
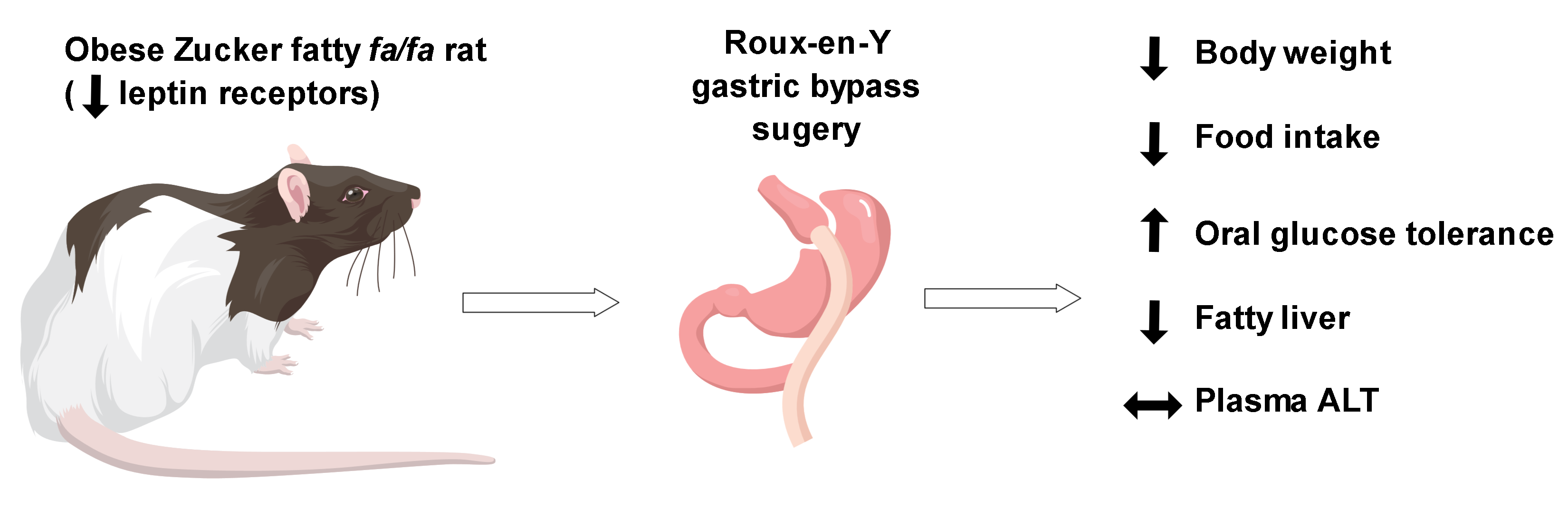
Leptin Receptors Are Not Required for Roux-en-Y Gastric Bypass Surgery to Normalize Energy and Glucose Homeostasis in Rats



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
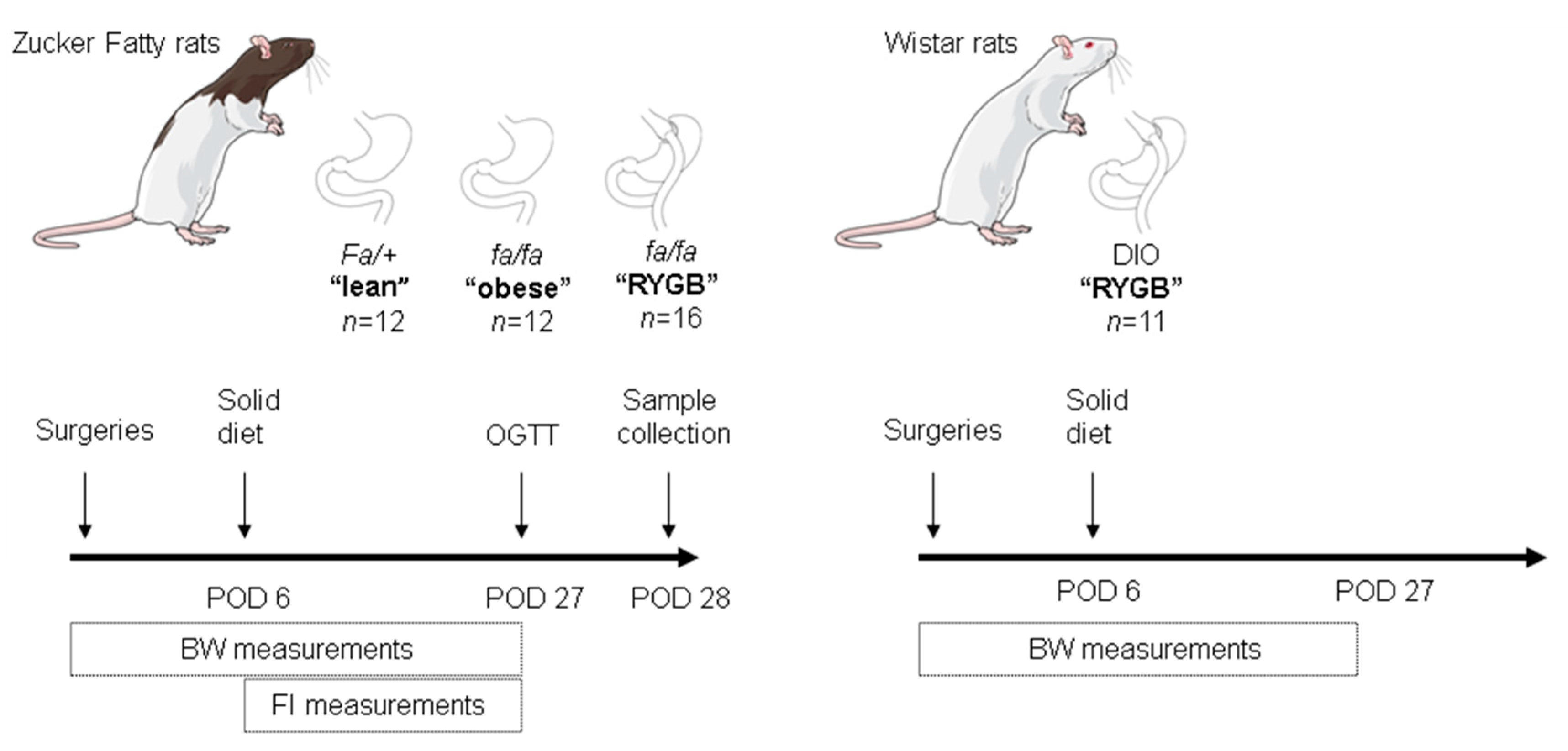
2. Materials and Methods
2.1. Animals
2.2. Surgeries
2.3. Perioperative Care
2.4. Metabolic Measurements
2.5. Tissue Harvesting
2.6. ELISAs
2.7. Bomb Calorimetry
2.8. Liver Histology
2.9. Statistics
3. Results
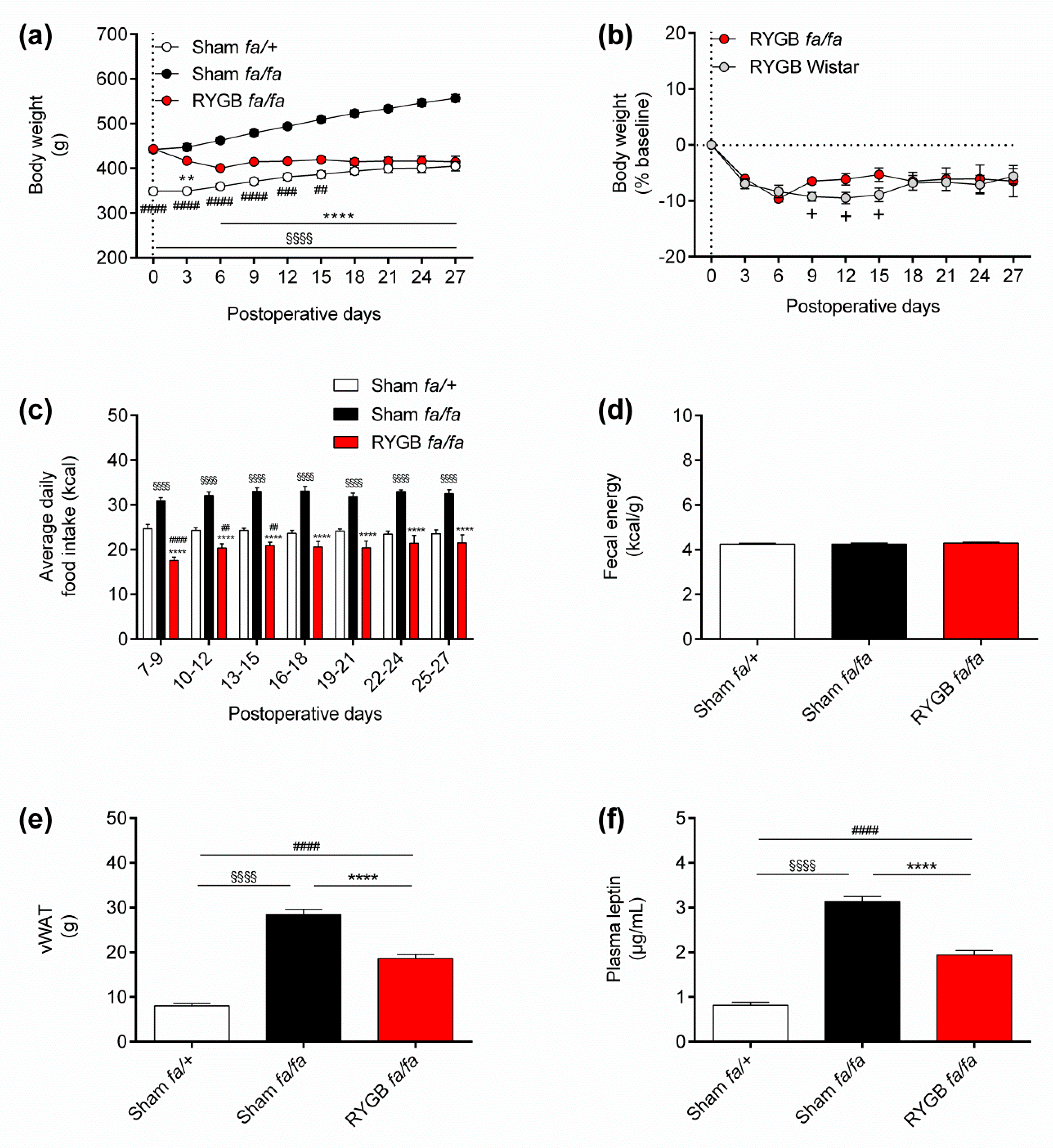
3.1. Leptin Receptors Are Not Required for RYGB to Normalize Energy Homeostasis
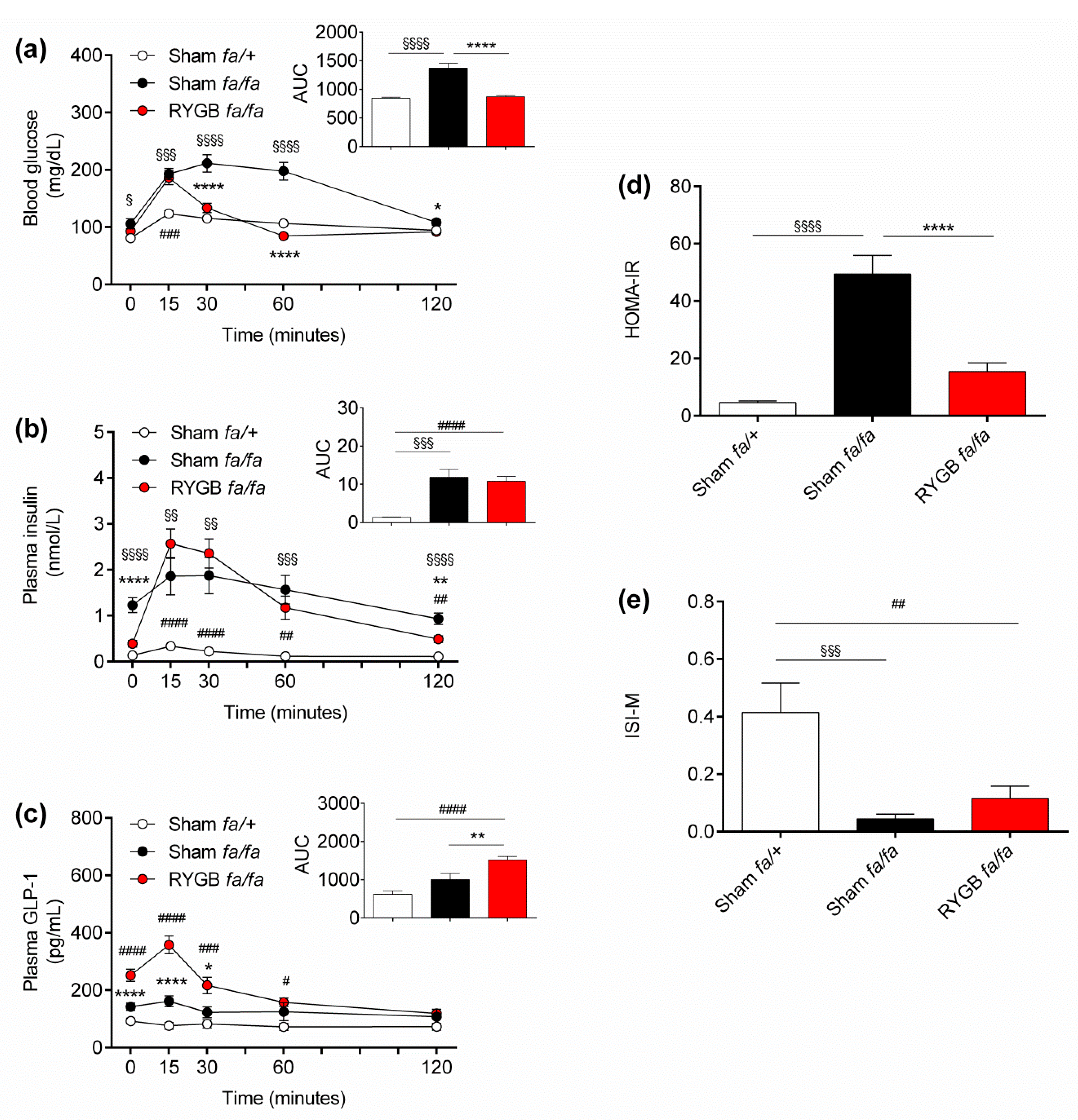
3.2. Leptin Receptors Are Not Required for RYGB to Normalize Oral Glucose Tolerance
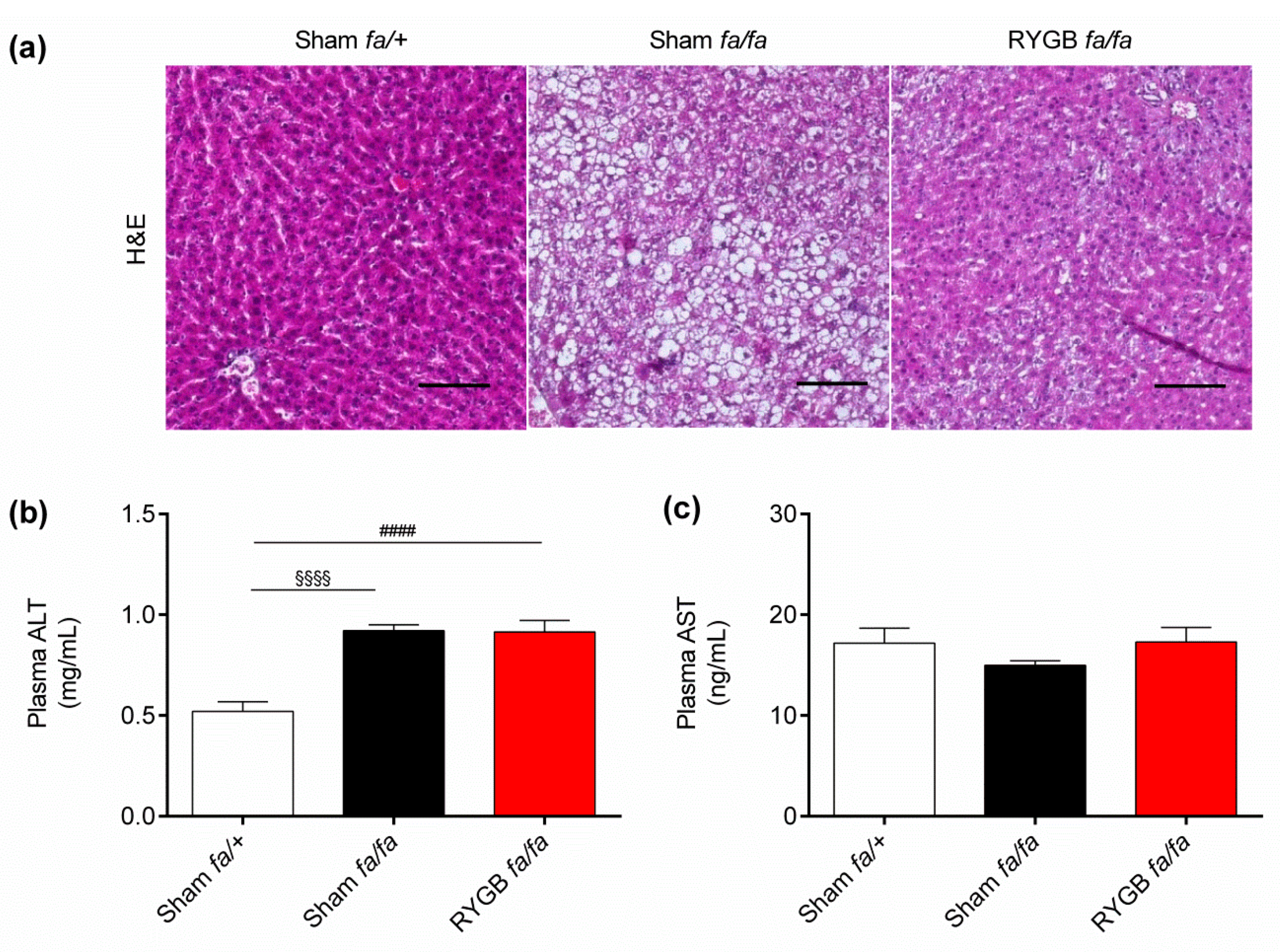
3.3. Leptin Receptors Might Be Required for RYGB to Normalize Liver Health
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lassailly, G.; Caiazzo, R.; Ntandja-Wandji, L.C.; Gnemmi, V.; Baud, G.; Verkindt, H.; Ningarhari, M.; Louvet, A.; Leteurtre, E.; Raverdy, V.; et al. Bariatric Surgery Provides Long-term Resolution of Nonalcoholic Steatohepatitis and Regression of Fibrosis. Gastroenterology 2020, 159, 1290–1301.e1295. [Google Scholar] [CrossRef]
- Mingrone, G.; Panunzi, S.; De Gaetano, A.; Guidone, C.; Iaconelli, A.; Capristo, E.; Chamseddine, G.; Bornstein, S.R.; Rubino, F. Metabolic surgery versus conventional medical therapy in patients with type 2 diabetes: 10-year follow-up of an open-label, single-centre, randomised controlled trial. Lancet 2021, 397, 293–304. [Google Scholar] [CrossRef]
- Adams, T.D.; Davidson, L.E.; Litwin, S.E.; Kim, J.; Kolotkin, R.L.; Nanjee, M.N.; Gutierrez, J.M.; Frogley, S.J.; Ibele, A.R.; Brinton, E.A.; et al. Weight and Metabolic Outcomes 12 Years after Gastric Bypass. N. Engl. J. Med. 2017, 377, 1143–1155. [Google Scholar] [CrossRef]
- Schauer, P.R.; Bhatt, D.L.; Kirwan, J.P.; Wolski, K.; Aminian, A.; Brethauer, S.A.; Navaneethan, S.D.; Singh, R.P.; Pothier, C.E.; Nissen, S.E.; et al. Bariatric Surgery versus Intensive Medical Therapy for Diabetes—5-Year Outcomes. N. Engl. J. Med. 2017, 376, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Sjostrom, L.; Narbro, K.; Sjostrom, C.D.; Karason, K.; Larsson, B.; Wedel, H.; Lystig, T.; Sullivan, M.; Bouchard, C.; Carlsson, B.; et al. Effects of bariatric surgery on mortality in Swedish obese subjects. N. Engl. J. Med. 2007, 357, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Evers, S.S.; Sandoval, D.A.; Seeley, R.J. The Physiology and Molecular Underpinnings of the Effects of Bariatric Surgery on Obesity and Diabetes. Annu. Rev. Physiol. 2017, 79, 313–334. [Google Scholar] [CrossRef]
- Kwon, I.G.; Kang, C.W.; Park, J.P.; Oh, J.H.; Wang, E.K.; Kim, T.Y.; Sung, J.S.; Park, N.; Lee, Y.J.; Sung, H.J.; et al. Serum glucose excretion after Roux-en-Y gastric bypass: A potential target for diabetes treatment. Gut 2020. [Google Scholar] [CrossRef]
- Wei, X.; Lu, Z.; Li, L.; Zhang, H.; Sun, F.; Ma, H.; Wang, L.; Hu, Y.; Yan, Z.; Zheng, H.; et al. Reducing NADPH Synthesis Counteracts Diabetic Nephropathy through Restoration of AMPK Activity in Type 1 Diabetic Rats. Cell Rep. 2020, 32, 108207. [Google Scholar] [CrossRef]
- Ye, Y.; Abu El Haija, M.; Morgan, D.A.; Guo, D.; Song, Y.; Frank, A.; Tian, L.; Riedl, R.A.; Burnett, C.M.L.; Gao, Z.; et al. Endocannabinoid Receptor-1 and Sympathetic Nervous System Mediate the Beneficial Metabolic Effects of Gastric Bypass. Cell Rep. 2020, 33, 108270. [Google Scholar] [CrossRef]
- Ben-Zvi, D.; Meoli, L.; Abidi, W.M.; Nestoridi, E.; Panciotti, C.; Castillo, E.; Pizarro, P.; Shirley, E.; Gourash, W.F.; Thompson, C.C.; et al. Time-Dependent Molecular Responses Differ between Gastric Bypass and Dieting but Are Conserved Across Species. Cell Metab. 2018, 28, 310–323.e316. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.M. Leptin and the endocrine control of energy balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Seoane-Collazo, P.; Martinez-Sanchez, N.; Milbank, E.; Contreras, C. Incendiary Leptin. Nutrients 2020, 12, 472. [Google Scholar] [CrossRef] [Green Version]
- D’Souza A, M.; Neumann, U.H.; Glavas, M.M.; Kieffer, T.J. The glucoregulatory actions of leptin. Mol. Metab. 2017, 6, 1052–1065. [Google Scholar] [CrossRef] [PubMed]
- Hackl, M.T.; Furnsinn, C.; Schuh, C.M.; Krssak, M.; Carli, F.; Guerra, S.; Freudenthaler, A.; Baumgartner-Parzer, S.; Helbich, T.H.; Luger, A.; et al. Brain leptin reduces liver lipids by increasing hepatic triglyceride secretion and lowering lipogenesis. Nat. Commun. 2019, 10, 2717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Su, H.; Wu, X.; Shen, H.; Kim, M.H.; Li, Y.; Myers, M.G., Jr.; Owyang, C.; Rui, L. Leptin receptor-expressing neuron Sh2b1 supports sympathetic nervous system and protects against obesity and metabolic disease. Nat. Commun. 2020, 11, 1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingalls, A.M.; Dickie, M.M.; Snell, G.D. Obese, a new mutation in the house mouse. J. Hered. 1950, 41, 317–318. [Google Scholar] [CrossRef]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Knight, Z.A.; Hannan, K.S.; Greenberg, M.L.; Friedman, J.M. Hyperleptinemia is required for the development of leptin resistance. PLoS ONE 2010, 5, e11376. [Google Scholar] [CrossRef]
- Zhao, S.; Zhu, Y.; Schultz, R.D.; Li, N.; He, Z.; Zhang, Z.; Caron, A.; Zhu, Q.; Sun, K.; Xiong, W.; et al. Partial Leptin Reduction as an Insulin Sensitization and Weight Loss Strategy. Cell Metab. 2019, 30, 706–719.e706. [Google Scholar] [CrossRef]
- Loh, K.; Fukushima, A.; Zhang, X.; Galic, S.; Briggs, D.; Enriori, P.J.; Simonds, S.; Wiede, F.; Reichenbach, A.; Hauser, C.; et al. Elevated hypothalamic TCPTP in obesity contributes to cellular leptin resistance. Cell Metab. 2011, 14, 684–699. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, L.E.; Unger, E.K.; Cheung, C.C.; Xu, A.W. Modulation of AgRP-neuronal function by SOCS3 as an initiating event in diet-induced hypothalamic leptin resistance. Proc. Natl. Acad. Sci. USA 2013, 110, E697–E706. [Google Scholar] [CrossRef] [Green Version]
- Valdearcos, M.; Robblee, M.M.; Benjamin, D.I.; Nomura, D.K.; Xu, A.W.; Koliwad, S.K. Microglia dictate the impact of saturated fat consumption on hypothalamic inflammation and neuronal function. Cell Rep. 2014, 9, 2124–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, G.; Zhang, H.; Karin, M.; Bai, H.; Cai, D. Hypothalamic IKKbeta/NF-kappaB and ER stress link overnutrition to energy imbalance and obesity. Cell 2008, 135, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Halaas, J.L.; Gajiwala, K.S.; Maffei, M.; Cohen, S.L.; Chait, B.T.; Rabinowitz, D.; Lallone, R.L.; Burley, S.K.; Friedman, J.M. Weight-reducing effects of the plasma protein encoded by the obese gene. Science 1995, 269, 543–546. [Google Scholar] [CrossRef]
- Pelleymounter, M.A.; Cullen, M.J.; Baker, M.B.; Hecht, R.; Winters, D.; Boone, T.; Collins, F. Effects of the obese gene product on body weight regulation in ob/ob mice. Science 1995, 269, 540–543. [Google Scholar] [CrossRef]
- Singh, A.; Wirtz, M.; Parker, N.; Hogan, M.; Strahler, J.; Michailidis, G.; Schmidt, S.; Vidal-Puig, A.; Diano, S.; Andrews, P.; et al. Leptin-mediated changes in hepatic mitochondrial metabolism, structure, and protein levels. Proc. Natl. Acad. Sci. USA 2009, 106, 13100–13105. [Google Scholar] [CrossRef] [Green Version]
- Campfield, L.A.; Smith, F.J.; Guisez, Y.; Devos, R.; Burn, P. Recombinant mouse OB protein: Evidence for a peripheral signal linking adiposity and central neural networks. Science 1995, 269, 546–549. [Google Scholar] [CrossRef]
- Lee, J.; Liu, J.; Feng, X.; Salazar Hernandez, M.A.; Mucka, P.; Ibi, D.; Choi, J.W.; Ozcan, U. Withaferin A is a leptin sensitizer with strong antidiabetic properties in mice. Nat. Med. 2016, 22, 1023–1032. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lee, J.; Salazar Hernandez, M.A.; Mazitschek, R.; Ozcan, U. Treatment of obesity with celastrol. Cell 2015, 161, 999–1011. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Guan, D.; Auen, T.; Choi, J.W.; Salazar Hernandez, M.A.; Lee, J.; Chun, H.; Faruk, F.; Kaplun, E.; Herbert, Z.; et al. IL1R1 is required for celastrol’s leptin-sensitization and antiobesity effects. Nat. Med. 2019, 25, 575–582. [Google Scholar] [CrossRef]
- Dodd, G.T.; Xirouchaki, C.E.; Eramo, M.; Mitchell, C.A.; Andrews, Z.B.; Henry, B.A.; Cowley, M.A.; Tiganis, T. Intranasal Targeting of Hypothalamic PTP1B and TCPTP Reinstates Leptin and Insulin Sensitivity and Promotes Weight Loss in Obesity. Cell Rep. 2019, 28, 2905–2922.e2905. [Google Scholar] [CrossRef]
- Tam, J.; Cinar, R.; Liu, J.; Godlewski, G.; Wesley, D.; Jourdan, T.; Szanda, G.; Mukhopadhyay, B.; Chedester, L.; Liow, J.S.; et al. Peripheral cannabinoid-1 receptor inverse agonism reduces obesity by reversing leptin resistance. Cell Metab. 2012, 16, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Haghighat, N.; Kazemi, A.; Asbaghi, O.; Jafarian, F.; Moeinvaziri, N.; Hosseini, B.; Amini, M. Long-term effect of bariatric surgery on body composition in patients with morbid obesity: A systematic review and meta-analysis. Clin. Nutr. 2020, 40, 1755–1766. [Google Scholar] [CrossRef]
- Arakawa, R.; Febres, G.; Cheng, B.; Krikhely, A.; Bessler, M.; Korner, J. Prospective study of gut hormone and metabolic changes after laparoscopic sleeve gastrectomy and Roux-en-Y gastric bypass. PLoS ONE 2020, 15, e0236133. [Google Scholar] [CrossRef]
- Salman, M.A.; El-Ghobary, M.; Soliman, A.; El Sherbiny, M.; Abouelregal, T.E.; Albitar, A.; Abdallah, A.; Mikhail, H.M.S.; Nafea, M.A.; Sultan, A.; et al. Long-Term Changes in Leptin, Chemerin, and Ghrelin Levels Following Roux-en-Y Gastric Bypass and Laparoscopic Sleeve Gastrectomy. Obes. Surg. 2020, 30, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Unamuno, X.; Izaguirre, M.; Gomez-Ambrosi, J.; Rodriguez, A.; Ramirez, B.; Becerril, S.; Valenti, V.; Moncada, R.; Silva, C.; Salvador, J.; et al. Increase of the Adiponectin/Leptin Ratio in Patients with Obesity and Type 2 Diabetes after Roux-en-Y Gastric Bypass. Nutrients 2019, 11, 2069. [Google Scholar] [CrossRef] [Green Version]
- Guijarro, A.; Osei-Hyiaman, D.; Harvey-White, J.; Kunos, G.; Suzuki, S.; Nadtochiy, S.; Brookes, P.S.; Meguid, M.M. Sustained weight loss after Roux-en-Y gastric bypass is characterized by down regulation of endocannabinoids and mitochondrial function. Ann. Surg. 2008, 247, 779–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, R.G.; Kumar, S.; Hall, C.E.; Palaia, T.; Rideout, D.A.; Hall, K.; Brathwaite, C.E.; Ragolia, L. Roux-en-Y gastric bypass attenuates the progression of cardiometabolic complications in obese diabetic rats via alteration in gastrointestinal hormones. Surg. Obes. Relat. Dis. 2015, 11, 1044–1053. [Google Scholar] [CrossRef]
- Lips, M.A.; Pijl, H.; van Klinken, J.B.; de Groot, G.H.; Janssen, I.M.; Van Ramshorst, B.; Van Wagensveld, B.A.; Swank, D.J.; Van Dielen, F.; Smit, J.W. Roux-en-Y gastric bypass and calorie restriction induce comparable time-dependent effects on thyroid hormone function tests in obese female subjects. Eur. J. Endocrinol. 2013, 169, 339–347. [Google Scholar] [CrossRef]
- Lips, M.A.; van Klinken, J.B.; Pijl, H.; Janssen, I.; van Dijk, K.W.; Koning, F.; van Harmelen, V. Weight loss induced by very low calorie diet is associated with a more beneficial systemic inflammatory profile than by Roux-en-Y gastric bypass. Metabolism 2016, 65, 1614–1620. [Google Scholar] [CrossRef]
- Neinast, M.D.; Frank, A.P.; Zechner, J.F.; Li, Q.; Vishvanath, L.; Palmer, B.F.; Aguirre, V.; Gupta, R.K.; Clegg, D.J. Activation of natriuretic peptides and the sympathetic nervous system following Roux-en-Y gastric bypass is associated with gonadal adipose tissues browning. Mol. Metab. 2015, 4, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Olivan, B.; Teixeira, J.; Bose, M.; Bawa, B.; Chang, T.; Summe, H.; Lee, H.; Laferrere, B. Effect of weight loss by diet or gastric bypass surgery on peptide YY3-36 levels. Ann. Surg. 2009, 249, 948–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Z.; Munzberg, H.; Rezai-Zadeh, K.; Keenan, M.; Coulon, D.; Lu, H.; Berthoud, H.R.; Ye, J. Leptin deficient ob/ob mice and diet-induced obese mice responded differently to Roux-en-Y bypass surgery. Int. J. Obes. 2015, 39, 798–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokadem, M.; Zechner, J.F.; Uchida, A.; Aguirre, V. Leptin Is Required for Glucose Homeostasis after Roux-en-Y Gastric Bypass in Mice. PLoS ONE 2015, 10, e0139960. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Haase, N.; Haange, S.B.; Sucher, R.; Munzker, J.; Jager, E.; Schischke, K.; Seyfried, F.; von Bergen, M.; Hankir, M.K.; et al. Roux-en-Y gastric bypass contributes to weight loss-independent improvement in hypothalamic inflammation and leptin sensitivity through gut-microglia-neuron-crosstalk. Mol. Metab. 2021, 48, 101214. [Google Scholar] [CrossRef]
- Hankir, M.K.; Seyfried, F. Partial Leptin Reduction: An Emerging Weight Loss Paradigm. Trends Endocrinol. Metab. 2020, 31, 395–397. [Google Scholar] [CrossRef]
- Natividad, J.M.; Agus, A.; Planchais, J.; Lamas, B.; Jarry, A.C.; Martin, R.; Michel, M.L.; Chong-Nguyen, C.; Roussel, R.; Straube, M.; et al. Impaired Aryl Hydrocarbon Receptor Ligand Production by the Gut Microbiota Is a Key Factor in Metabolic Syndrome. Cell Metab. 2018, 28, 737–749.e734. [Google Scholar] [CrossRef] [Green Version]
- Ceccarini, G.; Pelosini, C.; Ferrari, F.; Magno, S.; Vitti, J.; Salvetti, G.; Moretto, C.; Marioni, A.; Buccianti, P.; Piaggi, P.; et al. Serum IGF-binding protein 2 (IGFBP-2) concentrations change early after gastric bypass bariatric surgery revealing a possible marker of leptin sensitivity in obese subjects. Endocrine 2019, 65, 86–93. [Google Scholar] [CrossRef]
- Liu, B.; Kuang, L.; Liu, J. Bariatric surgery relieves type 2 diabetes and modulates inflammatory factors and coronary endothelium eNOS/iNOS expression in db/db mice. Can. J. Physiol. Pharmacol. 2014, 92, 70–77. [Google Scholar] [CrossRef]
- Huang, T.; Fu, J.; Zhang, Z.; Zhang, Y.; Liang, Y.; Ge, C.; Qin, X. Pancreatic islet regeneration through PDX-1/Notch-1/Ngn3 signaling after gastric bypass surgery in db/db mice. Exp. Ther. Med. 2017, 14, 2831–2838. [Google Scholar] [CrossRef] [Green Version]
- Hummel, K.P.; Dickie, M.M.; Coleman, D.L. Diabetes, a new mutation in the mouse. Science 1966, 153, 1127–1128. [Google Scholar] [CrossRef]
- Chen, H.; Charlat, O.; Tartaglia, L.A.; Woolf, E.A.; Weng, X.; Ellis, S.J.; Lakey, N.D.; Culpepper, J.; Moore, K.J.; Breitbart, R.E.; et al. Evidence that the diabetes gene encodes the leptin receptor: Identification of a mutation in the leptin receptor gene in db/db mice. Cell 1996, 84, 491–495. [Google Scholar] [CrossRef] [Green Version]
- Chua, S.C., Jr.; White, D.W.; Wu-Peng, X.S.; Liu, S.M.; Okada, N.; Kershaw, E.E.; Chung, W.K.; Power-Kehoe, L.; Chua, M.; Tartaglia, L.A.; et al. Phenotype of fatty due to Gln269Pro mutation in the leptin receptor (Lepr). Diabetes 1996, 45, 1141–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, M.; Murakami, T.; Ishida, K.; Mizuno, A.; Kuwajima, M.; Shima, K. Substitution at codon 269 (glutamine→proline) of the leptin receptor (OB-R) cDNA is the only mutation found in the Zucker fatty (fa/fa) rat. Biochem. Biophys. Res. Commun. 1996, 224, 597–604. [Google Scholar] [CrossRef]
- Phillips, M.S.; Liu, Q.; Hammond, H.A.; Dugan, V.; Hey, P.J.; Caskey, C.J.; Hess, J.F. Leptin receptor missense mutation in the fatty Zucker rat. Nat. Genet. 1996, 13, 18–19. [Google Scholar] [CrossRef]
- Zucker, L.M.; Zucker, T.F. Fatty, a new mutation in the rat. J. Hered. 1961, 52, 275–278. [Google Scholar] [CrossRef]
- LaBranche, T.P.; Kopec, A.K.; Mantena, S.R.; Hollingshead, B.D.; Harrington, A.W.; Stewart, Z.S.; Zhan, Y.; Hayes, K.D.; Whiteley, L.O.; Burdick, A.D.; et al. Zucker Lean Rats with Hepatic Steatosis Recapitulate Asymptomatic Metabolic Syndrome and Exhibit Greater Sensitivity to Drug-Induced Liver Injury Compared with Standard Nonclinical Sprague-Dawley Rat Model. Toxicol. Pathol. 2020, 48, 994–1007. [Google Scholar] [CrossRef]
- Ionescu, E.; Sauter, J.F.; Jeanrenaud, B. Abnormal oral glucose tolerance in genetically obese (fa/fa) rats. Am. J. Physiol. 1985, 248, E500–E506. [Google Scholar] [CrossRef] [PubMed]
- Carmiel-Haggai, M.; Cederbaum, A.I.; Nieto, N. A high-fat diet leads to the progression of non-alcoholic fatty liver disease in obese rats. FASEB J. 2005, 19, 136–138. [Google Scholar] [CrossRef]
- Bankoglu, E.E.; Seyfried, F.; Rotzinger, L.; Nordbeck, A.; Corteville, C.; Jurowich, C.; Germer, C.T.; Otto, C.; Stopper, H. Impact of weight loss induced by gastric bypass or caloric restriction on oxidative stress and genomic damage in obese Zucker rats. Free Radic. Biol. Med. 2016, 94, 208–217. [Google Scholar] [CrossRef]
- Meirelles, K.; Ahmed, T.; Culnan, D.M.; Lynch, C.J.; Lang, C.H.; Cooney, R.N. Mechanisms of glucose homeostasis after Roux-en-Y gastric bypass surgery in the obese, insulin-resistant Zucker rat. Ann. Surg. 2009, 249, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Seyfried, F.; Miras, A.D.; Rotzinger, L.; Nordbeck, A.; Corteville, C.; Li, J.V.; Schlegel, N.; Hankir, M.; Fenske, W.; Otto, C.; et al. Gastric Bypass-Related Effects on Glucose Control, beta Cell Function and Morphology in the Obese Zucker Rat. Obes. Surg. 2016, 26, 1228–1236. [Google Scholar] [CrossRef]
- Wolff, B.S.; Meirelles, K.; Meng, Q.; Pan, M.; Cooney, R.N. Roux-en-Y gastric bypass alters small intestine glutamine transport in the obese Zucker rat. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G594–G601. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Ohinata, K.; Meguid, M.M.; Marx, W.; Tada, T.; Chen, C.; Quinn, R.; Inui, A. Gastric bypass model in the obese rat to study metabolic mechanisms of weight loss. J. Surg. Res. 2002, 107, 56–63. [Google Scholar] [CrossRef]
- Clark, J.B.; Palmer, C.J.; Shaw, W.N. The diabetic Zucker fatty rat. Proc. Soc. Exp. Biol. Med. 1983, 173, 68–75. [Google Scholar] [CrossRef]
- Abegg, K.; Corteville, C.; Docherty, N.G.; Boza, C.; Lutz, T.A.; Munoz, R.; le Roux, C.W. Effect of bariatric surgery combined with medical therapy versus intensive medical therapy or calorie restriction and weight loss on glycemic control in Zucker diabetic fatty rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R321–R329. [Google Scholar] [CrossRef] [Green Version]
- Bhutta, H.Y.; Deelman, T.E.; le Roux, C.W.; Ashley, S.W.; Rhoads, D.B.; Tavakkoli, A. Intestinal sweet-sensing pathways and metabolic changes after Roux-en-Y gastric bypass surgery. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G588–G593. [Google Scholar] [CrossRef] [Green Version]
- Bhutta, H.Y.; Rajpal, N.; White, W.; Freudenberg, J.M.; Liu, Y.; Way, J.; Rajpal, D.; Cooper, D.C.; Young, A.; Tavakkoli, A.; et al. Effect of Roux-en-Y gastric bypass surgery on bile acid metabolism in normal and obese diabetic rats. PLoS ONE 2015, 10, e0122273. [Google Scholar] [CrossRef]
- Canney, A.L.; Cohen, R.V.; Elliott, J.A.; Aboud, C.M.; Martin, W.P.; Docherty, N.G.; le Roux, C.W. Improvements in diabetic albuminuria and podocyte differentiation following Roux-en-Y gastric bypass surgery. Diabetes Vasc. Dis. Res. 2020, 17, 1479164119879039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Liu, C.Q.; Liu, G.P.; Huang, Z.P.; Zou, D.J. Roux-en-Y gastric bypass decreases endotoxemia and inflammatory stress in association with improvements in gut permeability in obese diabetic rats. J. Diabetes 2019, 11, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Liu, C.Q.; Shan, C.X.; Chen, Y.; Li, H.H.; Huang, Z.P.; Zou, D.J. Gut microbiota after Roux-en-Y gastric bypass and sleeve gastrectomy in a diabetic rat model: Increased diversity and associations of discriminant genera with metabolic changes. Diabetes Metab. Res. Rev. 2017, 33. [Google Scholar] [CrossRef]
- Huang, H.; Aminian, A.; Hassan, M.; Dan, O.; Axelrod, C.L.; Schauer, P.R.; Brethauer, S.A.; Kirwan, J.P. Gastric Bypass Surgery Improves the Skeletal Muscle Ceramide/S1P Ratio and Upregulates the AMPK/ SIRT1/ PGC-1alpha Pathway in Zucker Diabetic Fatty Rats. Obes. Surg. 2019, 29, 2158–2165. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Lau, R.; Hall, C.; Palaia, T.; Brathwaite, C.E.; Ragolia, L. Bile acid elevation after Roux-en-Y gastric bypass is associated with cardio-protective effect in Zucker Diabetic Fatty rats. Int. J. Surg. 2015, 24, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Mocanu, A.O.; Mulya, A.; Huang, H.; Dan, O.; Schauer, P.R.; Dinischiotu, A.; Brethauer, S.A.; Kirwan, J.P. Effect of Roux-en-Y Gastric Bypass on the NLRP3 Inflammasome in Pancreatic Islets from Zucker Diabetic Fatty Rats. Obes. Surg. 2016, 26, 3076–3081. [Google Scholar] [CrossRef] [Green Version]
- Mosinski, J.D.; Aminian, A.; Axelrod, C.L.; Batayyah, E.; Talamas, H.R.; Daigle, C.; Mulya, A.; Scelsi, A.R.; Schauer, P.R.; Brethauer, S.; et al. Roux-en-Y Gastric Bypass Restores Islet Function and Morphology Independent of Body Weight in ZDF Rats. Am. J. Physiol. Endocrinol. Metab. 2021. [Google Scholar] [CrossRef]
- Nair, M.; Martin, W.P.; Zhernovkov, V.; Elliott, J.A.; Fearon, N.; Eckhardt, H.; McCormack, J.; Godson, C.; Brennan, E.P.; Fandriks, L.; et al. Characterization of the renal cortical transcriptome following Roux-en-Y gastric bypass surgery in experimental diabetic kidney disease. BMJ Open Diabetes Res. Care 2020, 8, e001113. [Google Scholar] [CrossRef]
- Pal, A.; Rhoads, D.B.; Tavakkoli, A. Effect of Portal Glucose Sensing on Systemic Glucose Levels in SD and ZDF Rats. PLoS ONE 2016, 11, e0165592. [Google Scholar] [CrossRef]
- Pal, A.; Rhoads, D.B.; Tavakkoli, A. Portal milieu and the interplay of multiple antidiabetic effects after gastric bypass surgery. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G668–G678. [Google Scholar] [CrossRef]
- Shimizu, H.; Eldar, S.; Heneghan, H.M.; Schauer, P.R.; Kirwan, J.P.; Brethauer, S.A. The effect of selective gut stimulation on glucose metabolism after gastric bypass in the Zucker diabetic fatty rat model. Surg. Obes. Relat. Dis. 2014, 10, 29–35. [Google Scholar] [CrossRef]
- Vangoitsenhoven, R.; Mulya, A.; Mosinski, J.D.; Brethauer, S.A.; Schauer, P.R.; Kirwan, J.P.; Aminian, A. Effects of gastric bypass surgery on expression of glucose transporters and fibrotic biomarkers in kidney of diabetic fatty rats. Surg. Obes. Relat. Dis. 2020, 16, 1242–1248. [Google Scholar] [CrossRef]
- Seyfried, F.; Bueter, M.; Spliethoff, K.; Miras, A.D.; Abegg, K.; Lutz, T.A.; le Roux, C.W. Roux-en Y gastric bypass is superior to duodeno-jejunal bypass in improving glycaemic control in Zucker diabetic fatty rats. Obes. Surg. 2014, 24, 1888–1895. [Google Scholar] [CrossRef] [PubMed]
- Dischinger, U.; Corteville, C.; Otto, C.; Fassnacht, M.; Seyfried, F.; Hankir, M.K. GLP-1 and PYY3-36 reduce high-fat food preference additively after Roux-en-Y gastric bypass in diet-induced obese rats. Surg. Obes. Relat. Dis. 2019, 15, 1483–1492. [Google Scholar] [CrossRef] [PubMed]
- Seyfried, F.; Miras, A.D.; Bueter, M.; Prechtl, C.G.; Spector, A.C.; le Roux, C.W. Effects of preoperative exposure to a high-fat versus a low-fat diet on ingestive behavior after gastric bypass surgery in rats. Surg. Endosc. 2013, 27, 4192–4201. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; DeFronzo, R.A. Insulin sensitivity indices obtained from oral glucose tolerance testing: Comparison with the euglycemic insulin clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef]
- Miras, A.D.; Seyfried, F.; Phinikaridou, A.; Andia, M.E.; Christakis, I.; Spector, A.C.; Botnar, R.M.; le Roux, C.W. Rats fed diets with different energy contribution from fat do not differ in adiposity. Obes. Facts 2014, 7, 302–310. [Google Scholar] [CrossRef]
- Camastra, S.; Muscelli, E.; Gastaldelli, A.; Holst, J.J.; Astiarraga, B.; Baldi, S.; Nannipieri, M.; Ciociaro, D.; Anselmino, M.; Mari, A.; et al. Long-term effects of bariatric surgery on meal disposal and beta-cell function in diabetic and nondiabetic patients. Diabetes 2013, 62, 3709–3717. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, S.H.; Bojsen-Moller, K.N.; Dirksen, C.; Jorgensen, N.B.; Clausen, T.R.; Wulff, B.S.; Kristiansen, V.B.; Worm, D.; Hansen, D.L.; Holst, J.J.; et al. Effects of gastric bypass surgery on glucose absorption and metabolism during a mixed meal in glucose-tolerant individuals. Diabetologia 2013, 56, 2250–2254. [Google Scholar] [CrossRef] [Green Version]
- Cusin, I.; Rohner-Jeanrenaud, F.; Stricker-Krongrad, A.; Jeanrenaud, B. The weight-reducing effect of an intracerebroventricular bolus injection of leptin in genetically obese fa/fa rats. Reduced sensitivity compared with lean animals. Diabetes 1996, 45, 1446–1450. [Google Scholar] [CrossRef] [Green Version]
- Dryden, S.; King, P.; Pickavance, L.; Doyle, P.; Williams, G. Divergent effects of intracerebroventricular and peripheral leptin administration on feeding and hypothalamic neuropeptide Y in lean and obese (fa/fa) Zucker rats. Clin. Sci. 1999, 96, 307–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyfried, F.; le Roux, C.W.; Bueter, M. Lessons learned from gastric bypass operations in rats. Obes. Facts 2011, 4 (Suppl. S1), 3–12. [Google Scholar] [CrossRef] [Green Version]
- Griffen, S.C.; Wang, J.; German, M.S. A genetic defect in beta-cell gene expression segregates independently from the fa locus in the ZDF rat. Diabetes 2001, 50, 63–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Dalbeni, A. Treatments for NAFLD: State of Art. Int. J. Mol. Sci. 2021, 22, 2350. [Google Scholar] [CrossRef]
- Fakhry, T.K.; Mhaskar, R.; Schwitalla, T.; Muradova, E.; Gonzalvo, J.P.; Murr, M.M. Bariatric surgery improves nonalcoholic fatty liver disease: A contemporary systematic review and meta-analysis. Surg. Obes. Relat. Dis. 2019, 15, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Una, M.; Lopez-Mancheno, Y.; Dieguez, C.; Fernandez-Rojo, M.A.; Novelle, M.G. Unraveling the Role of Leptin in Liver Function and Its Relationship with Liver Diseases. Int. J. Mol. Sci. 2020, 21, 9368. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Hao, Z.; Mumphrey, M.B.; Townsend, R.L.; Patterson, L.M.; Stylopoulos, N.; Munzberg, H.; Morrison, C.D.; Drucker, D.J.; Berthoud, H.R. GLP-1 receptor signaling is not required for reduced body weight after RYGB in rodents. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R352–R362. [Google Scholar] [CrossRef] [Green Version]
- Mokadem, M.; Zechner, J.F.; Margolskee, R.F.; Drucker, D.J.; Aguirre, V. Effects of Roux-en-Y gastric bypass on energy and glucose homeostasis are preserved in two mouse models of functional glucagon-like peptide-1 deficiency. Mol. Metab. 2014, 3, 191–201. [Google Scholar] [CrossRef]
- Carmody, J.S.; Munoz, R.; Yin, H.; Kaplan, L.M. Peripheral, but not central, GLP-1 receptor signaling is required for improvement in glucose tolerance after Roux-en-Y gastric bypass in mice. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E855–E861. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Zou, J.; Li, S.; Guo, J.; Shi, W.; Wang, B.; Han, X.; Zhang, H.; Zhang, P.; Miao, Z.; et al. Farnesoid X receptor contributes to body weight-independent improvements in glycemic control after Roux-en-Y gastric bypass surgery in diet-induced obese mice. Mol. Metab. 2020, 37, 100980. [Google Scholar] [CrossRef]
- Shin, A.C.; Zheng, H.; Townsend, R.L.; Sigalet, D.L.; Berthoud, H.R. Meal-induced hormone responses in a rat model of Roux-en-Y gastric bypass surgery. Endocrinology 2010, 151, 1588–1597. [Google Scholar] [CrossRef] [Green Version]
- Svane, M.S.; Jorgensen, N.B.; Bojsen-Moller, K.N.; Dirksen, C.; Nielsen, S.; Kristiansen, V.B.; Torang, S.; Wewer Albrechtsen, N.J.; Rehfeld, J.F.; Hartmann, B.; et al. Peptide YY and glucagon-like peptide-1 contribute to decreased food intake after Roux-en-Y gastric bypass surgery. Int. J. Obes. 2016, 40, 1699–1706. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hankir, M.K.; Rotzinger, L.; Nordbeck, A.; Corteville, C.; Dischinger, U.; Knop, J.-L.; Hoffmann, A.; Otto, C.; Seyfried, F. Leptin Receptors Are Not Required for Roux-en-Y Gastric Bypass Surgery to Normalize Energy and Glucose Homeostasis in Rats. Nutrients 2021, 13, 1544. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13051544
Hankir MK, Rotzinger L, Nordbeck A, Corteville C, Dischinger U, Knop J-L, Hoffmann A, Otto C, Seyfried F. Leptin Receptors Are Not Required for Roux-en-Y Gastric Bypass Surgery to Normalize Energy and Glucose Homeostasis in Rats. Nutrients. 2021; 13(5):1544. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13051544
Chicago/Turabian StyleHankir, Mohammed K., Laura Rotzinger, Arno Nordbeck, Caroline Corteville, Ulrich Dischinger, Juna-Lisa Knop, Annett Hoffmann, Christoph Otto, and Florian Seyfried. 2021. "Leptin Receptors Are Not Required for Roux-en-Y Gastric Bypass Surgery to Normalize Energy and Glucose Homeostasis in Rats" Nutrients 13, no. 5: 1544. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13051544