of inflammation markers, lipid metabolism, circulating glucose, insulin, triglycerides, and leptin levels [95]. In another study, Garcinia cambogia extract along with probiotics was observed to change the gut microbial community compared to the probiotic alone. The combination increased the Bifidobacteria and decreased Clostridium aminophilum, and other strains associated with diet induced obesity [96].
The organic acids, i.e., hydroxy citric acid (HCA) present in Garcinia, are also identified as potential weight loss supplements. Since HCA and G. cambogia are well-covered in numerous articles, they are not included in the present review.
Mechanism of action: AMP-activated protein kinase (AMPK) is a major regulator of energy metabolism Once activated, AMPK shifts the cellular metabolism from anabolic pathways to catabolic pathways. Activation of AMPK in adipocytes leads to a decreased fatty acid uptake, decreased triglyceride synthesis, and increased fatty acid oxidation. Garcinol activates the AMPK, and reduces cellular ER stress thereby improving metabolism. Garcinol was also reported to activate PGC-1α, PRDM16, and BMP-7, all involved in increasing UCP1 expression and in browning of white adipocytes. Furthermore, Garcinol shows profound influence on the gut microbiome, regulating the gut dysbiosis, and inflammation. The increase in A. mucinifila could be directly correlated to the anti-obesity activity of the molecule, remodulating the gut microbiome to reduce obesity.
5.3. Cyperus rotundus
Cyperus rotundus is a perennial herb native to India, used to treat various ailments such as diarrhea, diabetes, pyrosis, inflammation, malaria, bowel disorder, cancer, hypertension, and allergy (Figure 4) [97]. The major organic chemicals isolated from various parts of the Cyperus species include quinonoid pigments, sesquiterpenoids, flavonoids and stilbene derivatives [98].
Several monoterpenoids, amino acids and fatty acids are also reported in the plant. The ethyl acetate extract of dried, pulverized rhizomes of C. rotundus was reported to contain Scirpusin A, Scirpusin B & Piceatannol as major compounds [87]. The chloroform/methyl alcohol extraction of C. rotundus yielded novel enantiomeric and meso-stilbene trimers, such as Cyperusphenol A as well as other stilbenoids (Cyperusphenol C & D, Scirpusins A & B, Piceid and Luteolin) [99,100]. The plant rhizomes and the structures of its phytochemicals are presented in Figure 5.
The aqueous tuber extract of C. rotundus at a dose of 100, 200 and 300 mg/kg BW in rats, along with HFD for 40 days, showed a reduction in body weight gain, organ weight, and weight of fat pads. Serum triglycerides, TC, LDL, VLDL, and glucose were reduced while HDL cholesterol increased. The study reported normalization of liver enzymes and the reduction of oxidative stress in HFD obese rats [101]. Similar results were reported with 500 mg/kg BW of ethanolic extract of c. rotundus rhizome [102]. The hypolipidemic activity was reported at much lower doses (70, 140, and 280 mg/kg BW) [103]. The ethyl acetate extract of C. rotundus rhizome, having 5% total stilbenes with Scirpusin A, Scirpusin B, and Piceatannol as major compounds, showed a dose-dependent reduction of adipogenesis in vitro in 3T3-L1 preadipocyte cells. A reduction in body weight, leptin, corticosteroid concentration, and normalized lipid profile was observed in the HFD induced obese mice (Majeed et al., unpublished work). In a pilot clinical study C. rotundus extract showed bodyweight reduction, a significant decrease in waist circumference, and BMI with no adverse events [104]. Piceatannol and its dimer Scirpusin B are also found in passion fruit (Passiflora edulis) seeds in larger amounts [105,106]. Piceatannol has been shown to inhibit adipogenesis of preadipocytes 3T3-L1, especially in the first 24 h of adipogenesis by inhibiting the cell cycle progression of preadipocytes. Piceatannol suppressed mitotic clonal expansion by reducing the activation of the insulin-signaling pathways [107]. At a dose of 0.1% and 0.25%, piceatannol significantly lowered body weight, serum cholesterol, and LDL/HDL ratio and had an impact on gut microbiota by increasing the amount of Lactobacillus in high-fat-diet-treated mice [108]. Piceatannol was also reported to induce slight changes in the abundance of Lactobacillus, Clostridium, and Bacteroides in the gut, associated with a decrease in circulating non-esterified fatty acids, LDL-cholesterol and lactate in Zucker rats [109] The molecule also induced mild reno-protective effect in obese Zucker rats by reducing renal fibrosis and lipid peroxidation [110]. In a coculture of adipocyte and macrophage system, piceatannol significantly reduced the release of TNF-α and monocyte chemoattractant protein-1 (MCP-1) [111]. Stilbene, flavonoid and total (poly)phenol intake was associated with higher gut microbiome diversity, an increase in butyrate producing organisms and 20–23% lower prevalence of obesity [112].
Mechanism of action: Piceatannol inhibits adipogenesis by inhibiting mitotic clonal expansion and lowering the protein levels of adipogenic transcription factors such as PPARγ and C/EBPα in 3T3-L1 cells in vitro. They also regulate fatty acid synthase activity to regulate the triglyceride biosynthesis. Piceatannol also regulates the gut microbiome and reduces obesity mediated inflammation.
5.4. Curcuma longa
A rhizomatous perennial herb of the ginger family (Figure 5A), it is traditionally used for thousands of years in Ayurveda, Siddha medicine, traditional Chinese medicine, and Unani as a remedy to cure various illnesses related to inflammation, infectious diseases, gastric, hepatic and blood disorders [113,114]. The major ingredients of turmeric powder include 60–70% carbohydrates, 6–13% water, 6–8% protein, 5–10% fat, 3–7% dietary minerals, 3–7% essential oils, 2–7% dietary fiber and 1–6% curcuminoids. It has three major curcuminoids that include, Curcumin 50–60%, Demethoxy-curcumin 20–27%, and Bisdemethoxycurcumin (BDMC) 20–25% [115]. The habitat of the plant and the structures of its phytochemicals are presented in Figure 6.
BDMC is a beta-diketone existing nearly exclusively in its enolic form. It is reported to show a promising anti-tumor property by inducing caspase-dependent and independent apoptosis via the Smad and Akt signaling pathway [116,117]. The anti-obesity effect of BDMC and its mechanism of action was explored in 3T3-L1 mouse adipocytes in vitro and in the diet-induced obesity model in C57BL/6J mice in vivo. BDMC showed potent anti-adipogenic activity at 25 µM concentration. It was reported to suppress adipogenesis by attenuating mitotic clonal expansion by downregulating cyclin A, B, p21 and the mitogen-activated protein kinase (MAPK) signal. Additionally, the adipogenic transcription factors PPAR γ and C/EBP α were also downregulated by BDMC treatment. Similarly, the HFD induced mice treatment with 0.5% dietary BDMC (w/w) showed significantly lower adipose tissue mass [118].
Curcuma species contain several active phytochemicals with diverse biological activities (Figure 5C). Calebin A, a novel molecule with similar structural features to curcumin but devoid of its iconic 1,3-diketonic feature, was isolated from C. longa. It was shown to have anticancer and neuroprotective effects [119,120]. Calebin A belongs to a family of ferulate esters, rightfully called calebenoids, occurring naturally in Curcuma longa [121]. These calebenoids, especially Calebin A, seem more stable than Curcumin in a physiological medium with higher chemical stability in acidic and basic media, but sharing several physiological properties with Curcumin in contrast to the noted instability of Curcumin at higher pH values. The Calebenoids seem to arise from a biological Bayer-Villiger type of transformation of curcuminoids [122].
Calebin-A significantly inhibited the differentiation of preadipocytes and lipid accumulation in 3T3-L1 cells in vitro. It suppressed the expression of C/EBPα, C/EBPβ, and PPARγ proteins, which are the master regulators of adipogenesis. Calebin-A effectively decreased weight gain and relative perigonadal, retroperitoneal and mesenteric fat weight in a murine model of diet-induced adipogenesis. Further, Calebin-A reduced hepatic steatosis and restored the liver enzymes to normal levels. Calebin A was reported to mediate its anti-obesity effect through the activation of AMP-activated protein kinase signaling in both in vitro adipocytes and liver tissues [123].
Mechanism of Action: BDMC regulates the mitotic expansion of preadipocytes and regulates adipogenesis by downregulating PPAR γ and C/EBP α, while Calebin A mediates its anti-obesity effect by activation of AMPK signaling in both in vitro adipocytes and liver tissues. Calebin A also reduces leptin levels and increases adiponectin, and thus may also act through the adipokine regulation pathway to alleviate leptin resistance and increase metabolism.
5.5. Oroxylum indicum
Also known as “Sonapatha or Indian trumpet tree,” Oroxylum indicum belongs to the Bignoniaceae family (Figure 6). It is commonly found in tropical countries such as India, Japan, China, Sri Lanka, and Malaysia. Various plant segments are reported to be used in traditional medicine for cancer, diarrhea, fever, ulcer, jaundice, and arthritis [124]. The plant contains several secondary metabolites like polyphenols, flavonoids, tannins, terpenoids and alkaloids. It contains an essential oil that gives a specific aroma to the plant [125]. Various active ingredients have been isolated from O. indicum. The stem bark contains Oroxylin-A, Chrysin, Baicalein, Scutellarin-7-rutinoside, Tannic acid, Sitosterol, galactose, Baicalin, Biochanin-A and Ellagic acid [126,127]. The habitat of the plant and the structure of the active constituent are given in Figure 7.
Among the several flavonoids, Oroxylin A has been explored for its wide range of therapeutic applications ranging from anticancer, anti-obesity, antioxidant, anti-inflammatory, cardioprotective and neuroprotective roles [128].
The O. indicum extract composed of flavonoids, alkaloids, steroids, glycosides, and tannins exhibited a dose-dependent reduction of lipid accumulation in 3T3-L1 cell lines and showed an inhibitory effect on lipase activity [129]. Similarly, the ethyl acetate extract of O. indicum bark, containing three bioactive ingredients Oroxylin A, Chrysin and Baicalein, showed around 75% inhibition of lipid accumulation in in vitro cells at 50 µg/mL. The transcription factors PPAR γ and C/EBPα were downregulated by Oroxylin A and Chrysin [130]. The mRNA expression of lipogenic genes controlling adipogenesis, including SREBP-1c, GLUT4, FAS and leptin, was inhibited by O. indicum extract containing Quercetin, Apigenin, Kaempferol, Baicalein and Biochanin A [131]. Oroxylin A was also found to be more potent than non-methoxylated flavonoids Morin, Naringenin and Kaempferol in the same study.
Mechanism of Action: O. indicum extract containing Quercetin, Apigenin and Kaempferol prevents lipid uptake in the intestine by inhibiting pancreatic lipase and reduces the activity of adipogenic and lipogenic genes controlling adipogenesis, including PPARγ2, SREBP-1c, GLUT4, FAS and leptin, while Oroxylin A and Chrysin regulate adipogenesis by downregulating PPAR γ and C/EBP α.
5.6. Pterocarpus marsupium
Pterocarpus marsupium is a large deciduous tree, the heartwood extracts of which are extensively used as an antibiotic and hypoglycemic to control blood sugar (Figure 7). The Kino gum obtained from the bark is reported to have astringent, anti-diarrheal, and anti-hemorrhagic properties and the leaves are used to treat skin-related diseases. The tree is found in central and peninsular India. It can survive excessive temperatures in summer and prefers fertile, deep clayey loam soil with good drainage for growth [132]. Pterostilbene is the most actively studied ingredient of the extract [133]. Figure 8 shows the habitat of the plant and the structure of pterostilbene.
Pterostilbene administered at 15 and 30 mg/kg BW for six weeks in obesogenic rats reduced the adipose tissue mass by 15.1% and 22.9%, respectively, which was associated with a reduction in fatty acid synthase. The compound decreased the activity of acetyl-CoA carboxylases involved in regulating the biosynthesis and metabolism of fatty acids and activated the AMPK gene, thus, inducing protection against diet-induced obesity [134]. Pterostilbene was reported to inhibit adipocyte differentiation in 3T3-F442A preadipocytes and eliminate the lipogenic effect of insulin without inhibiting its antilipolytic action and glucose uptake, revealing a unique interaction with adipocytes [135].
Pterostilbene downregulated the expression of adipocyte conditioned medium (aCM)-induced fatty acid-binding protein 5 (FABP5) and pro-metastatic factors such as VEGF, MMP2, MMP9, and TNF-α by inhibiting NF-κB, β-catenin and PPAR-γ. It was also reported to suppress PI3K, Akt, p38 MAPK, ERK and JNK1/2 signaling pathways and to alleviate the adiposity-induced metastasis in obesity-related colorectal cancer cells [136].
The effect of Pterostilbene on brown adipose tissue thermogenic markers was analyzed in Zucker rats. A significant reduction in white adipose tissue and an increase in gene expression of UCP-1, PGC-1α, carnitine palmitoyl-transferase 1b (Cpt-1b), peroxisome proliferator-activated receptor α (PPAR-α), Nrf1 and Cox-2, were observed with oral administration of 15 and 30 mg/kg BW of Pterostilbene for six weeks. The increased expression of these adipocyte browning and anti-inflammatory genes resulted in thermogenesis and increased oxidative capacity in the obese rats [137]. A similar increase in genes associated with adipocyte browning i.e., Cidea, Ebf2, PGC1α, PPARγ, Sirt1, and Tbx1 were observed in HFD-fed mice [138]. In support of these observations, a recent study showed that both Pterostilbene and Resveratrol induce thermogenic activity by enhancing fatty acid oxidation and mitochondrio-genesis [139].
Oral supplementation of Pterostilbene at 15 mg/kg BW for 16 weeks to Zucker rats showed protective anti-obesity effects, improved metabolic function and structural changes in gut microbial composition. The Akkermansia and Odoribacter genus were modified in the intestine [140].
Mechanism of Action: Pterostilbene decelerates the progression of adipogenesis by decreasing the expression of C/EBPα, and PPARγ suppresses lipogenesis by decreasing fatty acid synthesis. Additionally, it acts by increasing UCP1 and NRF-1 mRNA levels in interscapular BATs from obese rats, as well as by changing the gut microbial profiles.
5.7. Withania somnifera
Withania somnifera or Ashwagandha (winter cherry) is a well-known medicinal plant in Ayurvedic medicine used for anxiety, depression and to relieve stress (Figure 8). Ashwagandha’s medicinal properties are mainly associated with bioactive ingredients such as withanolides and their glycoconjugates [141]. Withaferin A (WA) is a natural steroidal lactone present in various parts of the plant [142]. Figure 9 shows the habitat of the plant and the structure of the active constituent.
Investigations to identify molecules that can alleviate leptin resistance is an under-explored area for controlling obesity. In an elegant experiment, Liu et.al. used a connectivity map using Endoplasmic reticulum (ER) stress reduction as a tool to identify small molecules which can act as leptin sensitizers, and identified Celastrol, a pentacyclic triterpene extracted from the Tripterygium wilfordi plant, as a potent leptin sensitizer [143]. By analyzing a library of small molecules that have mRNA expression profiles similar to that of Celastrol, the same group identified another naturally occurring compound, Withaferin A, as a leptin sensitizer [143]. Withaferin A was reported to be a potent leptin sensitizer, reducing ER stress with additional beneficial effects on diabetes. Obesity induces stress in different organelles and ER stress is one major consequence of excess energy. ER stress has recently been reported to play a central role in leptin resistance [144]. The ER is responsible for secretory protein synthesis and, under excess energy levels, the ER activity is perturbed, leading to an unfolded protein response that aims to reestablish ER homeostasis [145]. Prolonged ER stress causes metabolic, inflammatory, neurodegenerative and cardiovascular diseases [146]. In diet-induced obese mice, treatment with Withaferin A was found to reduce body weight, food intake and hepatic steatosis with beneficial effects on glucose metabolism, independently of its leptin-sensitizing effect [143]. Withaferin A showed only marginal effects in ob/ob and db/db mice which are deficient in leptin signaling. The study suggested that Withaferin A may be a promising candidate for the treatment of obesity. Dietary Withaferin A for 12 weeks, significantly improved hepatic insulin sensitivity, and adipocytokines with enhanced glucose tolerance were seen in HFD induced obese mice. These results were observed along with attenuation of hepatic inflammation, oxidative stress, and insulin resistance in mice [147]. An ethanol extract of WS enriched with Withaferin A was found to suppress increases in bodyweight, serum lipids, and lipid accumulation in the liver and promote browning of subcutaneous fat in mice by increasing mitochondrial uncoupling protein 1 (UCP1) expression [148]. Withania somnifera extract was also found to increase energy expenditure by increasing oxygen consumption and thermogenesis in HFD-fed mice.
The overall mechanism action of the herbs and their active ingredients are described in Figure 9.
Mechanism of Action: Withaferin A from W. somnifera is a small molecule, which acts as a leptin sensitizer by reducing cellular ER stress. The molecule also reduces oxidative stress and inflammation. Withaferin A enriched extract induces adipocyte browning and increases oxygen consumption and energy expenditure.
The mechanism of action of the phytochemicals are given in Figure 10.
6. Other Plants and Phytochemicals with Anti-Obesity Activity
Several plant extracts with active phytochemicals have been explored for their anti-obesity activity and the list is exhaustive. Anthocyanins, the pigmented flavonoids, have anti-inflammatory, anti-diabetic and anti-obesity activities. In animals, anthocyanin supplementation reduced body weight gain, visceral fat and lipid levels, and alleviated leptin resistance [149]. Purified cyanidin-3-glucoside was found to induce thermogenesis by increasing mitochondrial UCP1 expression and activating AMPK phosphorylation [149]. Anthocyanins also have a positive interaction with gut microbiome and reduce obesity associated inflammation [150]. The fruits of Poncirus trifoliata L. contain several flavanones. Poncirin, one of the major flavanones, was found to have anti-obesity and anti-hyperglycemic effects in HFD included obese mice by modulating lipid and glucose metabolism [151].
We have covered a few of the plant extracts and their active phytochemicals with anti-obesity activity and hope that this review will provide a platform to understand the use of phytochemicals to tackle this global health issue.
7. Conclusions
Obesity, a multifactorial disorder, is the world’s third-largest prevalent disease. Various pathological features, including energy intake, energy expenditure, genetic disorder, gut microbiota imbalance and hormonal imbalance contribute to its complexity. The physiological pathogenesis, in turn, contributes to various comorbidity disorders affecting multiple parts of the body. To address the same, lifestyle management, surgical and pharmacological therapies are being widely used. The use of natural anti-obesity products may provide an alternative and safer tool in achieving weight-loss goals. Studies using isolated pure compounds and standardized natural extracts to understand the molecular mechanisms of action will enable a more scientific blending of these products for a synergistic activity for weight management. One major advantage of natural products is their multiple actions in controlling the comorbidities associated with obesity. While natural products generally show an effect in vitro and animal models, the major challenge has been in translating these results to clinical data. Future research focusing on clinical studies with well-defined endpoints would enable some of these herbal products to see the light of day in the market for human consumption.
8. Patents
US patent 10085963 (Forskolin), 10172903 (Scirpusin A, Scirpusin B and Piceatannol), 9328330 (Calebin A) are patents assigned to Sami Labs limited and Dr Muhammed Majeed for their anti-obesity activity.
Author Contributions
Conceptualization, M.M. and K.N.; resources, M.M.; writing—original draft preparation, L.M. and M.G.; writing—review and editing, M.G., K.N., L.M., S.M. and M.M. All authors have read and agreed to the published version of the manuscript.
Funding
The project was funded by Sami labs Limited and Sabinsa Corporation.
Acknowledgments
The authors acknowledge the support of all the scientists who helped in the preparation of the manuscript.
Conflicts of Interest
All the authors are affiliated to Sami Labs Limited or Sabinsa Corporation.
References
- Engin, A. The Definition and Prevalence of Obesity and Metabolic Syndrome. In Obesity and Lipotoxicity. Advances in Experimental Medicine and Biology; Ayse Basak, E., Atilla, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; Volume 960, pp. 1–17. [Google Scholar]
- Zhang, Y.; Liu, J.; Yao, J.; Ji, G.; Qian, L.; Wang, J.; Zhang, G.; Tian, J.; Nie, Y.; Zhang, Y.E.; et al. Obesity: Pathophysiology and intervention. Nutrients 2014, 6, 5153–5183. [Google Scholar] [CrossRef] [Green Version]
- Purnell, J.Q. Definitions, Classification, and Epidemiology of Obesity. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dungan, K., Grossman, A., Hershman, J.M., Hofland, H.J., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and Cardiovascular Disease: Pathophysiology, Evaluation, and Effect of Weight Loss. Circulation 2006, 113, 898–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavie, C.J.; Milani, R.V.; Ventura, H.O. Obesity and cardiovascular disease: Risk factor, paradox, and impact of weight loss. J. Am. Coll. Cardiol. 2009, 53, 1925–1932. [Google Scholar] [CrossRef] [Green Version]
- NIH. Clinical guidelines for the identification, evaluation, and treatment of overweight and obesity in adults-the evidence report. Obes. Res. 1998, 6, 51S–209S. [Google Scholar]
- Basen-Engquist, K.; Chang, M. Obesity and cancer risk: Recent review and evidence. Curr. Oncol. Rep. 2011, 13, 71–76. [Google Scholar] [CrossRef]
- Farnier, C.; Krief, S.; Blache, M.; Diot-Dupuy, F.; Mory, G.; Ferre, P.; Bazin, R. Adipocyte functions are modulated by cell size change: Potential involvement of an integrin/ERK signalling pathway. Int. J. Obes. 2003, 27, 1178–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 10 August 2020).
- Sun, N.N.; Wu, T.Y.; Chau, C.F. Natural Dietary and Herbal Products in Anti-Obesity Treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Jiang, Y.; Guo, J.; Su, Z. Natural Products with Anti-obesity Effects and Different Mechanisms of Action. J. Agric. Food Chem. 2016, 64, 9571–9585. [Google Scholar] [CrossRef]
- Karri, S.; Sharma, S.; Hatware, K.; Patil, K. Natural anti-obesity agents and their therapeutic role in management of obesity: A future trend perspective. Biomed. Pharmacother. 2019, 110, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Muller, T.D.; Clemmensen, C.; Finan, B.; DiMarchi, R.D.; Tschop, M.H. Anti-Obesity Therapy: From Rainbow Pills to Polyagonists. Pharm. Rev. 2018, 70, 712–746. [Google Scholar] [CrossRef] [Green Version]
- Kabiri, M.; Sexton Ward, A.; Ramasamy, A.; van Eijndhoven, E.; Ganguly, R.; Smolarz, B.G.; Zvenyach, T.; Goldman, D.P.; Baumgardner, J.R. The Societal Value of Broader Access to Antiobesity Medications. Obesity 2020, 28, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Onakpoya, I.J.; Heneghan, C.J.; Aronson, J.K. Post-marketing withdrawal of anti-obesity medicinal products because of adverse drug reactions: A systematic review. BMC Med. 2016, 14, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.F.; Cheung, B.M. Rise and fall of anti-obesity drugs. World J. Diabetes 2011, 2, 19–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodard, K.; Louque, L.; Hsia, D.S. Medications for the treatment of obesity in adolescents. Ther. Adv. Endocrinol. Metab. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Oussaada, S.M.; van Galen, K.A.; Cooiman, M.I.; Kleinendorst, L.; Hazebroek, E.J.; van Haelst, M.M.; ter Horst, K.W.; Serlie, M.J. The pathogenesis of obesity. Metab. Clin. Exp. 2019, 92, 26–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro Volp, A.C.; Esteves de Oliveira, F.C.; Duarte Moreira Alves, R.; Esteves, E.A.; Bressan, J. Energy expenditure: Components and evaluation methods. Nutr. Hosp. 2011, 26, 430–440. [Google Scholar] [CrossRef]
- Boron, W.; Boulpaep, E. Medical Physiology; Elsevier: Philadelphia, PA, USA, 2017. [Google Scholar]
- Sandoval, D.; Cota, D.; Seeley, R.J. The integrative role of CNS fuel-sensing mechanisms in energy balance and glucose regulation. Annu. Rev. Physiol. 2008, 70, 513–535. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, K.; Maretich, P.; Kajimura, S. The Common and Distinct FeatuRes. of Brown and Beige Adipocytes. Trends Endocrinol. Metab. 2018, 29, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Altshuler-Keylin, S.; Shinoda, K.; Hasegawa, Y.; Ikeda, K.; Hong, H.; Kang, Q.; Yang, Y.; Perera, R.M.; Debnath, J.; Kajimura, S. Beige Adipocyte Maintenance Is Regulated by Autophagy-Induced Mitochondrial Clearance. Cell Metab. 2016, 24, 402–419. [Google Scholar] [CrossRef] [Green Version]
- Cypess, A.M.; Kahn, C.R. Brown fat as a therapy for obesity and diabetes. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 143–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gesta, S.; Tseng, Y.H.; Kahn, C.R. Developmental origin of fat: Tracking obesity to its source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Hadi, H.; Di Vincenzo, A.; Vettor, R.; Rossato, M. Food Ingredients Involved in White-to-Brown Adipose Tissue Conversion and in Calorie Burning. Front. Physiol. 2019, 9, 1954. [Google Scholar] [CrossRef] [Green Version]
- Majeed, M.; Bani, S.; Pandey, A.; Nagabhushanam, K. Process and Compositions For Achieving Mammalian Energy Balance. U.S. Patent 10,085,963, 2018. [Google Scholar]
- Berthoud, H.R.; Münzberg, H.; Morrison, C.D. Blaming the Brain for Obesity: Integration of Hedonic and Homeostatic Mechanisms. Gastroenterology 2017, 152, 1728–1738. [Google Scholar] [CrossRef] [Green Version]
- Carnell, S.; Gibson, C.; Benson, L.; Ochner, C.N.; Geliebter, A. Neuroimaging and obesity: Current knowledge and future directions. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2012, 13, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Hesse, S.; van de Giessen, E.; Zientek, F.; Petroff, D.; Winter, K.; Dickson, J.C.; Tossici-Bolt, L.; Sera, T.; Asenbaum, S.; Darcourt, J.; et al. Association of central serotonin transporter availability and body mass index in healthy Europeans. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2014, 24, 1240–1247. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Thanos, P.K.; Gold, M.S. Dopamine and glucose, obesity, and reward deficiency syndrome. Front. Psychol. 2014, 5, 919. [Google Scholar] [CrossRef] [Green Version]
- Latorre, R.; Sternini, C.; De Giorgio, R.; Greenwood-Van Meerveld, B. Enteroendocrine cells: A review of their role in brain-gut communication. Neurogastroenterol. Motil. 2016, 28, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Mok, J.K.W.; Makaronidis, J.M.; Batterham, R.L. The role of gut hormones in obesity. Curr. Opin. Endocr. Metab. Res. 2019, 4, 4–13. [Google Scholar] [CrossRef]
- Dalamaga, M.; Chou, S.H.; Shields, K.; Papageorgiou, P.; Polyzos, S.A.; Mantzoros, C.S. Leptin at the intersection of neuroendocrinology and metabolism: Current evidence and therapeutic perspectives. Cell Metab. 2013, 18, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Frederich, R.C.; Hamann, A.; Anderson, S.; Lollmann, B.; Lowell, B.B.; Flier, J.S. Leptin levels reflect body lipid content in mice: Evidence for diet-induced resistance to leptin action. Nat. Med. 1995, 1, 1311–1314. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, T.; Kelesidis, I.; Chou, S.; Mantzoros, C.S. Narrative review: The role of leptin in human physiology: Emerging clinical applications. Ann. Intern. Med. 2010, 152, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Myers, M.G., Jr.; Heymsfield, S.B.; Haft, C.; Kahn, B.B.; Laughlin, M.; Leibel, R.L.; Tschop, M.H.; Yanovski, J.A. Challenges and opportunities of defining clinical leptin resistance. Cell Metab. 2012, 15, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakansson, A.; Molin, G. Gut microbiota and inflammation. Nutrients 2011, 3, 637–682. [Google Scholar] [CrossRef]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D.; Dewever, C.; Delzenne, N.M. Inulin-type fructans modulate gastrointestinal peptides involved in appetite regulation (glucagon-like peptide-1 and ghrelin) in rats. Br. J. Nutr. 2004, 92, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020. [Google Scholar] [CrossRef]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Backhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishino, S.; Takeuchi, M.; Park, S.B.; Hirata, A.; Kitamura, N.; Kunisawa, J.; Kiyono, H.; Iwamoto, R.; Isobe, Y.; Arita, M.; et al. Polyunsaturated fatty acid saturation by gut lactic acid bacteria affecting host lipid composition. Proc. Natl. Acad. Sci. USA 2013, 110, 17808–17813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Sanmiguel, C.; Gupta, A.; Mayer, E.A. Gut Microbiome and Obesity: A Plausible Explanation for Obesity. Curr. Obes. Rep. 2015, 4, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Bahceci, M.; Gokalp, D.; Bahceci, S.; Tuzcu, A.; Atmaca, S.; Arikan, S. The correlation between adiposity and adiponectin, tumor necrosis factor alpha, interleukin-6 and high sensitivity C-reactive protein levels. Is adipocyte size associated with inflammation in adults? J. Endocrinol. Investig. 2007, 30, 210–214. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Sabater, M.; Ortega, F.; Ricart, W.; Fernandez-Real, J.M. Circulating zonulin, a marker of intestinal permeability, is increased in association with obesity-associated insulin resistance. PLoS ONE 2012, 7, e37160. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, R.; Newman, T.M.; Wang, S.; Jain, S.; Lovato, J.F.; Yadav, H. Obesity-Linked Gut Microbiome Dysbiosis Associated with Derangements in Gut Permeability and Intestinal Cellular Homeostasis Independent of Diet. J. Diabetes Res. 2018, 2018, 3462092. [Google Scholar] [CrossRef] [PubMed]
- Kurashima, Y.; Goto, Y.; Kiyono, H. Mucosal innate immune cells regulate both gut homeostasis and intestinal inflammation. Eur. J. Immunol 2013, 43, 3108–3115. [Google Scholar] [CrossRef] [PubMed]
- Mutch, D.; Clément, K. Unraveling the Genetics of Human Obesity. PLoS Genet. 2007, 2, e188. [Google Scholar] [CrossRef] [Green Version]
- Farooqi, I.S.; Bullmore, E.; Keogh, J.; Gillard, J.; O’Rahilly, S.; Fletcher, P.C. Leptin regulates striatal regions and human eating behavior. Science 2007, 317, 1355. [Google Scholar] [CrossRef] [Green Version]
- Loos, R.J.; Yeo, G.S. The bigger picture of FTO: The first GWAS-identified obesity gene. Nat. Rev. Endocrinol. 2014, 10, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Thaker, V.V. Genetic and Epigentic causes of obesity. Adolesc Med State Art Rev 2017, 28, 379–405. [Google Scholar]
- Wabitsch, M.; Funcke, J.B.; Lennerz, B.; Kuhnle-Krahl, U.; Lahr, G.; Debatin, K.M.; Vatter, P.; Gierschik, P.; Moepps, B.; Fischer-Posovszky, P. Biologically inactive leptin and early-onset extreme obesity. N. Engl. J. Med. 2015, 372, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, I.S.; Wangensteen, T.; Collins, S.; Kimber, W.; Matarese, G.; Keogh, J.M.; Lank, E.; Bottomley, B.; Lopez-Fernandez, J.; Ferraz-Amaro, I.; et al. Clinical and molecular genetic spectrum of congenital deficiency of the leptin receptor. N. Engl. J. Med. 2007, 356, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Krude, H.; Biebermann, H.; Luck, W.; Horn, R.; Brabant, G.; Gruters, A. Severe early-onset obesity, adrenal insufficiency and red hair pigmentation caused by POMC mutations in humans. Nat. Genet. 1998, 19, 155–157. [Google Scholar] [CrossRef]
- Fan, W.; Boston, B.A.; Kesterson, R.A.; Hruby, V.J.; Cone, R.D. Role of melanocortinergic neurons in feeding and the agouti obesity syndrome. Nature 1997, 385, 165–168. [Google Scholar] [CrossRef]
- Ramachandrappa, S.; Raimondo, A.; Cali, A.M.; Keogh, J.M.; Henning, E.; Saeed, S.; Thompson, A.; Garg, S.; Bochukova, E.G.; Brage, S.; et al. Rare variants in single-minded 1 (SIM1) are associated with severe obesity. J. Clin. Investig. 2013, 123, 3042–3050. [Google Scholar] [CrossRef] [Green Version]
- Han, J.C.; Liu, Q.R.; Jones, M.; Levinn, R.L.; Menzie, C.M.; Jefferson-George, K.S.; Adler-Wailes, D.C.; Sanford, E.L.; Lacbawan, F.L.; Uhl, G.R.; et al. Brain-derived neurotrophic factor and obesity in the WAGR syndrome. N. Engl. J. Med. 2008, 359, 918–927. [Google Scholar] [CrossRef] [Green Version]
- Doche, M.E.; Bochukova, E.G.; Su, H.W.; Pearce, L.R.; Keogh, J.M.; Henning, E.; Cline, J.M.; Saeed, S.; Dale, A.; Cheetham, T.; et al. Human SH2B1 mutations are associated with maladaptive behaviors and obesity. J. Clin. Investig. 2012, 122, 4732–4736. [Google Scholar] [CrossRef] [Green Version]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Sarjeant, K.; Stephens, J.M. Adipogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scimè, A.; Devarakonda, S.; Conroe, H.M.; Erdjument-Bromage, H.; et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008, 454, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Frontini, A.; Cinti, S. Distribution and development of brown adipocytes in the murine and human adipose organ. Cell Metab. 2010, 11, 253–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saper, R.B.; Eisenberg, D.M.; Phillips, R.S. Common dietary supplements for weight loss. Am. Fam. Physician 2004, 70, 1731–1738. [Google Scholar]
- Dyck, D.J. Dietary fat intake, supplements, and weight loss. Can. J. Appl. Physiol. Rev. Can. Physiol. Appl. 2000, 25, 495–523. [Google Scholar] [CrossRef]
- Burns, T.W.; Langley, P.E.; Terry, B.E.; Bylund, D.B.; Forte, L.R. Comparative effects of forskolin and isoproterenol on the cyclic AMP content of human adipocytes. Life Sci. 1987, 40, 145–154. [Google Scholar] [CrossRef]
- Kamohara, S. An evidence-based review: Anti-obesity effects of Coleus forskohlii. Pers. Med. Universe 2016, 5, 16–20. [Google Scholar] [CrossRef]
- Tsuguyoshi, A. Clinical Report on Root Extract of Perilla Plant (Coleus forskohlii) ForsLean® in Reducing Body Fat. Available online: http://www.herbamedicabg.com/media/forslean/01-abstract-japan.pdf (accessed on 26 August 2015).
- Badmaev, V.; Majeed, M.; Conte, A.A.; Parker, J.E. Diterpene Forskolin (Coleus forskohlii, Benth.): A Possible New Compound for Reduction of Body Weight by Increasing Lean Body Mass. NutraCos 2002, 1, 6–7. [Google Scholar]
- Loftus, H.L.; Astell, K.J.; Mathai, M.L.; Su, X.Q. Coleus forskohlii Extract Supplementation in Conjunction with a Hypocaloric Diet Reduces the Risk Factors of Metabolic Syndrome in Overweight and Obese Subjects: A Randomized Controlled Trial. Nutrients 2015, 7, 9508–9522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godard, M.P.; Johnson, B.A.; Richmond, S.R. Body Composition and Hormonal Adaptations Associated with Forskolin Consumption in Overweight and Obese Men. Obes. Res. 2005, 13, 1335–1343. [Google Scholar] [CrossRef] [Green Version]
- Henderson, S.; Magu, B.; Rasmussen, C.; Lancaster, S.; Kerksick, C.; Smith, P.; Melton, C.; Cowan, P.; Greenwood, M.; Earnest, C.; et al. Effects of Coleus forskohlii supplementation on body composition and hematological profiles in mildly overweight women. J. Int. Soc. Sports Nutr. 2005, 2, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majeed, M. Coleus forskohlii Extract in the Management of Obesity, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Lampidonis, A.D.; Rogdakis, E.; Voutsinas, G.E.; Stravopodis, D.J. The resurgence of Hormone-Sensitive Lipase (HSL) in mammalian lipolysis. Gene 2011, 477, 1–11. [Google Scholar] [CrossRef]
- Montanari, T.; Pošćić, N.; Colitti, M. Factors involved in white-to-brown adipose tissue conversion and in thermogenesis: A review. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2017, 18, 495–513. [Google Scholar] [CrossRef]
- Yin, W.; Mu, J.; Birnbaum, M.J. Role of AMP-activated protein kinase in cyclic AMP-dependent lipolysis in 3T3-L1 adipocytes. J. Biol. Chem. 2003, 278, 43074–43080. [Google Scholar] [CrossRef] [Green Version]
- Sergio, W. A natural food, the malabar tamarind, may be effective in the treatment of obesity. Med. Hypotheses 1988, 27, 39–40. [Google Scholar] [CrossRef]
- Ananthakrishnan, R.; Rameshkumar, K. Phytochemicals and bioactivities of Garcinia indica (Thouars) Choisy-A review. Divers. Garcinia Species West. Ghats: Phytochem. Perspect. 2016, 142, 151–161. [Google Scholar]
- Parthasarathy, U.; Nandakishore, O.; Kumar, R.S.; Babu, K.N.; Zachariah, T.J.; Parthasarathy, V. Chromatographic fingerprinting and estimation of organic acids in selected Garcinia species. Int. J. Innov. Hortic. 2012, 1, 68–73. [Google Scholar]
- Kaur, R.; Chattopadhyay, S.K.; Tandon, S.; Sharma, S. Large scale extraction of the fruits of Garcinia indica for the isolation of new and known polyisoprenylated benzophenone derivatives. Ind. Crop. Prod. 2012, 37, 420–426. [Google Scholar] [CrossRef]
- Pandey, R.; Chandra, P.; Kumar, B.; Srivastva, M.; Aravind, A.A.; Shameer, P.; Rameshkumar, K. Simultaneous determination of multi-class bioactive constituents for quality assessment of Garcinia species using UHPLC–QqQLIT–MS/MS. Ind. Crop. Prod. 2015, 77, 861–872. [Google Scholar] [CrossRef]
- Jena, B.; Jayaprakasha, G.; Singh, R.; Sakariah, K. Chemistry and biochemistry of (−)-hydroxycitric acid from Garcinia. J. Agric. Food Chem. 2002, 50, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, P.; Bhise, K.; Prakya, V. A phytopharmacological review on Garcinia indica. Int. J. Herb. Med. 2015, 3, 2–7. [Google Scholar]
- Majeed, M.; Majeed, S.; Nagabhushanam, K.; Lawrence, L.; Mundkur, L. Garcinia indica extract standardized for 20% Garcinol reduces adipogenesis and high fat diet-induced obesity in mice by alleviating endoplasmic reticulum stress. J. Funct. Foods 2020, 67, 103863. [Google Scholar] [CrossRef]
- Lee, P.S.; Teng, C.Y.; Kalyanam, N.; Ho, C.T.; Pan, M.H. Garcinol Reduces Obesity in High-Fat-Diet-Fed Mice by Modulating Gut Microbiota Composition. Mol. Nutr. Food Res. 2019, 63, e1800390. [Google Scholar] [CrossRef]
- Schneeberger, M.; Everard, A.; Gomez-Valades, A.G.; Matamoros, S.; Ramirez, S.; Delzenne, N.M.; Gomis, R.; Claret, M.; Cani, P.D. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci. Rep. 2015, 5, 16643. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.; Seo, M.; Park, H.; Lee, W.K.; Guan, L.L.; Yoon, J.; Caetano-Anolles, K.; Ahn, H.; Kim, S.Y.; Kang, Y.M.; et al. Gut microbiota Modulated by Probiotics and Garcinia cambogia Extract Correlate with Weight Gain and Adipocyte Sizes in High Fat-Fed Mice. Sci. Rep. 2016, 6, 33566. [Google Scholar] [CrossRef]
- Peerzada, A.M.; Ali, H.H.; Naeem, M.; Latif, M.; Bukhari, A.H.; Tanveer, A. Cyperus rotundus L.: Traditional uses, phytochemistry, and pharmacological activities. J. Ethnopharmacol. 2015, 174, 540–560. [Google Scholar] [CrossRef]
- Allan, R.D.; Wells, R.J.; Correll, R.L.; MacLeod, J.K. The presence of quinones in the genus Cyperus as an aid to classification. Phytochemistry 1978, 17, 263–266. [Google Scholar] [CrossRef]
- Ito, T.; Endo, H.; Shinohara, H.; Oyama, M.; Akao, Y.; Iinuma, M. Occurrence of stilbene oligomers in Cyperus rhizomes. Fitoterapia 2012, 83, 1420–1429. [Google Scholar] [CrossRef] [PubMed]
- Lemaure, B.; Touche, A.; Zbinden, I.; Moulin, J.; Courtois, D.; Mace, K.; Darimont, C. Administration of Cyperus rotundus tubers extract prevents weight gain in obese Zucker rats. Phytother.Res. 2007, 21, 724–730. [Google Scholar] [CrossRef]
- Athesh, K.; Divakar, M.; Brindha, P. Anti-obesity potential of Cyperus rotundus L. aqueous tuber extract in rats fed on high fat cafeteria diet. Asian J. Pharm.Clin. Res. 2014, 7, 88–92. [Google Scholar]
- Nagulendran, K.; Ramalingam, M.; Vava Mohideen, H. Preventive role of Cyperus rotundus rhizomes extract on age associated changes in glucose and lipids. Pharmacologyonline 2007, 2, 318–325. [Google Scholar]
- Chandratre, R.; Chandarana, S.; Mengi, S. Lipid lowering activity of alcoholic extract of Cyperus rotundus. IJRPC 2011, 1, 1042–1045. [Google Scholar]
- Majeed, M.; Nagabhushanam, K.; Kalman, D.; Bhat, B.; Vaidyanathan, P.; Bani, S.; Anjali, P.; Karri, S. Composition Comprising Scirpusin A and Scirpusin B and Anti-Obesity Potential Thereof. U.S. Patent 10,172,903, 2019. [Google Scholar]
- Sano, S.; Sugiyama, K.; Ito, T.; Katano, Y.; Ishihata, A. Identification of the strong vasorelaxing substance scirpusin B, a dimer of piceatannol, from passion fruit (Passiflora edulis) seeds. J. Agric. Food Chem. 2011, 59, 6209–6213. [Google Scholar] [CrossRef]
- Pan, Z.-H.; Ning, D.-S.; Fu, Y.-X.; Li, D.-P.; Zou, Z.-Q.; Xie, Y.-C.; Yu, L.-L.; Li, L.-C. Preparative Isolation of Piceatannol Derivatives from Passion Fruit (Passiflora edulis) Seeds by High-Speed Countercurrent Chromatography Combined with High-Performance Liquid Chromatography and Screening for α-Glucosidase Inhibitory Activities. J. Agric. Food Chem. 2020, 68, 1555–1562. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Seo, S.G.; Heo, Y.S.; Yue, S.; Cheng, J.X.; Lee, K.W.; Kim, K.H. Piceatannol, natural polyphenolic stilbene, inhibits adipogenesis via modulation of mitotic clonal expansion and insulin receptor-dependent insulin signaling in early phase of differentiation. J. Biol. Chem. 2012, 287, 11566–11578. [Google Scholar] [CrossRef] [Green Version]
- Tung, Y.C.; Lin, Y.H.; Chen, H.J.; Chou, S.C.; Cheng, A.C.; Kalyanam, N.; Ho, C.T.; Pan, M.H. Piceatannol Exerts Anti-Obesity Effects in C57BL/6 Mice through Modulating Adipogenic Proteins and Gut Microbiota. Molecules 2016, 21, 1419. [Google Scholar] [CrossRef] [Green Version]
- Hijona, E.; Aguirre, L.; Perez-Matute, P.; Villanueva-Millan, M.J.; Mosqueda-Solis, A.; Hasnaoui, M.; Nepveu, F.; Senard, J.M.; Bujanda, L.; Aldamiz-Echevarria, L.; et al. Limited beneficial effects of piceatannol supplementation on obesity complications in the obese Zucker rat: Gut microbiota, metabolic, endocrine, and cardiac aspects. J. Physiol. Biochem. 2016, 72, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Llarena, M.; Andrade, F.; Hasnaoui, M.; Portillo, M.P.; Perez-Matute, P.; Arbones-Mainar, J.M.; Hijona, E.; Villanueva-Millan, M.J.; Aguirre, L.; Carpene, C.; et al. Potential renoprotective effects of piceatannol in ameliorating the early-stage nephropathy associated with obesity in obese Zucker rats. J. Physiol. Biochem. 2016, 72, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, P.; Chang, Q.; Wang, J.; Liu, J.; Lv, Y.; Wang, T.T.Y.; Gao, B.; Zhang, Y.; Yu, L.L. Inhibitory Effect of Piceatannol on TNF-α-Mediated Inflammation and Insulin Resistance in 3T3-L1 Adipocytes. J. Agric. Food Chem. 2017, 65, 4634–4641. [Google Scholar] [CrossRef] [PubMed]
- Mompeo, O.; Spector, T.D.; Matey Hernandez, M.; Le Roy, C.; Istas, G.; Le Sayec, M.; Mangino, M.; Jennings, A.; Rodriguez-Mateos, A.; Valdes, A.M.; et al. Consumption of Stilbenes and Flavonoids is Linked to Reduced Risk of Obesity Independently of Fiber Intake. Nutrients 2020, 12, 1871. [Google Scholar] [CrossRef]
- Tung, B.T.; Nham, D.T.; Hai, N.T.; Thu, D.K. Chapter 10—Curcuma longa, the Polyphenolic Curcumin Compound and Pharmacological Effects on Liver. In Dietary Interventions in Liver Disease; Watson, R.R., Preedy, V.R., Eds.; Academic Press: San Diego, CA, USA, 2019; pp. 125–134. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Nema, N.K.; Pandit, S.; Mukherjee, K. Chapter 23—Indian Medicinal Plants with Hypoglycemic Potential. In Bioactive Food as Dietary Interventions for Diabetes; Watson, R.R., Preedy, V.R., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 235–264. [Google Scholar] [CrossRef]
- Prasad, S.; Turmeric, A.B. The Golden Spice: From Traditional Medicine to Modern Medicine. In Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; Benzie IFF, W.-G.S., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2011. [Google Scholar]
- Huang, C.; Lu, H.F.; Chen, Y.H.; Chen, J.C.; Chou, W.H.; Huang, H.C. Curcumin, demethoxycurcumin, and bisdemethoxycurcumin induced caspase-dependent and -independent apoptosis via Smad or Akt signaling pathways in HOS cells. BMC Complement. Med. Ther. 2020, 20, 68. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, M.; Hatamipour, M.; Sahebkar, A. Promising anti-tumor properties of bisdemethoxycurcumin: A naturally occurring curcumin analogue. J. Cell. Physiol. 2018, 233, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.S.; Chen, Y.Y.; Lee, P.S.; Kalyanam, N.; Ho, C.T.; Liou, W.S.; Yu, R.C.; Pan, M.H. Bisdemethoxycurcumin Inhibits Adipogenesis in 3T3-L1 Preadipocytes and Suppresses Obesity in High-Fat Diet-Fed C57BL/6 Mice. J. Agric. Food Chem. 2016, 64, 821–830. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, D.S. Discovery of natural products from Curcuma longa that protect cells from beta-amyloid insult: A drug discovery effort against Alzheimer’s disease. J. Nat. Prod. 2002, 65, 1227–1231. [Google Scholar] [CrossRef]
- Li, Y.; Li, S.; Han, Y.; Liu, J.; Zhang, J.; Li, F.; Wang, Y.; Liu, X.; Yao, L. Calebin-A induces apoptosis and modulates MAPK family activity in drug resistant human gastric cancer cells. Eur. J. Pharm. 2008, 591, 252–258. [Google Scholar] [CrossRef]
- Zeng, Y.; Qiu, F.; Takahashi, K.; Liang, J.; Qu, G.; Yao, X. New sesquiterpenes and calebin derivatives from Curcuma longa. Chem. Pharm. Bull. 2007, 55, 940–943. [Google Scholar] [CrossRef] [Green Version]
- Leisch, H.; Morley, K.; Lau, P.C. Baeyer-Villiger monooxygenases: More than just green chemistry. Chem. Rev. 2011, 111, 4165–4222. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.S.; Liao, S.N.; Tsai, M.L.; Kalyanam, N.; Majeed, M.; Majeed, A.; Ho, C.T.; Pan, M.H. Calebin-A inhibits adipogenesis and hepatic steatosis in high-fat diet-induced obesity via activation of AMPK signaling. Mol. Nutr. Food Res. 2015, 59, 1883–1895. [Google Scholar] [CrossRef]
- Harminder, V.S.; Chaudhary, A.K. A Review on the Taxonomy, Ethnobotany, Chemistry and Pharmacology of Oroxylum indicum Vent. Indian J. Pharm. Sci. 2011, 73, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Ahad, A.; Ganai, A.; Sareer, O.; Najm, M.; Kausar, A.; Mujeeb, M.; Siddiqui, W. Therapeutic potential of Oroxylum indicum: A review. J. Pharm. Res. Opin. 2012, 2, 163–172. [Google Scholar]
- Kottapalli, J.; Majeti, P. Genetic diversity in Oroxylum indicum (L.) Vent. (Bignoniaceae), a vulnerable medicinal plant by random amplified polymorphic DNA marker. Afr. J. Biotechnol. 2007, 7, 254–262. [Google Scholar]
- Dinda, B.; Mohanta, B.C.; Arima, S.; Sato, N.; Harigaya, Y. Flavonoids from the stem-bark of Oroxylum indicum. Nat. Prod. Sci. 2007, 13, 190–194. [Google Scholar]
- Majeed, M.; Nagabhushanam, K.; Ramanujam, R.; Hemantha, H.P. Process for synthesis of Oroxylin A. Google Patents WO2020032913A1, 2019. [Google Scholar]
- Hengpratom, T.; Lowe, G.M.; Thumanu, K.; Suknasang, S.; Tiamyom, K.; Eumkeb, G. Oroxylum indicum (L.) Kurz extract inhibits adipogenesis and lipase activity in vitro. BMC Complement. Altern. Med. 2018, 18, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangal, P.; Khare, P.; Jagtap, S.; Bishnoi, M.; Kondepudi, K.K.; Bhutani, K.K. Screening of six Ayurvedic medicinal plants for anti-obesity potential: An investigation on bioactive constituents from Oroxylum indicum (L.) Kurz bark. J. Ethnopharmacol. 2017, 197, 138–146. [Google Scholar] [CrossRef]
- Hengpratom, T.; Ngernsoungnern, A.; Ngernsoungnern, P.; Lowe, G.M.; Eumkeb, G. Antiadipogenesis of Oroxylum indicum (L.) Kurz Extract via PPARγ2 in 3T3-L1 Adipocytes. Evid. Based Complement. Altern. Med. 2020, 2020, 6720205. [Google Scholar] [CrossRef] [Green Version]
- Katiyar, D.; Singh, V.; Ali, M. Phytochemical and pharmacological profile of Pterocarpus marsupium: A review. Pharm. Innov. J. 2016, 5, 31–39. [Google Scholar]
- Tiwari, M.; Sharma, M.; Khare, H. Chemical constituents and medicinal uses of Pterocarpus marsupium roxb. Flora Fauna 2015, 21, 5559. [Google Scholar]
- Gómez-Zorita, S.; Fernández-Quintela, A.; Lasa, A.; Aguirre, L.; Rimando, A.M.; Portillo, M.P. Pterostilbene, a dimethyl ether derivative of resveratrol, reduces fat accumulation in rats fed an obesogenic diet. J. Agric. Food Chem. 2014, 62, 8371–8378. [Google Scholar] [CrossRef]
- Gomez-Zorita, S.; Belles, C.; Briot, A.; Fernández-Quintela, A.; Portillo, M.P.; Carpéné, C. Pterostilbene inhibits lipogenic activity similar to resveratrol or caffeine but differently modulates lipolysis in adipocytes. Phytother. Res. 2017, 31, 1273–1282. [Google Scholar] [CrossRef]
- Hsiao, Y.H.; Chen, N.C.; Koh, Y.C.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Pterostilbene Inhibits Adipocyte Conditioned-Medium-Induced Colorectal Cancer Cell Migration through Targeting FABP5-Related Signaling Pathway. J. Agric. Food Chem. 2019, 67, 10321–10329. [Google Scholar] [CrossRef]
- Aguirre, L.; Milton-Laskibar, I.; Hijona, E.; Bujanda, L.; Rimando, A.M.; Portillo, M.P. Effects of pterostilbene in brown adipose tissue from obese rats. J. Physiol. Biochem. 2016, 73, 457–464. [Google Scholar] [CrossRef]
- La Spina, M.; Galletta, E.; Azzolini, M.; Gomez Zorita, S.; Parrasia, S.; Salvalaio, M.; Salmaso, A.; Biasutto, L. Browning effects of a chronic pterostilbene supplementation in mice fed a high-fat diet. Int. J. Mol. Sci. 2019, 20, 5377. [Google Scholar] [CrossRef] [Green Version]
- Milton-Laskíbar, I.; Gómez-Zorita, S.; Arias, N.; Romo-Miguel, N.; González, M.; Fernández-Quintela, A.; Portillo, M.P. Effects of resveratrol and its derivative pterostilbene on brown adipose tissue thermogenic activation and on white adipose tissue browning process. J. Physiol. Biochem. 2020, 76, 269–278. [Google Scholar] [CrossRef]
- Etxeberria, U.; Hijona, E.; Aguirre, L.; Milagro, F.I.; Bujanda, L.; Rimando, A.M.; Martínez, J.A.; Portillo, M.P. Pterostilbene-induced changes in gut microbiota composition in relation to obesity. Mol. Nutr. Food Res. 2017, 61, 1500906. [Google Scholar] [CrossRef] [PubMed]
- Alam, N.; Hossain, M.; Khalil, M.I.; Moniruzzaman, M.; Sulaiman, S.A.; Gan, S.H. Recent advances in elucidating the biological properties of Withania somnifera and its potential role in health benefits. Phytochem. Rev. 2012, 11, 97–112. [Google Scholar] [CrossRef]
- Mirjalili, M.H.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Palazon, J. Steroidal lactones from Withania somnifera, an ancient plant for novel medicine. Molecules 2009, 14, 2373–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Liu, J.; Feng, X.; Salazar Hernandez, M.A.; Mucka, P.; Ibi, D.; Choi, J.W.; Ozcan, U. Withaferin A is a leptin sensitizer with strong antidiabetic properties in mice. Nat. Med. 2016, 22, 1023–1032. [Google Scholar] [CrossRef] [Green Version]
- Ozcan, U.; Cao, Q.; Yilmaz, E.; Lee, A.H.; Iwakoshi, N.N.; Ozdelen, E.; Tuncman, G.; Gorgun, C.; Glimcher, L.H.; Hotamisligil, G.S. Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science 2004, 306, 457–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Ozcan, U. Unfolded protein response signaling and metabolic diseases. J. Biol. Chem. 2014, 289, 1203–1211. [Google Scholar] [CrossRef] [Green Version]
- Abu Bakar, M.H.; Azmi, M.N.; Shariff, K.A.; Tan, J.S. Withaferin A Protects Against High-Fat Diet-Induced Obesity Via Attenuation of Oxidative Stress, Inflammation, and Insulin Resistance. Appl. Biochem. Biotechnol. 2019, 188, 241–259. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Ahn, J.; Jang, Y.-J.; Seo, H.-D.; Ha, T.-Y.; Kim, M.J.; Huh, Y.H.; Jung, C.H. Withania somnifera Extract Enhances Energy Expenditure via Improving Mitochondrial Function in Adipose Tissue and Skeletal Muscle. Nutrients 2020, 12, 431. [Google Scholar] [CrossRef] [Green Version]
- Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. The Influence of Supplementation of Anthocyanins on Obesity-Associated Comorbidities: A Concise Review. Foods 2020, 9, 687. [Google Scholar] [CrossRef]
- Xie, L.; Su, H.; Sun, C.; Zheng, X.; Chen, W. Recent advances in understanding the anti-obesity activity of anthocyanins and their biosynthesis in microorganisms. Trends Food Sci. Technol. 2018, 72, 13–24. [Google Scholar] [CrossRef]
- Jia, S.; Gao, Z.; Yan, S.; Chen, Y.; Sun, C.; Li, X.; Chen, K. Anti-Obesity and Hypoglycemic Effects of Poncirus trifoliata L. Extracts in High-Fat Diet C57BL/6 Mice. Molecules 2016, 21, 453. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Various parameters affecting the pathology of obesity. [SCFA: short chain fatty acids].
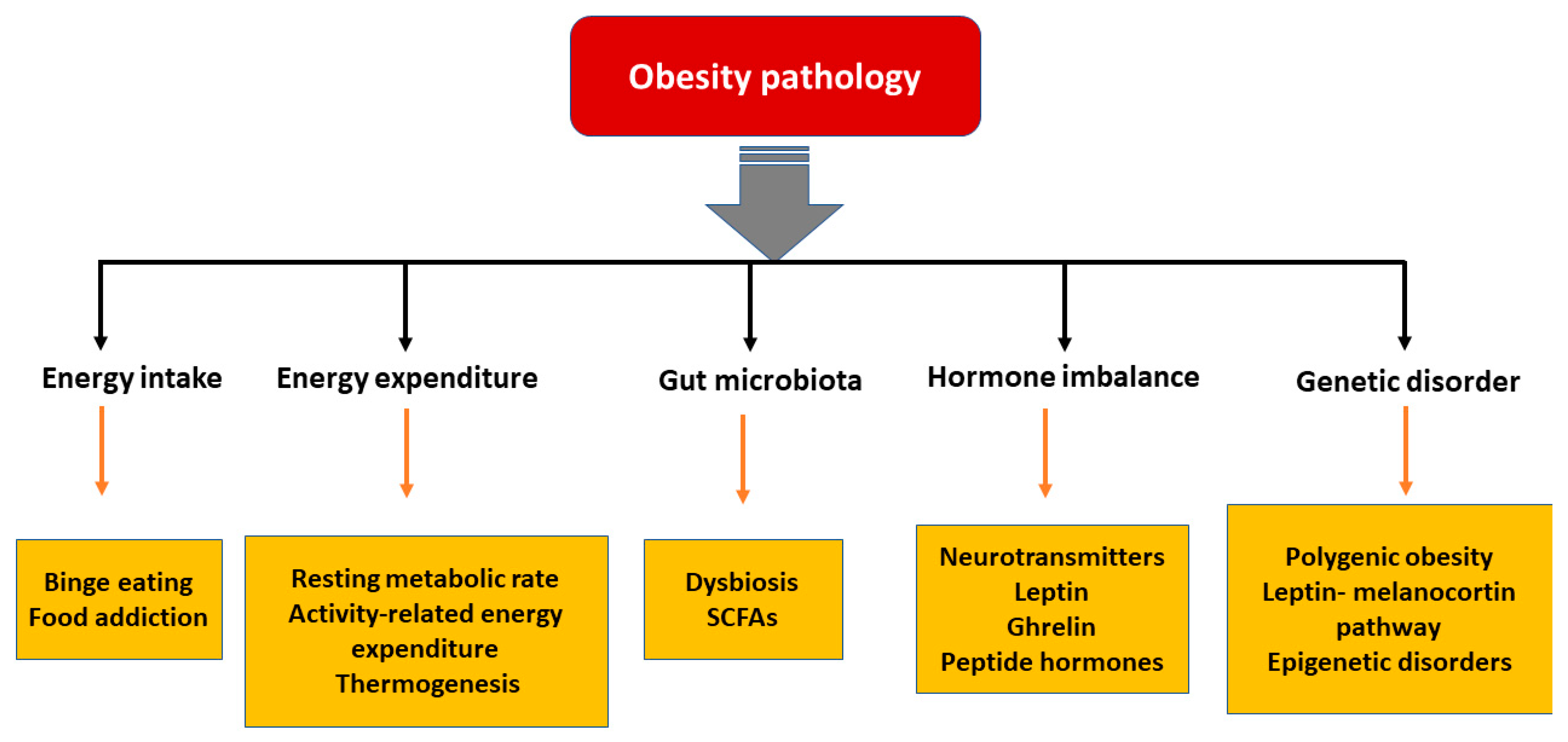
Figure 2.
Schematic representation of Gut microbiome and hormonal control of obesity: Ghrelin, produced in the stomach, is a potent stimulator of appetite in the brain. Insulin from the pancreas and leptin from adipose tissue act on the brain to induce satiety. Gut-derived peptides such as GLP-1 augment insulin release from the pancreas. The gut microbiome balances energy homeostasis by releasing short-chain fatty acids with numerous benefits. Dysbiosis of the gut decreases the peptide hormone and increases gut inflammation, which can induce insulin and leptin resistance preventing satiety. In the brain, the hypothalamus region controls energy homeostasis while the cortex and limbal areas are responsible for cognitive reward control of food intake. CCK, cholecystokinin; GLP-1, glucagon-like peptide–1; OXY, oxyntomodulin; PYY, peptide YY; SCFA, short-chain fatty acids. Blue lines represent positive stimulation to control fat storage while red lines represent the causes of obesity.
Figure 2.
Schematic representation of Gut microbiome and hormonal control of obesity: Ghrelin, produced in the stomach, is a potent stimulator of appetite in the brain. Insulin from the pancreas and leptin from adipose tissue act on the brain to induce satiety. Gut-derived peptides such as GLP-1 augment insulin release from the pancreas. The gut microbiome balances energy homeostasis by releasing short-chain fatty acids with numerous benefits. Dysbiosis of the gut decreases the peptide hormone and increases gut inflammation, which can induce insulin and leptin resistance preventing satiety. In the brain, the hypothalamus region controls energy homeostasis while the cortex and limbal areas are responsible for cognitive reward control of food intake. CCK, cholecystokinin; GLP-1, glucagon-like peptide–1; OXY, oxyntomodulin; PYY, peptide YY; SCFA, short-chain fatty acids. Blue lines represent positive stimulation to control fat storage while red lines represent the causes of obesity.
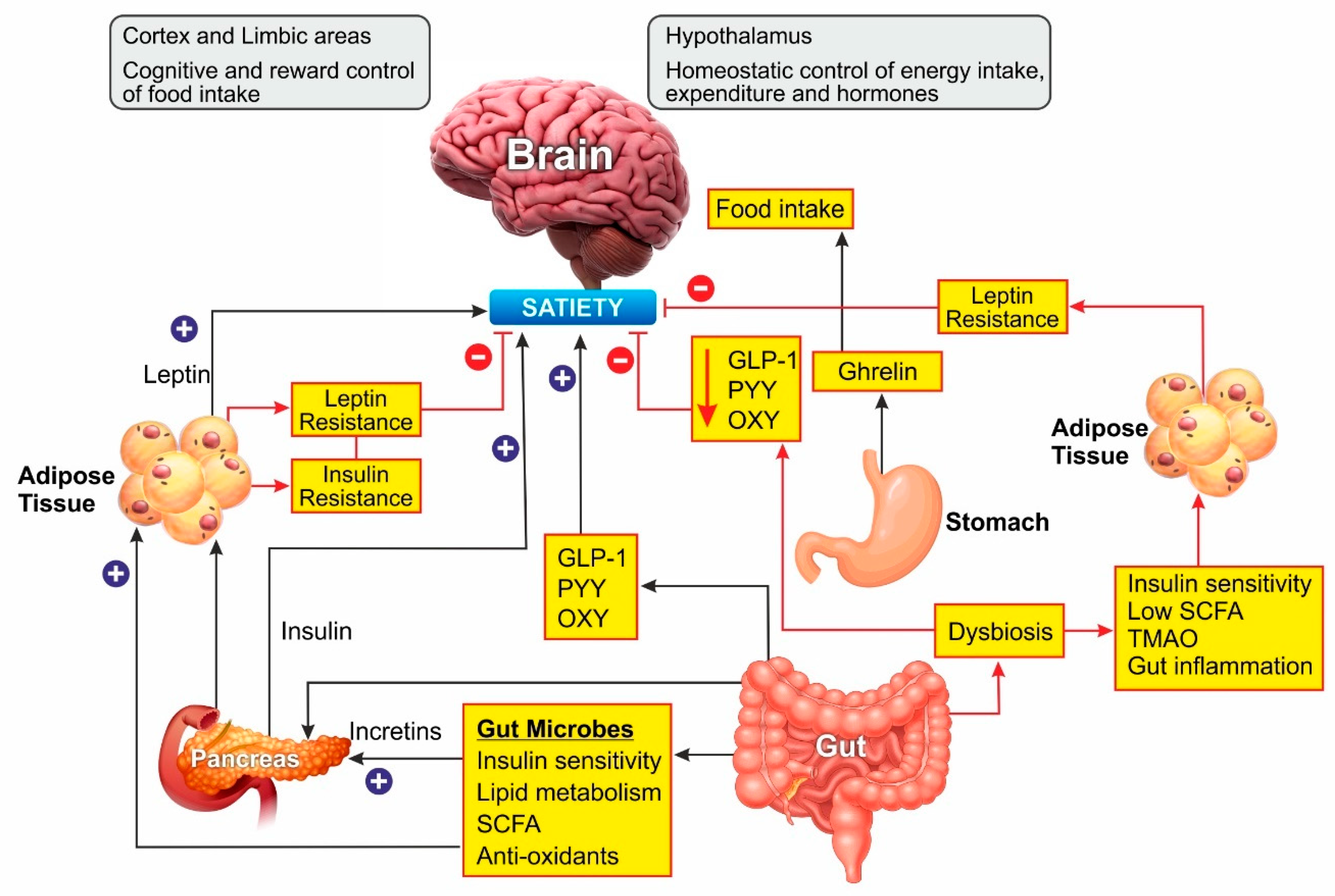
Figure 3.
Coleus forskohlii. (A) leaves; (B) roots; (C) Forskolin.
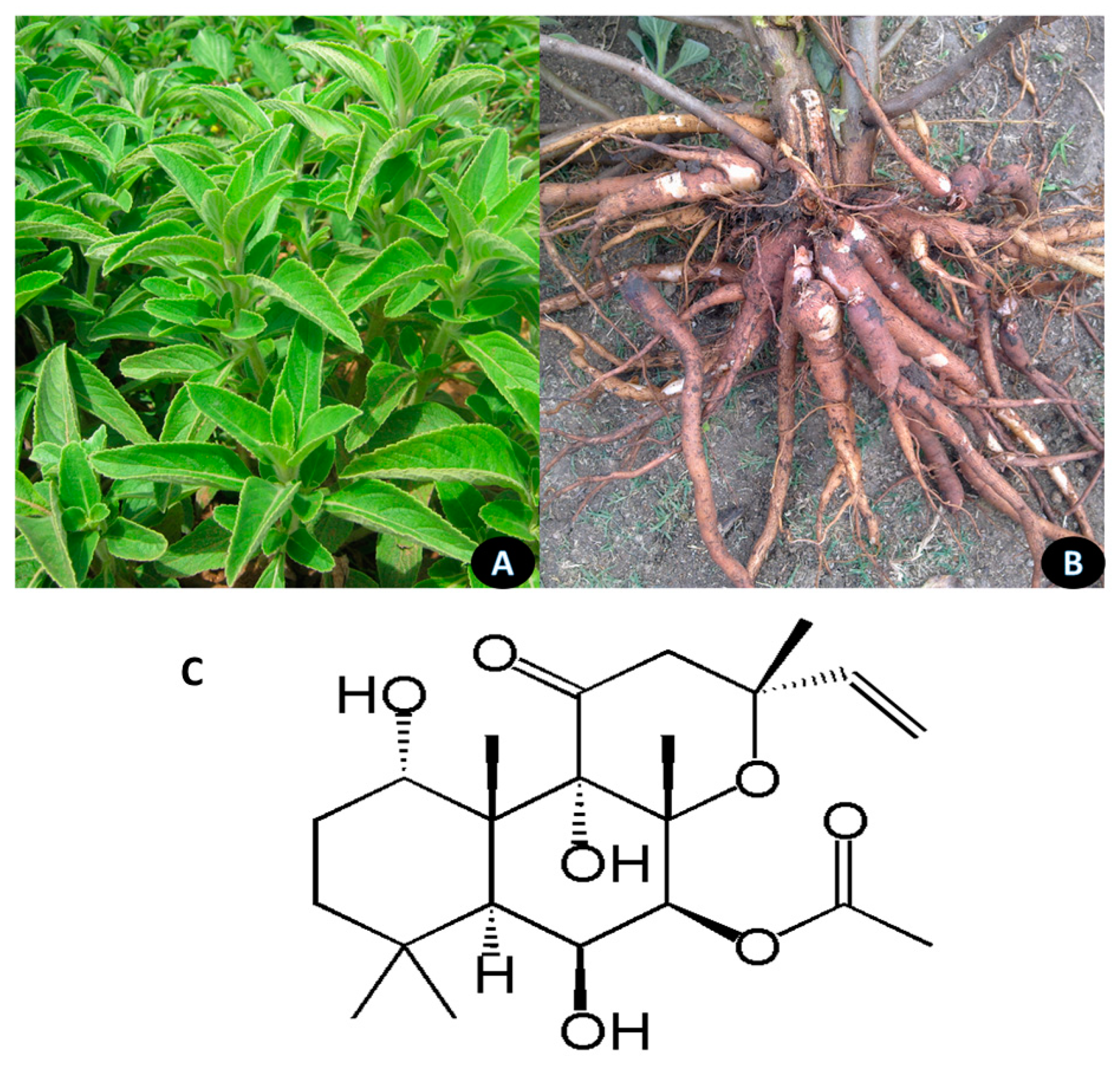
Figure 4.
Garcinia indica. (A) Habitat of the Plant; (B) Ripe fruits; (C) Dried fruit rinds; (D) Hydroxy citric acid; (E) Garcinol.
Figure 4.
Garcinia indica. (A) Habitat of the Plant; (B) Ripe fruits; (C) Dried fruit rinds; (D) Hydroxy citric acid; (E) Garcinol.

Figure 5.
(A) Cyperus rotundus. Habit of the Plant, and Enlarged view of rhizomes. (B) Passiflora edulis; habit, enlarged fruit, flower and seeds, (C) Piceatannol, (D) Scirpusin A, (E) Scirpusin B.
Figure 5.
(A) Cyperus rotundus. Habit of the Plant, and Enlarged view of rhizomes. (B) Passiflora edulis; habit, enlarged fruit, flower and seeds, (C) Piceatannol, (D) Scirpusin A, (E) Scirpusin B.

Figure 6.
Curcuma longa. (A) Habit of the Plant; Bunch of fingers; and dried rhizome slice (B) Bisdemethoxycurcumin (BDMC), (C) Calebin A.
Figure 6.
Curcuma longa. (A) Habit of the Plant; Bunch of fingers; and dried rhizome slice (B) Bisdemethoxycurcumin (BDMC), (C) Calebin A.

Figure 7.
Oroxylum indicum. (A) Habitat of the plant; (B) Fruits; (C) Dried bark; (D–F) Oroxylin A, Baicalein and Chrysin.
Figure 7.
Oroxylum indicum. (A) Habitat of the plant; (B) Fruits; (C) Dried bark; (D–F) Oroxylin A, Baicalein and Chrysin.

Figure 8.
Pterocarpus marsupium. (A) Habitat of the plant; (B) A portion of shoot showing inflorescence and flowers; (C) Enlarged view of leaves; (D) Wood slices and major actives, (E) Pterostilbene.
Figure 8.
Pterocarpus marsupium. (A) Habitat of the plant; (B) A portion of shoot showing inflorescence and flowers; (C) Enlarged view of leaves; (D) Wood slices and major actives, (E) Pterostilbene.
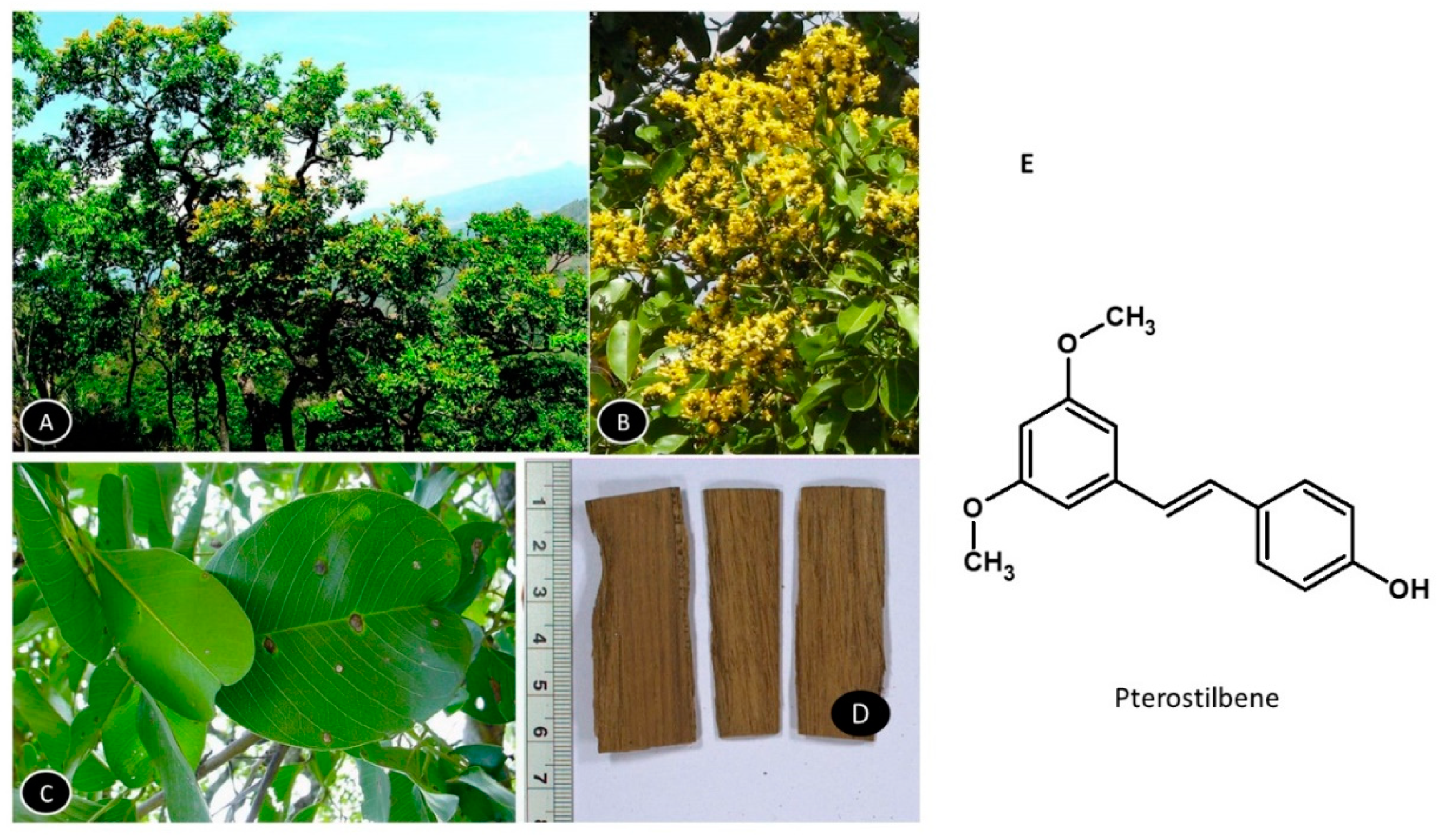
Figure 9.
Withania somnifera. (A) Habitat of the plant; (B) Dried stem; (C) Withaferin A.
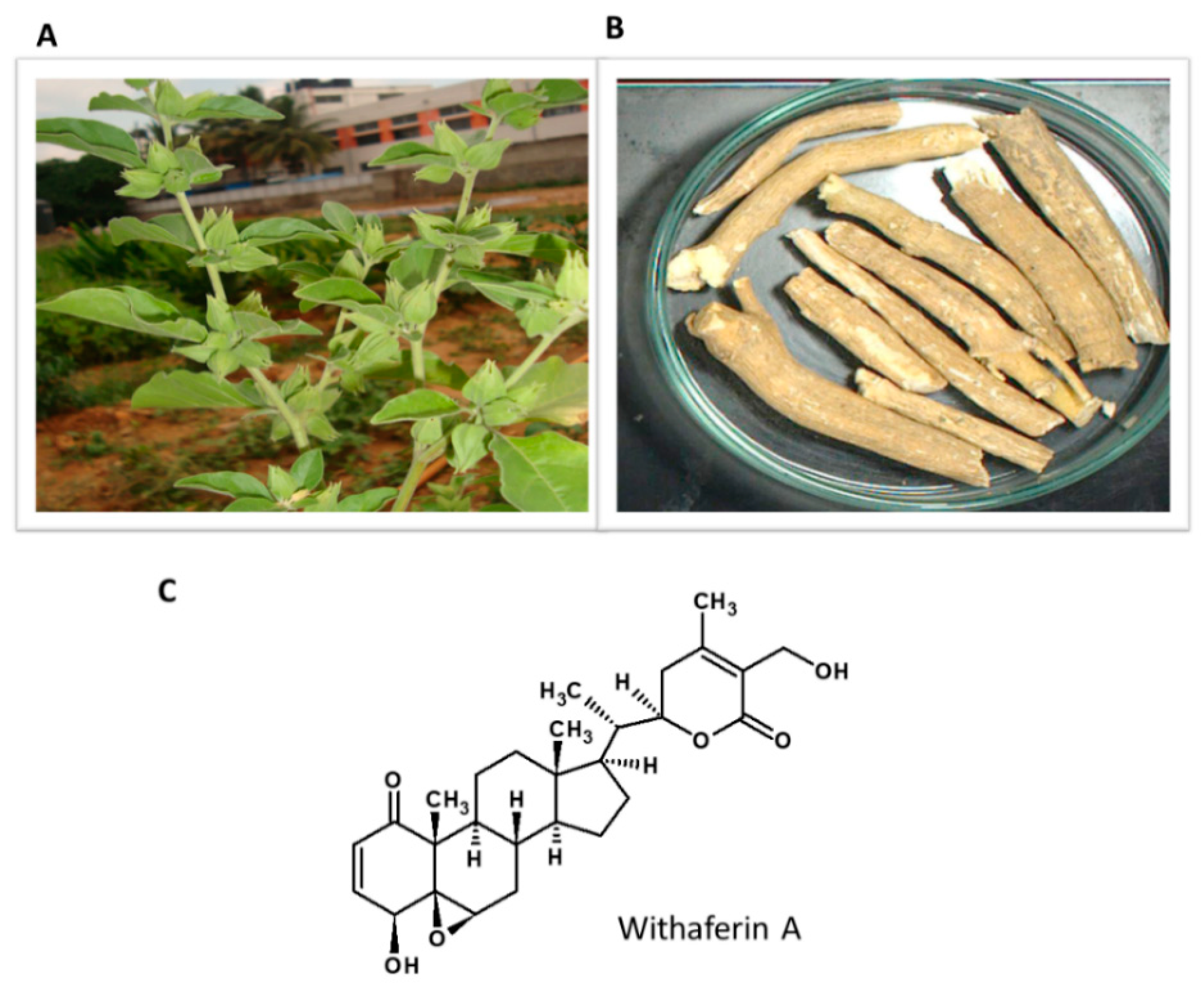
Figure 10.
Molecular pathways modulated by the active ingredients in modulating and regulating the obesity related genes and pathways. (┴) Inhibited; (↑) Promoted.
Figure 10.
Molecular pathways modulated by the active ingredients in modulating and regulating the obesity related genes and pathways. (┴) Inhibited; (↑) Promoted.
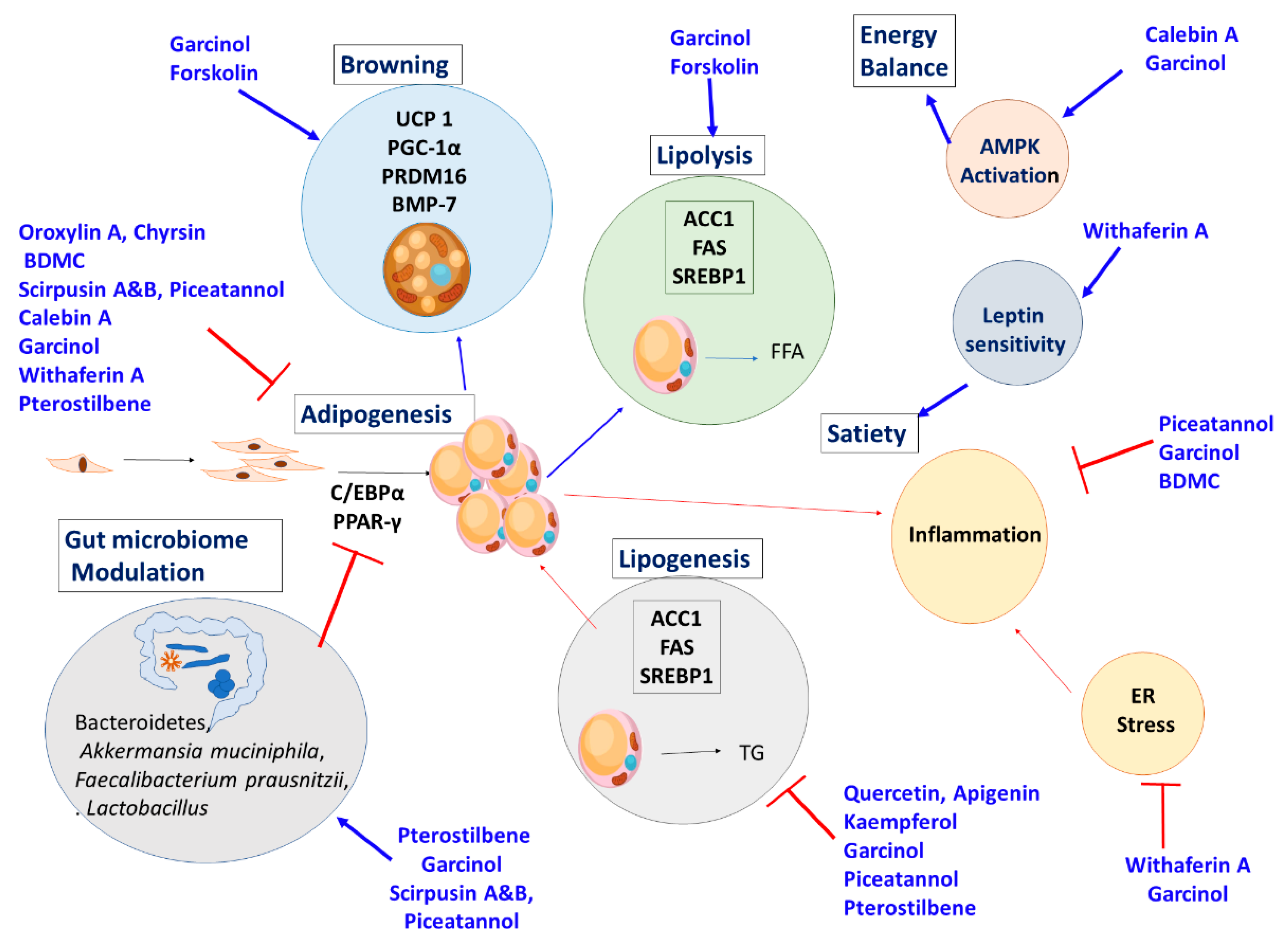

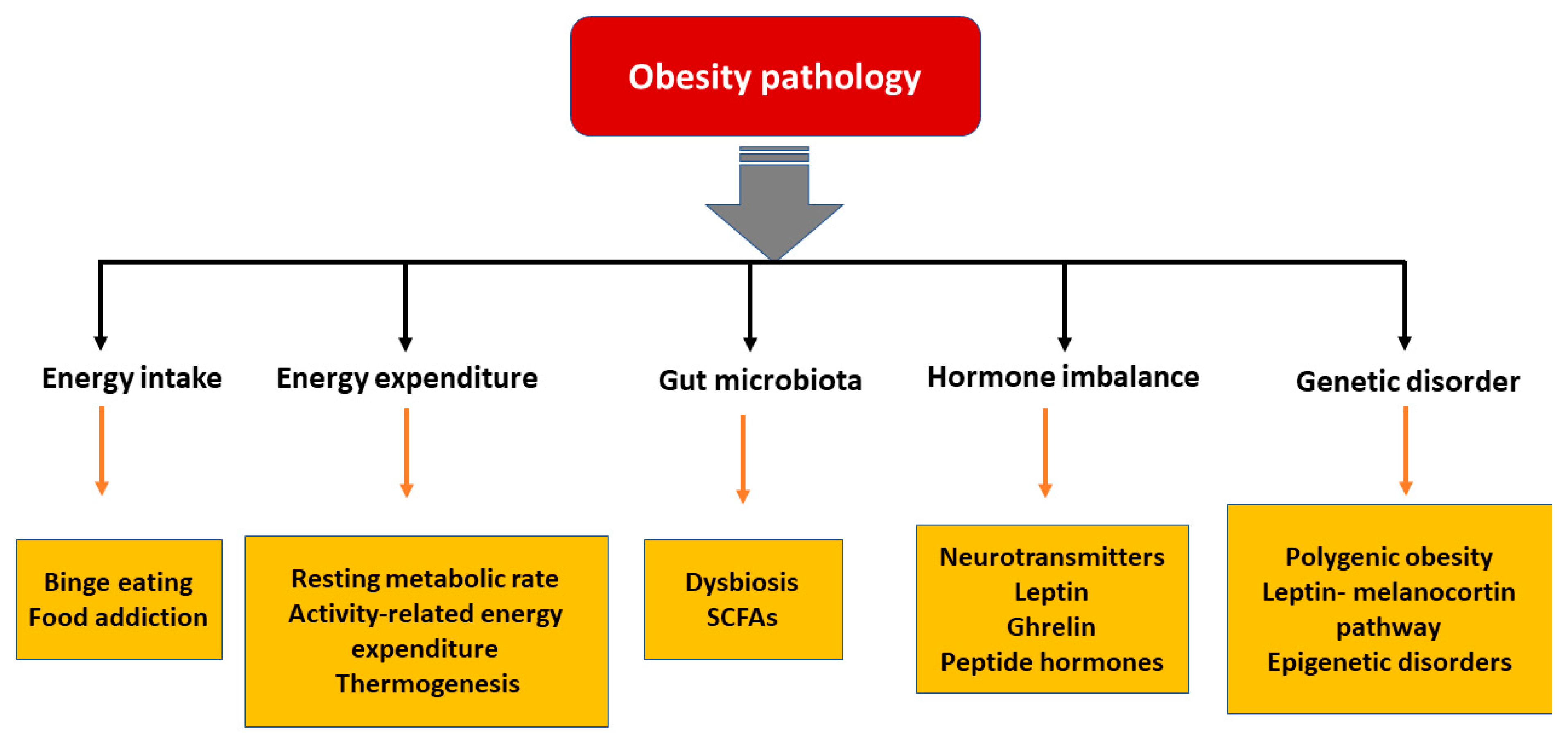{kind=link}
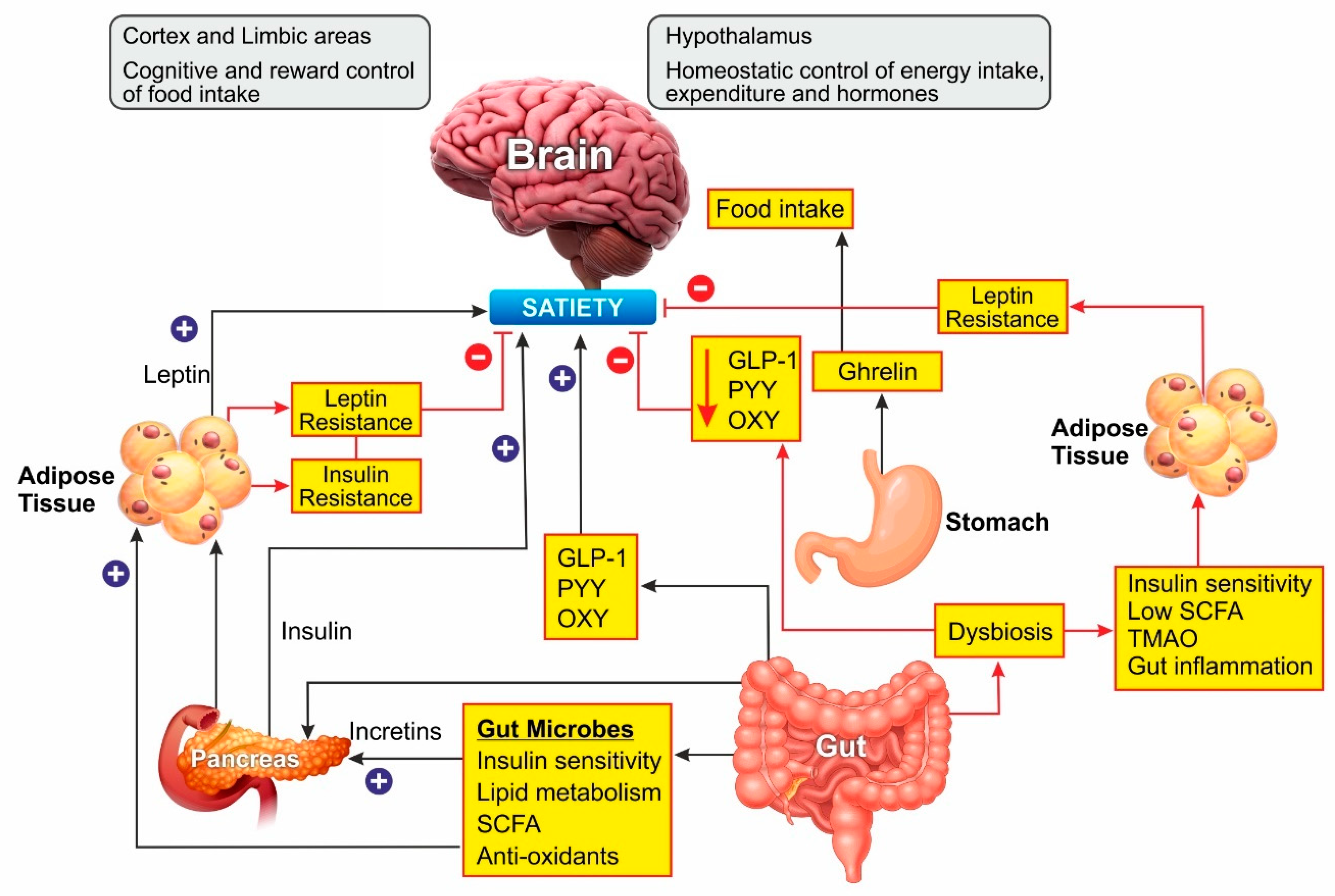{kind=link}
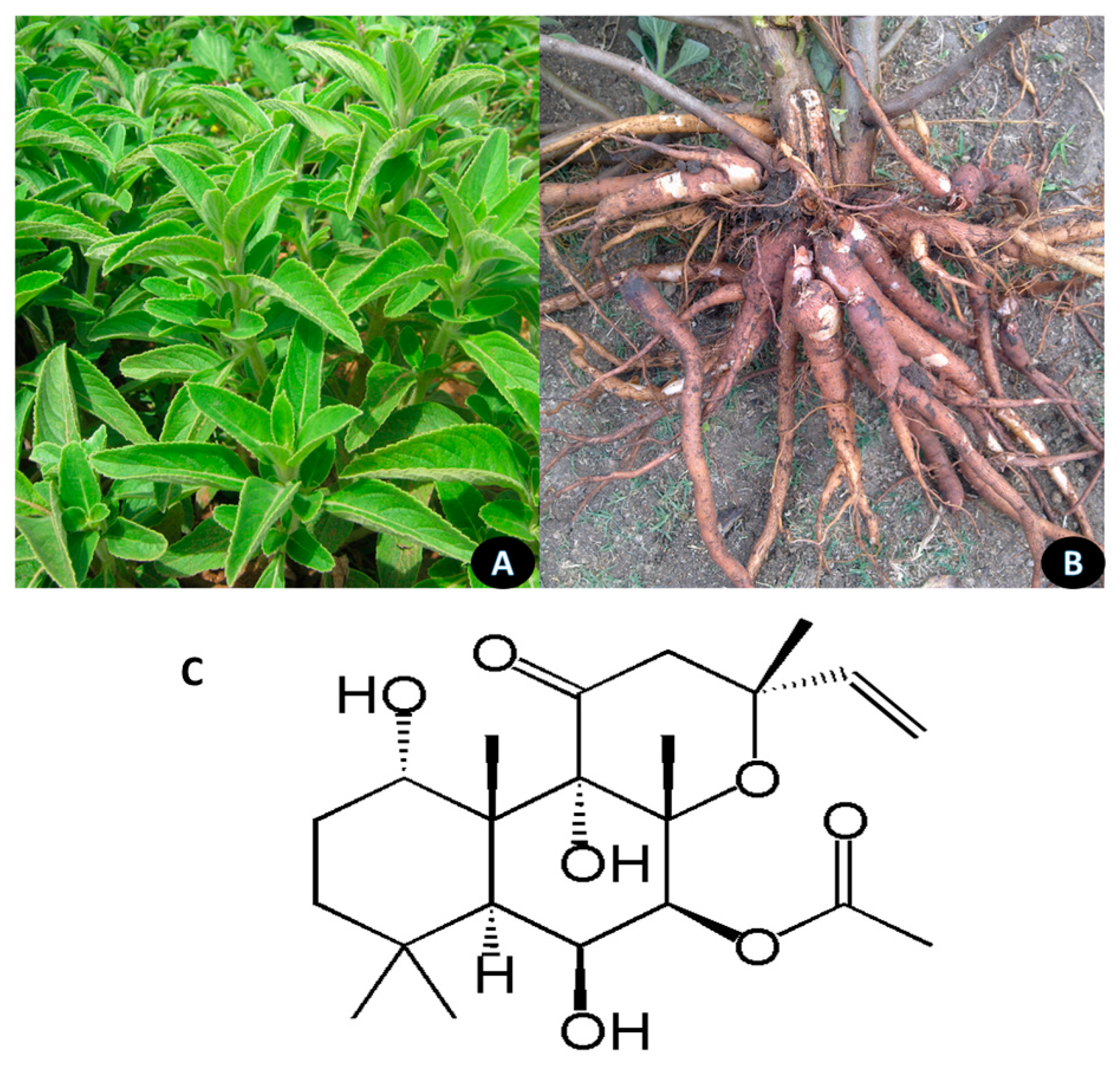{kind=link}
{kind=link}
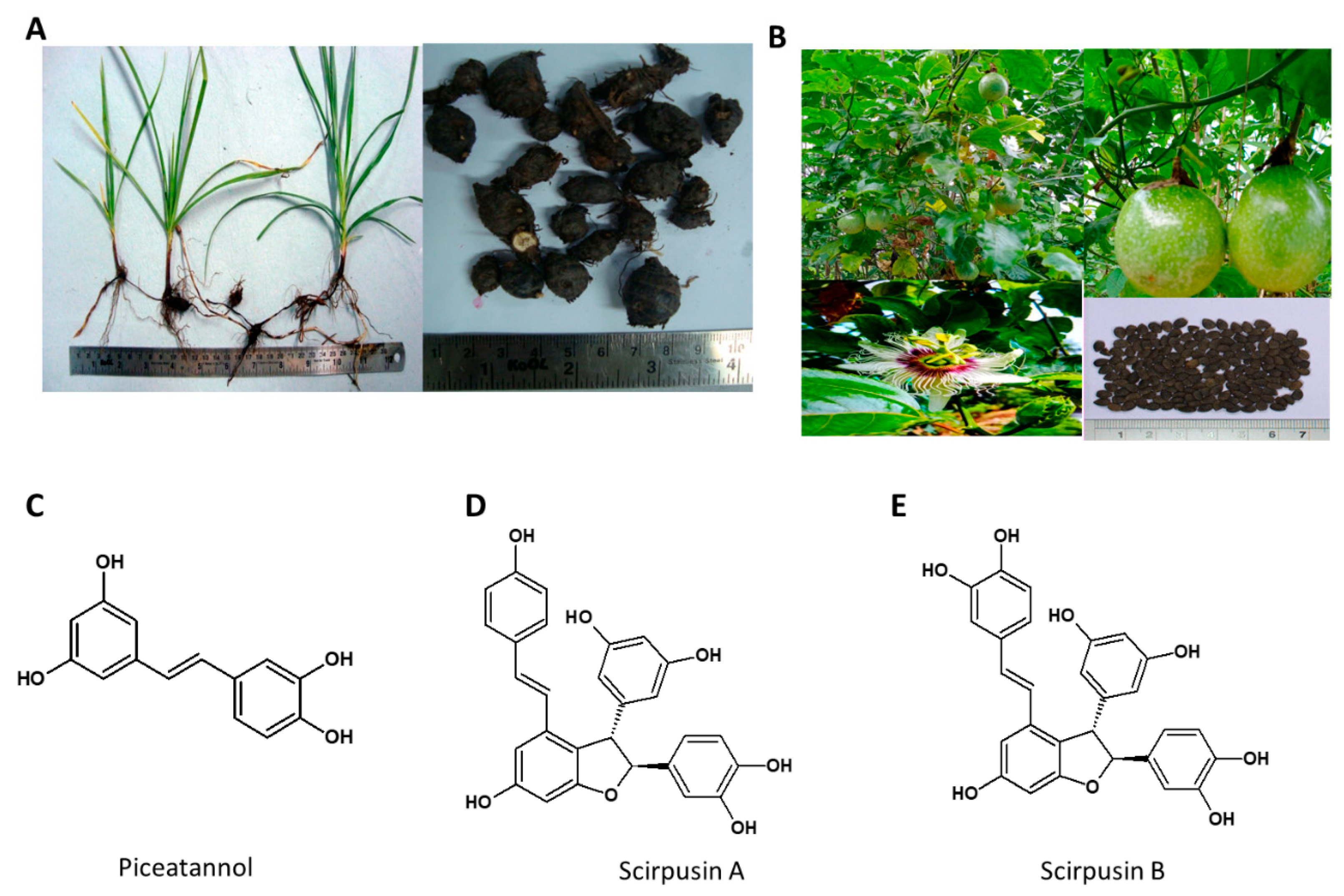{kind=link}
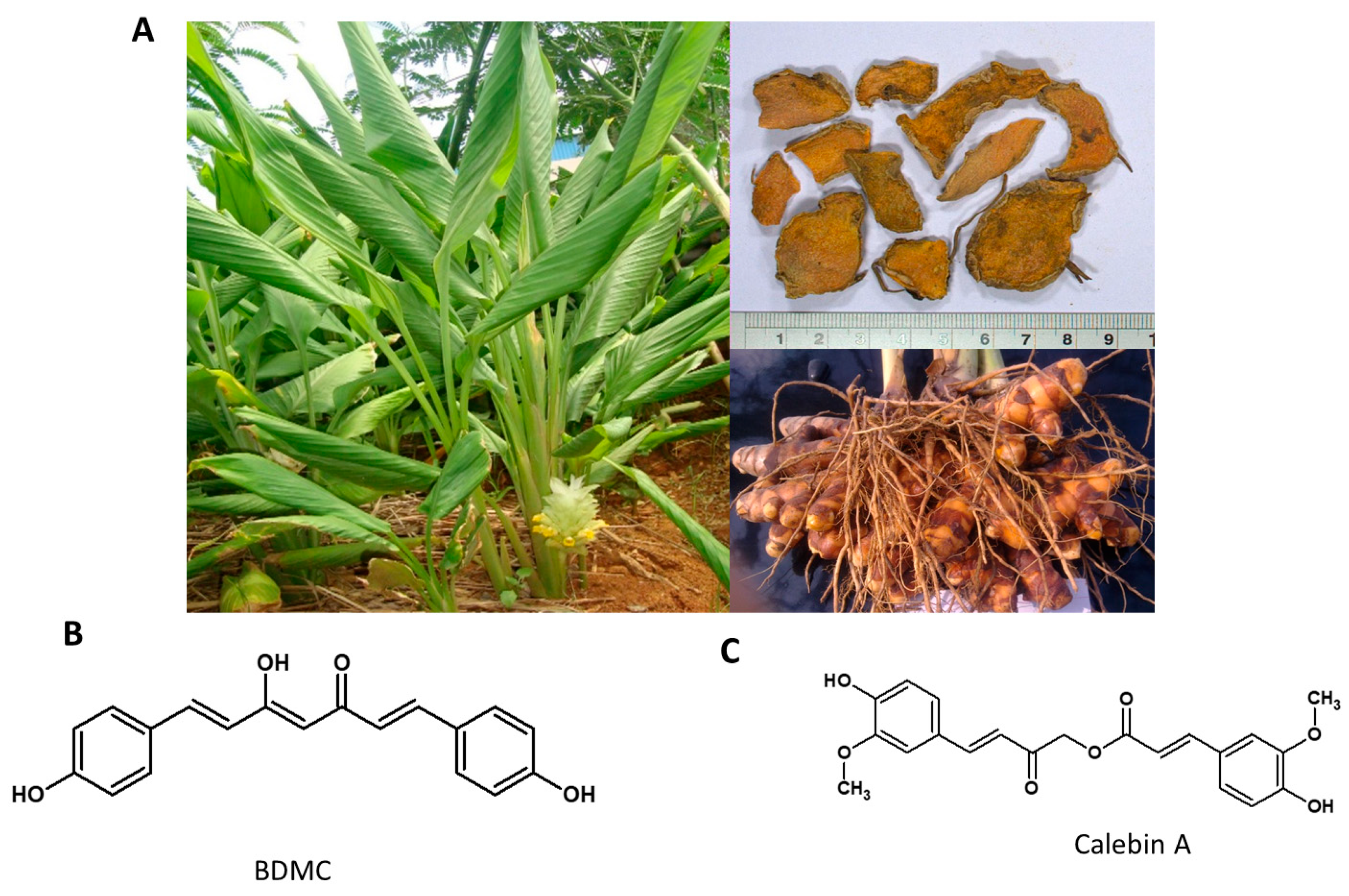{kind=link}
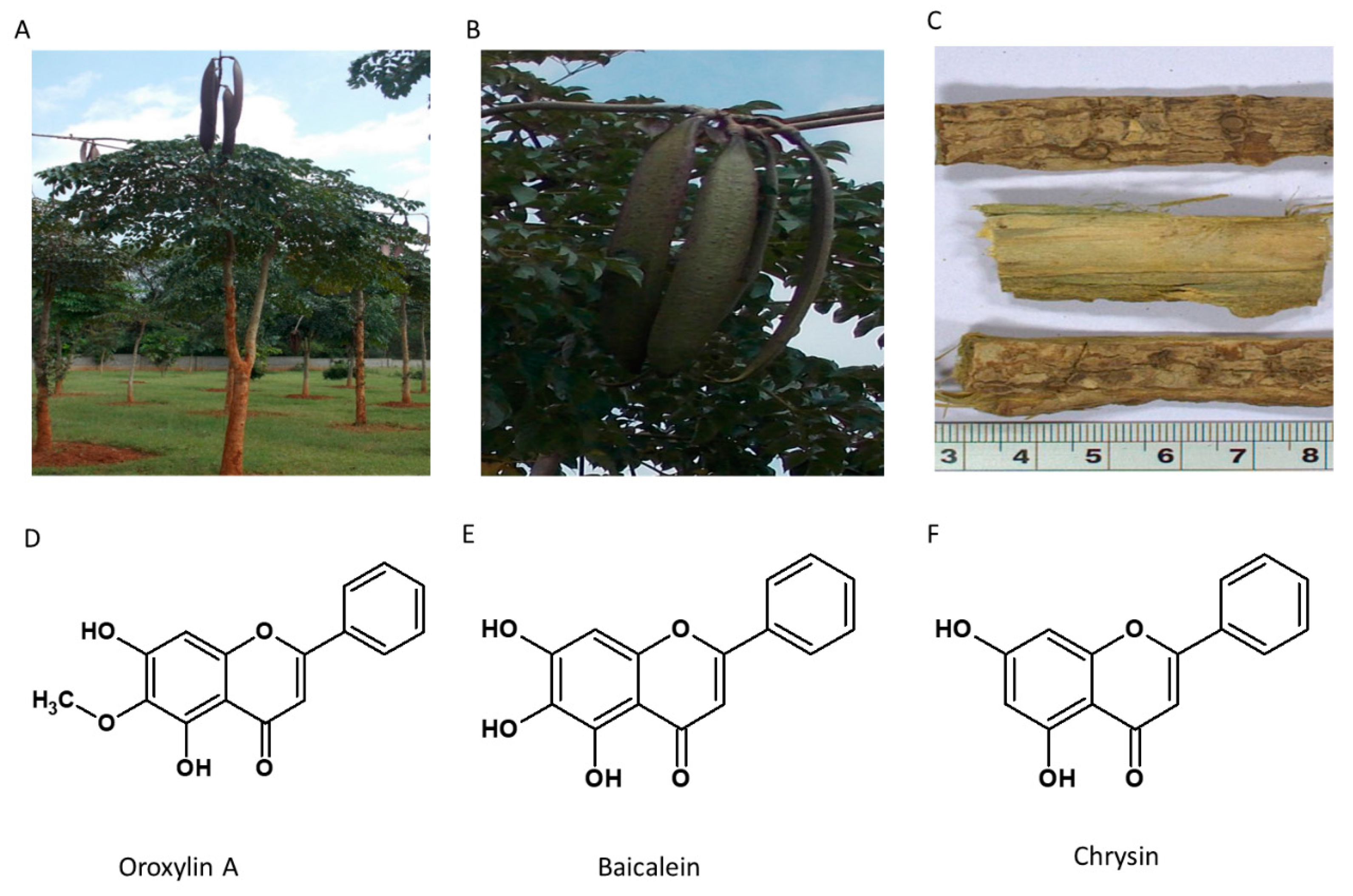{kind=link}
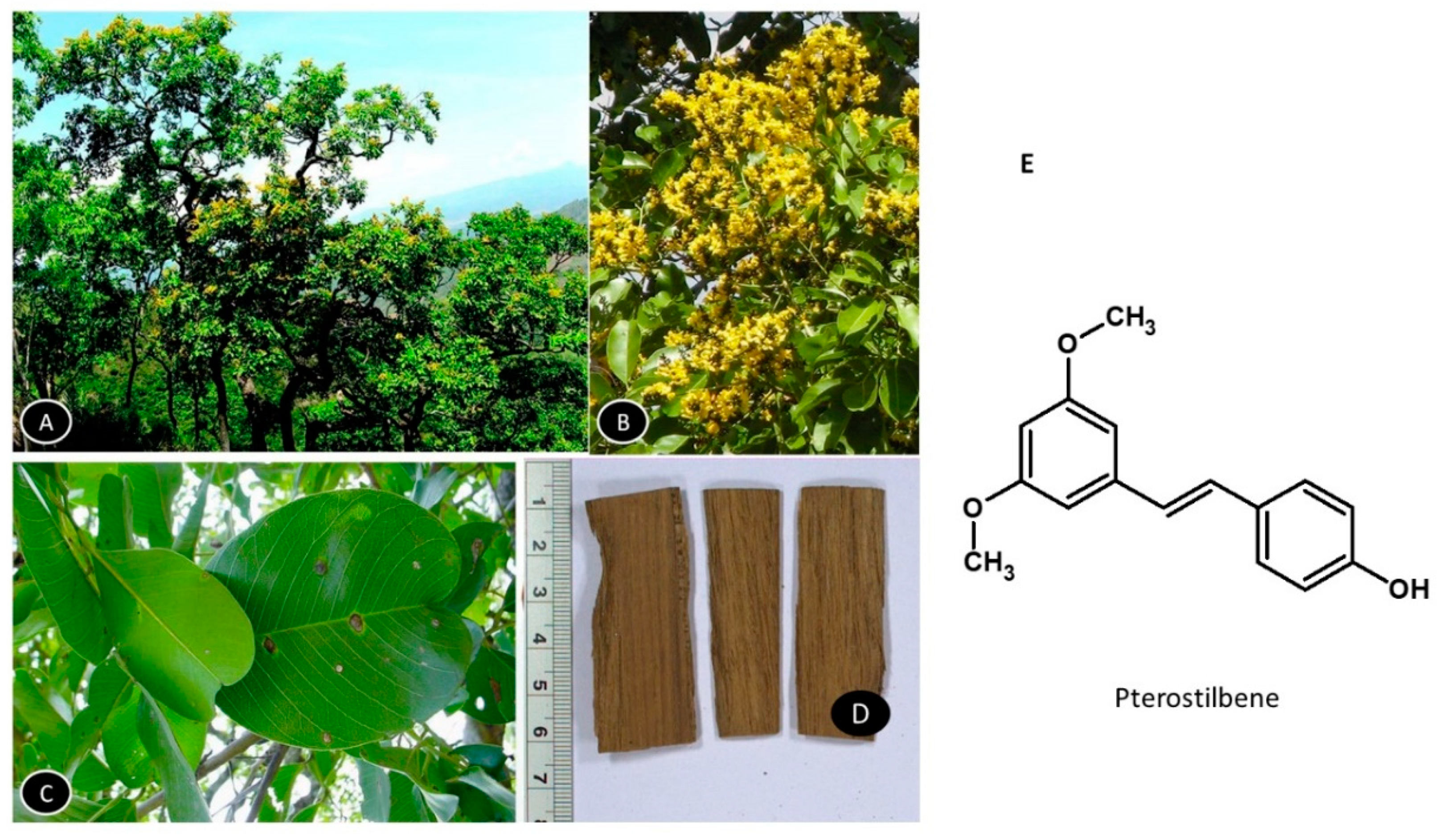{kind=link}
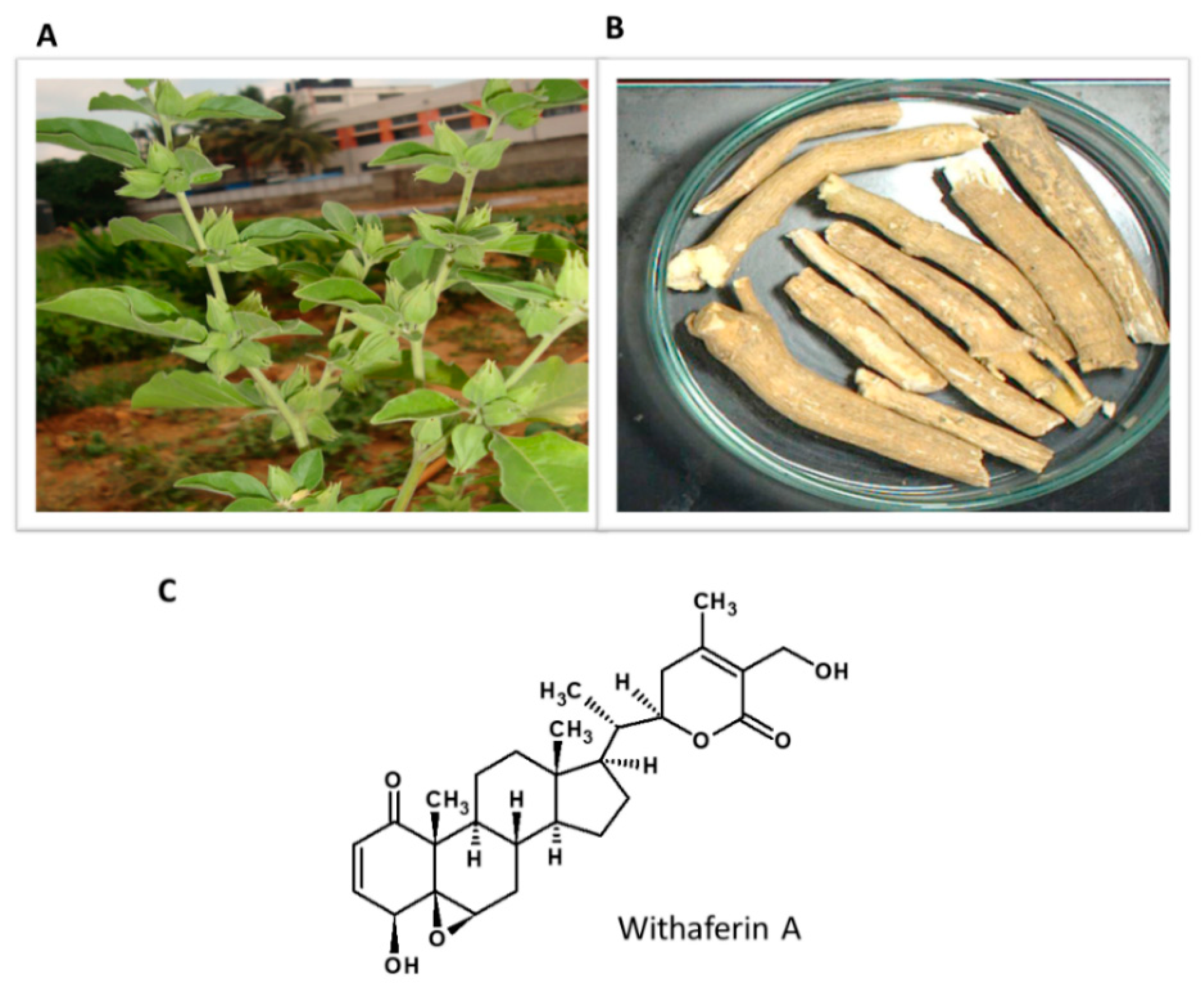{kind=link}
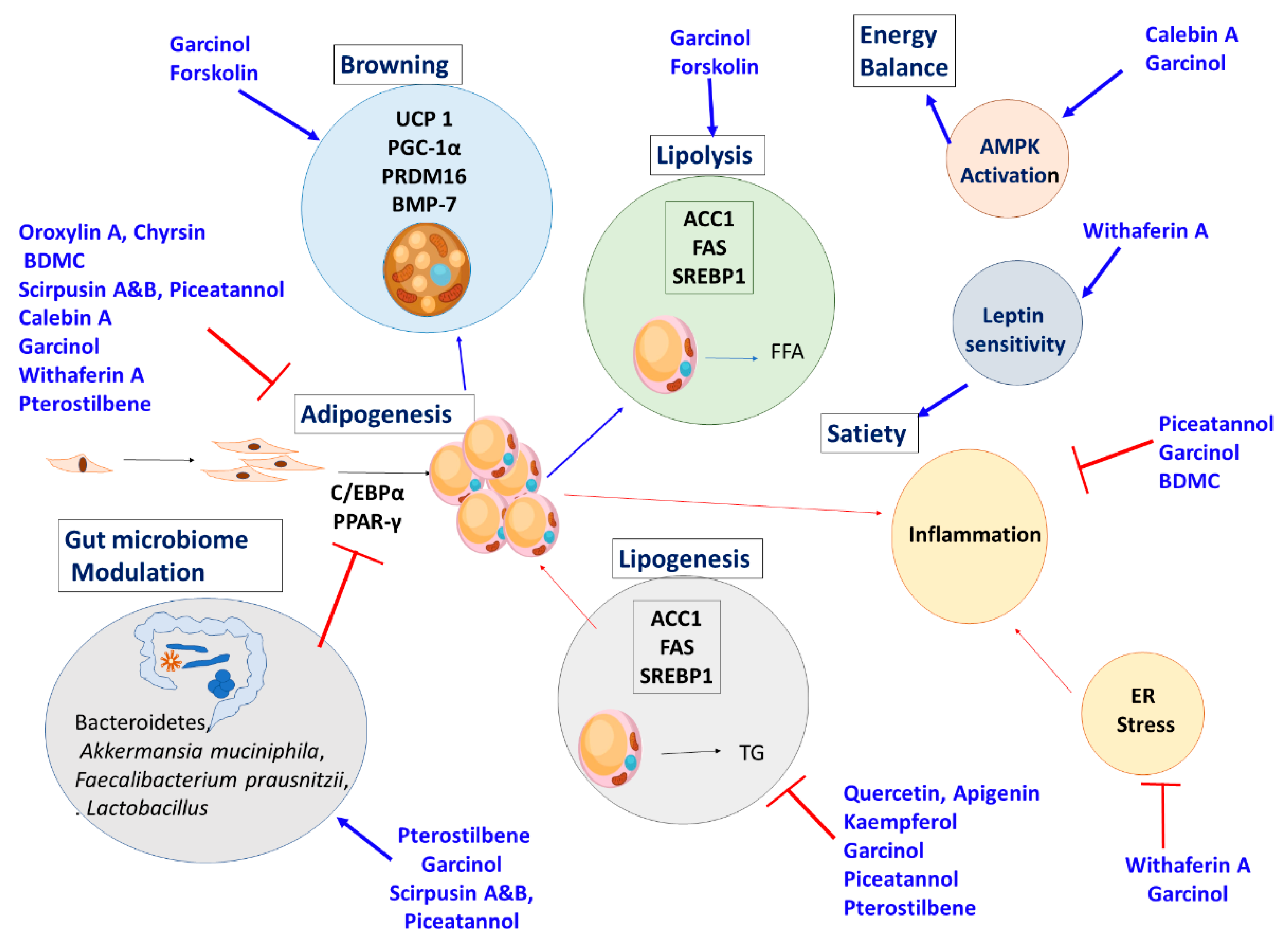{kind=link}
| Drug Name | Mechanism of Action | Contraindications and Side Effects | Year Approved and Present Status |
|---|---|---|---|
| Phentermine | Centrally acting sympathomimetic agent, appetite suppressant | Increased blood pressure and heart rate. | 1959 Approved for short term |
| Fenfluramine | Increasing serotonin levels through decreasing reuptake of serotonin | Heart valve damage and major adverse cardiovascular events | (1973–1997) Withdrawn |
| Dexfenfluramine | (1996–1997) Withdrawn | ||
| Sibutramine | (2001–2002) Withdrawn | ||
| Orlistat | Inhibiting pancreatic lipase | Hypertension, Diabetes, Hyperlipidemia | 1999 Approved |
| Rimonabant | Selective central cannabinoid (CB1) receptor antagonist | Psychiatric adverse events | (2006–2007) Withdrawn |
| Lorcaserin | Selective serotonin 2c (5-HT2c) receptor agonist | Occurrence of cancer | (2012–2020) Withdrawn |
| Phentermine-topiramate | Slowing gastric motility and suppressing appetite | Glaucoma, Hyperthyroidism | 2012 Approved |
| Liraglutide (saxenda) | GLP-1 receptor agonist control appetite by mimicking the natural hormone | Medullary thyroid cancer, multiple endocrine neoplasia type 2, C-cell hyperplasia of thyroid, decreased kidney function and pancreatitis | 2014 Approved |
| Semaglutide | Phase III |
Table 2.
Monogenic mutations linked to obesity.
| Name | Gene | Chromosomal Position | Action | Reference |
|---|---|---|---|---|
| Leptin | LEP | 7q32.1 | Secreted by adipocytes and functions as satiety signal in hypothalamus | [62] |
| Leptin receptor | LEPR | 1p31.2 | Functions as a receptor for leptin to mediate its effect | [63] |
| Proopiomelanocortin | POMC | 2p23.2 | Its deficiency results in the absence α MSH which regulates appetite | [64] |
| Melanocortin 4 receptor | MC4R | 18q21.32 | Appetite regulation, binds to α MSH | [65] |
| Single-minded Drosophila Homologue-1 | SIM1 | 6q16.3 | Transcriptional factor required for regulating appetite | [66] |
| Neurotrophic Tyrosine Kinase Receptor Type 2 and Brain Derived Neurotropic factor | NTRK2, BDNF | 9q21.33, 11p14.1 | These neuro-tropins are involved in regulation of food intake and body weight | [67] |
| SH2B adaptor protein | SH2B1 | 16p11.2 | Positive regulator of leptin sensitivity | [68] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Majeed, M.; Majeed, S.; Nagabhushanam, K.; Gnanamani, M.; Mundkur, L. Lesser Investigated Natural Ingredients for the Management of Obesity. Nutrients 2021, 13, 510. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020510
AMA Style
Majeed M, Majeed S, Nagabhushanam K, Gnanamani M, Mundkur L. Lesser Investigated Natural Ingredients for the Management of Obesity. Nutrients. 2021; 13(2):510. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020510
Chicago/Turabian StyleMajeed, Muhammed, Shaheen Majeed, Kalyanam Nagabhushanam, Muthuraman Gnanamani, and Lakshmi Mundkur. 2021. "Lesser Investigated Natural Ingredients for the Management of Obesity" Nutrients 13, no. 2: 510. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13020510
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.