
Effects of Bovine Colostrum with or without Egg on In Vitro Bacterial-Induced Intestinal Damage with Relevance for SIBO and Infectious Diarrhea

Abstract
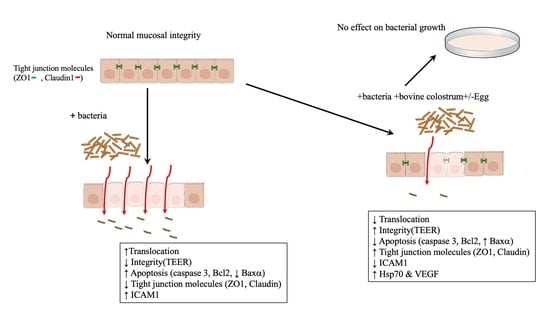
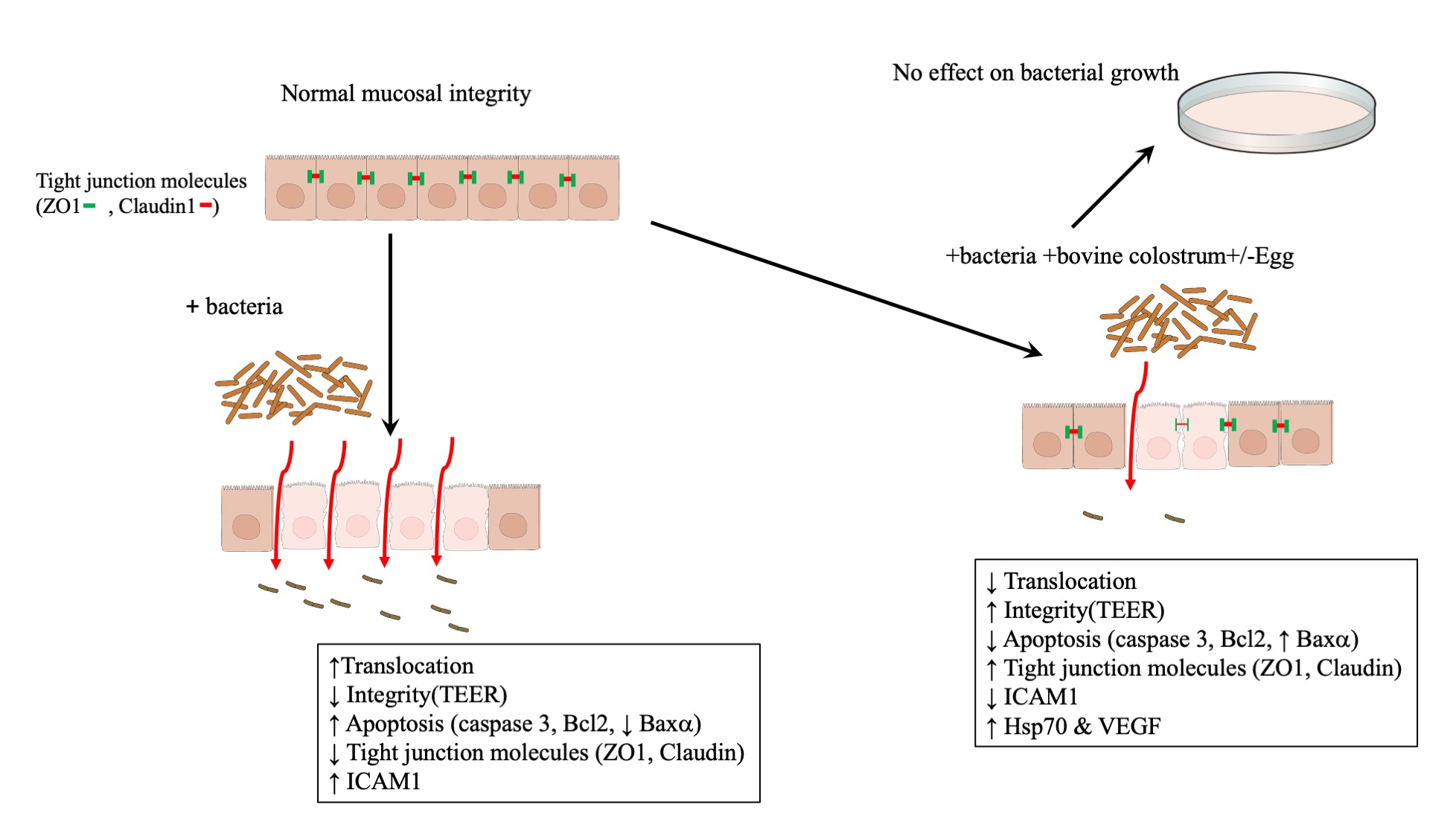
:
1. Introduction
2. Materials and Methods
2.1. Bovine Colostrum (BC) and Egg Samples
2.2. Cell Line
2.3. Bacterial Strains and Culture
2.4. Study Series 1: Effect of BC, Egg, or the Combination on Bacterial Growth
2.5. Study Series 2: Effect of BC, Egg, or the Combination on Transepithelial Passage of Bacteria and Transepithelial Electrical Resistance (TEER)
2.6. Study Series 3: Mechanisms of Action of BC, Egg, or the Combination for Maintaining Epithelial Integrity
2.6.1. Cell Lysate Preparation
2.6.2. Tight Junction Proteins
2.6.3. Cell Apoptosis Assays
2.6.4. ICAM-1, VEGF, and Hsp70
2.7. Statistical Analyses
3. Results
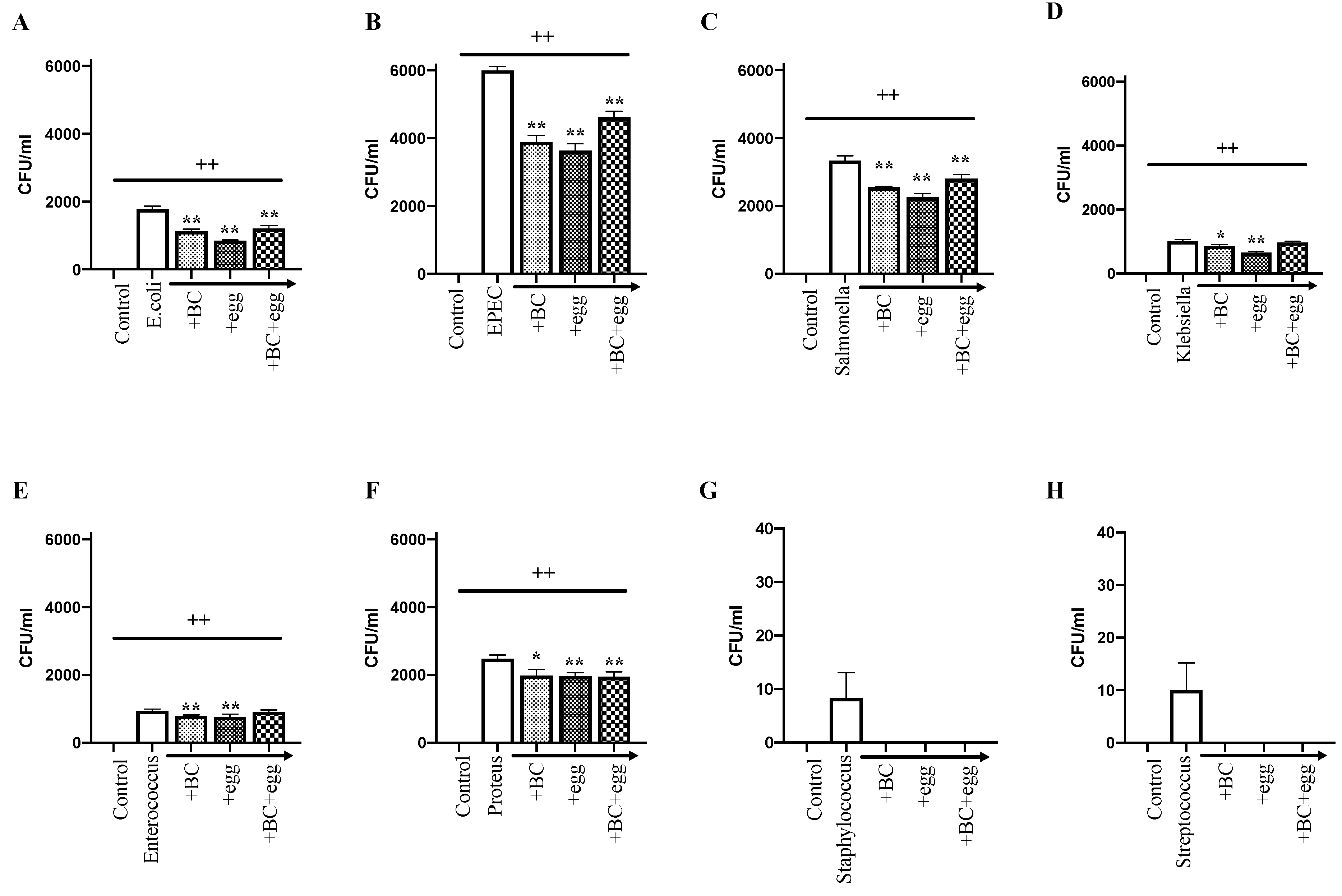
3.1. Study Series 1: Effect of BC, Egg or the Combination on Bacterial Growth
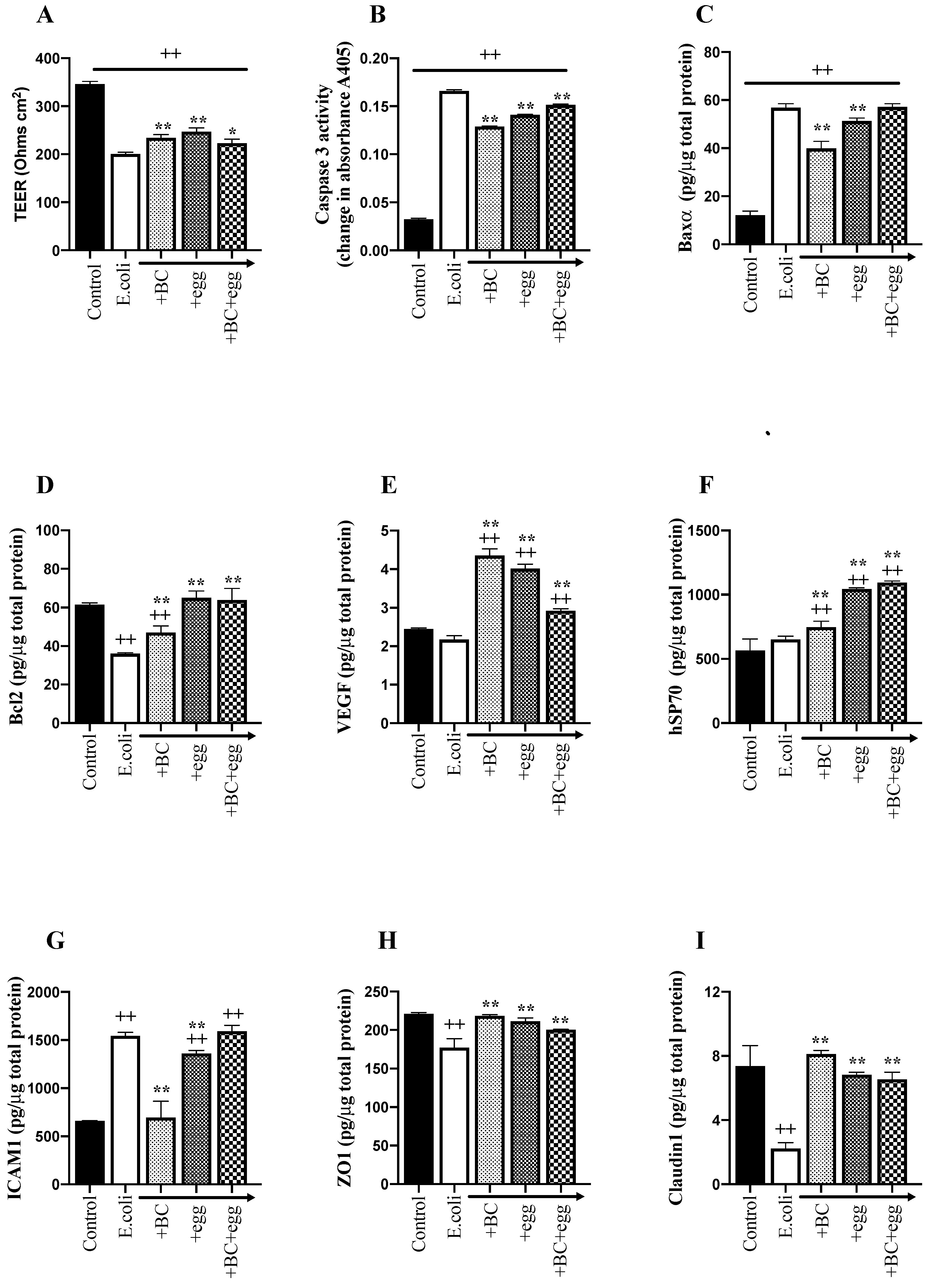
3.2. Study Series 2: Effect of BC, Egg, or the Combination on Transepithelial Passage of Bacteria and TEER
3.2.1. Bacterial Translocation
3.2.2. TEER
3.3. Study Series 3: Mechanisms of Action of BC, Egg, or the Combination for Maintaining Epithelial Integrity
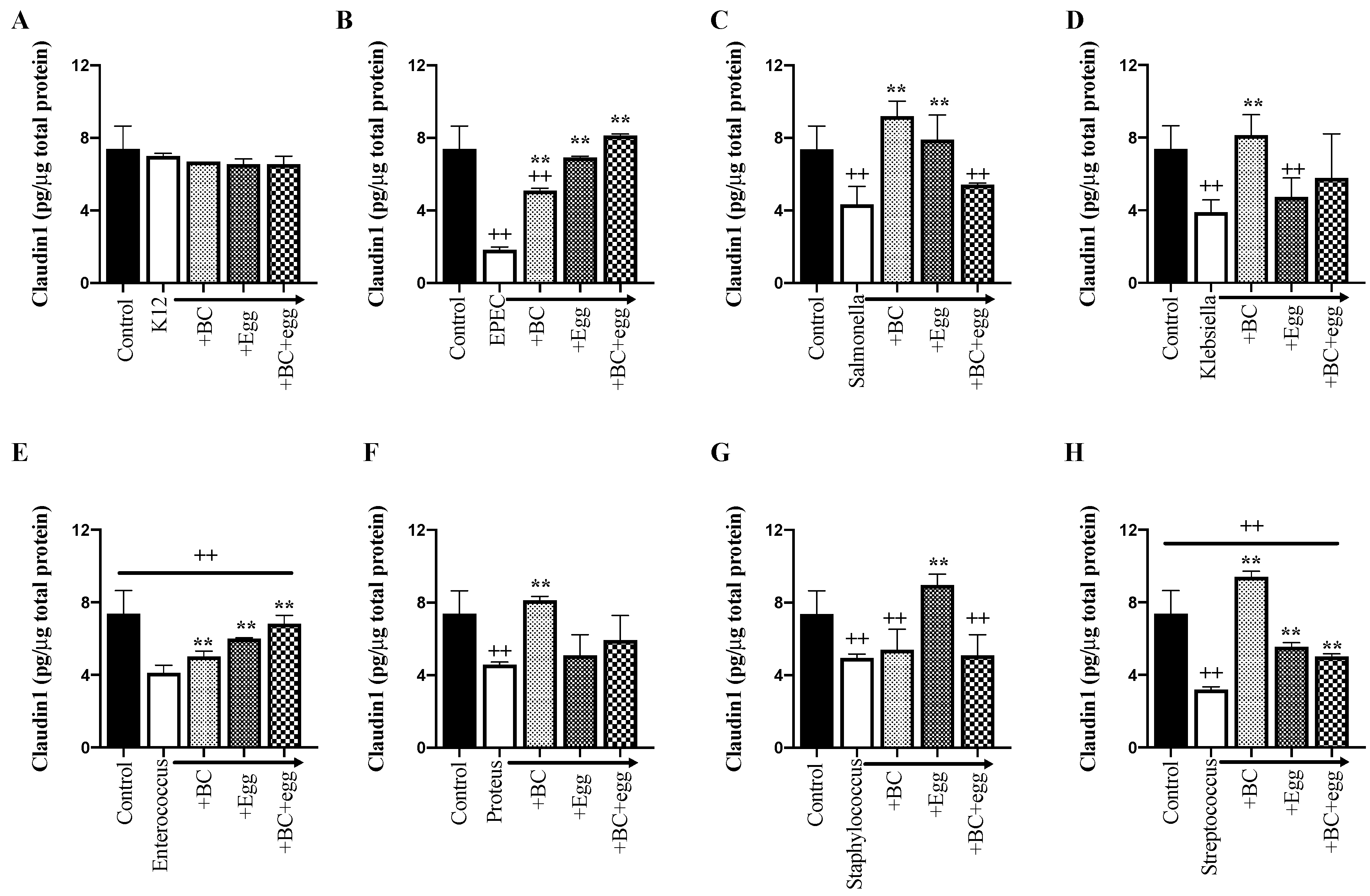
3.3.1. Tight Junction Proteins
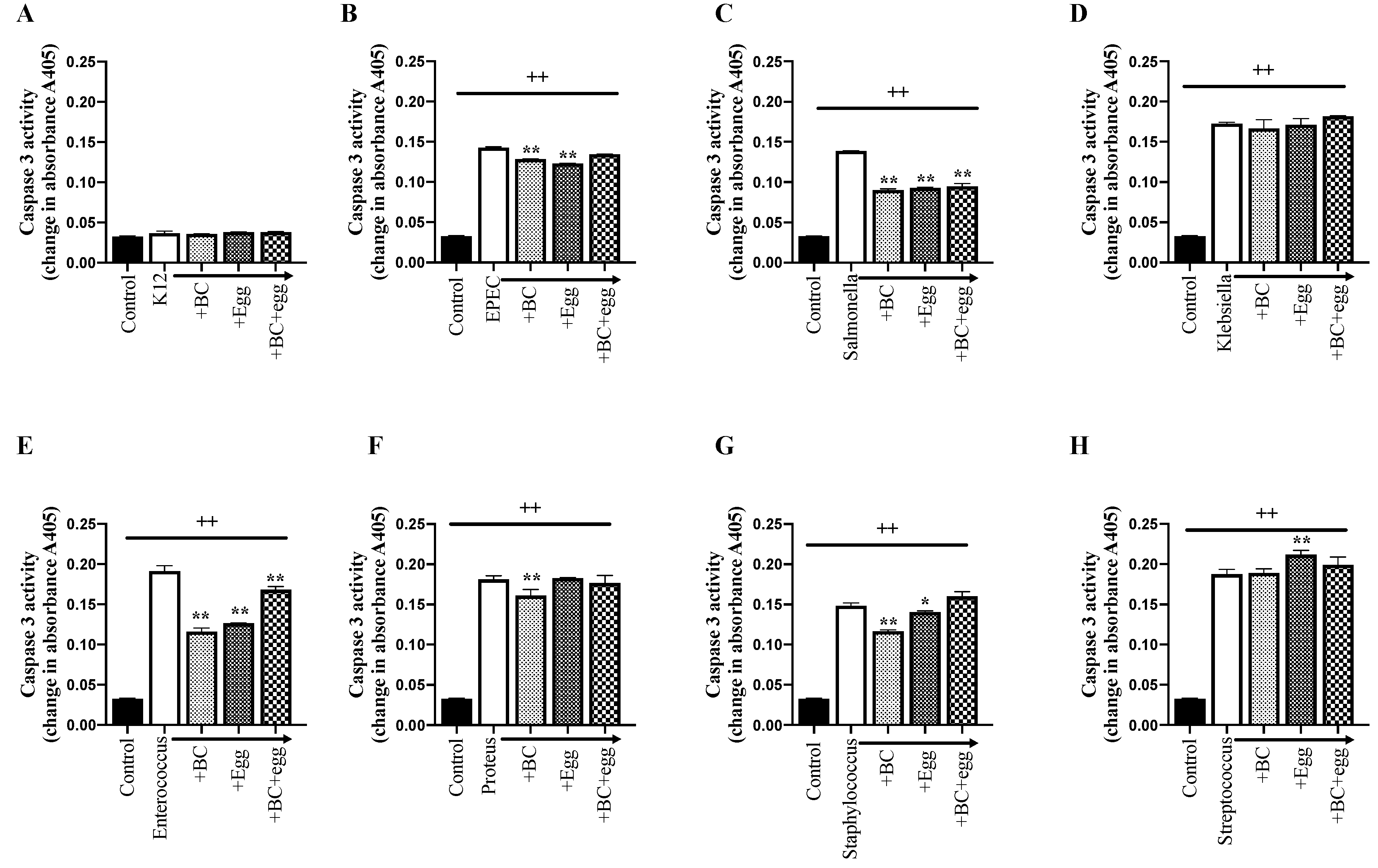
3.3.2. Cell Apoptosis Assays
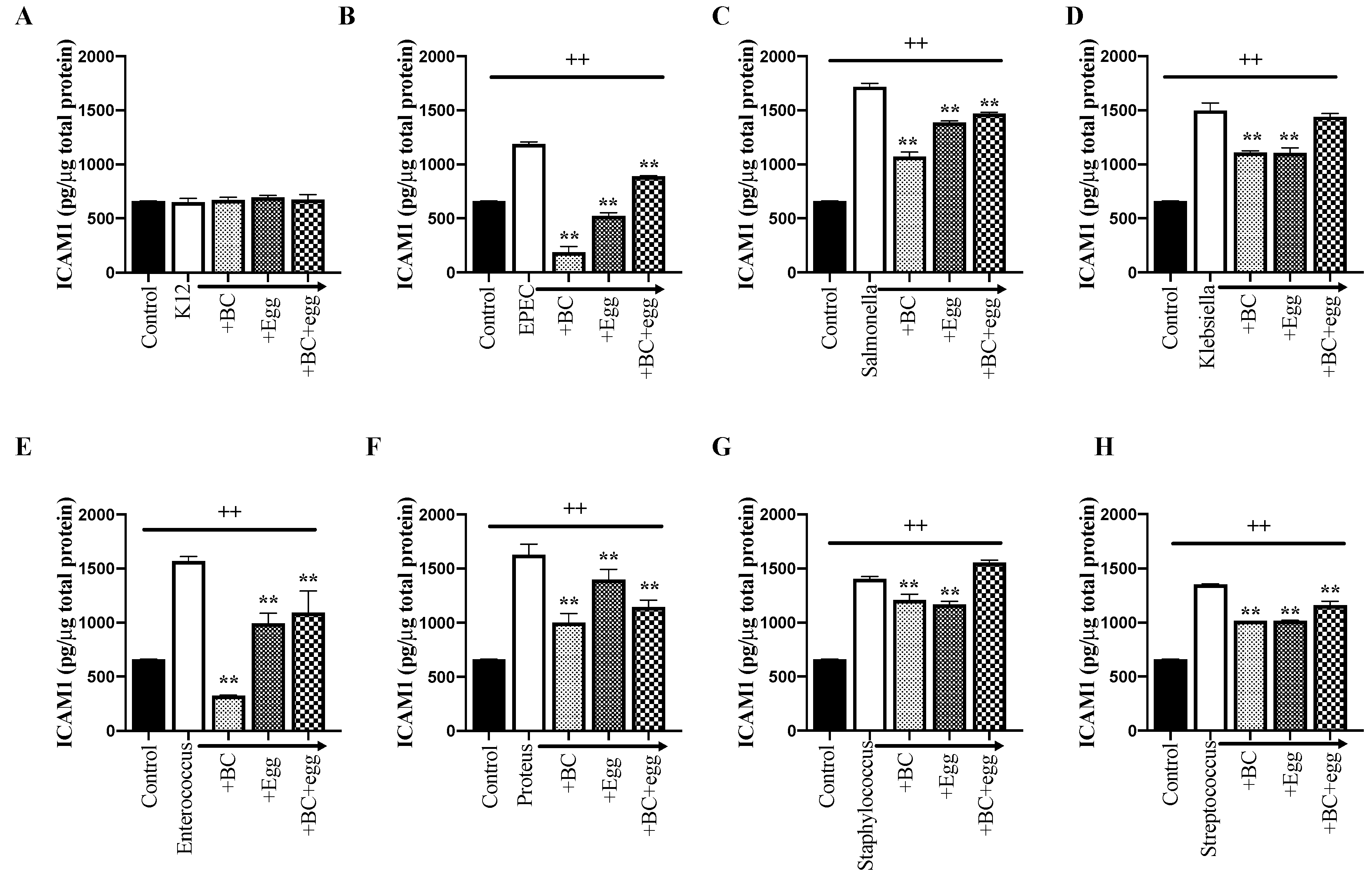
3.3.3. ICAM-1, VEGF, and Hsp70
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| BC | Bovine colostrum |
| EGF | Epidermal growth factor |
| EPEC | Enteropathogenic Escherichia coli |
| IGF | Insulin-like growth factor |
| SIBO | Small intestinal bacterial overgrowth |
| TEER | Transepithelial electrical resistance |
| TGF | Transforming growth factor |
References
- Middleton, S.J.; Playford, R.J. Bacterial overgrowth of the small intestine. In Oxford Textbook of Medicine, 6th ed.; Oxford University Press: Oxford, UK, 2020; pp. 2879–2883. [Google Scholar]
- Ghoshal, U.C.; Shukla, R.; Ghoshal, U. Small Intestinal Bacterial Overgrowth and Irritable Bowel Syndrome: A Bridge between Functional Organic Dichotomy. Gut Liver 2017, 11, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Dukowicz, A.C.; Lacy, B.E.; Levine, G.M. Small intestinal bacterial overgrowth: A comprehensive review. Gastroenterol. Hepatol. 2007, 3, 112–122. [Google Scholar]
- Pimentel, M.; Chow, E.J.; Lin, H.C. Eradication of small intestinal bacterial overgrowth reduces symptoms of irritable bowel syndrome. Am. J. Gastroenterol. 2000, 95, 3503–3506. [Google Scholar] [CrossRef]
- Mello, C.S.; Rodrigues, M.S.D.C.; Filho, H.B.D.A.; Melli, L.C.F.L.; Tahan, S.; Pignatari, A.C.C.; De Morais, M.B. Fecal microbiota analysis of children with small intestinal bacterial overgrowth among residents of an urban slum in Brazil. J. Pediatr. 2018, 94, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, C.; Adame, E.C.; Attaluri, A.; Valestin, J.; Rao, S.S.C. Dysmotility and proton pump inhibitor use are independent risk factors for small intestinal bacterial and/or fungal overgrowth. Aliment. Pharmacol. Ther. 2013, 37, 1103–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haboubi, N.Y.; Lee, G.S.; Montgomery, R.D. Duodenal Mucosal Morphometry of Elderly Patients with Small Intestinal Bacterial Overgrowth: Response to Antibiotic Treatment. Age Ageing 1991, 20, 29–32. [Google Scholar] [CrossRef]
- Gatta, L.; Scarpignato, C. Systematic review with meta-analysis: Rifaximin is effective and safe for the treatment of small intestine bacterial overgrowth. Aliment. Pharmacol. Ther. 2017, 45, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, E.C.; Gabrielli, M.; Scarpellini, E.; Lupascu, A.; Novi, M.; Sottili, S.; Vitale, G.; Cesario, V.; Serricchio, M.; Cammarota, G.; et al. Small Intestinal Bacterial Overgrowth Recurrence After Antibiotic Therapy. Am. J. Gastroenterol. 2008, 103, 2031–2035. [Google Scholar] [CrossRef]
- Pimentel, M.; Saad, R.J.; Long, M.D.; Rao, S.S.C. ACG Clinical Guideline: Small Intestinal Bacterial Overgrowth. Am. J. Gastroenterol. 2020, 115, 165–178. [Google Scholar] [CrossRef]
- Panahi, Y.; Falahi, G.; Falahpour, M.; Moharamzad, Y.; Khorasgani, M.R.; Beiraghdar, F.; Naghizadeh, M.M. Bovine Colostrum in the Management of Nonorganic Failure to Thrive: A Randomized Clinical Trial. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 551–554. [Google Scholar] [CrossRef] [PubMed]
- Playford, R.J.; Macdonald, C.E.; Calnan, D.P.; Floyd, D.N.; Podas, T.; Johnson, W.; Wicks, A.C.; Bashir, O.; Marchbank, T. Co-administration of the health food supplement, bovine colostrum, reduces the acute non-steroidal anti-inflammatory drug-induced increase in intestinal permeability. Clin. Sci. 2001, 100, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Buckley, J.D.; Brinkworth, G.D.; Abbott, M.J. Effect of bovine colostrum on anaerobic exercise performance and plasma insu-lin-like growth factor I. J. Sports Sci. 2003, 21, 577–588. [Google Scholar] [CrossRef]
- Réhault-Godbert, S.; Guyot, N.; Nys, Y. The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health. Nutrients 2019, 11, 684. [Google Scholar] [CrossRef] [Green Version]
- Kovacs-Nolan, J.; Phillips, M.; Mine, Y. Advances in the Value of Eggs and Egg Components for Human Health. J. Agric. Food Chem. 2005, 53, 8421–8431. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, L.; Tan, W.; Zhao, W. Clinical efficacy of oral immunoglobulin Y in infant rotavirus enteritis: Systematic re-view and meta-analysis. Medicine 2019, 98, e16100. [Google Scholar] [CrossRef]
- Playford, R.J.; Garbowsky, M.; Marchbank, T. Pasteurized Chicken Egg Powder Stimulates Proliferation and Migration of AGS, RIE1, and Caco-2 Cells and Reduces NSAID-Induced Injury in Mice and Colitis in Rats. J. Nutr. 2020, 150, 1434–1442. [Google Scholar] [CrossRef]
- Gaensbauer, J.T.; Melgar, M.A.; Calvimontes, D.M.; Lamb, M.M.; Asturias, E.J.; Contreras-Roldan, I.L.; Dominguez, S.R.; Rob-inson, C.C.; Berman, S. Efficacy of a bovine colostrum and egg-based intervention in acute childhood diarrhoea in Guatemala: A randomised, double-blind, placebo-controlled trial. BMJ Glob. Health 2017, 2, e000452. [Google Scholar] [CrossRef] [Green Version]
- Playford, R.J.; Cattell, M.; Marchbank, T. Marked variability in bioactivity between commercially available bovine colostrum for human use; implications for clinical trials. PLoS ONE 2020, 15, e0234719, Erratum in 2020, 15, e0240392. [Google Scholar]
- Lee, J.; Paik, H.-D. Anticancer and immunomodulatory activity of egg proteins and peptides: A review. Poult. Sci. 2019, 98, 6505–6516. [Google Scholar] [CrossRef]
- Fogh, J.; Fogh, J.M.; Orfeo, T. One Hundred and Twenty-Seven Cultured Human Tumor Cell Lines Producing Tumors in Nude Mice23. J. Natl. Cancer Inst. 1977, 59, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhry, N.; Scott, F.; Edgar, M.; Sanger, G.J.; Kelly, P. Reversal of Pathogen-Induced Barrier Defects in Intestinal Epithelial Cells by Contra-pathogenicity Agents. Dig. Dis. Sci. 2021, 66, 88–104. [Google Scholar] [CrossRef]
- Marchbank, T.; Davison, G.; Oakes, J.R.; Ghatei, M.A.; Patterson, M.; Moyer, M.P.; Playford, R.J. The nutriceutical bovine colostrum truncates the increase in gut permeability caused by heavy exercise in athletes. Am. J. Physiol. Liver Physiol. 2011, 300, G477–G484. [Google Scholar] [CrossRef] [Green Version]
- Playford, R.; Weiser, M. Bovine Colostrum: Its Constituents and Uses. Nutrients 2021, 13, 265. [Google Scholar] [CrossRef]
- Ebina, T.; Ohta, M.; Kanamaru, Y.; Yamamoto-Osumi, Y.; Baba, K. Passive immunizations of suckling mice and infants with bovine colostrum containing antibodies to human rotavirus. J. Med. Virol. 1992, 38, 117–123. [Google Scholar] [CrossRef]
- Rump, J.; Arndt, R.; Arnold, A.; Bendick, C.; Dichtelmüller, H.; Franke, M.; Helm, E.; Jäger, H.; Kampmann, B.; Kolb, P.; et al. Treatment of diarrhoea in human immunodeficiency virus-infected patients with immunoglobulins from bovine colostrum. J. Mol. Med. 1992, 70, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Brooks, H.J.; McConnell, M.A.; Corbett, J.; Buchan, G.S.; Fitzpatrick, C.E.; Broadbent, R.S. Potential prophylactic value of bovine colostrum in necrotizing enterocolitis in neonates: An in vitro study on bacterial attachment, antibody levels and cytokine production. FEMS Immunol. Med. Microbiol. 2006, 48, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lissner, R.; Thürmann, P.A.; Merz, G.; Karch, H. Antibody reactivity and fecal recovery of bovine immunoglobulins follow-ing oral administration of a colostrum concentrate from cows (Lactobin) to healthy volunteers. Int. J. Clin. Pharmacol. Ther. 1998, 36, 239–245. [Google Scholar] [PubMed]
- Mahdavi, A.; Rahmani, H.; Nili, N.; Samie, A.; Soleimania, S. Chicken Egg Yolk Antibody (IgY) Powder Against Escherichia coli O78:K80. J. Anim. Veter Adv. 2010, 9, 366–373. [Google Scholar] [CrossRef] [Green Version]
- Rokutan, K. Role of heat shock proteins in gastric mucosal protection. J. Gastroenterol. Hepatol. 2000, 15, 12–19. [Google Scholar] [CrossRef]
- Duffy, A.M.; Bouchier-Hayes, D.J.; Harmey, J.H. Vascular Endothelial Growth Factor (VEGF) and Its Role in Non-Endothelial Cells: Autocrine Signalling by VEGF. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2004; pp. 133–144. [Google Scholar]
- Hsu, C.-R.; Chang, I.-W.; Hsieh, P.-F.; Lin, T.-L.; Liu, P.-Y.; Huang, C.-H.; Li, K.-T.; Wang, J.-T. A Novel Role for the Klebsiella pneumoniae Sap (Sensitivity to Antimicrobial Peptides) Transporter in Intestinal Cell Interactions, Innate Immune Responses, Liver Abscess, and Virulence. J. Infect. Dis. 2019, 219, 1294–1306. [Google Scholar] [CrossRef] [PubMed]
- Kwak, Y.-K.; Vikström, E.; Magnusson, K.-E.; Vécsey-Semjén, B.; Colque-Navarro, P.; Möllby, R. The Staphylococcus aureus Alpha-Toxin Perturbs the Barrier Function in Caco-2 Epithelial Cell Monolayers by Altering Junctional Integrity. Infect. Immun. 2012, 80, 1670–1680. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Liu, H.; Yang, S.; Li, Z.; Zhong, J.; Fang, R. Epidermal Growth Factor and Intestinal Barrier Function. Mediat. Inflamm. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEntee, C.P.; Gunaltay, S.; Travis, M.A. Regulation of barrier immunity and homeostasis by integrin-mediated transforming growth factor β activation. Immunology 2019, 160, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, T.; Shinoda, I.; Fukuwatari, Y.; Shimamura, S. Effects of Lactoferrin and Its Peptides on Proliferation of Rat Intestinal Epithelial Cell Line, IEC-18, in the Presence of Epidermal Growth Factor. Biosci. Biotechnol. Biochem. 1995, 59, 1875–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otani, K.; Tanigawa, T.; Watanabe, T.; Shimada, S.; Nadatani, Y.; Nagami, Y.; Tanaka, F.; Kamata, N.; Yamagami, H.; Shiba, M.; et al. Microbiota Plays a Key Role in Non-Steroidal Anti-Inflammatory Drug-Induced Small Intestinal Damage. Digestion 2017, 95, 22–28. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| +Bacteria | Bacteria + BC | Bacteria + Egg | Bacteria + BC + Egg | |
|---|---|---|---|---|
| TEER (Ohm/cm2) Baseline without bacteria control value = 346 ± 5.9 | ||||
| E. coli K12 (-ve control) | 336 ± 10 | 353 ± 7 | 374 ± 12 | 356 ± 11 |
| EPEC | 167 ± 6 ++ | 201 ± 3 ++ ** | 220 ± 2 ++ ** | 195 ± 5 ++ ** |
| Salmonella | 173 ± 2 ++ | 197 ± 6 ++ ** | 229.5 ± 4 ++ ** | 186 ± 1 ++ ** |
| Klebsiella | 236 ± 10 ++ | 230 ± 6 ++ | 259 ± 1 ++ ** | 216 ± 9 ++ |
| Enterococcus | 219 ± 3 ++ | 219 ± 5 ++ | 231 ± 2 ++ ** | 220 ± 2 ++ |
| Proteus | 213 ± 8 ++ | 227 ± 2 ++ | 229.8 ± 1 ++ | 223 ± 1 ++ |
| Staphylococcus | 281 ± 5 ++ | 306 ± 14 ++ | 325 ± 6.8 ** | 291 ± 3 ++ |
| Streptococcus | 279 ± 6 ++ | 326 ± 16 * | 323 ± 19 * | 290 ± 3 ++ |
| ZO1 (pg/μg protein) Baseline without bacteria control value = 207 ± 5 | ||||
| E. coli K12 | 206 ± 3.5 | 205 ± 3 | 206 ± 1 | 204 ± 1 |
| EPEC | 156 ± 6 ++ | 205 ± 1 ++ ** | 201 ± 2 ++ ** | 183 ± 3 ++ ** |
| Salmonella | 177 ± 8 ++ | 205 ± 2 ++ ** | 201 ± 2 ++ ** | 171 ± 27 ++ |
| Klebsiella | 151 ± 1 ++ | 175 ± 0.5 ++ ** | 163 ± 4 ++ ** | 153 ± 2 ++ |
| Enterococcus | 71 ± 21.5 ++ | 190 ± 2 ++ ** | 183 ± 0.1 ++ ** | 175 ± 3 ++ ** |
| Proteus | 95 ± 1 ++ | 146 ± 1 ++ ** | 132 ± 5 ++ ** | 133 ± 4 ++ ** |
| Staphylococcus | 161 ± 2 ++ | 162 ± 1 ++ | 171 ± 1 ++ | 155 ± 30 ++ |
| Streptococcus | 173 ± 1 ++ | 156 ± 5 ++ | 175 ± 1 ++ | 165 ± 28 ++ |
| +Bacteria | Bacteria + BC | Bacteria + Egg | Bacteria + BC + Egg | |
|---|---|---|---|---|
| Baxα (pg/μg protein) Baseline without bacteria control value = 12.17 ± 1.67 | ||||
| E. coli K12 | 14.2 ± 1.67 | 12.8 ± 2 | 14.67 ± 1.5 | 12.3 ± 1.5 |
| EPEC | 49.8 ± 0.33 ++ | 36.7 ± 1.17 ++ ** | 36.5 ± 0.33 ++ ** | 44.3 ± 1.16 ++ ** |
| Salmonella | 44.3 ± 1.17 ++ | 36.5 ± 1 ++ ** | 36.5 ± 1 ++ ** | 44 ± 0.5 ++ |
| Klebsiella | 63.3 ± 1.17 ++ | 61.2 ± 1.33 ++ | 63.5 ± 1.76 ++ | 63.8 ± 3 ++ |
| Enterococcus | 67 ± 0.83 ++ | 37.7 ± 0.83 ++ ** | 39.3 ± 2.5 ++ ** | 46.2 ± 2.3 ++ ** |
| Proteus | 66 ± 0.17 ++ | 58.8 ± 0.33 ++ ** | 59.7 ± 0.5 ++ ** | 64.7 ± 0.8 ++ |
| Staphylococcus | 55.3 ± 0.83 ++ | 39.2 ± 1 ++ ** | 45.8 ± 2.7 ++ ** | 53.3 ± 3.5 ++ |
| Streptococcus | 66.8 ± 0.33 ++ | 64.5 ± 3.3 ++ | 73.6 ± 2.7 ++ | 69.3 ± 1.17 ++ |
| Bcl2 (pg/μg protein) Baseline without bacteria control value = 61.5 ± 0.87 | ||||
| E. coli K12 | 59.5 ± 0.3 | 61.5 ± 0.3 | 54.2 ± 1.7 | 56 ± 5.2 |
| EPEC | 45 ± 4 ++ | 61.5 ± 0.9 ** | 69.5 ± 1.4 ++ ** | 53 ± 2.9 ++ ** |
| Salmonella | 38 ± 4.6 ++ | 59.5 ± 0.9 ** | 57 ± 3.5 ** | 54 ± 5.2 ** |
| Klebsiella | 66 ± 3.5 | 55 ± 2.6 | 61.5 ± 2 | 57 ± 0.6 |
| Enterococcus | 37.5 ± 0.9 ++ | 36 ± 0.6 ++ | 38.5 ± 3.2 ++ | 35 ± 3.5 ++ |
| Proteus | 31.5 ± 0.3 ++ | 61.5 ± 0.3 ** | 53 ± 4 ** | 67 ± 3.5 ** |
| Staphylococcus | 54 ± 7.5 | 62 ± 5.8 | 72.5 ± 3.7 ++ * | 79.5 ± 6.6 ++ ** |
| Streptococcus | 63 ± 3.5 | 51.5 ± 2.6 | 66.5 ± 0.9 ++ ** | 81.8 ± 2.2 ++ ** |
| +Bacteria | Bacteria + BC | Bacteria + Egg | Bacteria + BC + Egg | |
|---|---|---|---|---|
| VEGF (pg/μg protein) Baseline without bacteria control value = 2.45 ± 0.03 | ||||
| E. coli K12 | 2.67 ± 0.55 | 2.57 ± 0.004 | 2.56 ± 0.01 | 2.79 ± 0.24 |
| EPEC | 2.52 ± 0.05 | 3.72 ± 0.03 ++ ** | 3.49 ± 0.01 ++ ** | 3.41 ± 0.04 ++ ** |
| Salmonella | 4.4 ± 0.03 ++ | 5.56 ± 0.01 ++ ** | 5.2 ± 0.02 ++ ** | 4.5 ± 0.07 ++ ** |
| Klebsiella | 2.31 ± 0.36 | 2.57 ± 0.3 | 2.62 ± 0.27 | 3.59 ± 0.26 ++ ** |
| Enterococcus | 5.11 ± 0.14 ++ | 5.48 ± 0.15 ++ ** | 5 ± 0.2 ++ | 4.8 ± 0.01 ++ |
| Proteus | 3.16 ± 0.01 ++ | 3.69 ± 0.14 ++ ** | 3.21 ± 0.19 ++ | 3.15 ± 0.01 ++ |
| Staphylococcus | 4.51 ± 0.06 ++ | 4.59 ± 0.11 ++ | 4.53 ± 0.04 ++ | 4.29 ± 0.29 ++ |
| Streptococcus | 4.96 ± 0.04 ++ | 4.58 ± 0.12 ++ | 4.46 ± 0.01 ++ | 4.75 ± 0.18 ++ |
| Hsp70 (pg/μg protein) Baseline without bacteria control value = 566 ± 90 | ||||
| E. coli K12 | 675 ± 57 | 682 ± 30 | 694 ± 72 | 677 ± 65 |
| EPEC | 492 ± 60 | 867 ± 87 ++ ** | 792 ± 40 ++ ** | 782 ± 4 ++ ** |
| Salmonella | 642 ± 40 | 966 ± 12 ++ ** | 1091 ± 23 ++ ** | 923 ± 119 ++ ** |
| Klebsiella | 1396 ± 10 ++ | 1167 ± 25 ++ | 1422 ± 28 ++ | 1234 ± 2 ++ |
| Enterococcus | 1195 ± 129 ++ | 1589 ± 15 ++ ** | 1599 ± 67 ++ ** | 1425 ± 5 ++ ** |
| Proteus | 860 ± 28 ++ | 1083 ± 27 ++ ** | 1135 ± 11 ++ ** | 963 ± 21 ++ ** |
| Staphylococcus | 1172 ± 82 ++ | 1165 ± 185 ++ | 1031 ± 101 ++ | 1185 ± 147 ++ |
| Streptococcus | 1280 ± 86 ++ | 1540 ± 10 ++ ** | 1333 ± 97 ++ | 1157 ± 235 ++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Playford, R.J.; Choudhry, N.; Kelly, P.; Marchbank, T. Effects of Bovine Colostrum with or without Egg on In Vitro Bacterial-Induced Intestinal Damage with Relevance for SIBO and Infectious Diarrhea. Nutrients 2021, 13, 1024. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13031024
Playford RJ, Choudhry N, Kelly P, Marchbank T. Effects of Bovine Colostrum with or without Egg on In Vitro Bacterial-Induced Intestinal Damage with Relevance for SIBO and Infectious Diarrhea. Nutrients. 2021; 13(3):1024. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13031024
Chicago/Turabian StylePlayford, Raymond J., Naheed Choudhry, Paul Kelly, and Tania Marchbank. 2021. "Effects of Bovine Colostrum with or without Egg on In Vitro Bacterial-Induced Intestinal Damage with Relevance for SIBO and Infectious Diarrhea" Nutrients 13, no. 3: 1024. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13031024