
Microbiota and Metabolite Modifications after Dietary Exclusion of Dairy Products and Reduced Consumption of Fermented Food in Young and Older Men

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
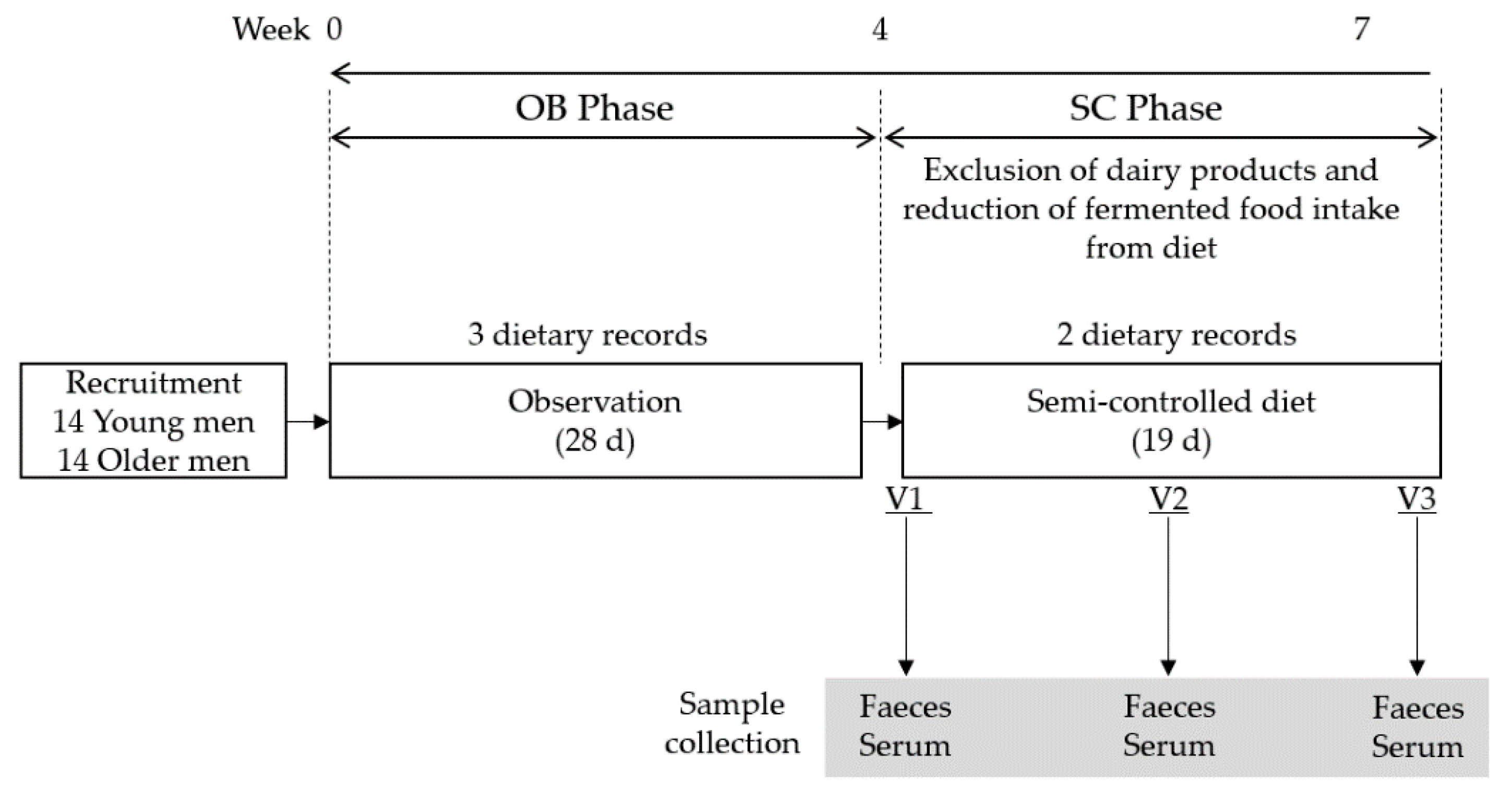
2.1. Study Design
2.2. Study Population
2.3. Dietary Assessment
2.4. Serum Analyses
2.4.1. Biochemical Analyses
2.4.2. Gas Chromatography Analyses
2.5. Microbiota Analyses
2.6. Statistical Analysis
2.6.1. Statistical Analyses on Dietary and Circulating Markers
2.6.2. Statistical Analyses on Microbiota Data
3. Results
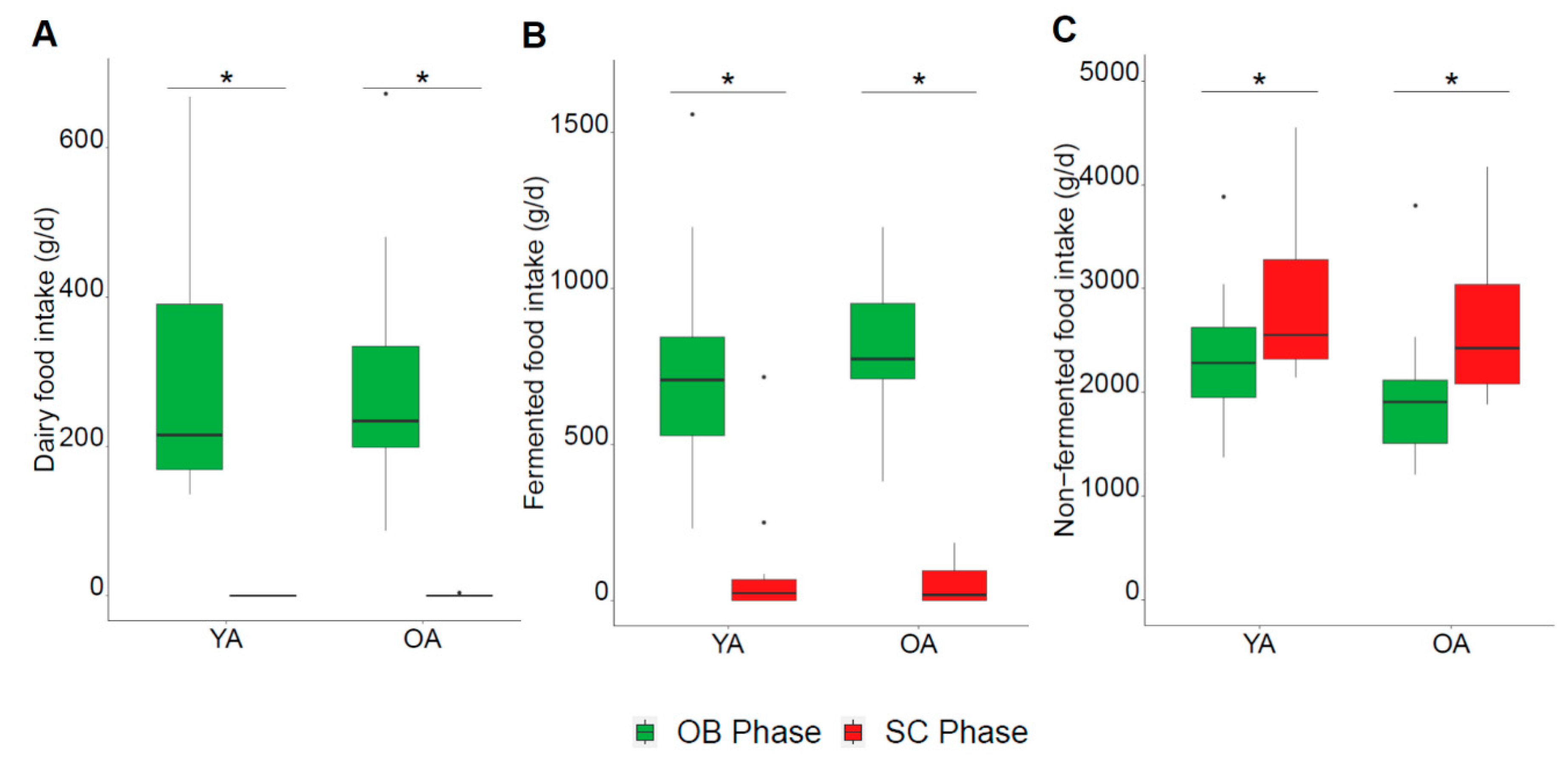
3.1. Dietary Characteristics
3.2. Biochemical Parameters and Serum Metabolites
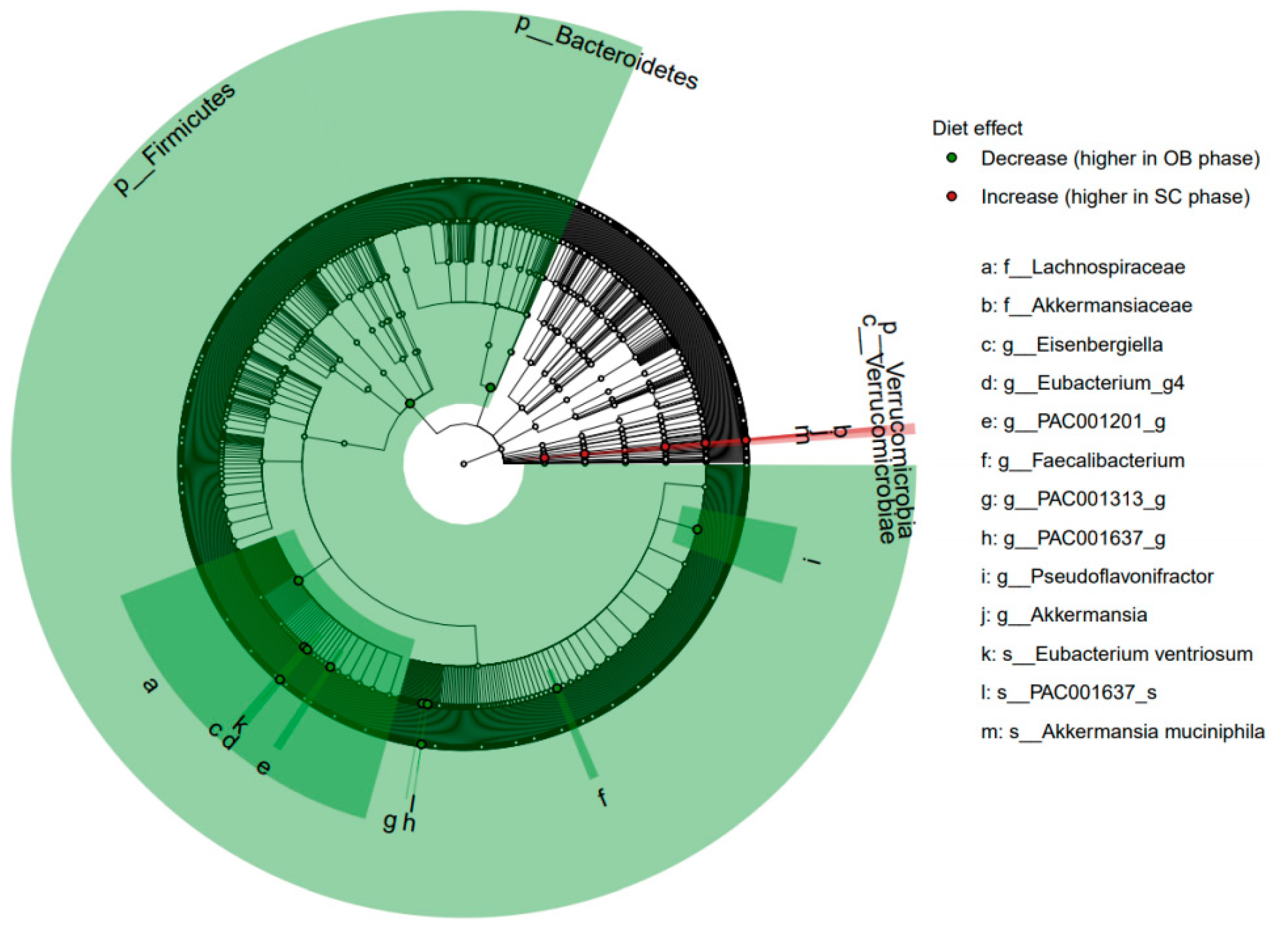
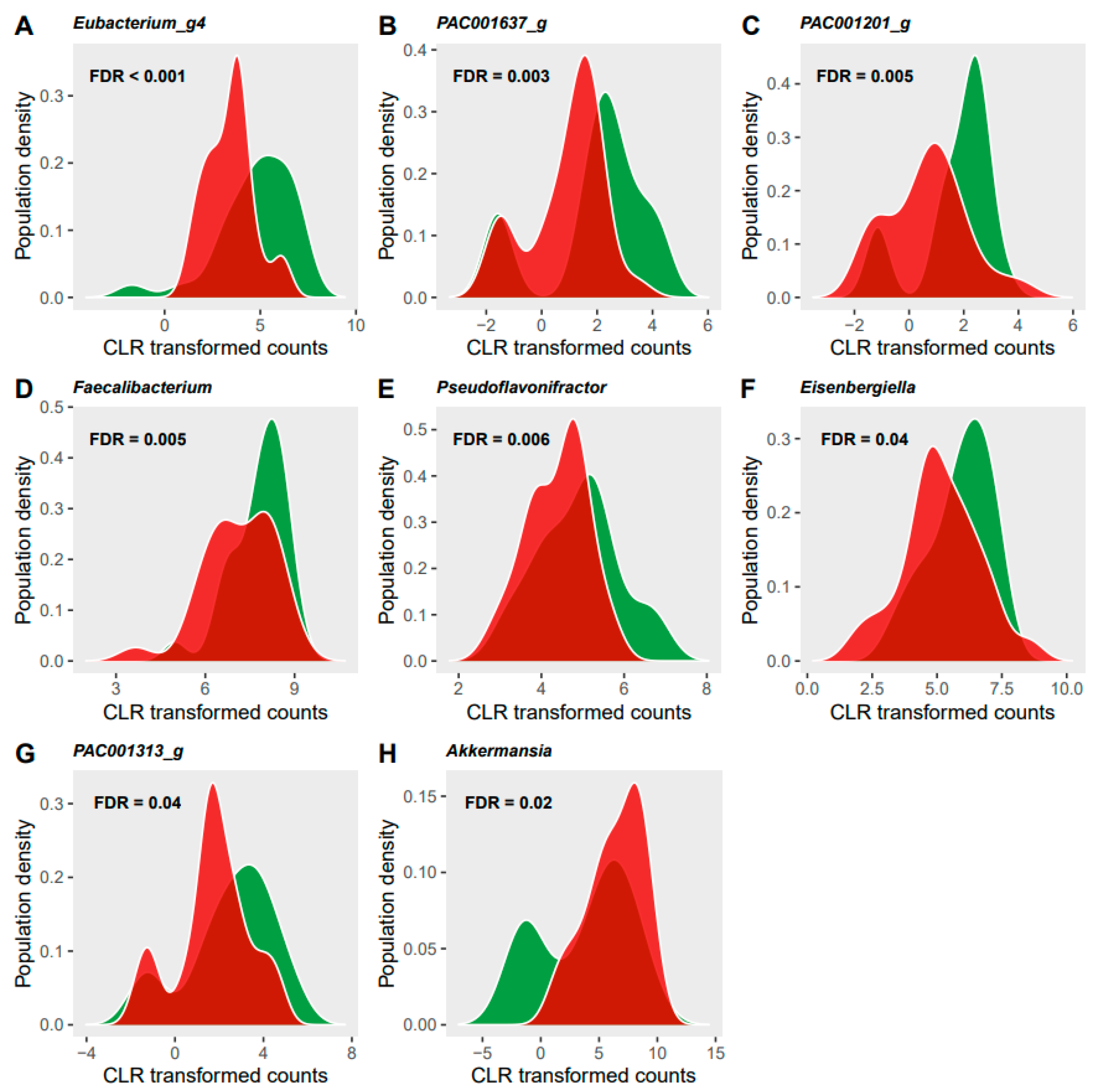
3.3. Microbiome Analyses
4. Discussion
4.1. Serum Metabolites Modification by Exclusion of Dairy and Limitation of Fermented Foods
4.2. Gut Microbiota Modifications and Its Potential Associations with Diet and Circulating Metabolites
4.3. Different Modifications between Young and Older Adults in Response to the Restriction on Dairy and Fermented Food Intake
4.4. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nat. Cell Biol. 2018, 555, 210–215. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Statovci, D.; Aguilera, M.; MacSharry, J.; Melgar, S. The Impact of Western Diet and Nutrients on the Microbiota and Immune Response at Mucosal Interfaces. Front. Immunol. 2017, 8, 838. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 69, 1218–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, V.; Ferrão, J.; Pimentel, L.; Pintado, M.; Fernandes, T. One Health, Fermented Foods, and Gut Microbiota. Foods 2018, 7, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiemsma, L.T.; Nakamura, R.; Nguyen, J.G.; Michels, K.B. Does Consumption of Fermented Foods Modify the Human Gut Microbiota? J. Nutr. 2020, 150, 1680–1692. [Google Scholar] [CrossRef]
- Charalampopoulos, D.; Rastall, R. Prebiotics in foods. Curr. Opin. Biotechnol. 2012, 23, 187–191. [Google Scholar] [CrossRef]
- Peng, M.; Tabashsum, Z.; Anderson, M.; Truong, A.; Houser, A.K.; Padilla, J.; Akmel, A.; Bhatti, J.; Rahaman, S.O.; Biswas, D. Effectiveness of probiotics, prebiotics, and prebiotic-like components in common functional foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1908–1933. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, A.M.; Barile, D. Bovine Milk as a Source of Functional Oligosaccharides for Improving Human Health. Adv. Nutr. 2011, 2, 284–289. [Google Scholar] [CrossRef]
- Chen, X.Y.; Gänzle, M.G. Lactose and lactose-derived oligosaccharides: More than prebiotics? Int. Dairy J. 2017, 67, 61–72. [Google Scholar] [CrossRef]
- Lukito, W.; Malik, S.G.; Surono, I.S.; Wahlqvist, M.L. From ’lactose intolerance’ to ’lactose nutrition’. Asia Pac. J. Clin. Nutr. 2015, 24, 1–8. [Google Scholar]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented Foods: Definitions and Characteristics, Impact on the Gut Microbiota and Effects on Gastrointestinal Health and Disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Khorraminezhad, L.; Leclercq, M.; O’Connor, S.; Julien, P.; Weisnagel, S.J.; Gagnon, C.; Droit, A.; Rudkowska, I. Dairy product intake modifies gut microbiota composition among hyperinsulinemic individuals. Eur. J. Nutr. 2021, 60, 159–167. [Google Scholar] [CrossRef]
- Volokh, O.; Klimenko, N.; Berezhnaya, Y.; Tyakht, A.; Nesterova, P.; Popenko, A.; Alexeev, D. Human Gut Microbiome Re-sponse Induced by Fermented Dairy Product Intake in Healthy Volunteers. Nutrients 2019, 11, 547. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Solares, S.; Fernandez-Navarro, T.; Arboleya, S.; Reyes-Gavilán, C.G.D.L.; Salazar, N.; Gueimonde, M. Fermented Dairy Foods: Impact on Intestinal Microbiota and Health-Linked Biomarkers. Front. Microbiol. 2019, 10, 1046. [Google Scholar] [CrossRef]
- Aslam, H.; Marx, W.; Rocks, T.; Loughman, A.; Chandrasekaran, V.; Ruusunen, A.; Dawson, S.L.; West, M.; Mullarkey, E.; Pasco, J.A.; et al. The effects of dairy and dairy derivatives on the gut microbiota: A systematic literature review. Gut Microbes 2020, 12, 1799533. [Google Scholar] [CrossRef]
- Jung, M.; Bourne, J.; Buchholz, A.; Ginis, K.A.M. Strategies for public health initiatives targeting dairy consumption in young children: A qualitative formative investigation of parent perceptions. Public Health Nutr. 2017, 20, 2893–2908. [Google Scholar] [CrossRef] [Green Version]
- Iuliano, S.; Poon, S.; Wang, X.; Bui, M.; Seeman, E. Dairy food supplementation may reduce malnutrition risk in institutionalised elderly. Br. J. Nutr. 2017, 117, 142–147. [Google Scholar] [CrossRef] [Green Version]
- Cuesta-Triana, F.; Verdejo-Bravo, C.; Fernández-Pérez, C.; Martín-Sánchez, F.J. Effect of Milk and Other Dairy Products on the Risk of Frailty, Sarcopenia, and Cognitive Performance Decline in the Elderly: A Systematic Review. Adv. Nutr. 2019, 10, S105–S119. [Google Scholar] [CrossRef]
- Rémond, D.; Shahar, D.R.; Gille, D.; Pinto, P.; Kachal, J.; Peyron, M.-A.; Dos Santos, C.N.; Walther, B.; Bordoni, A.; Dupont, D.; et al. Understanding the gastrointestinal tract of the elderly to develop dietary solutions that prevent malnutrition. Oncotarget 2015, 6, 13858–13898. [Google Scholar] [CrossRef] [Green Version]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.B.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef]
- Maynard, C.; Weinkove, D. The Gut Microbiota and Ageing. Subcell Biochem. 2018, 90, 351–371. [Google Scholar]
- Quercia, S.; Candela, M.; Giuliani, C.; Turroni, S.; Luiselli, D.; Rampelli, S.; Brigidi, P.; Franceschi, C.; Bacalini, M.G.; Garagnani, P.; et al. From lifetime to evolution: Timescales of human gut microbiota adaptation. Front. Microbiol. 2014, 5, 587. [Google Scholar] [CrossRef] [Green Version]
- Durack, J.; Lynch, S.V. The gut microbiome: Relationships with disease and opportunities for therapy. J. Exp. Med. 2018, 216, 20–40. [Google Scholar] [CrossRef] [Green Version]
- Li, K.J.; Brouwer-Brolsma, E.M.; Burton, K.J.; Vergères, G.; Feskens, E.J.M. Prevalence of fermented foods in the Dutch adult diet and validation of a food frequency questionnaire for estimating their intake in the NQplus cohort. BMC Nutr. 2020, 6, 69. [Google Scholar] [CrossRef]
- Trimigno, A.; Münger, L.; Picone, G.; Freiburghaus, C.; Pimentel, G.; Vionnet, N.; Pralong, F.; Capozzi, F.; Badertscher, R.; Vergères, G. GC-MS Based Metabolomics and NMR Spectroscopy Investigation of Food Intake Biomarkers for Milk and Cheese in Serum of Healthy Humans. Metabolites 2018, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Münger, L.H.; Trimigno, A.; Picone, G.; Freiburghaus, C.; Pimentel, G.; Burton, K.J.; Pralong, F.P.; Vionnet, N.; Capozzi, F.; Badertscher, R.; et al. Identification of Urinary Food Intake Biomarkers for Milk, Cheese, and Soy-Based Drink by Untargeted GC-MS and NMR in Healthy Humans. J. Proteome Res. 2017, 16, 3321–3335. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Collomb, M.; Bühler, T. Analyse de la Composition en Acides Gras de la Graisse de Lait. Mitteilungen aus Lebensmitteluntersuchung und Hygiene 2000, 91, 306–332. [Google Scholar]
- Burton, K.J.; Rosikiewicz, M.; Pimentel, G.; Bütikofer, U.; Von Ah, U.; Voirol, M.-J.; Croxatto, A.; Aeby, S.; Drai, J.; McTernan, P.G.; et al. Probiotic yogurt and acidified milk similarly reduce postprandial inflammation and both alter the gut microbiota of healthy, young men. Br. J. Nutr. 2017, 117, 1312–1322. [Google Scholar] [CrossRef] [Green Version]
- Köster, J.; Rahmann, S. Snakemake—A scalable bioinformatics workflow engine. Bioinformatics 2018, 34, 3600. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet 2011, 17, 3. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasolli, E.; De Filippis, F.; Mauriello, I.E.; Cumbo, F.; Walsh, A.M.; Leech, J.; Cotter, P.D.; Segata, N.; Ercolini, D. Large-scale genome-wide analysis links lactic acid bacteria from food with the gut microbiome. Nat. Commun. 2020, 11, 2610. [Google Scholar] [CrossRef]
- Gardner, S.N.; Slezak, T. Simulate_PCR for amplicon prediction and annotation from multiplex, degenerate primers and probes. BMC Bioinform. 2014, 15, 237. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: AnRPackage for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.7; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Noguchi, Y.R.G.K.; Brunner, E.; Konietschke, F. nparLD: An R Software Package for the Nonparametric Analysis of Longitudinal Data in Factorial Experiments. J. Stat. Softw. 2012, 50. [Google Scholar] [CrossRef] [Green Version]
- Willett, W. Nutritional Epidemiology; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0.12. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 1 November 2020).
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, A.M.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 November 2020).
- Fernandes, A.D.; Reid, J.N.; Macklaim, J.M.; McMurrough, T.; Edgell, D.R.; Gloor, G.B. Unifying the analysis of high-throughput sequencing datasets: Characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome 2014, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.D.; Macklaim, J.M.; Linn, T.G.; Reid, G.; Gloor, G.B. ANOVA-Like Differential Expression (ALDEx) Analysis for Mixed Population RNA-Seq. PLoS ONE 2013, 8, e67019. [Google Scholar] [CrossRef]
- Gloor, G.B.; Macklaim, J.M.; Fernandes, A.D. Displaying Variation in Large Datasets: Plotting a Visual Summary of Effect Sizes. J. Comput. Graph. Stat. 2016, 25, 971–979. [Google Scholar] [CrossRef]
- Yu, G. Using ggtree to Visualize Data on Tree-Like Structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef]
- Yu, G.; Lam, T.T.-Y.; Zhu, H.; Guan, Y. Two Methods for Mapping and Visualizing Associated Data on Phylogeny Using Ggtree. Mol. Biol. Evol. 2018, 35, 3041–3043. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Xu, S.; Yu, G. MicrobiotaProcess: An R Package for Analysis, Visualization and Biomarker Discovery of Microbiome. Available online: https://github.com/YuLab-SMU/MicrobiotaProcess/ (accessed on 1 November 2020).
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression pro-files. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anses. Actualisation des Repères du PNNS: Révision des Repères de Consommations Alimentaires; Anses: Maisons-Alfort, France, 2016. [Google Scholar]
- Claesson, M.J.; Cusack, S.; O’Sullivan, O.; Greene-Diniz, R.; De Weerd, H.; Flannery, E.; Marchesi, J.R.; Falush, D.; Dinan, T.G.; Fitzgerald, G.F.; et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc. Natl. Acad. Sci. USA 2011, 108, 4586–4591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venn-Watson, S.; Lumpkin, R.; Dennis, E.A. Efficacy of dietary odd-chain saturated fatty acid pentadecanoic acid parallels broad associated health benefits in humans: Could it be essential? Sci. Rep. 2020, 10, 8161. [Google Scholar] [CrossRef]
- Taormina, V.M.; Unger, A.L.; Schiksnis, M.R.; Torres-Gonzalez, M.; Kraft, J. Branched-Chain Fatty Acids—An Underexplored Class of Dairy-Derived Fatty Acids. Nutrients 2020, 12, 2875. [Google Scholar] [CrossRef]
- Chartrand, D.; Da Silva, M.S.; Julien, P.; Rudkowska, I. Influence of Amino Acids in Dairy Products on Glucose Homeostasis: The Clinical Evidence. Can. J. Diabetes 2017, 41, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, G.; Burnand, D.; Munger, L.H.; Pralong, F.P.; Vionnet, N.; Portmann, R.; Vergeres, G. Identification of Milk and Cheese Intake Biomarkers in Healthy Adults Reveals High Interindividual Variability of Lewis System-Related Oligosaccharides. J. Nutr. 2020, 150, 1058–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anses. Étude Individuelle Nationale des Consommations Alimentaires 3; Anses: Maisons-Alfort, France, 2017. [Google Scholar]
- Pfeuffer, M.; Jaudszus, A. Pentadecanoic and Heptadecanoic Acids: Multifaceted Odd-Chain Fatty Acids. Adv. Nutr. 2016, 7, 730–734. [Google Scholar] [CrossRef] [Green Version]
- Pranger, I.G.; Corpeleijn, E.; Muskiet, F.A.J.; Kema, I.P.; Singh-Povel, C.; Bakker, S.J.L. Circulating fatty acids as biomarkers of dairy fat intake: Data from the lifelines biobank and cohort study. Biomarkers 2019, 24, 360–372. [Google Scholar] [CrossRef]
- Soerensen, K.V.; Thorning, T.K.; Astrup, A.; Kristensen, M.; Lorenzen, J.K. Effect of dairy calcium from cheese and milk on fecal fat excretion, blood lipids, and appetite in young men. Am. J. Clin. Nutr. 2014, 99, 984–991. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Poon, S.; Seeman, E.; Hare, D.L.; Bui, M.; Iuliano, S. Fat from dairy foods and ’meat’ consumed within recommended levels is associated with favourable serum cholesterol levels in institutionalised older adults. J. Nutr. Sci. 2019, 8, e10. [Google Scholar] [CrossRef] [Green Version]
- Lordan, R.; Tsoupras, A.; Mitra, B.; Zabetakis, I. Dairy Fats and Cardiovascular Disease: Do We Really Need to Be Concerned? Foods 2018, 7, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prodhan, U.K.; Milan, A.M.; Thorstensen, E.B.; Barnett, M.P.G.; Stewart, R.A.H.; Benatar, J.R.; Cameron-Smith, D. Altered Dairy Protein Intake Does Not Alter Circulatory Branched Chain Amino Acids in Healthy Adults: A Randomized Controlled Trial. Nutrients 2018, 10, 1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vionnet, N.; Münger, L.H.; Freiburghaus, C.; Burton, K.J.; Pimentel, G.; Pralong, F.P.; Badertscher, R.; Vergères, G. Assessment of lactase activity in humans by measurement of galactitol and galactonate in serum and urine after milk intake. Am. J. Clin. Nutr. 2019, 109, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Afandi, F.; Wijaya, C.; Faridah, D.; Suyatma, N.; Jayanegara, A. Evaluation of Various Starchy Foods: A Systematic Review and Meta-Analysis on Chemical Properties Affecting the Glycemic Index Values Based on In Vitro and In Vivo Experiments. Foods 2021, 10, 364. [Google Scholar] [CrossRef] [PubMed]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized Nutrition by Prediction of Glycemic Responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphilaand improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2015, 65, 426–436. [Google Scholar] [CrossRef] [Green Version]
- Naito, Y.; Uchiyama, K.; Takagi, T. A next-generation beneficial microbe: Akkermansia muciniphila. J. Clin. Biochem. Nutr. 2018, 63, 33–35. [Google Scholar] [CrossRef] [Green Version]
- Greer, R.L.; Dong, X.; Moraes, A.C.; Zielke, R.A.; Fernandes, G.R.; Peremyslova, E.; Vasquez-Perez, S.; Schoenborn, A.A.; Gomes, E.P.; Pereira, A.C.; et al. Akkermansia muciniphila mediates negative effects of IFNgamma on glucose metabolism. Nat. Commun. 2016, 7, 13329. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Zhang, S.; Wu, J.; Ye, T.; Wang, S.; Wang, P.; Xing, D. Butyrate-producing bacteria and the gut-heart axis in ath-erosclerosis. Clin. Chim. Acta 2020, 507, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Belenguer, A.; Holtrop, G.; Johnstone, A.M.; Flint, H.J.; Lobley, G.E. Reduced Dietary Intake of Carbohydrates by Obese Subjects Results in Decreased Concentrations of Butyrate and Butyrate-Producing Bacteria in Feces. Appl. Environ. Microbiol. 2006, 73, 1073–1078. [Google Scholar] [CrossRef] [Green Version]
- Crowley, E.; Long-Smith, C.; Murphy, A.; Patterson, E.; Murphy, K.; O’Gorman, D.; Stanton, C.; Nolan, Y. Dietary Supple-mentation with a Magnesium-Rich Marine Mineral Blend Enhances the Diversity of Gastrointestinal Microbiota. Marine Drugs 2018, 16, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantino, S.; Paneni, F.; Cosentino, F. Ageing, metabolism and cardiovascular disease. J. Physiol. 2016, 594, 2061–2073. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, A.N.; Segal, S. The metabolic fate of [i−14C]galactitol in mammalian tissue. Biochim. Biophys. Acta Gen. Subj. 1968, 156, 9–16. [Google Scholar] [CrossRef]
- Musso, C.G.; Oreopoulos, D.G. Aging and Physiological Changes of the Kidneys Including Changes in Glomerular Filtration Rate. Nephron 2011, 119, 1–5. [Google Scholar] [CrossRef]
- Schadewaldt, P.; Killius, S.; Kamalanathan, L.; Hammen, H.-W.; Strasburger, K.; Wendel, U. Renal excretion of galactose and galactitol in patients with classical galactosaemia, obligate heterozygous parents and healthy subjects. J. Inherit. Metab. Dis. 2003, 26, 459–479. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, M.; Mazur, A.; Berry, G.T.; Ning, C.; Wehrli, S.; Yager, C.; Reynolds, R.; Singh, R.; Muralidharan, K.; Langley, S.; et al. Urine and plasma galactitol in patients with galactose-1-phosphate uridyltransferase deficiency galactosemia. Metabolism 1999, 48, 1294–1302. [Google Scholar] [CrossRef]
- Liu, H.-H.; Li, J.-J. Aging and dyslipidemia: A review of potential mechanisms. Ageing Res. Rev. 2015, 19, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.; Saunier, K.; Hanisch, C.; Norin, E.; Alm, L.; Midtvedt, T.; Cresci, A.; Silvi, S.; Orpianesi, C.; Verdenelli, M.C.; et al. Differences in Fecal Microbiota in Different European Study Populations in Relation to Age, Gender, and Country: A Cross-Sectional Study. Appl. Environ. Microbiol. 2006, 72, 1027–1033. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, P.W.; Jeffery, I.B. Gut microbiota and aging. Science 2015, 350, 1214–1215. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YA | OA | p-Value (Wilcoxon 1) | p-Value (Wald Test) a | |||||
|---|---|---|---|---|---|---|---|---|
| OB Phase | SC Phase | OB Phase | SC Phase | Baseline OA vs. YA | Age Effect | Diet Effect | Interaction | |
| Dairy food subgroups | ||||||||
| Cheese | 42.7 (34.0, 67.3) | 0.0 (0.0, 0.0) * | 51.8 (44.7, 73.0) | 0.0 (0.0, 0.0) * | 0.220 | 0.195 | <0.001 | 0.195 |
| Cream | 6.7 (0.5, 17.2) | 0.0 (0.0, 0.0) * | 11.8 (4.7, 17.8) | 0.0 (0.0, 0.0) * | 0.394 | 0.231 | <0.001 | 0.231 |
| Dairy fats | 5.0 (1.2, 7.9) | 0.0 (0.0, 0.0) * | 8.1 (2.6, 18.2) | 0.0 (0.0, 0.0) * | 0.126 | 0.097 | <0.001 | 0.041 |
| Fresh fermented dairy products | 102.2 (60.1, 161.5) | 0.0 (0.0, 0.0) * | 111.1 (63.8, 133.3) | 0.0 (0.0, 0.0) * | 1.000 | 0.785 | <0.001 | 0.785 |
| Milk | 78.7 (4.3, 122.1) | 0.0 (0.0, 0.0) * | 88.9 (16.1, 127.9) | 0.0 (0.0, 0.0) * | 0.827 | 0.682 | <0.001 | 0.868 |
| Non-dairy fermented foods subgroups | ||||||||
| Alcohol | 185.2 (39.9, 259.7) | 0.0 (0.0, 0.0) * | 147.8 (61.1, 339.4) | 0.0 (0.0, 0.0) * | 0.903 | 0.989 | <0.001 | 0.900 |
| Bread products | 87.8 (55.4, 124.8) | 0.0 (0.0, 0.0) * | 167.4 (129.4, 197.0) # | 0.0 (0.0, 3.3) * | 0.002 | 0.001 | <0.001 | 0.048 |
| Cake and pastries | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.0) * | 0.0 (0.0, 5.6) | 0.0 (0.0, 17.7) # | 0.777 | 0.027 | 0.887 | 0.022 |
| Chocolate products | 2.2 (0.3, 7.4) | 0.0 (0.0, 0.0) * | 6.1 (4.8, 10.9) | 0.0 (0.0, 0.0) * | 0.125 | 0.092 | <0.001 | 0.092 |
| Coffee, tea, cocoa | 136.0 (37.2, 244.3) | 0.0 (0.0, 50.0) * | 255.6 (91.2, 420.4) | 0.0 (0.0, 0.0) * | 0.234 | 0.642 | <0.001 | 0.085 |
| Processed meat | 54.8 (44.0, 64.6) | 0.0 (0.0, 12.0) * | 38.2 (10.2, 58.8) | 0.0 (0.0, 6.0) * | 0.202 | 0.257 | <0.001 | 0.491 |
| Viennese pastries | 4.3 (0.0, 24.6) | 0.0 (0.0, 0.0) * | 6.7 (0.0, 12.8) | 0.0 (0.0, 0.0) * | 0.860 | NA | <0.001 | NA |
| Non-dairy and non-fermented foods subgroups | ||||||||
| Breakfast cereals | 0.0 (0.0, 21.6) | 7.9 (0.0, 17.8) | 0.0 (0.0, 0.0) | 0.0 (0.0, 12.3) | 0.218 | 0.374 | 0.066 | 0.354 |
| Eggs | 25.3 (14.8, 55.1) | 25.0 (11.2, 42.1) | 24.6 (17.7, 34.4) | 41.7 (25.0, 50.0) | 0.905 | 0.444 | 0.159 | 0.008 |
| Fish and seafood | 28.1 (16.7, 61.5) | 51.4 (14.2, 78.3) | 45.4 (26.1, 64.6) | 75.2 (29.2, 83.8) | 0.234 | 0.330 | 0.249 | 0.959 |
| Fruits | 93.9 (50.6, 201.5) | 359.4 (249.2, 563.8) * | 230.1 (186.8, 293.7) # | 295.7 (260.7, 339.7) * | 0.025 | 0.382 | <0.001 | 0.021 |
| Ice cream and sorbet | 0.0 (0.0, 16.9) | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.0) | 0.352 | 0.970 | 0.022 | 0.117 |
| Ingredients | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.1) | 0.0 (0.0, 0.0) | 0.529 | 0.429 | 0.683 | 0.881 |
| Jam | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.0) | 0.0 (0.0, 5.7) | 0.0 (0.0, 0.0) | 0.067 | 0.074 | 0.202 | 0.278 |
| Juice | 47.3 (9.4, 62.8) | 44.1 (4.6, 118.7) | 14.1 (3.1, 39.8) | 59.8 (11.3, 103.8) * | 0.307 | 0.744 | 0.164 | 0.269 |
| Margarines | 0.3 (0, 0.6) | 2.3 (0.0, 4.8) * | 0.0 (0.0, 3.6) | 6.7 (2.7, 7.7) | 0.979 | 0.286 | 0.001 | 0.272 |
| Non-alcoholic drinks | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.0) | 0.0 (0.0, 26.7) | 0.591 | 0.430 | 0.243 | 0.772 |
| Non-dairy desserts | 0.0 (0.0, 0.0) | 0.0 (0.0, 91.7) | 0.0 (0.0, 0.0) | 0.0 (0.0, 166.7) | 0.563 | 0.766 | 0.019 | 0.813 |
| Non-fermented bread | 0.0 (0.0, 0.0) | 30.5 (11.6, 53.0) * | 0.0 (0.0, 0.0) | 54.0 (40.8, 133.0) *# | 0.969 | 0.045 | <0.001 | 0.019 |
| Non-fermented tea | 0.0 (0.0, 54.2) | 54.2 (0.0, 213.5) | 0.0 (0.0, 0.0) | 260.0 (100.0, 300.0) * | 0.089 | 0.826 | <0.001 | 0.004 |
| Nuts | 2.4 (0.0, 8.5) | 0.0 (0.0, 3.5) | 3.3 (0.0, 9.1) | 9.2 (0.0, 18.8) | 0.654 | 0.262 | 0.881 | 0.290 |
| Pasta, rice, and cereals | 126.9 (85.1, 136.1) | 137.4 (88.5, 184.1) | 85.4 (47.8, 102.6) # | 82.5 (42.5, 106.0) # | 0.012 | 0.002 | 0.741 | 0.721 |
| Plant-based drinks | 0.0 (0.0, 0.0) | 53.1 (0.0, 102.5) * | 0.0 (0.0, 0.0) | 67.0 (6.7, 96.0) * | 0.625 | 0.919 | <0.001 | 0.549 |
| Potatoes | 66.4 (44.8, 107.1) | 60.8 (34.4, 133.0) | 52.3 (41.3, 72.6) | 62.0 (31.8, 113.3) | 0.423 | 0.649 | 0.827 | 0.659 |
| Poultry | 25.3 (15.7, 42.0) | 80.0 (46.0, 96.7) * | 11.1 (0.0, 26.8) | 27.5 (20.3, 48.7) # | 0.163 | 0.002 | 0.001 | 0.402 |
| Red meat | 48.2 (34.6, 63.9) | 57.7 (46.8, 66.7) | 30.3 (23.4, 38.0) # | 69.7 (18.5, 96.7) * | 0.044 | 0.325 | 0.009 | 0.238 |
| Soft drinks | 97.2 (37.9, 135.4) | 0.0 (0.0, 0.0) * | 0.0 (0.0, 0.0) # | 0.0 (0.0, 0.0) | 0.001 | 0.007 | 0.003 | 0.001 |
| Soup | 0.0 (0.0, 0.2) | 0.0 (0.0, 0.0) | 11.2 (0.0, 75.6) # | 0.0 (0.0, 1.5) | 0.031 | 0.012 | 0.010 | 0.505 |
| Spices | 0.6 (0.0, 1.2) | 0.0 (0.0, 0.7) | 0.6 (0.0, 1.0) | 0.0 (0.0, 0.8) | 0.633 | 0.846 | 0.145 | 0.580 |
| Sugars and honey | 5.8 (1.8, 8.2) | 1.3 (0.0, 8.1) | 13.2 (7.9, 20.3) # | 13.7 (6.0, 22.0) # | 0.033 | 0.002 | 0.578 | 0.497 |
| Vegetable fats | 8.8 (6.3, 14.4) | 20.0 (11.9, 27.7) * | 20.3 (10.9, 23.0) # | 27.5 (22.3, 34.2) * | 0.026 | 0.033 | <0.001 | 0.592 |
| Vegetables | 158.8 (130.1, 178.9) | 203.8 (157.4, 226.7) * | 210.9 (168.2, 250.3) | 276.7 (218.8, 347.5) * | 0.068 | 0.048 | 0.001 | 0.912 |
| Water | 1183.4 (766.0, 1812.4) | 1216.3 (771.7, 1799.3) | 832.9 (765.6, 1066.7) | 864.7 (688.7, 1513.5) | 0.128 | 0.140 | 0.453 | 0.927 |
| Non-dairy, fermented/non-fermented foods subgroups | ||||||||
| Cereal composite dish | 0.0 (0.0, 6.4) | 0.0 (0.0, 0.0) | 0.0 (0.0, 32.0) | 0.0 (0.0, 20.2) | 0.300 | 0.181 | 0.878 | 0.971 |
| Condiments | 3.6 (0.3, 10.3) | 0.0 (0.0, 0.0) * | 11.1 (8.4, 14.1) # | 0.0 (0.0, 0.0) * | 0.027 | 0.120 | <0.001 | 0.010 |
| Offal | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.0) | 0.0 (0.0, 10.0) | 0.0 (0.0, 14.3) | 0.098 | 0.006 | 0.675 | 0.970 |
| Flour | 4.7 (0.4, 10.2) | 0.0 (0.0, 5.0) | 6.4 (1.8, 12.0) | 2.3 (0.0, 10.5) | 0.465 | 0.209 | 0.045 | 0.618 |
| Sauces | 10.8 (2.9, 19.1) | 0.0 (0.0, 0.0) * | 3.3 (2.2, 6.7) | 0.0 (0.0, 1.7) # | 0.103 | 0.557 | <0.001 | 0.050 |
| Starters | 0.0 (0.0, 0.0) | 0.0 (0.0, 0.0) | 0.0 (0.0, 3.9) | 0.0 (0.0, 0.0) | 0.176 | 0.261 | 0.016 | 0.164 |
| Sweet biscuits | 1.3 (0.0, 15.6) | 0.0 (0.0, 7.9) | 0.0 (0.0, 3.3) | 0.0 (0.0, 18.0) | 0.379 | 0.932 | 0.799 | 0.274 |
| YA | OA | p-Value (Wilcoxon 1) | p-Value (Wald Test) a | |||||
|---|---|---|---|---|---|---|---|---|
| OB Phase | SC Phase | OB Phase | SC Phase | Baseline OA vs. YA | Age Effect | Diet Effect | Interaction | |
| Energy and macronutrients | ||||||||
| Energy, kcal/d | 2321 (2021, 2465) | 1693 (1319, 1889) * | 2433 (2227, 2588) | 2000 (1868, 2564) *# | 0.583 | 0.101 | <0.001 | 0.031 |
| Energy, kJ/d | 9710 (8454, 10,314) | 7081 (5519, 7901) * | 10,178 (9316, 10,827) | 8367 (7815, 10,726) *# | 0.583 | 0.101 | <0.001 | 0.031 |
| Total Fat, g/d | 86.1 (74.4, 99.8) | 51.2 (36.5, 70.9) * | 92.2 (84.7, 98.9) | 73.4 (69.8, 95.6) *# | 0.616 | 0.112 | <0.001 | 0.018 |
| Fat, %E | 34.5 (32.9, 37.0) | 29.9 (24.5, 33.3) * | 35.1 (32.3, 36.1) | 32.2 (30.3, 34.6) * | 0.867 | 0.574 | <0.001 | 0.342 |
| SFA, g/d | 35.3 (29.6, 40.0) | 12.6 (8.0, 14.5) * | 35.6 (27.3, 45.9) | 14.0 (11.5, 18.3) * | 0.650 | 0.173 | <0.001 | 0.211 |
| SFA, % of total fat | 40.4 (35.2, 41.5) | 22.4 (20.8, 24.6) * | 41.0 (35.2, 43.9) | 19.6 (18.4, 21.9) * | 0.830 | 0.529 | <0.001 | 0.192 |
| MUFA, g/d | 27.3 (24.5, 32.2) | 24.0 (14.4, 32.4) | 31.4 (26.3, 36.9) | 37.5 (30.0, 42.8) # | 0.350 | 0.049 | 0.723 | 0.018 |
| MUFA, % of total fat | 32.0 (30.9, 34.3) | 45.1 (39.7, 46.0) * | 34.1 (32.4, 37.8) | 46.0 (42.7, 50.0) * | 0.076 | 0.044 | <0.001 | 0.871 |
| PUFA, g/d | 10.7 (7.8, 12.2) | 9.4 (6.0, 11.5) | 12.3 (9.6, 14.1) | 11.6 (8.9, 14.7) | 0.280 | 0.187 | 0.435 | 0.854 |
| PUFA, % of total fat | 10.9 (9.3, 15.1) | 17.7 (15.3, 22.1) * | 13.5 (11.3, 14.4) | 15.7 (13.2, 19.4) * | 0.185 | 0.869 | <0.001 | 0.085 |
| Carbohydrate, g/d | 214.3 (195.2, 270.0) | 184.4 (163.3, 253.1) * | 240.9 (230.5, 256.8) | 252.5 (199.6, 287.4) # | 0.302 | 0.124 | 0.041 | 0.019 |
| Carbohydrate, %E | 41.8 (35.2, 43.9) | 48.3 (41.9, 52.0) * | 42.8 (39.5, 44.0) | 50.1 (47.2, 51.5) * | 0.830 | 0.590 | <0.001 | 0.761 |
| Protein, g/d | 100.1 (83.7, 115.2) | 76.6 (62.2, 99.2) * | 94.5 (75.7, 105.0) | 85.9 (64.2, 97.1) * | 0.259 | 0.587 | <0.001 | 0.313 |
| Protein, %E | 17.4 (16.1, 19.3) | 20.8 (18.2, 22.5) * | 15.4 (14.1, 16.3) # | 15.9 (13.8, 16.2) # | 0.033 | 0.002 | 0.140 | 0.236 |
| Starch, g/d | 100.0 (80.9, 110.0) | 71.8 (54.0, 110.0) * | 102.8 (85.5, 127.6) | 78.2 (60.3, 114.7) | 0.430 | 0.607 | 0.002 | 0.745 |
| Sugars, g/d | 89.9 (76.7, 101.5) | 89.4 (61.9, 101.1) | 93.5 (83.5, 100.2) | 98.9 (92.7, 116.8) | 0.458 | 0.179 | 0.231 | 0.089 |
| Alcohol, g/d | 10.2 (2.1, 18.6) | 0.0 (0.0, 0.0) * | 13.3 (4.8, 32.3) | 0.0 (0.0, 0.0) * | 0.905 | 0.878 | <0.001 | 0.975 |
| Alcohol, %E | 3.3 (0.6, 5.0) | 0.0 (0.0, 0.0) * | 3.7 (1.0, 8.0) | 0.0 (0.0, 0.0) * | 0.830 | 0.907 | <0.001 | 0.970 |
| Water, g | 2594 (2005, 2900) | 2299 (2069, 2844) | 2314 (1962, 2645) | 2101 (1864, 2682) | 0.402 | 0.466 | 0.359 | 0.909 |
| Fibre, g/d | 19.5 (16.8, 21.9) | 19.5 (16.7, 22.2) | 21.9 (19.9, 28.0) | 26.8 (17.1, 31.0) | 0.068 | 0.063 | 0.684 | 0.841 |
| Cholesterol, mg/d | 355 (282, 479) | 262 (195, 343) | 323 (267, 435) | 335 (297, 369) | 0.458 | 0.778 | 0.040 | 0.115 |
| Minerals & trace elements | ||||||||
| Na, mg/d | 2571 (2246, 2901) | 887 (684, 1151) * | 2482 (2309, 2622) | 1152 (871, 1615) * | 0.943 | 0.245 | <0.001 | 0.122 |
| K, mg/d | 2932 (2462, 3513) | 2529 (2151, 3257) | 3186 (3132, 3605) | 2873 (2502, 3236) * | 0.259 | 0.346 | 0.001 | 0.695 |
| Mg, mg/d | 302 (268, 383) | 251 (209, 311) * | 347 (292, 395) | 324 (236, 420) | 0.375 | 0.221 | 0.001 | 0.514 |
| P, mg/d | 1456 (1182, 1562) | 1055 (762, 1144) * | 1302 (1240, 1448) | 999 (801, 1190) * | 0.302 | 0.836 | <0.001 | 0.289 |
| Ca, mg/d | 1071 (830, 1261) | 399 (350, 607) * | 1032 (900, 1137) | 498 (415, 664) * | 0.756 | 0.435 | <0.001 | 0.044 |
| Fe, mg/d | 11.9 (9.5, 14.0) | 9.1 (6.6, 12.1) * | 12.3 (10.9, 13.6) | 11.3 (9.3, 12.2) * | 0.650 | 0.323 | 0.001 | 0.525 |
| Vitamins | ||||||||
| Vitamin A, µg RE/d | 773 (559, 1093) | 778 (392, 831) | 1281 (870, 1590) # | 850 (746, 1414) | 0.022 | 0.031 | 0.103 | 0.400 |
| Vitamin B1, mg/d | 1.3 (1.1, 1.5) | 1.1 (0.8, 1.2) * | 1.2 (1, 1.3) | 1.1 (0.9, 1.2) | 0.239 | 0.637 | 0.001 | 0.068 |
| Vitamin B2, mg/d | 2.0 (1.6, 2.1) | 0.9 (0.7, 1.1) * | 1.8 (1.7, 2.1) | 1.1 (0.9, 1.4) * | 0.905 | 0.471 | <0.001 | 0.177 |
| Vitamin B3 mg/d | 19.7 (15.8, 22.8) | 23.5 (16.5, 26.4) | 17.6 (14.5, 22.5) | 19.0 (14.9, 23.4) | 0.488 | 0.251 | 0.090 | 0.396 |
| Vitamin B5, mg/d | 6.1 (5.5, 7.1) | 4.7 (3.9, 6.0) * | 5.6 (4.6, 6.5) | 4.6 (4.2, 6.2) | 0.350 | 0.696 | 0.001 | 0.312 |
| Vitamin B6, mg/d | 1.9 (1.6, 2.3) | 2.0 (1.7, 2.3) | 1.6 (1.5, 2.3) | 2.0 (1.7, 2.3) | 0.488 | 0.514 | 0.090 | 0.804 |
| Vitamin B9, µg/d | 308.9 (260.7, 356.7) | 272.3 (219.7, 386.2) | 375.0 (329.9, 390.3) # | 336.7 (257.8, 441.8) | 0.025 | 0.079 | 0.201 | 0.704 |
| Vitamin B12, µg/d | 4.7 (4.0, 8.0) | 4.3 (3.2, 4.9) * | 5.8 (4.2, 14.2) | 9.5 (6.5, 12.5) # | 0.302 | 0.007 | 0.950 | 0.056 |
| Vitamin C, mg/d | 62.7 (45.7, 98.4) | 105.1 (51.1, 182.9) | 95.2 (81.5, 114.9) | 133.0 (98.4, 156.0) * | 0.085 | 0.142 | 0.001 | 0.993 |
| Vitamin D, µg/d | 4.2 (3.4, 5.4) | 3.3 (2.2, 3.8) | 4.2 (3.6, 5.1) | 5.0 (3.1, 6.1) | 0.943 | 0.235 | 0.159 | 0.209 |
| Vitamin E, mg/d | 9.7 (9.5, 11.7) | 13.5 (10.6, 16.3) * | 12.0 (10.5, 13.5) | 18.1 (14.0, 23.8) * | 0.116 | 0.082 | <0.001 | 0.520 |
| YA | OA | p-Value (Wilcoxon 1) | p-Value (Wald Test) a | |||||
|---|---|---|---|---|---|---|---|---|
| OB Phase | SC Phase | OB Phase | SC Phase | Baseline OA vs. YA | Age Effect | Diet Effect | Interaction | |
| Body weight, kg | 80.7 (76.0, 84.4) | 79.1 (75.1, 83.6) | 73.0 (68.5, 76.4) # | 71.4 (67.0, 74.9) # | 0.049 | 0.029 | <0.001 | 0.963 |
| BMI, kg/m2 | 25.7 (22.6, 26.3) | 25.1 (22.3, 25.9) | 24.3 (23.2, 26.8) | 23.8 (22.6, 26.9) | 0.981 | 0.990 | <0.001 | 0.725 |
| Biochemical parameters | ||||||||
| Insulin, pM | 40.16 (24.34, 45.39) | 26.32 (15.06, 33.93) | 22.60 (19.10, 39.00) | 21.60 (14.34, 27.43) | 0.202 | 0.251 | 0.002 | 0.515 |
| Glucose, mM | 5.31 (4.95, 5.56) | 4.98 (4.87, 5.21) | 5.40 (5.20, 5.70) | 5.19 (5.03, 5.35) | 0.275 | 0.106 | <0.001 | 0.850 |
| Triglycerides, mM | 0.99 (0.76, 1.27) | 0.67 (0.61, 1.06) | 0.90 (0.80, 1.10) | 0.93 (0.74, 1.20) | 0.734 | 0.495 | 0.135 | 0.091 |
| Total cholesterol, mM | 4.43 (4.20, 4.95) | 3.98 (3.59, 4.53) * | 5.70 (5.30, 6.40) # | 5.06 (4.88, 5.84) # | 0.001 | <0.001 | <0.001 | 0.698 |
| HDL, mM | 1.23 (1.11, 1.41) | 1.14 (1.01, 1.37) | 1.40 (1.30, 1.60) | 1.21 (1.15, 1.41) | 0.126 | 0.207 | <0.001 | 0.117 |
| LDL, mM | 2.71 (2.52, 3.08) | 2.28 (2.06, 2.62) | 3.60 (3.10, 3.80) # | 3.36 (2.79, 3.56) # | 0.001 | <0.001 | <0.001 | 0.815 |
| Total cholesterol/HDL ratio | 1.72 (1.60, 1.79) | 1.72 (1.691, 1.857) | 1.70 (1.50, 1.80) | 1.640 (1.46, 1.86) | 0.402 | 0.271 | 0.604 | 0.304 |
| Total cholesterol/LDL ratio | 3.75 (3.27, 4.28) | 3.31 (3.24, 4.06) | 3.90 (3.70, 4.30) | 4.132 (3.90, 4.58) # | 0.350 | 0.045 | 0.461 | 0.018 |
| LDL/HDL ratio | 2.27 (1.99, 2.57) | 1.96 (1.80, 2.33) | 2.40 (2.10, 2.70) | 2.78 (2.07, 3.10) # | 0.375 | 0.060 | 0.202 | 0.036 |
| NEFA, mM | 0.20 (0.14, 0.23) | 0.21 (0.12, 0.26) | 0.20 (0.10, 0.20) | 0.19 (0.15, 0.23) | 0.495 | 0.907 | 0.906 | 0.519 |
| Urea, mM | 5.89 (5.44, 6.22) | 5.26 (4.89, 6.22) | 6.40 (5.70, 7.20) | 6.28 (5.09, 7.52) | 0.141 | 0.070 | 0.109 | 0.772 |
| Lactate, mM | 1.55 (1.37, 1.80) | 1.61 (1.41, 1.88) | 1.60 (1.30, 1.70) | 1.52 (1.43, 1.71) | 0.771 | 0.796 | 0.554 | 0.958 |
| Inflammation parameters | ||||||||
| CRP, ng/mL | 19.36 (16.71, 28.80) | 19.327 (12.42, 21.48) | 39.20 (24.80, 54.50) # | 40.65 (23.20, 69.84) # | 0.038 | 0.002 | 0.087 | 0.087 |
| Adiponectin, ng/mL | 8.99 (8.47, 10.71) | 9.13 (7.18, 10.67) | 7.70 (6.20, 7.90) # | 6.29 (5.81, 7.86) # | 0.012 | 0.003 | 0.737 | 0.920 |
| YA | OA | p-Value (Wilcoxon 1) | p-Value (Wald Test) a | |||||
|---|---|---|---|---|---|---|---|---|
| OB Phase | SC Phase | OB Phase (IQR) | SC Phase (IQR) | Baseline OA vs. YA | Age Effect | Diet Effect | Interaction | |
| Amino acids | ||||||||
| Total amino acids, A.U. | 14.53 (12.64, 18.54) | 13.21 (10.59, 16.21) | 13.40 (10.32, 18.48) | 14.41 (10.65, 16.02) | 0.650 | 0.678 | 0.203 | 0.777 |
| Essential amino acids, A.U. | 6.60 (6.19, 7.61) | 5.70 (4.83, 7.46) | 5.79 (5.48, 8.00) | 6.93 (4.87, 7.41) | 0.793 | 0.891 | 0.283 | 0.849 |
| BCAA, A.U. | 2.11 (1.81, 2.55) | 2.10 (1.60, 2.31) | 2.36 (2.06, 3.00) | 2.09 (1.67, 2.49) | 0.202 | 0.498 | 0.016 | 0.297 |
| Alanine, A.U. | 0.97 (0.84, 1.35) | 0.92 (0.67, 1.21) | 0.95 (0.66, 1.67) | 1.10 (0.73, 1.27) | 0.943 | 0.724 | 0.809 | 0.600 |
| Asparagine, A.U. | 0.45 (0.25, 0.69) | 0.41 (0.29, 0.74) | 0.62 (0.38, 0.78) | 0.65 (0.42, 0.73) | 0.259 | 0.227 | 0.942 | 0.844 |
| Aspartic acid, A.U. | 0.58 (0.41, 0.81) | 0.48 (0.46, 0.68) | 0.47 (0.35, 0.68) | 0.49 (0.29, 0.72) | 0.519 | 0.431 | 0.631 | 0.934 |
| Cysteine, A.U. | 0.76 (0.47, 1.30) | 0.82 (0.59, 1.35) | 0.90 (0.68, 1.20) | 0.86 (0.41, 1.28) | 0.793 | 0.918 | 0.765 | 0.518 |
| Glutamic acid, A.U. | 0.64 (0.57, 0.73) | 0.59 (0.46, 0.91) | 0.43 (0.36, 0.72) | 0.55 (0.31, 0.63) | 0.350 | 0.231 | 0.479 | 0.994 |
| Glycine, A.U. | 1.20 (0.87, 1.31) | 1.05 (0.75, 1.32) | 0.86 (0.69, 1.02) | 0.92 (0.66, 1.13) | 0.169 | 0.168 | 0.671 | 0.615 |
| Isoleucine, A.U. | 0.74 (0.58, 0.83) | 0.70 (0.58, 0.86) | 0.62 (0.58, 1.09) | 0.65 (0.52, 0.80) | 0.793 | 0.750 | 0.194 | 0.341 |
| Leucine, A.U. | 0.61 (0.52, 0.84) | 0.55 (0.44, 0.70) | 0.84 (0.58, 1.00) | 0.65 (0.49, 0.81) | 0.169 | 0.318 | 0.005 | 0.307 |
| Lysine, A.U. | 1.00 (0.81, 1.13) | 0.89 (0.71, 1.11) | 0.93 (0.64, 1.00) | 1.03 (0.90, 1.11) | 0.550 | 0.866 | 0.497 | 0.172 |
| Methionine, A.U. | 0.85 (0.67, 1.11) | 0.78 (0.61, 1.03) | 0.77 (0.52, 1.00) | 0.87 (0.45, 1.00) | 0.402 | 0.646 | 0.725 | 0.424 |
| Phenylalanine, A.U. | 0.95 (0.77, 1.05) | 0.80 (0.69, 0.99) | 0.81 (0.69, 0.98) | 0.98 (0.59, 1.06) | 0.488 | 0.839 | 0.842 | 0.274 |
| Proline, A.U. | 1.18 (0.60, 2.28) | 0.82 (0.29, 1.50) | 1.02 (0.42, 1.81) | 0.94 (0.30, 1.40) | 0.375 | 0.463 | 0.127 | 0.846 |
| Serine, A.U. | 0.87 (0.50, 1.05) | 0.75 (0.44, 1.03) | 0.60 (0.48, 1.11) | 0.54 (0.50, 0.91) | 0.720 | 0.534 | 0.327 | 0.769 |
| Taurine, A.U. | 1.36 (1.17, 2.04) | 1.39 (1.12, 1.56) | 0.99 (0.69, 1.70) | 1.36 (0.79, 1.57) | 0.155 | 0.176 | 0.658 | 0.584 |
| Threonine, A.U. | 0.85 (0.65, 1.05) | 0.83 (0.62, 1.08) | 0.80 (0.55, 0.96) | 0.68 (0.50, 0.89) | 0.430 | 0.267 | 0.503 | 0.728 |
| Tryptophan, A.U. | 0.86 (0.62, 1.22) | 0.80 (0.52, 0.96) | 0.87 (0.60, 0.92) | 0.63 (0.60, 0.87) | 0.458 | 0.551 | 0.135 | 0.522 |
| Tyrosine, A.U. | 0.87 (0.70, 0.99) | 0.78 (0.63, 0.84) | 0.94 (0.83, 0.99) | 0.88 (0.79, 0.98) | 0.519 | 0.181 | 0.018 | 0.507 |
| Valine, A.U. | 0.84 (0.74, 0.94) | 0.75 (0.65, 0.83) | 0.93 (0.89, 1.01) | 0.85 (0.67, 0.88) * | 0.105 | 0.253 | 0.001 | 0.274 |
| Carbohydrates and derivatives | ||||||||
| Lactose, A.U. | 0.15 (0.13, 0.25) | 0.12 (0.10, 0.16) * | 0.17 (0.16, 0.23) | 0.13 (0.11, 0.17) | 0.720 | 0.517 | <0.001 | 0.731 |
| Galactose, A.U. | 0.00 (0.00, 0.00) | 0.00 (0.00, 0.00) | 0.00 (0.00, 0.00) | 0.00 (0.00, 0.00) | NA | NA | NA | NA |
| Galactonate, A.U. | 0.08 (0.07, 0.13) | 0.07 (0.04, 0.09) | 0.14 (0.12, 0.15) | 0.08 (0.07, 0.14) | 0.054 | 0.005 | 0.067 | 0.767 |
| Galactitol, A.U. | 0.45 (0.40, 0.57) | 0.46 (0.33, 0.57) | 0.71 (0.54, 0.77) # | 0.72 (0.52, 0.84) # | 0.033 | 0.001 | 0.779 | 0.514 |
| Maltose, A.U. | 1.18 (0.94, 1.99) | 1.12 (0.63, 1.69) | 1.04 (0.64, 1.48) | 0.79 (0.65, 1.17) | 0.350 | 0.254 | 0.070 | 0.926 |
| Fatty acids (sums of individual FA) | ||||||||
| Total fatty acids, mg/L | 257.4 (247.1, 296.6) | 269.7 (259.9, 302.0) | 291.5 (273.4, 328.6) # | 301.5 (261.2, 345.8) | 0.025 | 0.079 | 0.626 | 0.131 |
| SCFA (C4-C10), mg/L | 0.0 (0.0, 0.1) | 0.1 (0.0, 0.1) | 0.1 (0.0, 0.1) | 0.1 (0.0, 0.1) | 0.756 | 0.686 | 0.703 | 0.987 |
| MCFA (C11-C16), mg/L | 77.6 (72.0, 92.4) | 78.3 (72.4, 84.3) | 91.2 (88.5, 104.5) # | 86.0 (72.7, 102.6) | 0.048 | 0.068 | 0.265 | 0.241 |
| LCFA (>C17), mg/L | 159.6 (151.2, 181.0) | 174.5 (160.6, 194.3) | 180.7 (164.8, 194.7) # | 190.2 (159.4, 214.5) | 0.038 | 0.167 | 0.236 | 0.078 |
| SFA, mg/L | 106.0 (99.6, 121.6) | 102.0 (96.4, 110.1) | 121.1 (117.8, 132.3) # | 113.5 (97.6, 134.4) | 0.022 | 0.043 | 0.060 | 0.203 |
| SFA, % of total fatty acids | 40.7 (40.1, 41.9) | 37.8 (36.1, 38.7) * | 41.5 (40.5, 41.7) | 38.0 (37.6, 38.7) * | 0.616 | 0.480 | <0.001 | 0.889 |
| USFA, mg/L | 131.8 (124.0, 150.9) | 151.4 (134.8, 172.2) * | 152.3 (133.6, 162.5) | 160.5 (134.6, 181.7) | 0.068 | 0.202 | 0.040 | 0.143 |
| MUFA, mg/L | 64.6 (50.8, 72.6) | 79.6 (69.0, 95.0) * | 68.2 (61.7, 82.8) | 81.9 (61.9, 100.0) | 0.325 | 0.530 | 0.004 | 0.201 |
| MUFA, % of total fatty acids | 23.4 (21.6, 25.7) | 28.0 (26.4, 30.2) * | 22.5 (21.6, 26.6) | 26.3 (24.4, 31.2) | 0.943 | 0.869 | <0.001 | 0.571 |
| PUFA, mg/L | 72.5 (67.2, 74.0) | 75.3 (64.6, 79.4) | 80.0 (75.3, 90.7) # | 74.3 (71.4, 80.8) | 0.003 | 0.034 | 0.564 | 0.058 |
| PUFA, % of total fatty acids | 27.6 (26.6, 28.0) | 26.1 (25.2, 27.8) | 27.6 (25.0, 28.1) | 26.6 (24.5, 27.1) | 0.720 | 0.616 | 0.140 | 0.826 |
| Omega 3 fatty acids, mg/L | 10.0 (8.6, 11.6) | 11.8 (9.1, 12.7) | 16.6 (13.5, 18.9) # | 12.4 (12.1, 15.3) | 0.001 | <0.001 | 0.696 | 0.047 |
| Omega 6 fatty acids, mg/L | 61.0 (56.8, 66.6) | 63.2 (56.4, 70.5) | 65.8 (62.9, 69.0) | 62.0 (58.0, 67.9) | 0.128 | 0.389 | 0.752 | 0.161 |
| BCFA, mg/L | 1.4 (1.0, 1.5) | 1.2 (0.8, 1.4) | 1.8 (1.3, 2.2) # | 1.2 (0.9, 1.3) * | 0.022 | 0.141 | <0.001 | 0.082 |
| TFA, mg/L | 4.9 (4.0, 5.6) | 4.4 (4.1, 5.1) | 5.3 (4.8, 6.5) | 4.9 (4.3, 5.7) | 0.202 | 0.189 | 0.080 | 0.789 |
| TFA without CLA, mg/L | 4.4 (3.8, 4.9) | 4.1 (3.8, 4.7) | 4.5 (3.9, 5.9) | 4.4 (3.7, 5.4) | 0.350 | 0.424 | 0.206 | 0.706 |
| CLA, mg/L | 0.5 (0.4, 0.6) | 0.4 (0.2, 0.5) | 0.7 (0.5, 0.9) | 0.5 (0.4, 0.6) | 0.061 | 0.096 | 0.001 | 0.639 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Burton-Pimentel, K.J.; Fleuti, C.; Blaser, C.; Scherz, V.; Badertscher, R.; Marmonier, C.; Lyon-Belgy, N.; Caille, A.; Pidou, V.; et al. Microbiota and Metabolite Modifications after Dietary Exclusion of Dairy Products and Reduced Consumption of Fermented Food in Young and Older Men. Nutrients 2021, 13, 1905. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13061905
Kim J, Burton-Pimentel KJ, Fleuti C, Blaser C, Scherz V, Badertscher R, Marmonier C, Lyon-Belgy N, Caille A, Pidou V, et al. Microbiota and Metabolite Modifications after Dietary Exclusion of Dairy Products and Reduced Consumption of Fermented Food in Young and Older Men. Nutrients. 2021; 13(6):1905. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13061905
Chicago/Turabian StyleKim, Jinyoung, Kathryn J. Burton-Pimentel, Charlotte Fleuti, Carola Blaser, Valentin Scherz, René Badertscher, Corinne Marmonier, Noëlle Lyon-Belgy, Aurélie Caille, Véronique Pidou, and et al. 2021. "Microbiota and Metabolite Modifications after Dietary Exclusion of Dairy Products and Reduced Consumption of Fermented Food in Young and Older Men" Nutrients 13, no. 6: 1905. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13061905