
Effects of a Diet Based on Foods from Symbiotic Agriculture on the Gut Microbiota of Subjects at Risk for Metabolic Syndrome

 , , , , , , , , ,
, , , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Setting and Participants
2.2. Study Intervention
2.3. Collection of Participants’ Information and Samples
2.4. Dietary Habits at Baseline
2.5. Gut Microbiota Analysis through Illumina Sequencing
2.6. Urine Metabolomics by Nuclear Magnetic Resonance Spectroscopy
2.7. Statistical Analyses
3. Results
3.1. Description of the Study Participants at Baseline
3.1.1. Anthropometric, Biochemical and Immunological Characteristics
3.1.2. Dietary Habits
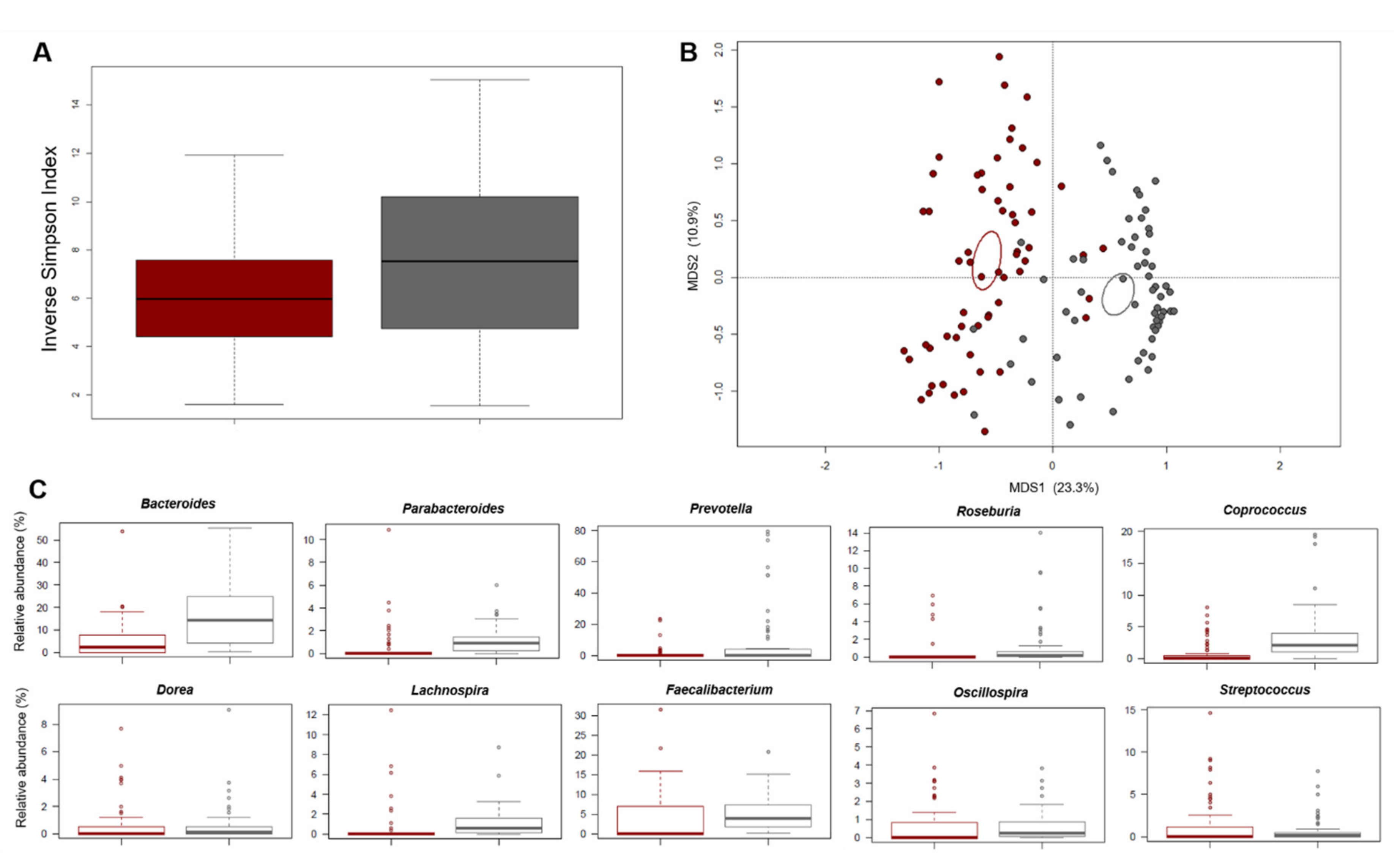
3.1.3. Gut Microbiota Profiling
3.1.4. Urine Nuclear Magnetic Resonance-Based Metabolomics
3.2. Effects of the Dietary Intervention
3.2.1. Impact on Anthropometric, Biochemical and Immunological Parameters
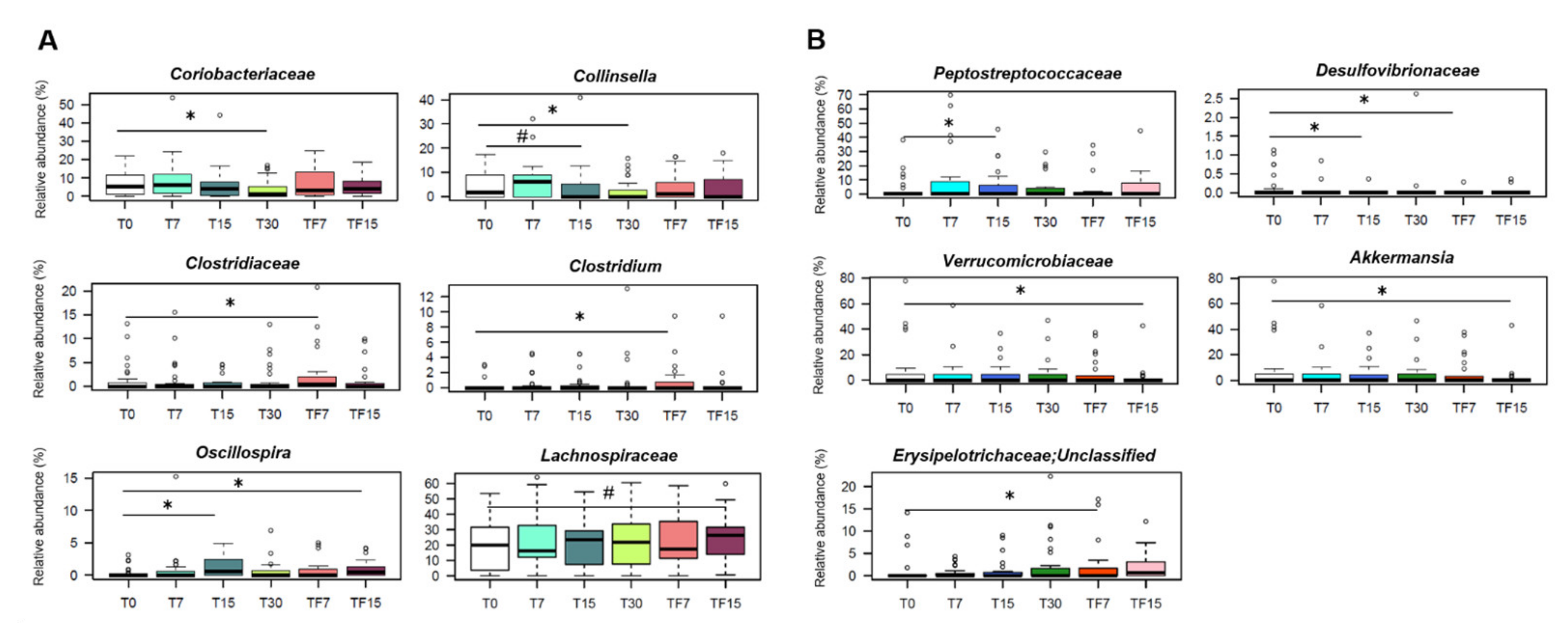
3.2.2. Impact on the Gut Microbiota Composition
3.2.3. Impact on the Urine Metabolome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lemieux, I.; Despres, J.P. Metabolic Syndrome: Past, Present and Future. Nutrients 2020, 12, 3501. [Google Scholar] [CrossRef] [PubMed]
- Dabke, K.; Hendrick, G.; Devkota, S. The gut microbiome and metabolic syndrome. J. Clin. Investig. 2019, 129, 4050–4057. [Google Scholar] [CrossRef] [PubMed]
- Vishram, J.K.; Borglykke, A.; Andreasen, A.H.; Jeppesen, J.; Ibsen, H.; Jorgensen, T.; Palmieri, L.; Giampaoli, S.; Donfrancesco, C.; Kee, F.; et al. Impact of age and gender on the prevalence and prognostic importance of the metabolic syndrome and its components in Europeans. The MORGAM Prospective Cohort Project. PLoS ONE 2014, 9, e107294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mottillo, S.; Filion, K.B.; Genest, J.; Joseph, L.; Pilote, L.; Poirier, P.; Rinfret, S.; Schiffrin, E.L.; Eisenberg, M.J. The metabolic syndrome and cardiovascular risk a systematic review and meta-analysis. J. Am. Coll. Cardiol. 2010, 56, 1113–1132. [Google Scholar] [CrossRef] [Green Version]
- Choi, I.Y.; Chun, S.; Shin, D.W.; Han, K.; Jeon, K.H.; Yu, J.; Chae, B.J.; Suh, M.; Park, Y.M. Changes in Metabolic Syndrome Status and Breast Cancer Risk: A Nationwide Cohort Study. Cancers 2021, 13, 1177. [Google Scholar] [CrossRef]
- Esposito, K.; Chiodini, P.; Colao, A.; Lenzi, A.; Giugliano, D. Metabolic syndrome and risk of cancer: A systematic review and meta-analysis. Diabetes Care 2012, 35, 2402–2411. [Google Scholar] [CrossRef] [Green Version]
- Han, F.; Wu, G.; Zhang, S.; Zhang, J.; Zhao, Y.; Xu, J. The association of Metabolic Syndrome and its Components with the Incidence and Survival of Colorectal Cancer: A Systematic Review and Meta-analysis. Int. J. Biol. Sci. 2021, 17, 487–497. [Google Scholar] [CrossRef]
- Mendonca, F.M.; de Sousa, F.R.; Barbosa, A.L.; Martins, S.C.; Araujo, R.L.; Soares, R.; Abreu, C. Metabolic syndrome and risk of cancer: Which link? Metabolism 2015, 64, 182–189. [Google Scholar] [CrossRef]
- Xia, B.; He, Q.; Pan, Y.; Gao, F.; Liu, A.; Tang, Y.; Chong, C.; Teoh, A.Y.B.; Li, F.; He, Y.; et al. Metabolic syndrome and risk of pancreatic cancer: A population-based prospective cohort study. Int. J. Cancer 2020, 147, 3384–3393. [Google Scholar] [CrossRef]
- Gallardo-Becerra, L.; Cornejo-Granados, F.; Garcia-Lopez, R.; Valdez-Lara, A.; Bikel, S.; Canizales-Quinteros, S.; Lopez-Contreras, B.E.; Mendoza-Vargas, A.; Nielsen, H.; Ochoa-Leyva, A. Metatranscriptomic analysis to define the Secrebiome, and 16S rRNA profiling of the gut microbiome in obesity and metabolic syndrome of Mexican children. Microb. Cell Fact. 2020, 19, 61. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Wu, W.; Wu, S.; Zheng, H.M.; Li, P.; Sheng, H.F.; Chen, M.X.; Chen, Z.H.; Ji, G.Y.; Zheng, Z.D.; et al. Linking gut microbiota, metabolic syndrome and economic status based on a population-level analysis. Microbiome 2018, 6, 172. [Google Scholar] [CrossRef]
- Kootte, R.S.; Levin, E.; Salojarvi, J.; Smits, L.P.; Hartstra, A.V.; Udayappan, S.D.; Hermes, G.; Bouter, K.E.; Koopen, A.M.; Holst, J.J.; et al. Improvement of Insulin Sensitivity after Lean Donor Feces in Metabolic Syndrome Is Driven by Baseline Intestinal Microbiota Composition. Cell Metab. 2017, 26, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Org, E.; Blum, Y.; Kasela, S.; Mehrabian, M.; Kuusisto, J.; Kangas, A.J.; Soininen, P.; Wang, Z.; Ala-Korpela, M.; Hazen, S.L.; et al. Relationships between gut microbiota, plasma metabolites, and metabolic syndrome traits in the METSIM cohort. Genome Biol. 2017, 18, 70. [Google Scholar] [CrossRef]
- Wutthi-In, M.; Cheevadhanarak, S.; Yasom, S.; Kerdphoo, S.; Thiennimitr, P.; Phrommintikul, A.; Chattipakorn, N.; Kittichotirat, W.; Chattipakorn, S. Gut Microbiota Profiles of Treated Metabolic Syndrome Patients and their Relationship with Metabolic Health. Sci. Rep. 2020, 10, 10085. [Google Scholar] [CrossRef]
- Cancello, R.; Turroni, S.; Rampelli, S.; Cattaldo, S.; Candela, M.; Cattani, L.; Mai, S.; Vietti, R.; Scacchi, M.; Brigidi, P.; et al. Effect of Short-Term Dietary Intervention and Probiotic Mix Supplementation on the Gut Microbiota of Elderly Obese Women. Nutrients 2019, 11, 3011. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D. Microbiota and metabolites in metabolic diseases. Nat. Rev. Endocrinol. 2019, 15, 69–70. [Google Scholar] [CrossRef]
- Gomez-Arango, L.F.; Barrett, H.L.; Wilkinson, S.A.; Callaway, L.K.; McIntyre, H.D.; Morrison, M.; Dekker Nitert, M. Low dietary fiber intake increases Collinsella abundance in the gut microbiota of overweight and obese pregnant women. Gut Microbes 2018, 9, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Silveira-Nunes, G.; Durso, D.F.; de Oliveira, L.R.A., Jr.; Cunha, E.H.M.; Maioli, T.U.; Vieira, A.T.; Speziali, E.; Correa-Oliveira, R.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; et al. Hypertension Is Associated With Intestinal Microbiota Dysbiosis and Inflammation in a Brazilian Population. Front. Pharmacol. 2020, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Kolodziejczyk, A.A.; Zheng, D.; Elinav, E. Diet-microbiota interactions and personalized nutrition. Nat. Rev. Micro Biol. 2019, 17, 742–753. [Google Scholar] [CrossRef]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef]
- Moszak, M.; Szulinska, M.; Bogdanski, P. You Are What You Eat-The Relationship between Diet, Microbiota, and Metabolic Disorders—A Review. Nutrients 2020, 12, 1096. [Google Scholar] [CrossRef] [Green Version]
- Hirt, H. Healthy soils for healthy plants for healthy humans: How beneficial microbes in the soil, food and gut are interconnected and how agriculture can contribute to human health. EMBO Rep. 2020, 21, e51069. [Google Scholar] [CrossRef]
- Raiola, A.; Tenore, G.C.; Petito, R.; Ciampaglia, R.; Ritieni, A. Improving of nutraceutical features of many important mediterranean vegetables by inoculation with a new commercial product. Curr. Pharm. Biotechnol. 2015, 16, 738–746. [Google Scholar] [CrossRef]
- Longo, V.C.M.; Della Croce, C.M.; Giovannetti, G.; Masciandaro, G. Assessment of nutraceutical features of different foods from conventional and mycorrhized farming. Bull. Sci. Inform. 2018, 36, 40–46. [Google Scholar]
- Giovannetti, G.; Polo, F.; Nutricato, S.; Masoero, G.; Nuti, M. Efficacy of a commercial symbiotic bio-fertilizer consortium for mitigating the olive quick decline syndrome (OQDS). J. Agron. Res. 2019, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.; Loria, C.M.; Smith, S.C., Jr.; et al. Harmonizing the metabolic syndrome. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.S.; Ho, H.H.; Hsieh, S.H.; Kuo, Y.W.; Tseng, H.Y.; Kao, H.F.; Wang, J.Y. Lactobacillus salivarius AP-32 and Lactobacillus reuteri GL-104 decrease glycemic levels and attenuate diabetes-mediated liver and kidney injury in db/db mice. BMJ Open Diabetes Res. Care 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.J.; Kim, H.J.; Kim, T.S.; Yeo, I.H.; Ji, G.E. Effects of Lactobacillus plantarum PMO 08 Alone and Combined with Chia Seeds on Metabolic Syndrome and Parameters Related to Gut Health in High-Fat Diet-Induced Obese Mice. J. Med. Food 2019, 22, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Marquez, A.; Herrera, H.; Abeijon-Mukdsi, C.; Saavedra, L.; Hebert, E.; Gauffin-Cano, P.; Medina, R. Oral administration of Lactobacillus fermentum CRL1446 improves biomarkers of metabolic syndrome in mice fed a high-fat diet supplemented with wheat bran. Food Funct. 2020, 11, 3879–3894. [Google Scholar] [CrossRef] [PubMed]
- Pala, V.; Sieri, S.; Palli, D.; Salvini, S.; Berrino, F.; Bellegotti, M.; Frasca, G.; Tumino, R.; Sacerdote, C.; Fiorini, L.; et al. Diet in the Italian EPIC cohorts: Presentation of data and methodological issues. Tumori 2003, 89, 594–607. [Google Scholar] [CrossRef]
- Pisani, P.; Faggiano, F.; Krogh, V.; Palli, D.; Vineis, P.; Berrino, F. Relative validity and reproducibility of a food frequency dietary questionnaire for use in the Italian EPIC centres. Int. J. Epidemiol. 1997, 26 (Suppl. S1), S152–S160. [Google Scholar] [CrossRef] [Green Version]
- Riboli, E.; Hunt, K.J.; Slimani, N.; Ferrari, P.; Norat, T.; Fahey, M.; Charrondiere, U.R.; Hemon, B.; Casagrande, C.; Vignat, J.; et al. European Prospective Investigation into Cancer and Nutrition (EPIC): Study populations and data collection. Public Health Nutr. 2002, 5, 1113–1124. [Google Scholar] [CrossRef]
- Salvini, S.; Parpinel, M.; Gnagnarella, P.; Maisonneuve, P.; Turrini, A. Banca di Composizione degli Alimenti per Studi Epidemiologici in Italia; Istituto Europeo di Oncologia: Milano, Italy, 1998. [Google Scholar]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef]
- D’Amico, F.; Biagi, E.; Rampelli, S.; Fiori, J.; Zama, D.; Soverini, M.; Barone, M.; Leardini, D.; Muratore, E.; Prete, A.; et al. Enteral Nutrition in Pediatric Patients Undergoing Hematopoietic SCT Promotes the Recovery of Gut Microbiome Homeostasis. Nutrients 2019, 11, 2958. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Schnorr, S.L.; Candela, M.; Rampelli, S.; Centanni, M.; Consolandi, C.; Basaglia, G.; Turroni, S.; Biagi, E.; Peano, C.; Severgnini, M.; et al. Gut microbiome of the Hadza hunter-gatherers. Nat. Commun. 2014, 5, 3654. [Google Scholar] [CrossRef] [PubMed]
- Biagi, E.; Franceschi, C.; Rampelli, S.; Severgnini, M.; Ostan, R.; Turroni, S.; Consolandi, C.; Quercia, S.; Scurti, M.; Monti, D.; et al. Gut Microbiota and Extreme Longevity. Curr. Biol. 2016, 26, 1480–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The human urine metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef]
- Agnoli, C.; Krogh, V.; Grioni, S.; Sieri, S.; Palli, D.; Masala, G.; Sacerdote, C.; Vineis, P.; Tumino, R.; Frasca, G.; et al. A priori-defined dietary patterns are associated with reduced risk of stroke in a large Italian cohort. J. Nutr. 2011, 141, 1552–1558. [Google Scholar] [CrossRef] [Green Version]
- Dalmartello, M.; Bravi, F.; Serraino, D.; Crispo, A.; Ferraroni, M.; La Vecchia, C.; Edefonti, V. Dietary Patterns in Italy and the Risk of Renal Cell Carcinoma. Nutrients 2020, 12, 134. [Google Scholar] [CrossRef] [Green Version]
- Edefonti, V.; La Vecchia, C.; Di Maso, M.; Crispo, A.; Polesel, J.; Libra, M.; Parpinel, M.; Serraino, D.; Ferraroni, M.; Bravi, F. Association between Nutrient-Based Dietary Patterns and Bladder Cancer in Italy. Nutrients 2020, 12, 1584. [Google Scholar] [CrossRef]
- Kaufman, L.; Rousseeuw, P.J. Finding Groups in Data: An Introduction to Cluster Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2009; Volume 344. [Google Scholar]
- Noguchi, K.; Gel, Y.R.; Brunner, E.; Konietschke, F. nparLD: An R software package for the nonparametric analysis of longitudinal data in factorial experiments. J. Stat. Softw. 2012, 50. [Google Scholar] [CrossRef] [Green Version]
- Vignoli, A.; Rodio, D.M.; Bellizzi, A.; Sobolev, A.P.; Anzivino, E.; Mischitelli, M.; Tenori, L.; Marini, F.; Priori, R.; Scrivo, R.; et al. NMR-based metabolomic approach to study urine samples of chronic inflammatory rheumatic disease patients. Anal. Bioanal. Chem. 2017, 409, 1405–1413. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing 2021; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.R-project.org (accessed on 7 April 2021).
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; R Package Version 2.1.3. 2021. Available online: https://CRAN.R-project.org/package=psych (accessed on 7 April 2021).
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions; R Package Version 2.1.1. 2021. Available online: https://CRAN.R-project.org/package=cluster (accessed on 7 April 2021).
- Dimitriadou, E.; Hornik, K.; Hornik, M.K. Package ‘cclust’. 2017. Available online: https://mran.microsoft.com/snapshot/2017-02-04/web/packages/cclust/index.html (accessed on 7 April 2021).
- Scrucca, L.; Fop, M.; Murphy, T.B.; Raftery, A.E. mclust 5: Clustering, classification and density estimation using Gaussian finite mixture models. R J. 2016, 8, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culhane, A.C.; Thioulouse, J.; Perriere, G.; Higgins, D.G. MADE4: An R package for multivariate analysis of gene expression data. Bioinformatics 2005, 21, 2789–2790. [Google Scholar] [CrossRef] [Green Version]
- Vujkovic-Cvijin, I.; Sklar, J.; Jiang, L.; Natarajan, L.; Knight, R.; Belkaid, Y. Host variables confound gut microbiota studies of human disease. Nature 2020, 587, 448–454. [Google Scholar] [CrossRef]
- Candela, M.; Biagi, E.; Soverini, M.; Consolandi, C.; Quercia, S.; Severgnini, M.; Peano, C.; Turroni, S.; Rampelli, S.; Pozzilli, P.; et al. Modulation of gut microbiota dysbioses in type 2 diabetic patients by macrobiotic Ma-Pi 2 diet. Br. J. Nutr. 2016, 116, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Smits, L.P.; Kootte, R.S.; Levin, E.; Prodan, A.; Fuentes, S.; Zoetendal, E.G.; Wang, Z.; Levison, B.S.; Cleophas, M.C.P.; Kemper, E.M.; et al. Effect of Vegan Fecal Microbiota Transplantation on Carnitine- and Choline-Derived Trimethylamine-N-Oxide Production and Vascular Inflammation in Patients With Metabolic Syndrome. J. Am. Heart Assoc. 2018, 7. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojarvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916. [Google Scholar] [CrossRef]
- Wu, T.R.; Lin, C.S.; Chang, C.J.; Lin, T.L.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Lu, C.C.; Young, J.D.; Lai, H.C. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 2019, 68, 248–262. [Google Scholar] [CrossRef]
- Duvallet, C.; Gibbons, S.M.; Gurry, T.; Irizarry, R.A.; Alm, E.J. Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat. Commun. 2017, 8, 1784. [Google Scholar] [CrossRef] [Green Version]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Ozato, N.; Saito, S.; Yamaguchi, T.; Katashima, M.; Tokuda, I.; Sawada, K.; Katsuragi, Y.; Kakuta, M.; Imoto, S.; Ihara, K.; et al. Blautia genus associated with visceral fat accumulation in adults 20–76 years of age. NPJ Biofilms Microbiomes 2019, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Tavella, T.; Rampelli, S.; Guidarelli, G.; Bazzocchi, A.; Gasperini, C.; Pujos-Guillot, E.; Comte, B.; Barone, M.; Biagi, E.; Candela, M.; et al. Elevated gut microbiome abundance of Christensenellaceae, Porphyromonadaceae and Rikenellaceae is associated with reduced visceral adipose tissue and healthier metabolic profile in Italian elderly. Gut Microbes 2021, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chan, L.; Zhou, S. Trigonelline: A plant alkaloid with therapeutic potential for diabetes and central nervous system disease. Curr. Med. Chem. 2012, 19, 3523–3531. [Google Scholar] [CrossRef] [PubMed]
- Frost, F.; Storck, L.J.; Kacprowski, T.; Gartner, S.; Ruhlemann, M.; Bang, C.; Franke, A.; Volker, U.; Aghdassi, A.A.; Steveling, A.; et al. A structured weight loss program increases gut microbiota phylogenetic diversity and reduces levels of Collinsella in obese type 2 diabetics: A pilot study. PLoS ONE 2019, 14, e0219489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Micro Biol. 2016, 24, 523–524. [Google Scholar] [CrossRef]
- Fassarella, M.; Blaak, E.E.; Penders, J.; Nauta, A.; Smidt, H.; Zoetendal, E.G. Gut microbiome stability and resilience: Elucidating the response to perturbations in order to modulate gut health. Gut 2021, 70, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Naderpoor, N.; Mousa, A.; Gomez-Arango, L.F.; Barrett, H.L.; Dekker Nitert, M.; de Courten, B. Faecal Microbiota Are Related to Insulin Sensitivity and Secretion in Overweight or Obese Adults. J. Clin. Med. 2019, 8, 452. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.; Bassot, A.; Bulteau, A.L.; Pirola, L.; Morio, B. Glycine Metabolism and Its Alterations in Obesity and Metabolic Diseases. Nutrients 2019, 11, 1356. [Google Scholar] [CrossRef] [Green Version]
- Altorf-van der Kuil, W.; Brink, E.J.; Boetje, M.; Siebelink, E.; Bijlsma, S.; Engberink, M.F.; van’t Veer, P.; Tome, D.; Bakker, S.J.; van Baak, M.A.; et al. Identification of biomarkers for intake of protein from meat, dairy products and grains: A controlled dietary intervention study. Br. J. Nutr. 2013, 110, 810–822. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, C.L.; Onnerfalt, J.; Xu, J.; Molin, G.; Ahrne, S.; Thorngren-Jerneck, K. The microbiota of the gut in preschool children with normal and excessive body weight. Obesity 2012, 20, 2257–2261. [Google Scholar] [CrossRef]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.B. Dietary pattern analysis: A new direction in nutritional epidemiology. Curr. Opin. Lipidol. 2002, 13, 3–9. [Google Scholar] [CrossRef]
- Moeller, S.M.; Reedy, J.; Millen, A.E.; Dixon, L.B.; Newby, P.K.; Tucker, K.L.; Krebs-Smith, S.M.; Guenther, P.M. Dietary patterns: Challenges and opportunities in dietary patterns research an Experimental Biology workshop, April 1, 2006. J. Am. Diet. Assoc. 2007, 107, 1233–1239. [Google Scholar] [CrossRef]
- Edefonti, V.; Nicolussi, F.; Polesel, J.; Bravi, F.; Bosetti, C.; Garavello, W.; La Vecchia, C.; Bidoli, E.; Decarli, A.; Serraino, D.; et al. Nutrient-based dietary patterns and nasopharyngeal cancer: Evidence from an exploratory factor analysis. Br. J. Cancer 2015, 112, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Edefonti, V.; Randi, G.; Decarli, A.; La Vecchia, C.; Bosetti, C.; Franceschi, S.; Dal Maso, L.; Ferraroni, M. Clustering dietary habits and the risk of breast and ovarian cancers. Ann. Oncol. 2009, 20, 581–590. [Google Scholar] [CrossRef]
- Masala, G.; Ceroti, M.; Pala, V.; Krogh, V.; Vineis, P.; Sacerdote, C.; Saieva, C.; Salvini, S.; Sieri, S.; Berrino, F.; et al. A dietary pattern rich in olive oil and raw vegetables is associated with lower mortality in Italian elderly subjects. Br. J. Nutr. 2007, 98, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Pala, V.; Sieri, S.; Masala, G.; Palli, D.; Panico, S.; Vineis, P.; Sacerdote, C.; Mattiello, A.; Galasso, R.; Salvini, S.; et al. Associations between dietary pattern and lifestyle, anthropometry and other health indicators in the elderly participants of the EPIC-Italy cohort. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 186–201. [Google Scholar] [CrossRef]
- Edefonti, V.; De Vito, R.; Dalmartello, M.; Patel, L.; Salvatori, A.; Ferraroni, M. Reproducibility and Validity of A Posteriori Dietary Patterns: A Systematic Review. Adv. Nutr. 2020, 11, 293–326. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Goodrich, J.K.; Davenport, E.R.; Beaumont, M.; Jackson, M.A.; Knight, R.; Ober, C.; Spector, T.D.; Bell, J.T.; Clark, A.G.; Ley, R.E. Genetic Determinants of the Gut Microbiome in UK Twins. Cell Host Microbe 2016, 19, 731–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Rowan, F.; Docherty, N.G.; Murphy, M.; Murphy, B.; Calvin Coffey, J.; O’Connell, P.R. Desulfovibrio bacterial species are increased in ulcerative colitis. Dis. Colon Rectum 2010, 53, 1530–1536. [Google Scholar] [CrossRef]
- Ahn, J.; Sinha, R.; Pei, Z.; Dominianni, C.; Wu, J.; Shi, J.; Goedert, J.J.; Hayes, R.B.; Yang, L. Human gut microbiome and risk for colorectal cancer. J. Natl. Cancer Inst. 2013, 105, 1907–1911. [Google Scholar] [CrossRef] [Green Version]
- Kaakoush, N.O. Insights into the Role of Erysipelotrichaceae in the Human Host. Front. Cell Infect. Micro Biol. 2015, 5, 84. [Google Scholar] [CrossRef] [Green Version]
- Granchi, D.; Baldini, N.; Ulivieri, F.M.; Caudarella, R. Role of Citrate in Pathophysiology and Medical Management of Bone Diseases. Nutrients 2019, 11, 2576. [Google Scholar] [CrossRef] [Green Version]
- Pallister, T.; Jackson, M.A.; Martin, T.C.; Zierer, J.; Jennings, A.; Mohney, R.P.; MacGregor, A.; Steves, C.J.; Cassidy, A.; Spector, T.D.; et al. Hippurate as a metabolomic marker of gut microbiome diversity: Modulation by diet and relationship to metabolic syndrome. Sci. Rep. 2017, 7, 13670. [Google Scholar] [CrossRef] [Green Version]
- Palau-Rodriguez, M.; Tulipani, S.; Isabel Queipo-Ortuno, M.; Urpi-Sarda, M.; Tinahones, F.J.; Andres-Lacueva, C. Metabolomic insights into the intricate gut microbial-host interaction in the development of obesity and type 2 diabetes. Front. Micro Biol. 2015, 6, 1151. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 69, 1218–1228. [Google Scholar] [CrossRef] [Green Version]
- Haro, C.; Montes-Borrego, M.; Rangel-Zuniga, O.A.; Alcala-Diaz, J.F.; Gomez-Delgado, F.; Perez-Martinez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Tinahones, F.J.; Landa, B.B.; et al. Two Healthy Diets Modulate Gut Microbial Community Improving Insulin Sensitivity in a Human Obese Population. J. Clin. Endocrinol. Metab. 2016, 101, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.D.; Nguyen, L.H.; Li, Y.; Yan, Y.; Ma, W.; Rinott, E.; Ivey, K.L.; Shai, I.; Willett, W.C.; Hu, F.B.; et al. The gut microbiome modulates the protective association between a Mediterranean diet and cardiometabolic disease risk. Nat. Med. 2021, 27, 333–343. [Google Scholar] [CrossRef]
- Tenorio-Jimenez, C.; Martinez-Ramirez, M.J.; Gil, A.; Gomez-Llorente, C. Effects of Probiotics on Metabolic Syndrome: A Systematic Review of Randomized Clinical Trials. Nutrients 2020, 12, 124. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| All (n = 60) | SA-Group (n = 33) | PROB-Group (n = 27) | p | ||||
|---|---|---|---|---|---|---|---|
| Median | [Min–Max] | Median | [Min–Max] | Median | [Min–Max] | ||
| Gender, n (%) | 0.176 | ||||||
| Male | 13 | (21.7) | 5 | (15.2) | 8 | (29.6) | |
| Female | 47 | (78.3) | 28 | (84.8) | 19 | (70.4) | |
| Smoking habit, n (%) 1 | 0.860 | ||||||
| Never smoker | 26 | (48.1) | 14 | (46.7) | 12 | (50.0) | |
| Ex-smoker | 22 | (40.7) | 12 | (40.0) | 10 | (41.7) | |
| Current smoker | 6 | (11.1) | 4 | (13.3) | 2 | (8.3) | |
| Age at enrollment, years | 46.9 | [18.3–86.4] | 52.7 | [34.6–86.4] | 45.3 | [18.3–64.2] | 0.015 |
| Weight, kg | 70.5 | [44.0–103.0] | 70.0 | [44.0–103.0] | 72.0 | [47–94.5] | 0.953 |
| Height, m | 1.65 | [1.4–1.8] | 1.7 | [1.4–1.8] | 1.7 | [1.5–1.8] | 0.183 |
| BMI, kg/m2 | 25.7 | [19.2–36.8] | 26.1 | [19.2–36.8] | 25.3 | [19.8–33.3] | 0.427 |
| Waist circumference, cm | 85.0 | [64.0–113.0] | 85.0 | [67.0–113.0] | 84.0 | [64.0–102.0] | 0.639 |
| Hip circumference, cm | 105.0 | [89.0–123.0] | 105.0 | [89.0–123] | 104.0 | [90.0–116.0] | 0.312 |
| WHR | 0.8 | [0.7–1.0] | 0.8 | [0.7–1.0] | 0.8 | [0.7–1.0] | 0.783 |
| Abdomen circumference, cm | 98.5 | [69.0–120] | 98.0 | [78.0–120.0] | 99.0 | [69.0–111.0] | 0.582 |
| Glucose, mg/dL | 82.5 | [66.0–212.0] | 83.0 | [66.0–212.0] | 82.0 | [72.0–103.0] | 0.271 |
| Cholesterol, mg/dL 1 | 193.0 | [136.0–269.0] | 190.0 | [136.0–269.0] | 195.0 | [139.0–269.0] | 0.345 |
| HDL, mg/dL 1 | 59.0 | [31.0–94.0] | 65.0 | [31.0–94.0] | 55.0 | [34.0–86.0] | 0.061 |
| LDL, mg/dL 1 | 113.0 | [55.0–171.0] | 106.0 | [67.0–171.0] | 119.0 | [55.0–167.0] | 0.064 |
| Triglycerides, mg/dL 1 | 90.0 | [43.0–365.0] | 91.5 | [43.0–365.0] | 90.0 | [44.0–243.0] | 0.879 |
| Cortisol, µg/L 1 | 125.0 | [61.0–268.0] | 124.5 | [68.0–206.0] | 129.0 | [61.0–268.0] | 0.744 |
| Insulin, mU/L 1 | 9.2 | [3.0–93.3] | 8.9 | [3.0–28.2] | 10.1 | [5.1–93.3] | 0.169 |
| Systolic BP, mmHg 1 | 120.0 | [97.0–155.0] | 120.0 | [100.0–155.0] | 115.0 | [97.0–150.0] | 0.072 |
| Diastolic BP, mmHg 1 | 70.0 | [55.0–90.0] | 70.0 | [60.0–90.0] | 70.0 | [55.0–90.0] | 1.000 |
| MS, n (%) 1 | 0.488 | ||||||
| No | 50 | (84.7) | 26 | (81.2) | 24 | (88.9) | |
| Yes | 9 | (15.3) | 6 | (18.8) | 3 | (11.1) | |
| INF-γ 1 | 0 | [0.0–7.5] | 0 | [0–2.8] | 0 | [0–7.5] | 0.646 |
| IL-6 1 | 1.3 | [0.0–254.4] | 1.3 | [0–55.5] | 1.6 | [0–254.4] | 0.613 |
| IL-10 1 | 0.3 | [0.0–15.0] | 0 | [0–15.0] | 0.6 | [0–4.5] | 0.087 |
| IL-17A 1 | 0 | [0.0–18.8] | 0 | [0–2.6] | 0.8 | [0–18.8] | 0.004 |
| TNFα 1 | 0.2 | [0.0–67.9] | 0 | [0–67.9] | 0.3 | [0–11.9] | 0.419 |
| IMI categories, n (%) 1 | 0.265 | ||||||
| 0–3 | 24 | (40.7) | 16 | (50.0) | 8 | (29.6) | |
| 4–5 | 23 | (39.0) | 10 | (31.3) | 13 | (48.2) | |
| 6–8 | 12 | (20.3) | 6 | (18.7) | 6 | (22.2) | |
| Low Adherence (Index Range: 0–3) (n = 24, 40.7%) | Medium Adherence (Index Range: 4–5) (n = 23, 39.0%) | High Adherence (Index Range 6–8) 2 (n = 12, 20.3%) | p | ||||
|---|---|---|---|---|---|---|---|
| Median | [Min–Max] | Median | [Min–Max] | Median | [Min–Max] | ||
| Gender, n (%) | 0.848 | ||||||
| Male | 5 | (20.8) | 6 | (26.1) | 2 | (16.7) | |
| Female | 19 | (79.0) | 17 | (73.9) | 10 | (83.3) | |
| Smoking habit, n (%) 1 | 0.797 | ||||||
| Never smoker | 11 | (50.0) | 10 | (47.6) | 5 | (45.5) | |
| Ex-smoker | 8 | (36.4) | 10 | (47.6) | 4 | (36.4) | |
| Current smoker | 3 | (13.6) | 1 | (4.8) | 2 | (18.2) | |
| Age at enrollment, years | 46.2 | [18.3–86.4] | 53.7 | [35.4–84.8] | 46.1 | [40.5–55.0] | 0.389 |
| Weight, kg | 73.5 | [47.0–103.0] | 67.0 | [44.0–94.5] | 66.5 | [56.0–84.5] | 0.431 |
| Height, m | 1.7 | [1.43–1.8] | 1.7 | [1.4–1.8] | 1.7 | [1.58–1.8] | 0.949 |
| BMI, kg/m2 | 26.5 | [19.2–36.8] | 25.4 | [20.0–31.9] | 24.7 | [20.3–31.8] | 0.380 |
| Waist circumference, cm | 85.5 | [64.0–113.0] | 83.0 | [70.0–105.0] | 84.0 | [68.0–101.0] | 0.543 |
| Hip circumference, cm | 107.0 | [90.0–123.0] | 104.0 | [89.0–115.0] | 101.5 | [90.0–122.0] | 0.391 |
| WHR | 0.8 | [0.7–1.0] | 0.8 | [0.7–1.0] | 0.8 | [0.7–1.0] | 0.952 |
| Abdomen circumference, cm | 99.5 | [69.0–120.0] | 98.0 | [81.0–110.0] | 97.0 | [75–115.0] | 0.501 |
| Glucose, mg/dL | 86.5 | [72.0–212.0] | 82.0 | [72.0–123.0] | 79.0 | [66.0–88.0] | 0.051 |
| Cholesterol, mg/dL 1 | 193.5 | [136.0–269] | 193.5 | [139.0–228.0] | 185.0 | [145–269.0] | 0.486 |
| HDL, mg/dL 1 | 55.5 | [31.0–94.0] | 58.0 | [34.0–79.0] | 63.0 | [41.0–82.0] | 0.176 |
| LDL, mg/dL 1 | 117.0 | [67.0–171.0] | 111.0 | [67.0–157.0] | 110.5 | [55.0–167.0] | 0.535 |
| Triglycerides, mg/dL 1 | 96.0 | [44.0–365.0] | 90.0 | [50.0–267.0] | 90.5 | [43.0–164.0] | 0.740 |
| Cortisol, µg/L 1 | 119.5 | [61.0–268.0] | 141.5 | [85.0–226.0] | 121.0 | [68.0–254.0] | 0.471 |
| Insulin, mU/L 1 | 8.9 | [3.0–93.3] | 10.0 | [5.4–35.0] | 8.1 | [3.2–27.7] | 0.334 |
| Systolic BP, mmHg 1 | 120.0 | [100.0–140.0] | 118.0 | [97.0–155.0] | 120.0 | [107.0–130.0] | 0.984 |
| Diastolic BP, mmHg 1 | 72.5 | [60.0–90] | 70.0 | [55.0–90.0] | 70.0 | [60.0–90.0] | 0.514 |
| MS, n (%) 1 | 0.719 | ||||||
| No | 19 | (79.2) | 19 | (86.4) | 11 | (91.7) | |
| Yes | 5 | (20.8) | 3 | (13.6) | 1 | (8.3) | |
| INF-γ 1 | 0 | [0–2.3] | 0 | [0–2.8] | 0.2 | [0–7.5] | 0.151 |
| IL-6 1 | 1.0 | [0–55.5] | 1.6 | [0–5.7] | 1.3 | [0–254.4] | 0.489 |
| IL-10 1 | 0.1 | [0–15.0] | 0.4 | [0–1.9] | 0.7 | [0–4.5] | 0.520 |
| IL-17A 1 | 0 | [0–3.6] | 0 | [0–3.3] | 0.5 | [0–18.8] | 0.411 |
| TNFα 1 | 0 | [0–11.0] | 0.345 | [0–5.3] | 0.6 | [0–11.9] | |
| Nutrient | Dietary Pattern | ||
|---|---|---|---|
| Animal Products | Vitamins and Fiber | Regional | |
| Animal protein | 0.96 | - | - |
| Vegetable protein | 0.34 | 0.48 | 0.73 |
| Cholesterol | 0.88 | 0.15 | 0.13 |
| Saturated fatty acids | 0.80 | 0.43 | 0.13 |
| Monounsaturated fatty acids | 0.48 | 0.66 | 0.42 |
| Linoleic acid | 0.64 | 0.40 | 0.44 |
| Linolenic acid | 0.49 | 0.61 | 0.40 |
| Other polyunsaturated fatty acids | - | - | 0.70 |
| Soluble carbohydrates | 0.43 | 0.66 | 0.29 |
| Starch | 0.46 | 0.19 | 0.70 |
| Sodium | 0.78 | 0.30 | 0.27 |
| Calcium | 0.66 | 0.54 | - |
| Potassium | 0.61 | 0.72 | 0.25 |
| Phosphorus | 0.80 | 0.44 | 0.31 |
| Iron | 0.54 | 0.57 | 0.57 |
| Zinc | 0.82 | 0.29 | 0.42 |
| Thiamin (vitamin B1) | 0.69 | 0.44 | 0.41 |
| Riboflavin (vitamin B2) | 0.70 | 0.43 | - |
| Vitamin B6 | 0.73 | 0.46 | 0.32 |
| Total folate | 0.33 | 0.77 | 0.44 |
| Niacin | 0.83 | 0.29 | 0.27 |
| Vitamin C | 0.21 | 0.88 | - |
| Retinol | 0.72 | - | 0.17 |
| Beta-carotene | - | 0.87 | 0.24 |
| Vitamin D | 0.79 | 0.10 | - |
| Vitamin E | 0.28 | 0.76 | 0.47 |
| Total fiber | 0.22 | 0.80 | 0.47 |
| Proportion of explained variance (%) | 37.99 | 27.28 | 15.09 |
| Cumulative explained variance (%) | 37.99 | 65.27 | 80.36 |
| Cluster Name 2 | Cluster Size | Cluster Center (Medoid) | ||
|---|---|---|---|---|
| Animal Products | Vitamins and Fiber | Regional | ||
| C1-High consumers | 11 | 0.11 | 0.24 | 0.883 |
| C2-Low consumers | 19 | −0.72 | −0.59 | −0.41 |
| C3-Omnivorous with meat prevalence | 18 | 0.813 | −0.30 | −0.30 |
| C4-Omnivorous with plant-based foods prevalence | 11 | −0.69 | 0.703 | −0.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turroni, S.; Petracci, E.; Edefonti, V.; Giudetti, A.M.; D’Amico, F.; Paganelli, L.; Giovannetti, G.; Del Coco, L.; Fanizzi, F.P.; Rampelli, S.; et al. Effects of a Diet Based on Foods from Symbiotic Agriculture on the Gut Microbiota of Subjects at Risk for Metabolic Syndrome. Nutrients 2021, 13, 2081. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13062081
Turroni S, Petracci E, Edefonti V, Giudetti AM, D’Amico F, Paganelli L, Giovannetti G, Del Coco L, Fanizzi FP, Rampelli S, et al. Effects of a Diet Based on Foods from Symbiotic Agriculture on the Gut Microbiota of Subjects at Risk for Metabolic Syndrome. Nutrients. 2021; 13(6):2081. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13062081
Chicago/Turabian StyleTurroni, Silvia, Elisabetta Petracci, Valeria Edefonti, Anna M. Giudetti, Federica D’Amico, Lisa Paganelli, Giusto Giovannetti, Laura Del Coco, Francesco P. Fanizzi, Simone Rampelli, and et al. 2021. "Effects of a Diet Based on Foods from Symbiotic Agriculture on the Gut Microbiota of Subjects at Risk for Metabolic Syndrome" Nutrients 13, no. 6: 2081. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13062081