
Can Cranberry Juice Protect against Rotenone-Induced Toxicity in Rats?


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
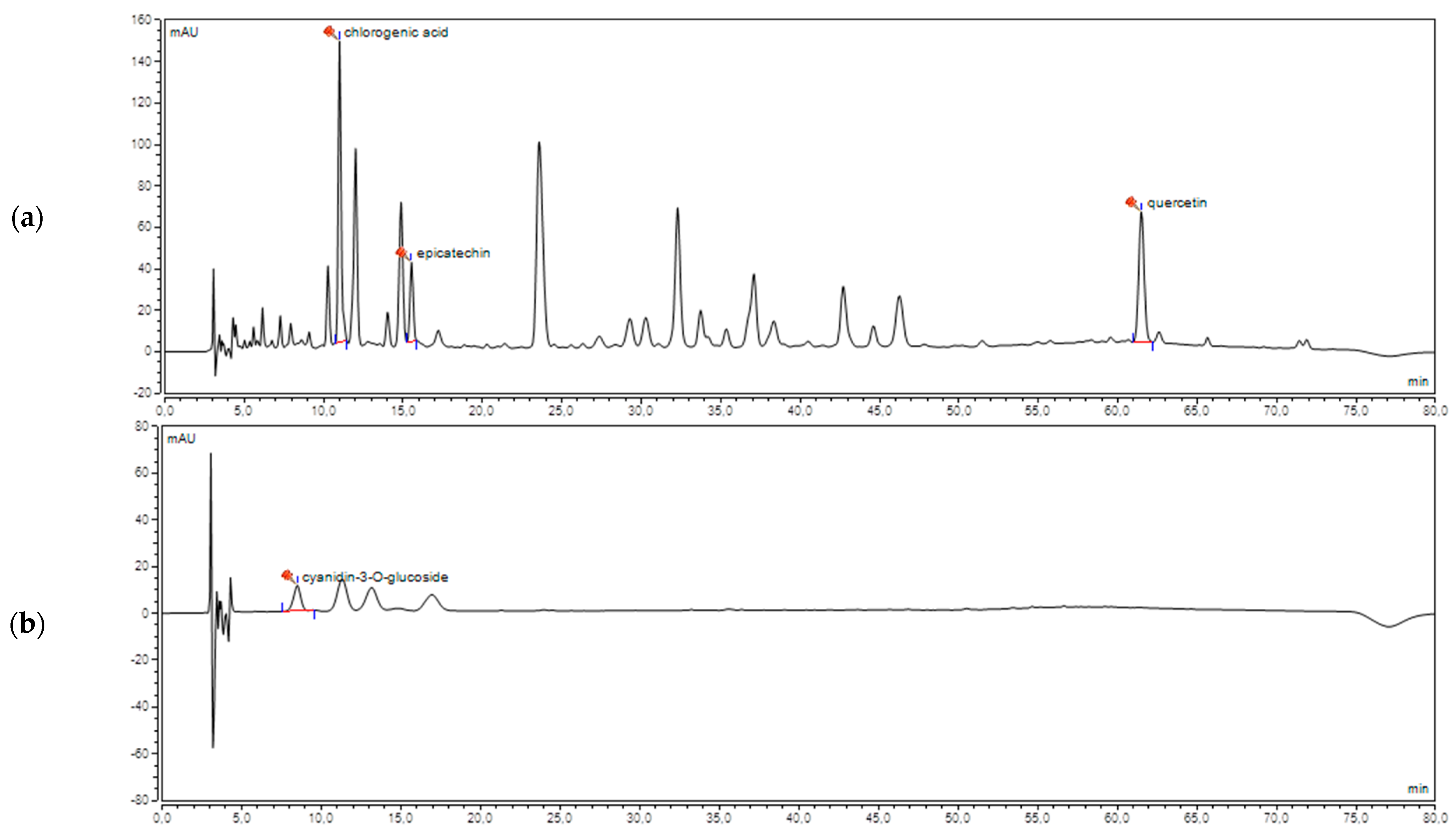
2.1. HPLC Analysis of Cranberry Juice (CJ)
2.2. Animals
2.3. Experimental Design
2.4. Biochemical Examinations
2.4.1. Sample Collection
2.4.2. Oxidative Stress Markers
2.4.3. Mitochondrial Aldehyde Dehydrogenase (ALDH2) Activity
2.4.4. Protein Determination
2.4.5. Liver Function Markers
3. Results
3.1. Determination of Polyphenols in Cranberry Juice (CJ)
3.2. Biochemical Measurements
3.2.1. Effects of Cranberry Juice on the Oxidative Stress Parameters in the Brain of Rats Treated with Rotenone
3.2.2. Effects of Cranberry Juice on the Oxidative Stress Parameters in the Liver of Rats Treated with Rotenone
3.2.3. Effects of Cranberry Juice on the Oxidative Stress Parameters in the Kidneys of Rats Treated with Rotenone
3.2.4. Effects of Cranberry Juice on the Oxidative Stress Parameters in the Serum of Rats Treated with Rotenone
4. Discussion
4.1. Brain
4.2. Liver
4.3. Kidneys
4.4. Serum
5. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jurikova, T.; Skrovankova, S.; Mlcek, J.; Balla, S.; Snopek, L. Bioactive Compounds, Antioxidant Activity, and Biological Effects of European Cranberry. Molecules 2018, 24, 24. [Google Scholar] [CrossRef] [Green Version]
- Blumberg, J.B.; Camesano, T.A.; Cassidy, A.; Kris-Etherton, P.; Howell, A.; Manach, C.; Ostertag, L.M.; Sies, H.; Skulas-Ray, A.; Vita, J.A. Cranberries and their bioactive constituents in human health. Adv. Nutr. 2013, 4, 618–632. [Google Scholar] [CrossRef] [Green Version]
- Palikova, I.; Vostalova, J.; Zdarilova, A.; Svobodova, A.; Kosina, P.; Vecera, R.; Stejskal, D.; Proskova, J.; Hrbac, J.; Bednar, P.; et al. Long-term effects of three commercial cranberry products on the antioxidative status in rats: A pilot study. J. Agric. Food Chem. 2010, 58, 1672–1678. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, J.H.; Kwak, H.K. Antioxidant effects of cranberry powder in lipopolysaccharide treated hypercholesterolemic rats. Prev. Nutr. Food Sci. 2014, 19, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elberry, A.A.; Abdel-Naim, A.B.; Abdel-Sattar, E.A.; Nagy, A.A.; Mosli, H.A.; Mohamadin, A.M.; Ashour, O.M. Cranberry (Vaccinium macrocarpon) protects against doxorubicin-induced cardiotoxicity in rats. Food Chem. Toxicol. 2010, 48, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Hussain, F.; Malik, A.; Ayyaz, U.; Shafique, H.; Rana, Z.; Hussain, Z. Efficient hepatoprotective activity of cranberry extract against CCl. Asian Pac. J. Trop. Med. 2017, 10, 1054–1058. [Google Scholar] [CrossRef] [PubMed]
- Faheem, S.A.; Saeed, N.M.; El-Naga, R.N.; Ayoub, I.M.; Azab, S.S. Hepatoprotective Effect of Cranberry Nutraceutical Extract in Non-alcoholic Fatty Liver Model in Rats: Impact on Insulin Resistance and Nrf-2 Expression. Front. Pharmacol. 2020, 11, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boušová, I.; Bártíková, H.; Matoušková, P.; Lněničková, K.; Zappe, L.; Valentová, K.; Szotáková, B.; Martin, J.; Skálová, L. Cranberry extract-enriched diets increase NAD(P)H:quinone oxidoreductase and catalase activities in obese but not in nonobese mice. Nutr. Res. 2015, 35, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Chew, B.; Mathison, B.; Kimble, L.; McKay, D.; Kaspar, K.; Khoo, C.; Chen, C.O.; Blumberg, J. Chronic consumption of a low calorie, high polyphenol cranberry beverage attenuates inflammation and improves glucoregulation and HDL cholesterol in healthy overweight humans: A randomized controlled trial. Eur. J. Nutr. 2019, 58, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Pourmasoumi, M.; Hadi, A.; Najafgholizadeh, A.; Joukar, F.; Mansour-Ghanaei, F. The effects of cranberry on cardiovascular metabolic risk factors: A systematic review and meta-analysis. Clin. Nutr. 2020, 39, 774–788. [Google Scholar] [CrossRef]
- Wang, Y.; Gallegos, J.L.; Haskell-Ramsay, C.; Lodge, J.K. Effects of chronic consumption of specific fruit (berries, citrus and cherries) on CVD risk factors: A systematic review and meta-analysis of randomised controlled trials. Eur. J. Nutr. 2020. [Google Scholar] [CrossRef]
- Ferguson, P.J.; Kurowska, E.; Freeman, D.J.; Chambers, A.F.; Koropatnick, D.J. A flavonoid fraction from cranberry extract inhibits proliferation of human tumor cell lines. J. Nutr. 2004, 134, 1529–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeram, N.P.; Adams, L.S.; Hardy, M.L.; Heber, D. Total cranberry extract versus its phytochemical constituents: Antiproliferative and synergistic effects against human tumor cell lines. J. Agric. Food Chem. 2004, 52, 2512–2517. [Google Scholar] [CrossRef] [PubMed]
- Neto, C.C.; Amoroso, J.W.; Liberty, A.M. Anticancer activities of cranberry phytochemicals: An update. Mol. Nutr. Food Res. 2008, 52 (Suppl. 1), S18–27. [Google Scholar] [CrossRef]
- Prasain, J.K.; Jones, K.; Moore, R.; Barnes, S.; Leahy, M.; Roderick, R.; Juliana, M.M.; Grubbs, C.J. Effect of cranberry juice concentrate on chemically-induced urinary bladder cancers. Oncol. Rep. 2008, 19, 1565–1570. [Google Scholar] [PubMed] [Green Version]
- Sanders, L.H.; Timothy Greenamyre, J. Oxidative damage to macromolecules in human Parkinson disease and the rotenone model. Free Radic. Biol. Med. 2013, 62, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherer, T.B.; Betarbet, R.; Testa, C.M.; Seo, B.B.; Richardson, J.R.; Kim, J.H.; Miller, G.W.; Yagi, T.; Matsuno-Yagi, A.; Greenamyre, J.T. Mechanism of toxicity in rotenone models of Parkinson’s disease. J. Neurosci. 2003, 23, 10756–10764. [Google Scholar] [CrossRef] [PubMed]
- Bové, J.; Perier, C. Neurotoxin-based models of Parkinson’s disease. Neuroscience 2012, 211, 51–76. [Google Scholar] [CrossRef] [PubMed]
- Ravenstijn, P.G.M.; Merlini, M.; Hameetman, M.; Murray, T.K.; Ward, M.A.; Lewis, H.; Ball, G.; Mottart, C.; de Ville de Goyet, C.; Lemarchand, T.; et al. The exploration of rotenone as a toxin for inducing Parkinson’s disease in rats, for application in BBB transport and PK-PD experiments. J. Pharmacol. Toxicol. Methods 2008, 57, 114–130. [Google Scholar] [CrossRef]
- Xiong, N.; Xiong, J.; Khare, G.; Chen, C.; Huang, J.; Zhao, Y.; Zhang, Z.; Qiao, X.; Feng, Y.; Reesaul, H.; et al. Edaravone guards dopamine neurons in a rotenone model for Parkinson’s disease. PLoS ONE 2011, 6, e20677. [Google Scholar] [CrossRef]
- Zhang, Z.N.; Zhang, J.S.; Xiang, J.; Yu, Z.H.; Zhang, W.; Cai, M.; Li, X.T.; Wu, T.; Li, W.W.; Cai, D.F. Subcutaneous rotenone rat model of Parkinson’s disease: Dose exploration study. Brain Res. 2017, 1655, 104–113. [Google Scholar] [CrossRef]
- Terzi, A.; Iraz, M.; Sahin, S.; Ilhan, A.; Idiz, N.; Fadillioglu, E. Protective effects of erdosteine on rotenone-induced oxidant injury in liver tissue. Toxicol. Ind. Health 2004, 20, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Salam, O.M.E.; Youness, E.R.; Ahmed, N.A.; El-Toumy, S.A.; Souleman, A.M.A.; Shaffie, N.; Abouelfadl, D.M. Bougainvillea spectabilis flowers extract protects against the rotenone-induced toxicity. Asian Pac. J. Trop. Med. 2017, 10, 478–490. [Google Scholar] [CrossRef]
- Intararuchikul, T.; Teerapattarakan, N.; Rodsiri, R.; Tantisira, M.; Wohlgemuth, G.; Fiehn, O.; Tansawat, R. Effects of centella asiatica extract on antioxidant status and liver metabolome of rotenone-treated rats using GC-MS. Biomed. Chromatogr. 2019, 33, e4395. [Google Scholar] [CrossRef] [PubMed]
- Cho, T.; Wang, X.; Uetrecht, J. Rotenone increases isoniazid toxicity but does not cause significant liver injury: Implications for the hypothesis that inhibition of the mitochondrial electron transport chain is a common mechanism of idiosyncratic drug-induced liver injury. Chem. Res. Toxicol. 2019, 32, 1423–1431. [Google Scholar] [CrossRef]
- Bavli, D.; Prill, S.; Ezra, E.; Levy, G.; Cohen, M.; Vinken, M.; Vanfleteren, J.; Jaeger, M.; Nahmias, Y. Real-time monitoring of metabolic function in liver-on-chip microdevices tracks the dynamics of mitochondrial dysfunction. Proc. Natl. Acad. Sci. USA 2016, 113, E2231–E2240. [Google Scholar] [CrossRef] [Green Version]
- Kujawska, M.; Jourdes, M.; Kurpik, M.; Szulc, M.; Szaefer, H.; Chmielarz, P.; Kreiner, G.; Krajka-Kuźniak, V.; Mikołajczak, P.; Teissedre, P.L.; et al. Neuroprotective Effects of Pomegranate Juice against Parkinson’s Disease and Presence of Ellagitannins-Derived Metabolite-Urolithin A-In the Brain. Int. J. Mol. Sci. 2019, 21, 202. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, A.; Agurell, E.; Beevers, C.; Brendler-Schwaab, S.; Burlinson, B.; Clay, P.; Collins, A.; Smith, A.; Speit, G.; Thybaud, V.; et al. Recommendations for conducting the in vivo alkaline Comet assay. 4th International Comet Assay Workshop. Mutagenesis 2003, 18, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Kujawska, M.; Ignatowicz, E.; Ewertowska, M.; Adamska, T.; Markowski, J.; Jodynis-Liebert, J. Attenuation of KBrO3-induced renal and hepatic toxicity by cloudy apple juice in rat. Phytother. Res. 2013, 27, 1214–1219. [Google Scholar] [CrossRef] [PubMed]
- Kujawska, M.; Ewertowska, M.; Adamska, T.; Ignatowicz, E.; Gramza-Michałowska, A.; Jodynis-Liebert, J. Protective effect of yellow tea extract on N-nitrosodiethylamine-induced liver carcinogenesis. Pharm. Biol. 2016, 54, 1891–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurucz, A.; Bombicz, M.; Kiss, R.; Priksz, D.; Varga, B.; Hortobágyi, T.; Trencsényi, G.; Szabó, R.; Pósa, A.; Gesztelyi, R.; et al. Heme Oxygenase-1 Activity as a Correlate to Exercise-Mediated Amelioration of Cognitive Decline and Neuropathological Alterations in an Aging Rat Model of Dementia. Biomed. Res. Int. 2018, 2018, 7212861. [Google Scholar] [CrossRef]
- Chapman, D.E.; Holbrook, D.J.; Chaney, S.G.; Hall, I.H.; Lee, K.H. In vitro inhibition of mouse hepatic mixed-function oxidase enzymes by helenalin and alantolactone. Biochem. Pharmacol. 1989, 38, 3913–3923. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Gokul, K.; Muralidhara. Oral supplements of aqueous extract of tomato seeds alleviate motor abnormality, oxidative impairments and neurotoxicity induced by rotenone in mice: Relevance to Parkinson’s disease. Neurochem. Res. 2014, 39, 1382–1394. [Google Scholar] [CrossRef] [PubMed]
- Denny Joseph, K.M.; Muralidhara, M. Fish oil prophylaxis attenuates rotenone-induced oxidative impairments and mitochondrial dysfunctions in rat brain. Food Chem. Toxicol. 2012, 50, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Khurana, N.; Gajbhiye, A. Ameliorative effect of Sida cordifolia in rotenone induced oxidative stress model of Parkinson’s disease. Neurotoxicology 2013, 39, 57–64. [Google Scholar] [CrossRef]
- Kaur, H.; Chauhan, S.; Sandhir, R. Protective effect of lycopene on oxidative stress and cognitive decline in rotenone induced model of Parkinson’s disease. Neurochem. Res. 2011, 36, 1435–1443. [Google Scholar] [CrossRef]
- Javed, H.; Azimullah, S.; Meeran, M.F.N.; Ansari, S.A.; Ojha, S. Neuroprotective Effects of Thymol, a Dietary Monoterpene Against Dopaminergic Neurodegeneration in Rotenone-Induced Rat Model of Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarbishegi, M.; Charkhat Gorgich, E.A.; Khajavi, O.; Komeili, G.; Salimi, S. The neuroprotective effects of hydro-alcoholic extract of olive (Olea europaea L.) leaf on rotenone-induced Parkinson’s disease in rat. Metab. Brain Dis. 2018, 33, 79–88. [Google Scholar] [CrossRef]
- Zaitone, S.A.; Abo-Elmatty, D.M.; Shaalan, A.A. Acetyl-L-carnitine and α-lipoic acid affect rotenone-induced damage in nigral dopaminergic neurons of rat brain, implication for Parkinson’s disease therapy. Pharmacol. Biochem. Behav. 2012, 100, 347–360. [Google Scholar] [CrossRef]
- Verma, R.; Nehru, B. Effect of centrophenoxine against rotenone-induced oxidative stress in an animal model of Parkinson’s disease. Neurochem. Int. 2009, 55, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Palle, S.; Neerati, P. Improved neuroprotective effect of resveratrol nanoparticles as evinced by abrogation of rotenone-induced behavioral deficits and oxidative and mitochondrial dysfunctions in rat model of Parkinson’s disease. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2018, 391, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Ojha, S.; Javed, H.; Azimullah, S.; Abul Khair, S.B.; Haque, M.E. Glycyrrhizic acid Attenuates Neuroinflammation and Oxidative Stress in Rotenone Model of Parkinson’s Disease. Neurotox. Res. 2016, 29, 275–287. [Google Scholar] [CrossRef]
- Jayaraj, R.L.; Beiram, R.; Azimullah, S.; Meeran, M.F.N.; Ojha, S.K.; Adem, A.; Jalal, F.Y. Lycopodium Attenuates Loss of Dopaminergic Neurons by Suppressing Oxidative Stress and Neuroinflammation in a Rat Model of Parkinson’s Disease. Molecules 2019, 24, 2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandran, G.; Muralidhara. Neuroprotective effect of aqueous extract of Selaginella delicatula as evidenced by abrogation of rotenone-induced motor deficits, oxidative dysfunctions, and neurotoxicity in mice. Cell. Mol. Neurobiol. 2013, 33, 929–942. [Google Scholar] [CrossRef]
- Farombi, E.O.; Awogbindin, I.O.; Farombi, T.H.; Oladele, J.O.; Izomoh, E.R.; Aladelokun, O.B.; Ezekiel, I.O.; Adebambo, O.I.; Abah, V.O. Neuroprotective role of kolaviron in striatal redo-inflammation associated with rotenone model of Parkinson’s disease. Neurotoxicology 2019, 73, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.V.; Hemalatha, P.; Yetish, S.; Muralidhara, M.; Rajini, P.S. Prophylactic neuroprotective propensity of Crocin, a carotenoid against rotenone induced neurotoxicity in mice: Behavioural and biochemical evidence. Metab. Brain Dis. 2019, 34, 1341–1353. [Google Scholar] [CrossRef]
- Alzahrani, S.; Ezzat, W.; Elshaer, R.E.; Abd El-Lateef, A.S.; Mohammad, H.M.F.; Elkazaz, A.Y.; Toraih, E.; Zaitone, S.A. Standarized Tribulus terrestris extract protects against rotenone-induced oxidative damage and nigral dopamine neuronal loss in mice. J. Physiol. Pharmacol. 2018, 69. [Google Scholar] [CrossRef]
- Manjunath, M.J.; Muralidhara. Effect of Withania somnifera supplementation on rotenone-induced oxidative damage in cerebellum and striatum of the male mice brain. Cent. Nerv. Syst. Agents Med. Chem. 2013, 13, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca-Fonseca, L.A.; Wong-Guerra, M.; Ramírez-Sánchez, J.; Montano-Peguero, Y.; Padrón Yaquis, A.S.; Rodríguez, A.M.; da Silva, V.D.A.; Costa, S.L.; Pardo-Andreu, G.L.; Núñez-Figueredo, Y. JM-20, a novel hybrid molecule, protects against rotenone-induced neurotoxicity in experimental model of Parkinson’s disease. Neurosci. Lett. 2019, 690, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Reus, M.I.; Gómez del Rio, M.A.; Iglesias, I.; Elorza, M.; Slowing, K.; Benedí, J. Standardized Hypericum perforatum reduces oxidative stress and increases gene expression of antioxidant enzymes on rotenone-exposed rats. Neuropharmacology 2007, 52, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Zaminelli, T.; Gradowski, R.W.; Bassani, T.B.; Barbiero, J.K.; Santiago, R.M.; Maria-Ferreira, D.; Baggio, C.H.; Vital, M.A. Antidepressant and antioxidative effect of Ibuprofen in the rotenone model of Parkinson’s disease. Neurotox. Res. 2014, 26, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Zagoura, D.; Canovas-Jorda, D.; Pistollato, F.; Bremer-Hoffmann, S.; Bal-Price, A. Evaluation of the rotenone-induced activation of the Nrf2 pathway in a neuronal model derived from human induced pluripotent stem cells. Neurochem. Int. 2017, 106, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Deza-Ponzio, R.; Herrera, M.L.; Bellini, M.J.; Virgolini, M.B.; Hereñú, C.B. Aldehyde dehydrogenase 2 in the spotlight: The link between mitochondria and neurodegeneration. Neurotoxicology 2018, 68, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, D.S.; Sullivan, P.; Cooney, A.; Jinsmaa, Y.; Kopin, I.J.; Sharabi, Y. Rotenone decreases intracellular aldehyde dehydrogenase activity: Implications for the pathogenesis of Parkinson’s disease. J. Neurochem. 2015, 133, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Sthijns, M.M.; Weseler, A.R.; Bast, A.; Haenen, G.R. Time in Redox Adaptation Processes: From Evolution to Hormesis. Int. J. Mol. Sci. 2016, 17, 1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, M.; Bauer, I. Heme oxygenase-1: Redox regulation and role in the hepatic response to oxidative stress. Antioxid. Redox Signal. 2002, 4, 749–758. [Google Scholar] [CrossRef]
- Deshane, J.; Wright, M.; Agarwal, A. Heme oxygenase-1 expression in disease states. Acta Biochim. Pol. 2005, 52, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Nakahira, K.; Takahashi, T.; Shimizu, H.; Maeshima, K.; Uehara, K.; Fujii, H.; Nakatsuka, H.; Yokoyama, M.; Akagi, R.; Morita, K. Protective role of heme oxygenase-1 induction in carbon tetrachloride-induced hepatotoxicity. Biochem. Pharmacol. 2003, 66, 1091–1105. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Morita, K.; Akagi, R.; Sassa, S. Heme oxygenase-1: A novel therapeutic target in oxidative tissue injuries. Curr. Med. Chem. 2004, 11, 1545–1561. [Google Scholar] [CrossRef]
- Ross, D.; Siegel, D. Functions of NQO1 in Cellular Protection and CoQ. Front. Physiol. 2017, 8, 595. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [Green Version]
- Litvinov, D.; Mahini, H.; Garelnabi, M. Antioxidant and anti-inflammatory role of paraoxonase 1: Implication in arteriosclerosis diseases. N. Am. J. Med. Sci. 2012, 4, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.G.; Wang, T.; Vance, T.M.; Hubert, P.; Kim, D.O.; Koo, S.I.; Chun, O.K. Validation of Analytical Methods for Plasma Total Antioxidant Capacity by Comparing with Urinary 8-Isoprostane Level. J. Microbiol. Biotechnol. 2017, 27, 388–394. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Du, Y.Q.; Yuan, L.; Wang, N.N. Protective effect of puerarin on acute alcoholic liver injury. Am. J. Chin. Med. 2010, 38, 241–249. [Google Scholar] [CrossRef]
- Yeom, Y.; Kim, Y. The Sasa quelpaertensis Leaf Extract Inhibits the Dextran Sulfate Sodium-induced Mouse Colitis through Modulation of Antioxidant Enzyme Expression. J. Cancer Prev. 2015, 20, 136–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deyhim, F.; Patil, B.S.; Villarreal, A.; Lopez, E.; Garcia, K.; Rios, R.; Garcia, C.; Gonzales, C.; Mandadi, K. Cranberry juice increases antioxidant status without affecting cholesterol homeostasis in orchidectomized rats. J. Med. Food 2007, 10, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Karuna, R.; Reddy, S.S.; Baskar, R.; Saralakumari, D. Antioxidant potential of aqueous extract of Phyllanthus amarus in rats. Indian J. Pharmacol. 2009, 41, 64–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjbar, A.; Mohsenzadeh, F.; Chehregani, A.; Khajavi, F.; Zijoud, S.M.; Ghasemi, H. Ameliorative effect of Matricaria chamomilla .L on paraquat: Induced oxidative damage in lung rats. Pharmacogn. Res. 2014, 6, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Gumulec, J.; Raudenska, M.; Hlavna, M.; Stracina, T.; Sztalmachova, M.; Tanhauserova, V.; Pacal, L.; Ruttkay-Nedecky, B.; Sochor, J.; Zitka, O.; et al. Determination of oxidative stress and activities of antioxidant enzymes in guinea pigs treated with haloperidol. Exp. Ther. Med. 2013, 5, 479–484. [Google Scholar] [CrossRef] [Green Version]
- Gechev, T.; Gadjev, I.; Van Breusegem, F.; Inzé, D.; Dukiandjiev, S.; Toneva, V.; Minkov, I. Hydrogen peroxide protects tobacco from oxidative stress by inducing a set of antioxidant enzymes. Cell. Mol. Life Sci. 2002, 59, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.H.; Lee, H.C. Oxidative stress, mitochondrial DNA mutation, and impairment of antioxidant enzymes in aging. Exp. Biol. Med. 2002, 227, 671–682. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Parameters | Treatment | ||||
|---|---|---|---|---|---|
| Control | CJ II | ROT | CJ I + ROT | CJ II + ROT | |
| SOD [U/mg protein] | 39.40 ± 4.37 | 67.62 ± 16.48 (a) | 56.17 ± 10.91 (a) | 52.31 ± 12.38 | 66.45 ± 16.19 |
| CAT [U/mg protein] | 0.60 ± 0.23 | 0.85 ± 0.27 | 0.77 ± 0.23 | 1.06 ± 0.26 | 1.09 ± 0.24 |
| GR [nmol NADPH/min/mg protein] | 0.33 ± 0.09 | 0.58 ± 0.13 (a) | 0.51 ± 0.11 (a) | 0.18 ± 0.04 (b) | 0.15 ± 0.03 (b) |
| GST [nmol CDNB/min/mg protein | 362.4 ± 96.6 | 480.8 ± 110.3 | 523.4 ± 128.0 (a) | 577.8 ± 170.9 | 495.2 ± 92.2 |
| ALDH2 [nmol NADPH/min/mg protein] | 7.95 ± 0.65 | 6.93 ± 0.98 | 6.31 ± 0.43 (a) | 6.52 ± 1.12 | 7.11 ± 0.93 |
| MDA [nmol/g tissue] | 48.3 ± 8.7 | 16.5 ± 1.9 (a) | 89.65 ± 55.0 (a) | 18.8 ± 6.6a (b) | 22.3 ± 4.9 (b) |
| DNA damage [arbitrary points] | 112 ± 6 | 111 ± 3 | 172 ± 10 (a) | 140 ± 4 (b) | 124 ± 4 (b) |
| Parameters | Treatment | ||||
|---|---|---|---|---|---|
| Control | CJ II | ROT | CJ I + TOT | CJ II + ROT | |
| SOD [U/mg protein] | 27.7 ± 5.0 | 47.1 ± 12.4 (a) | 32.8 ± 11.0 | 52.8 ± 7.8 (b) | 60.8 ± 9.7 (b) |
| CAT [U/mg protein] | 2.55 ± 0.37 | 3.02 ± 0.67 | 2.83 ± 0.57 | 3.97 ± 0.73 (b) | 3.55 ± 0.50 |
| GPx [nmol NADPH/min/mg protein] | 285.7 ± 62.6 | 453.0 ± 90.7 (a) | 493.8 ± 95.2 (a) | 485.8 ± 71.8 | 502.9 ± 79.0 |
| GR [nmol NADPH/min/mg protein] | 72.1 ± 8.8 | 69.9 ± 5.9 | 56.4 ± 9.9 (a) | 54.3 ± 7.3 | 56.7 ± 6.7 |
| HO-1 [pmol bilirubin/min/mg protein] | 13.2 ± 3.2 | 17.9 ± 5.5 | 23.7 ± 7.2 (a) | 21.7 ± 8.4 | 20.0 ± 6.7 |
| NQO1 [nmol cyt c/min/mg protein] | 8.8 ± 2.9 | 17.6 ± 3.2 (a) | 21.2 ± 3.5 (a) | 23.4 ± 6.6 | 21.3 ± 6.2 |
| PON1 [μmol phenol/min/mg protein] | 722.5 ± 92.6 | 814.1 ± 92.3 | 807.3 ± 66.3 | 526.3 ± 39.1 (b) | 1470.9 ± 120.3 (b) |
| DNA damage [arbitrary points] | 73 ± 3 | 78 ± 3 | 123 ± 4 (a) | 118 ± 5 | 121 ± 4 |
| Parameters | Treatment | ||||
|---|---|---|---|---|---|
| Control | CJ II | ROT | CJ I + ROT | CJ II + ROT | |
| ALT [U/L] | 20.6 ± 1.8 | 22.7 ± 1.7 | 39.3 ± 5.5 (a) | 39.6 ± 3.9 | 27.6 ± 3.6 (b) |
| AST [U/L] | 82.6 ± 10.7 | 84.3 ± 12.6 | 236.7 ± 28.3 (a) | 97.6 ± 8.5 (b) | 115.6 ± 11.5 (b) |
| ALP [U/L] | 78.7 ± 9.2 | 104.3 ± 10.2 (a) | 102.9 ± 15.1 (a) | 114.6 ± 15.8 | 111.1 ± 17.9 |
| LDH [U/L] | 128.9 ± 16.7 | 138.2 ± 16.4 | 412.9 ± 54.5 (a) | 247.0 ± 38.0 (b) | 185.7 ± 29.1 (b) |
| Parameters | Treatment | ||||
|---|---|---|---|---|---|
| Control | CJ II | ROT | CJ I + TOT | CJ II + ROT | |
| CAT [U/mg protein] | 0.16 ± 0.01 | 0.17 ± 0.01 | 0.11 ± 0.01 (a) | 0.14 ± 0.01 (b) | 0.15 ± 0.01 (b) |
| GPx [nmol NADPH/min/mg protein] | 132.5 ± 16.0 | 136.9 ± 10.5 | 182.8 ± 17.4 (a) | 162 ± 24.1 | 162.1 ± 15.5 |
| GR [nmol NADPH/min/mg protein] | 56.9 ± 5.9 | 56.5 ± 5.0 | 63.3 ± 5.0 (a) | 54.9 ± 3.1 (b) | 50.5 ± 3.5 (b) |
| PON1 [μmol phenol/min/mg protein] | 138.1 ± 10.3 | 137.4 ± 8.0 | 117.6 ± 8.2 (a) | 143.4 ± 13.7 (b) | 139.9 ± 12.7 (b) |
| GSH [μmol/g tissue] | 3.08 ± 0.16 | 3.39 ± 0.33 | 5.55 ± 0.50 (a) | 5.42 ± 0.51 | 4.63 ± 0.32 (b) |
| DNA damage [arbitrary points] | 91.0 ± 4.0 | 90.9 ± 5.3 | 174.0 ± 14.4 (a) | 183.5 ± 3.0 | 180.3 ± 2.1 |
| Parameters | Treatment | ||||
|---|---|---|---|---|---|
| Control | CJ II | ROT | CJ I + ROT | CJ II + ROT | |
| SOD [U/mL] | 96.5 ± 10.3 | 107.1 ± 17.1 | 85.3 ± 11.2 | 74.2 ± 11.6 | 70.9 ± 10.7 (b) |
| GPx [nmol NADPH/min/mL] | 2045.2 ± 143.1 | 2089.4 ± 92.0 | 2053.3 ± 254.9 | 1331.4 ± 70.7 (b) | 175.9 ± 38.1 (b) |
| PON1 [μmol phenol/min/mL] | 31.9 ± 4.1 | 37.7 ± 1.7 | 34.9 ± 5.5 | 41.9 ± 5.9 (b) | 43.0 ± 5.3 (b) |
| TAS# [μmol vit C/min/mL] | 2.37 ± 0.03 | 2.30 ± 0.06 | 1.74 ± 0.09 (a) | 1.90 ± 0.04 | 2.00 ± 0.01 (b) |
| DNA damage * [arbitrary points] | 63 ± 3 | 72 ± 4 | 112 ± 4 (a) | 119 ± 5 | 120 ± 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurpik, M.; Zalewski, P.; Kujawska, M.; Ewertowska, M.; Ignatowicz, E.; Cielecka-Piontek, J.; Jodynis-Liebert, J. Can Cranberry Juice Protect against Rotenone-Induced Toxicity in Rats? Nutrients 2021, 13, 1050. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13041050
Kurpik M, Zalewski P, Kujawska M, Ewertowska M, Ignatowicz E, Cielecka-Piontek J, Jodynis-Liebert J. Can Cranberry Juice Protect against Rotenone-Induced Toxicity in Rats? Nutrients. 2021; 13(4):1050. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13041050
Chicago/Turabian StyleKurpik, Monika, Przemysław Zalewski, Małgorzata Kujawska, Małgorzata Ewertowska, Ewa Ignatowicz, Judyta Cielecka-Piontek, and Jadwiga Jodynis-Liebert. 2021. "Can Cranberry Juice Protect against Rotenone-Induced Toxicity in Rats?" Nutrients 13, no. 4: 1050. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13041050