
Protective Effects Induced by a Hydroalcoholic Allium sativum Extract in Isolated Mouse Heart


 , , , , ,
, , , , ,  ,
,  , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Garlic Extract
2.2. Color Analysis
2.3. GC–MS Analysis of VOC Fraction
2.3.1. HS-SPME/GC–MS of Garlic Powder
2.3.2. GC–MS of Garlic Hydroalcoholic Extract
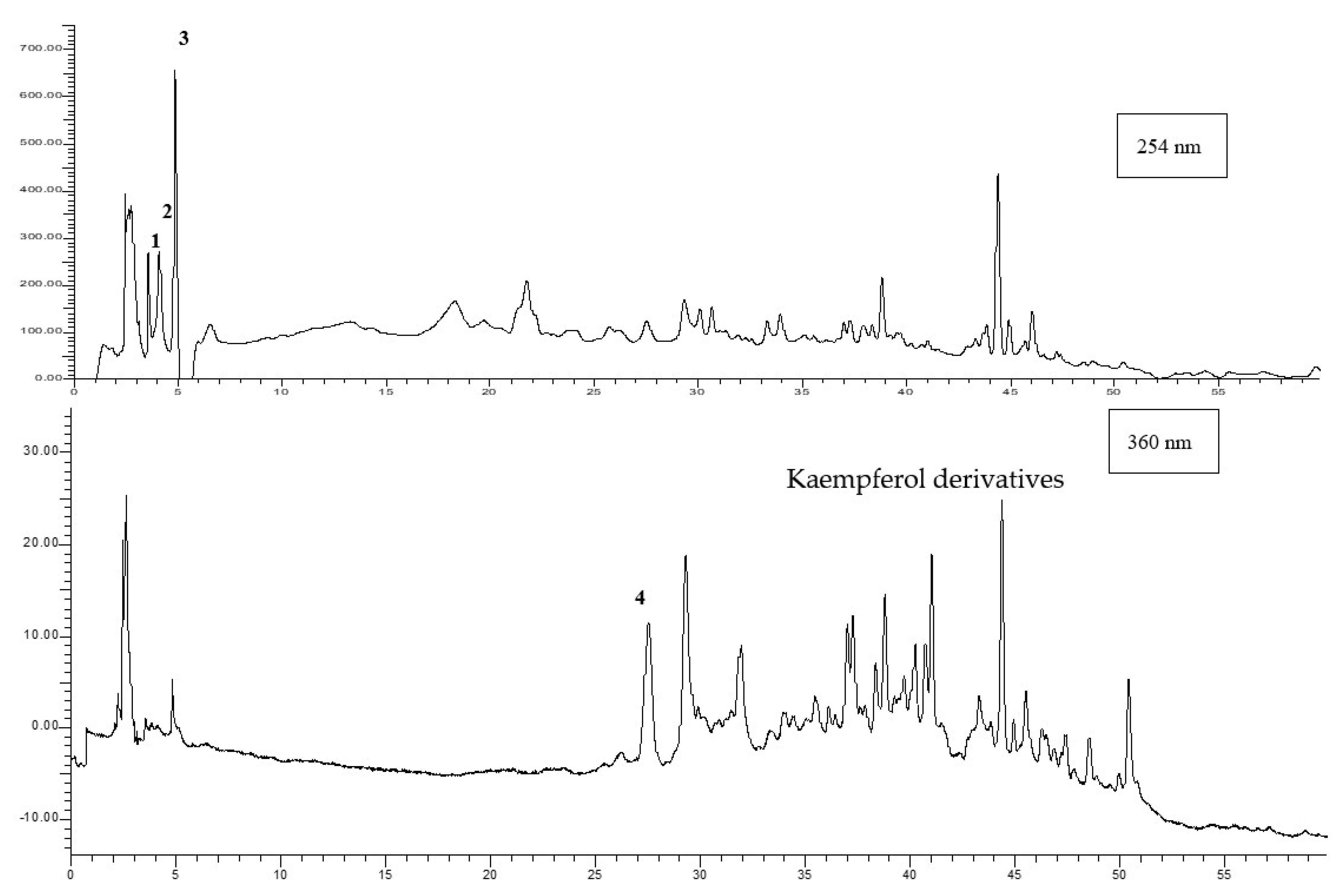
2.4. HPLC–DAD Analyses
2.5. Toxicological and Pharmacological Studies
2.5.1. Cell Cultures
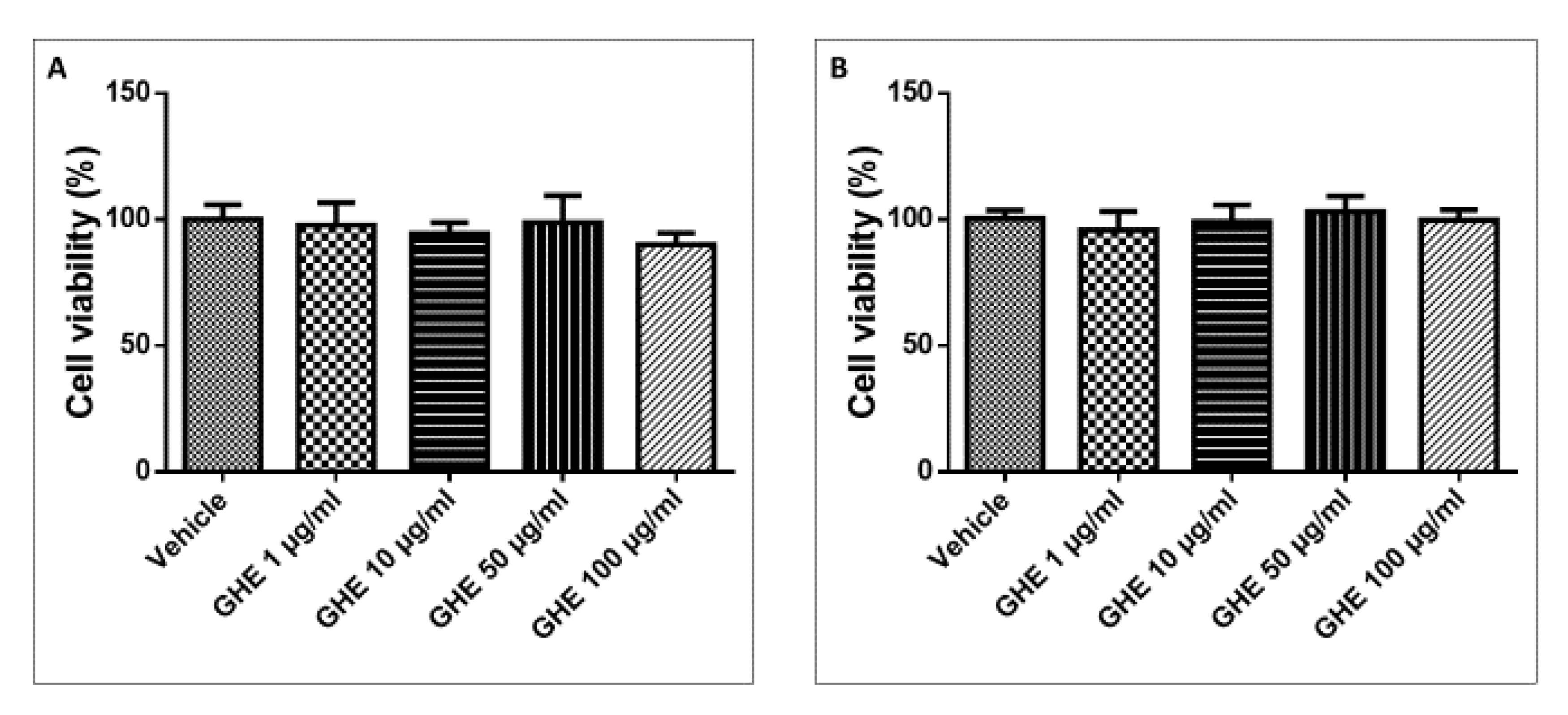
2.5.2. Assessment of Cell Toxicity by Garlic Hydroalcoholic Extract in 24 h and 48 h
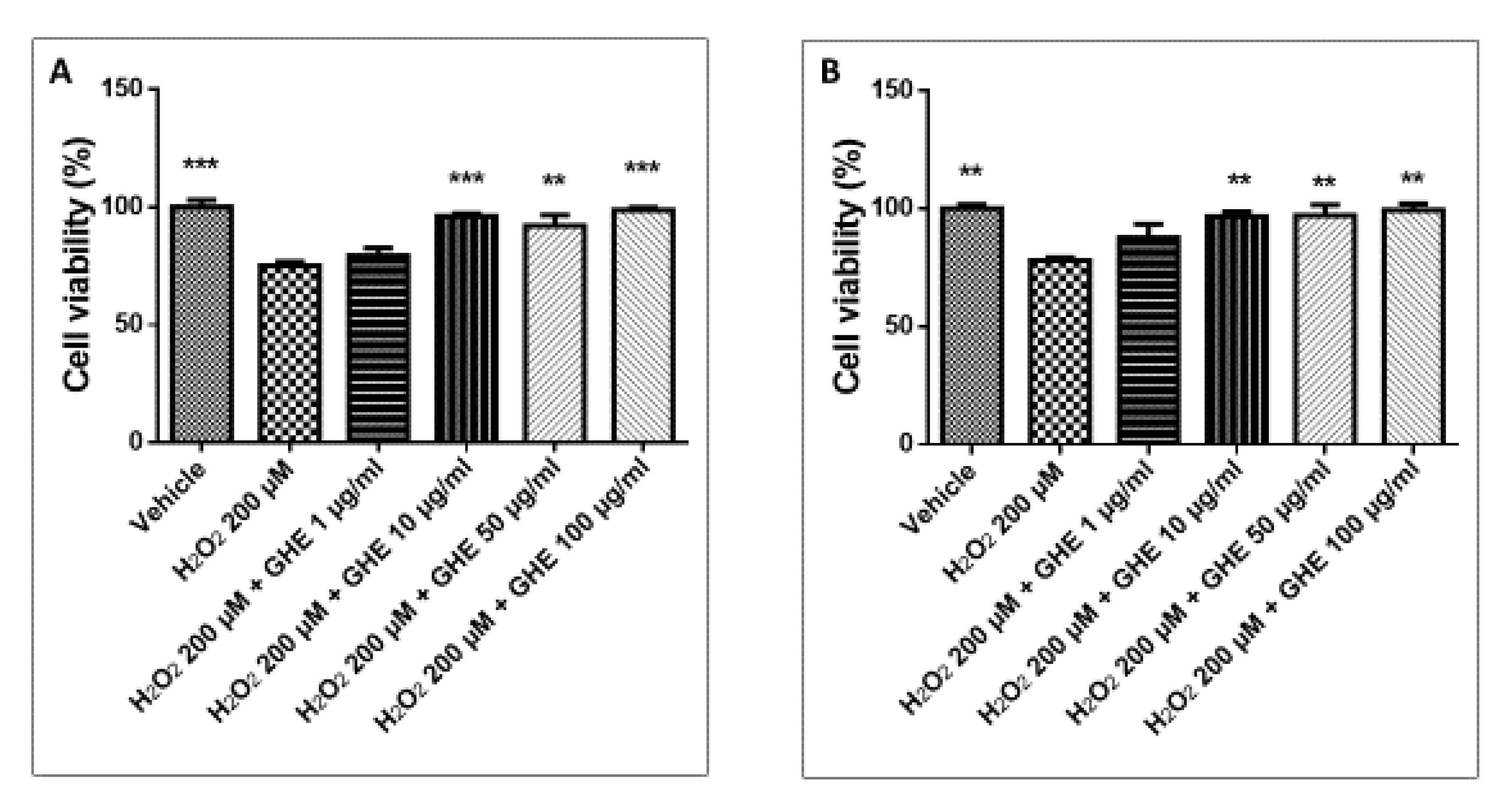
2.5.3. Evaluation of Cell Viability Preservation by Garlic Hydroalcoholic Extract against H2O2-Induced Cell Damage
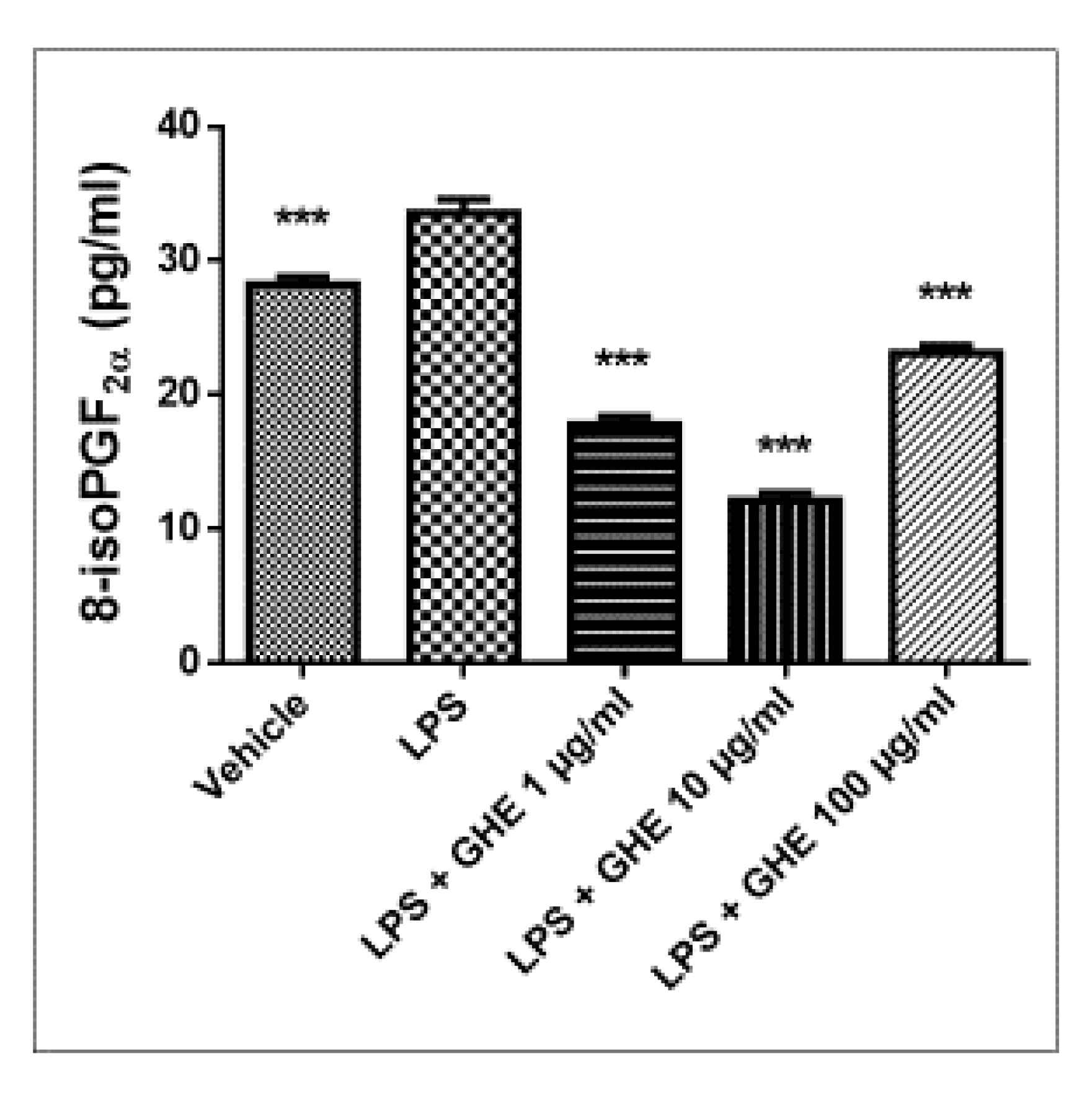
2.5.4. Ex Vivo Studies
2.6. Bioinformatics
2.7. Statistical Analysis
3. Results and Discussion
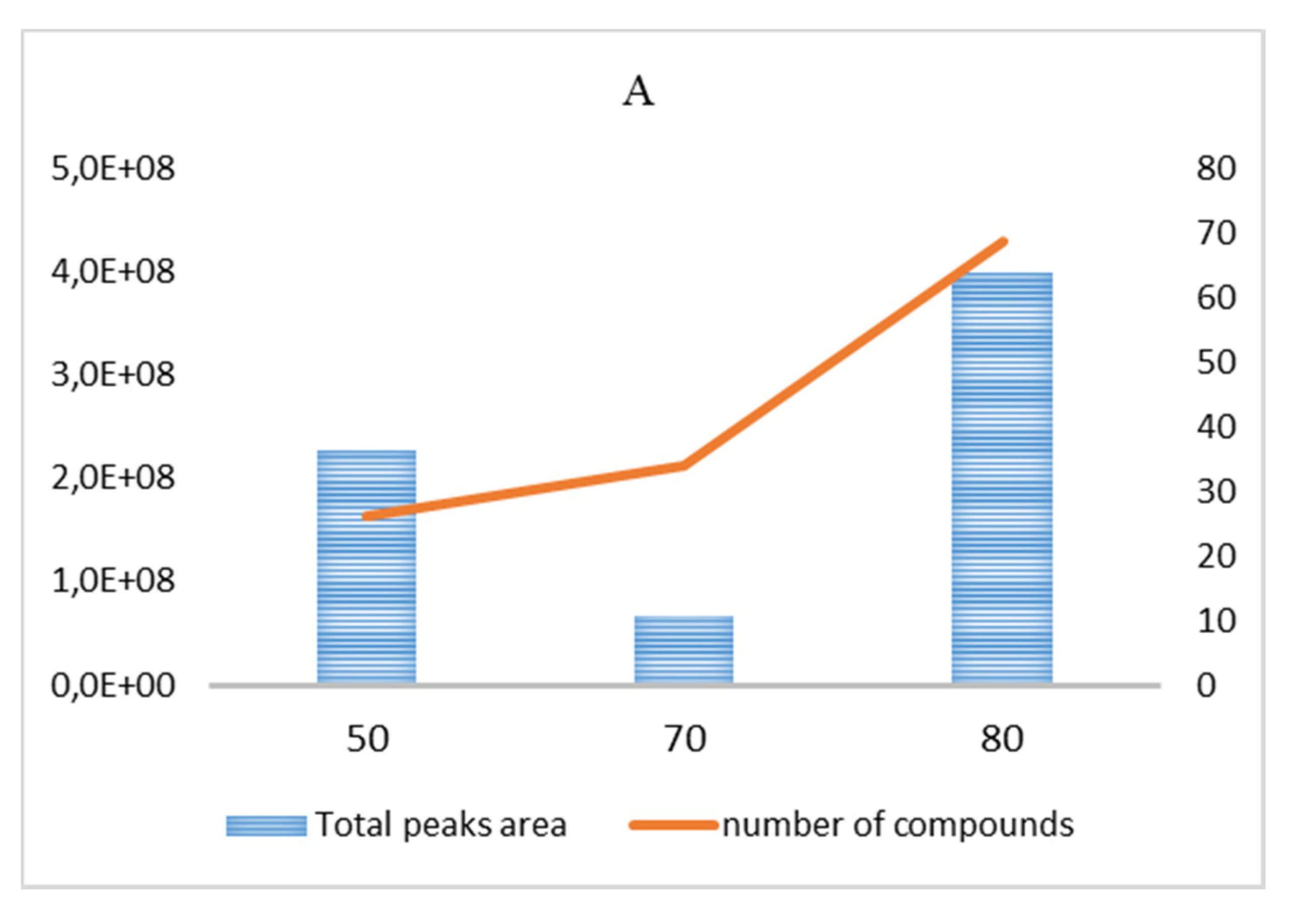
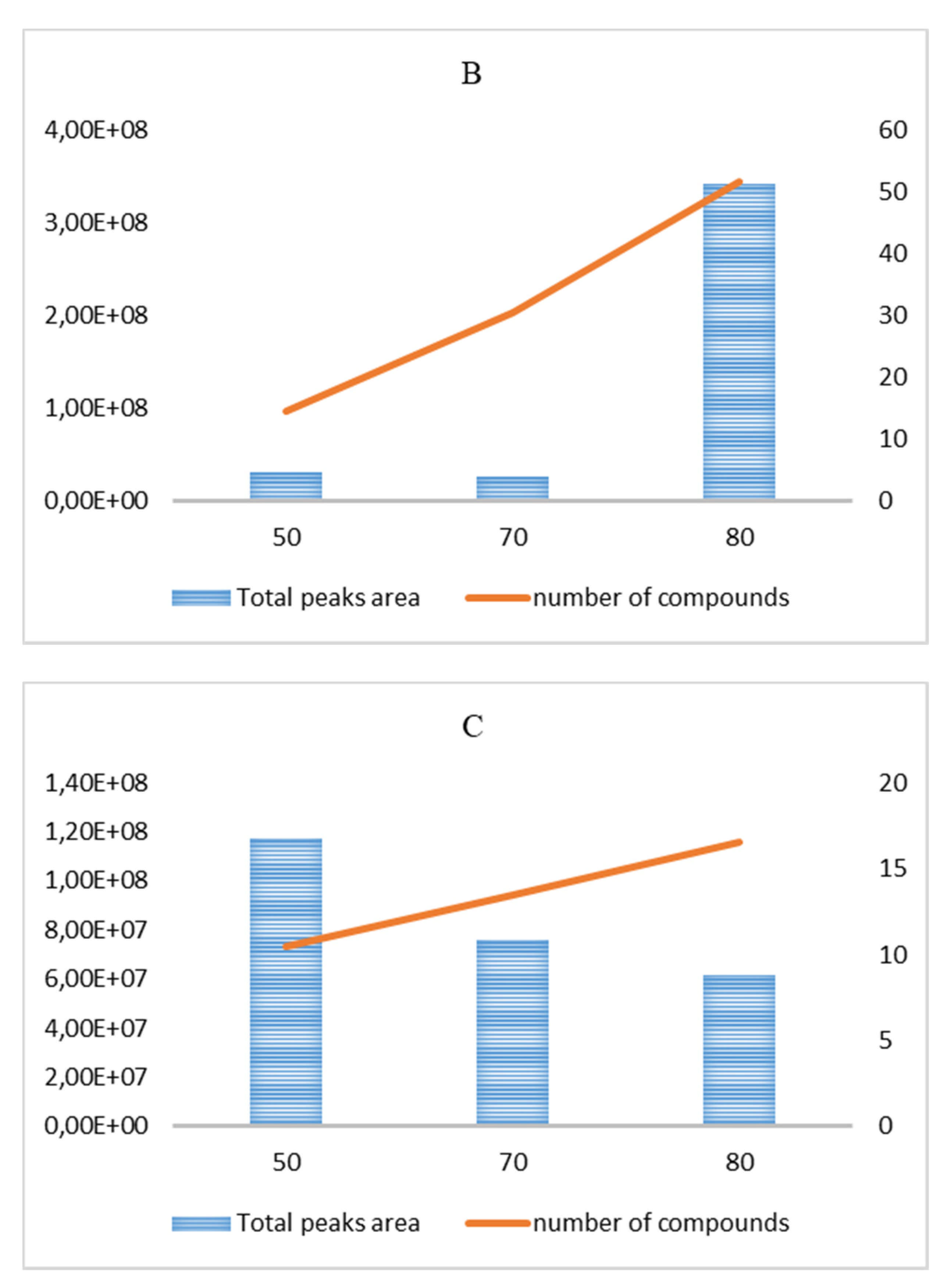
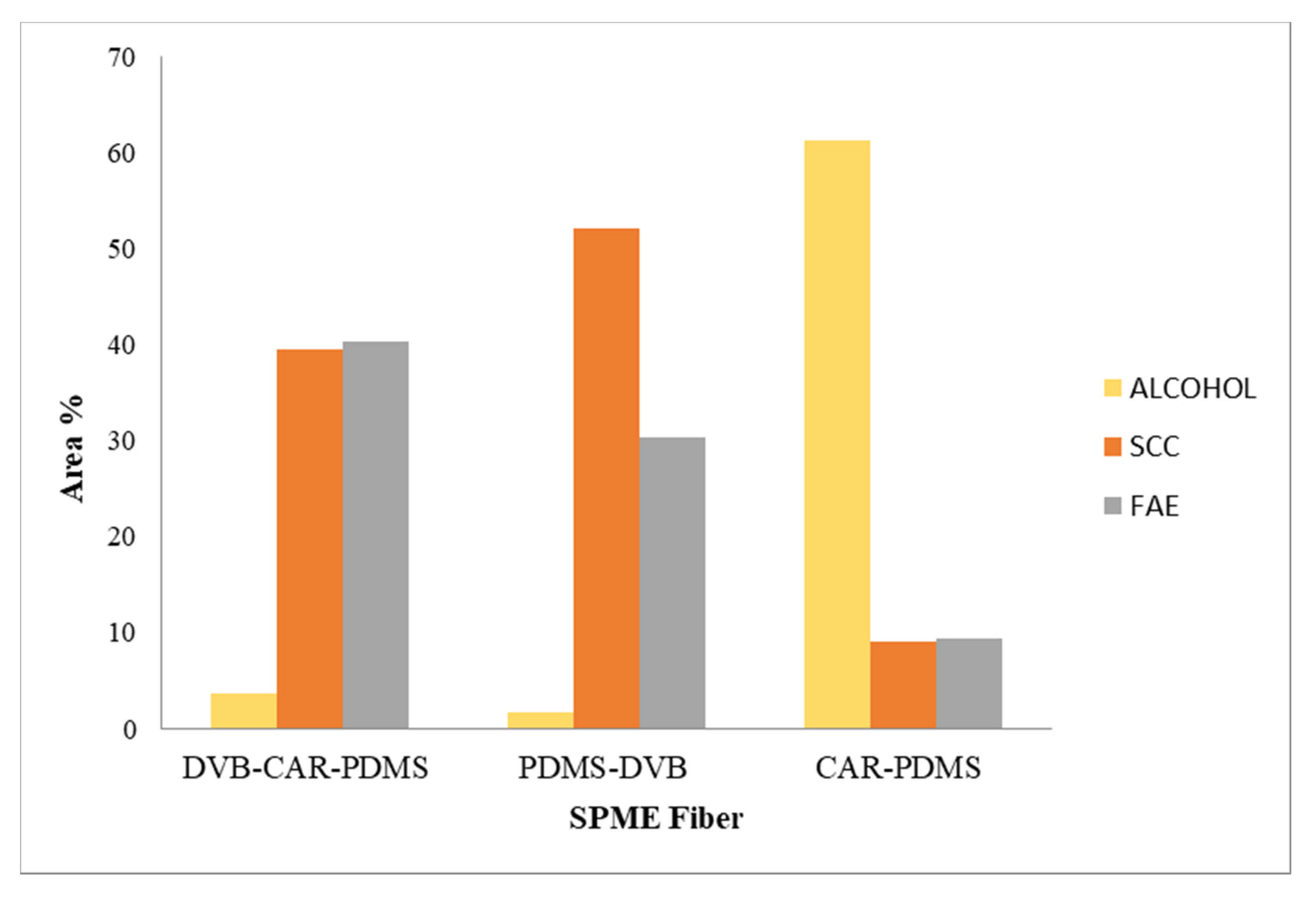
3.1. GC–MS Analysis of VOC Fraction
3.1.1. Garlic Powder
3.1.2. Garlic Hydroalcoholic Extract
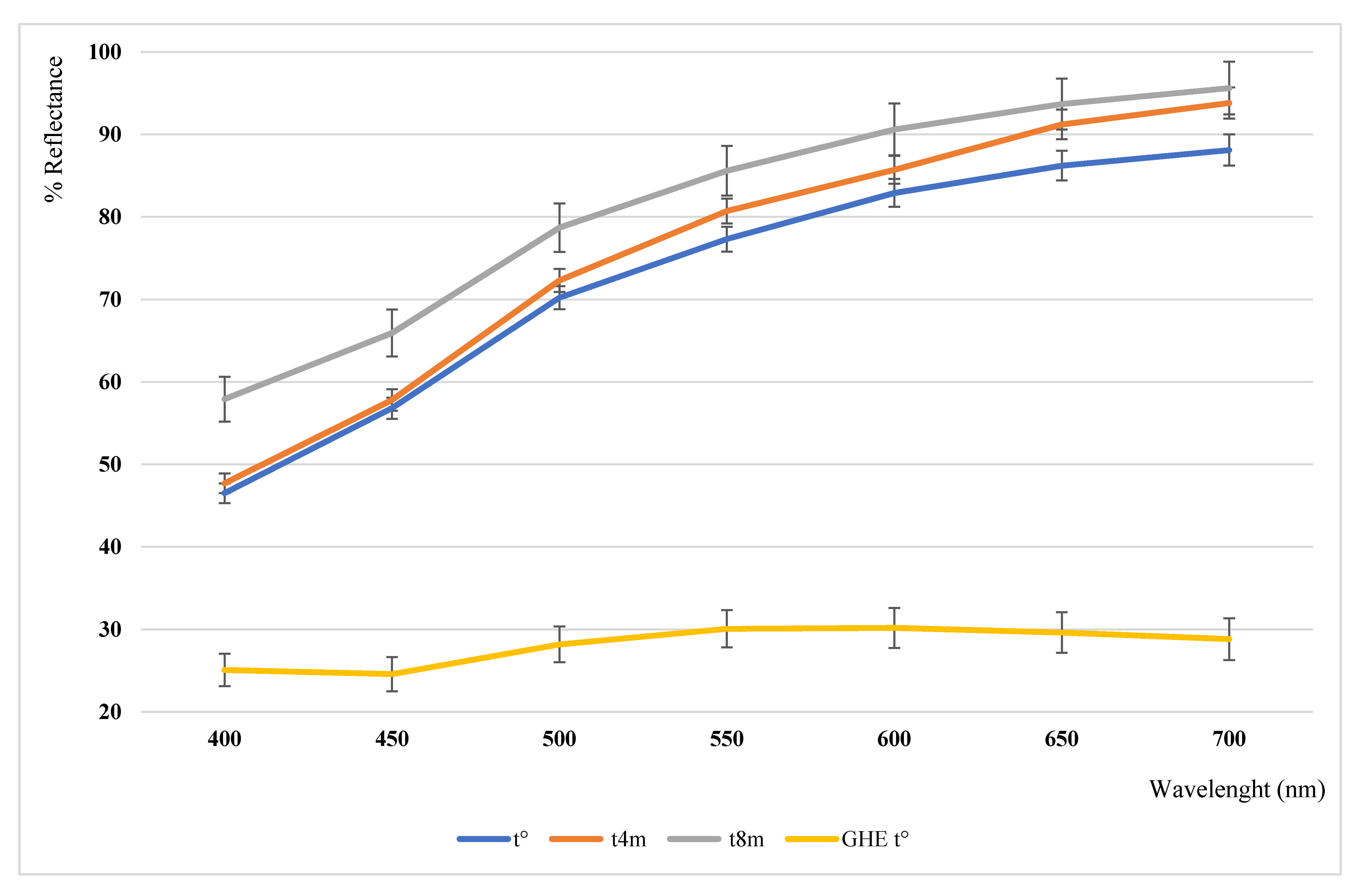
3.2. Colorimetric Analysis
3.3. HPLC–DAD Analysis
3.4. Bioinformatics
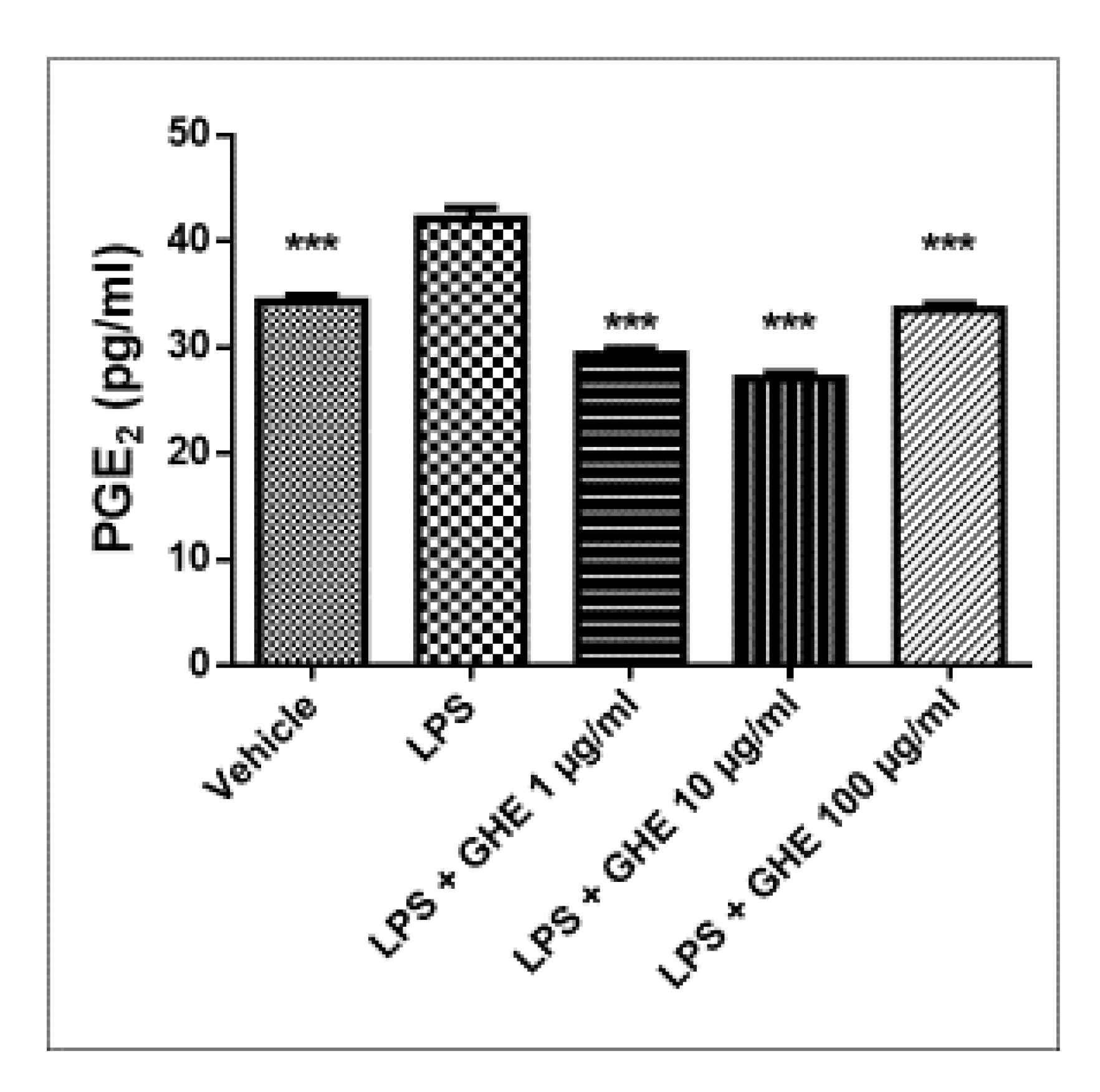
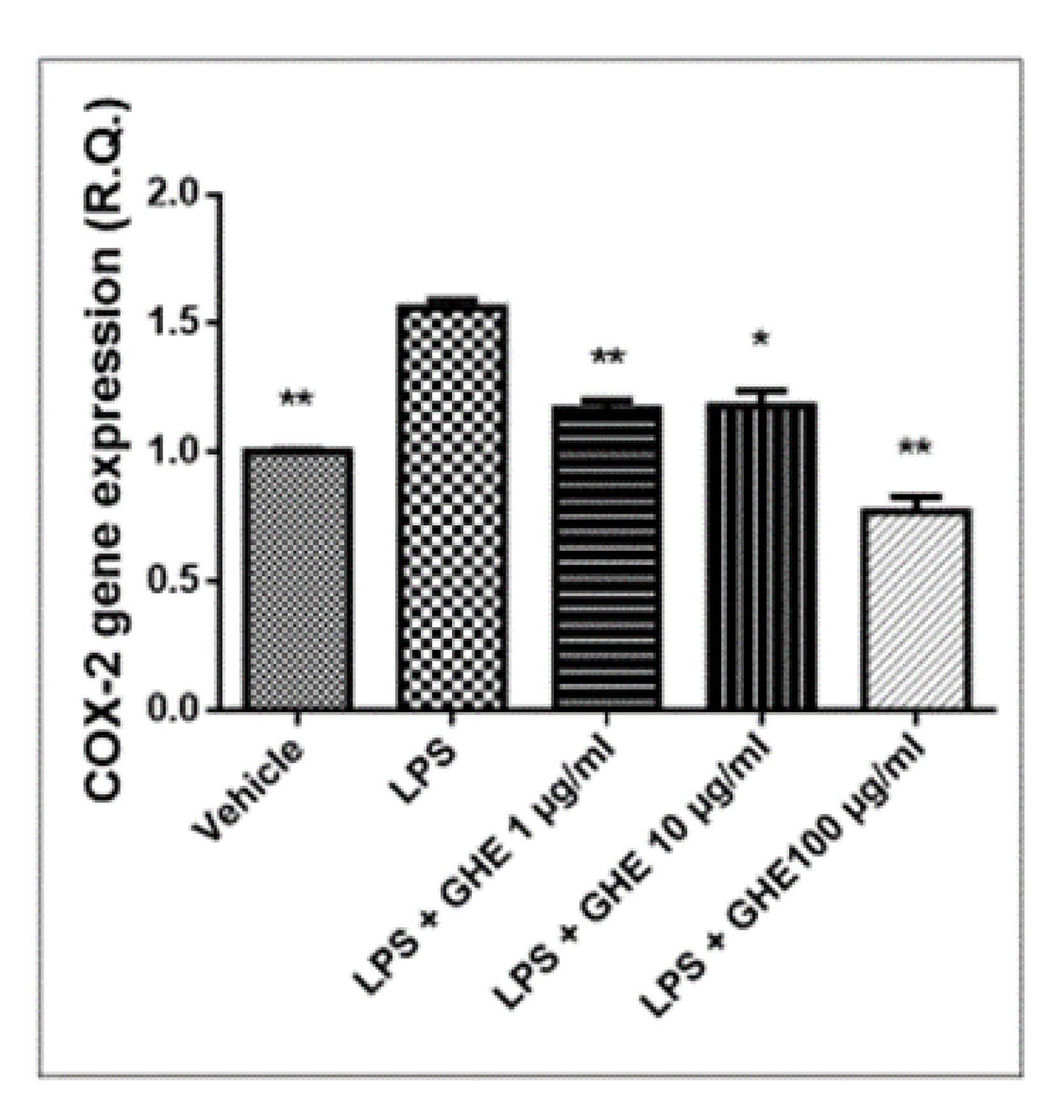
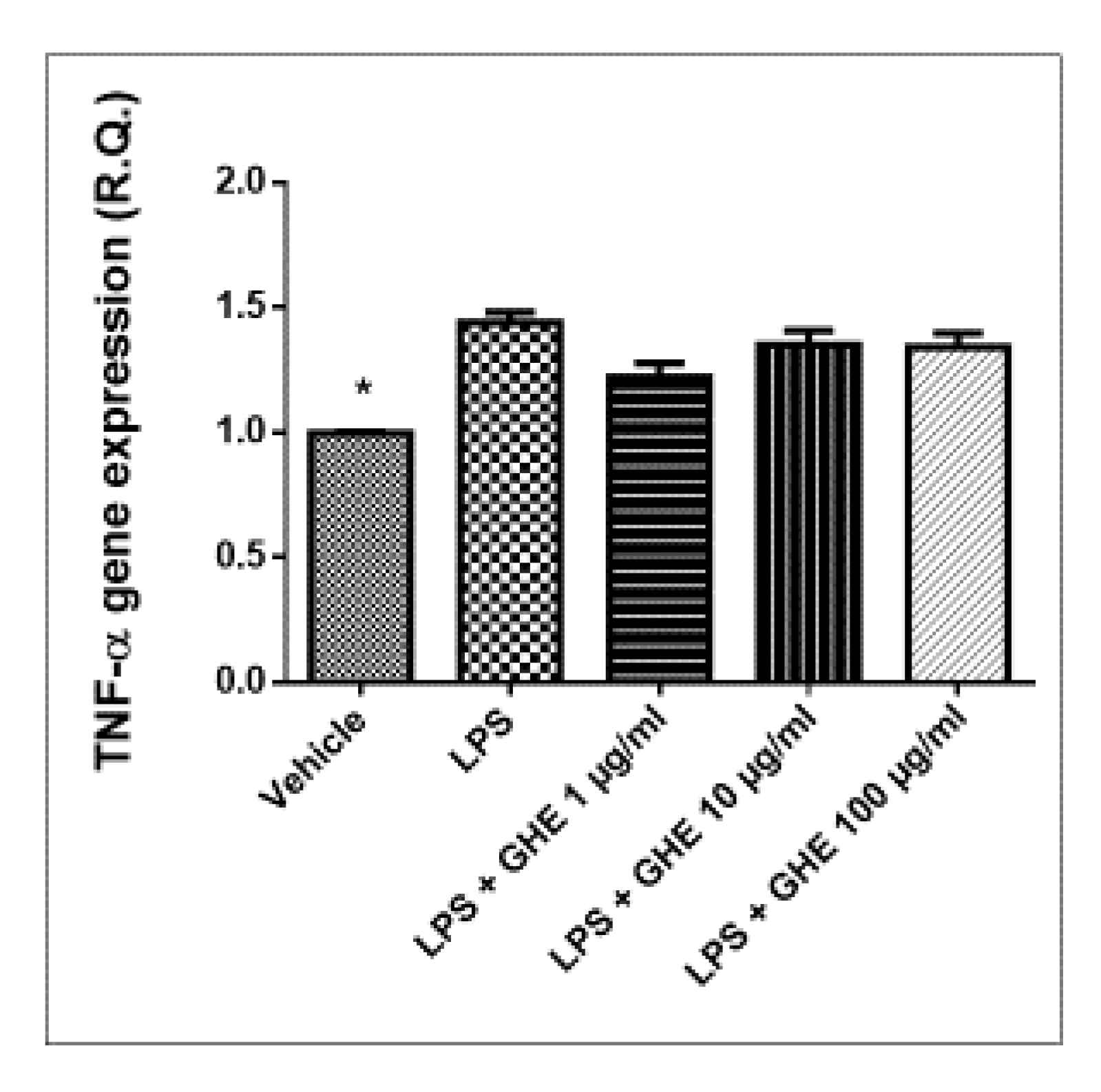
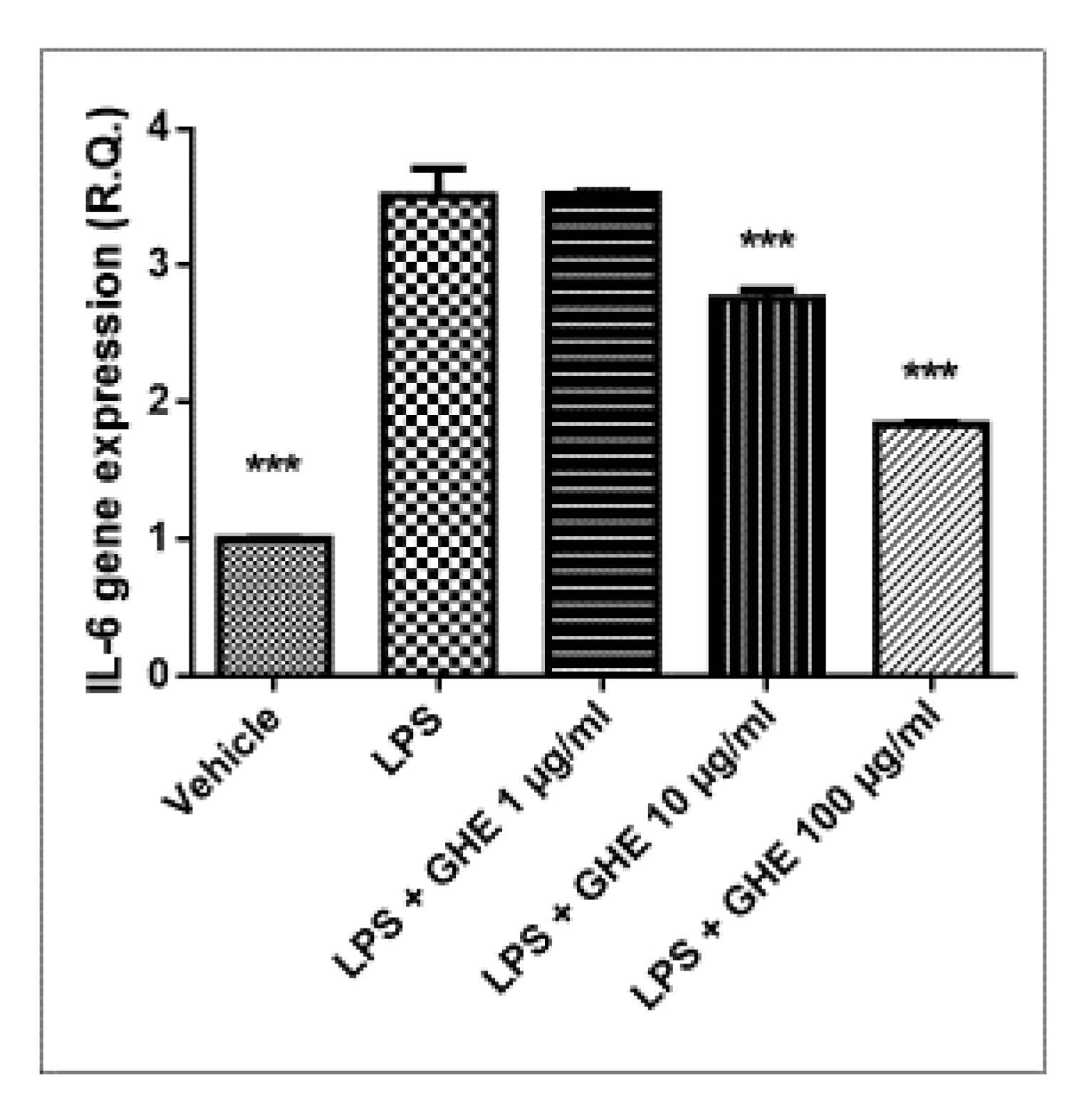
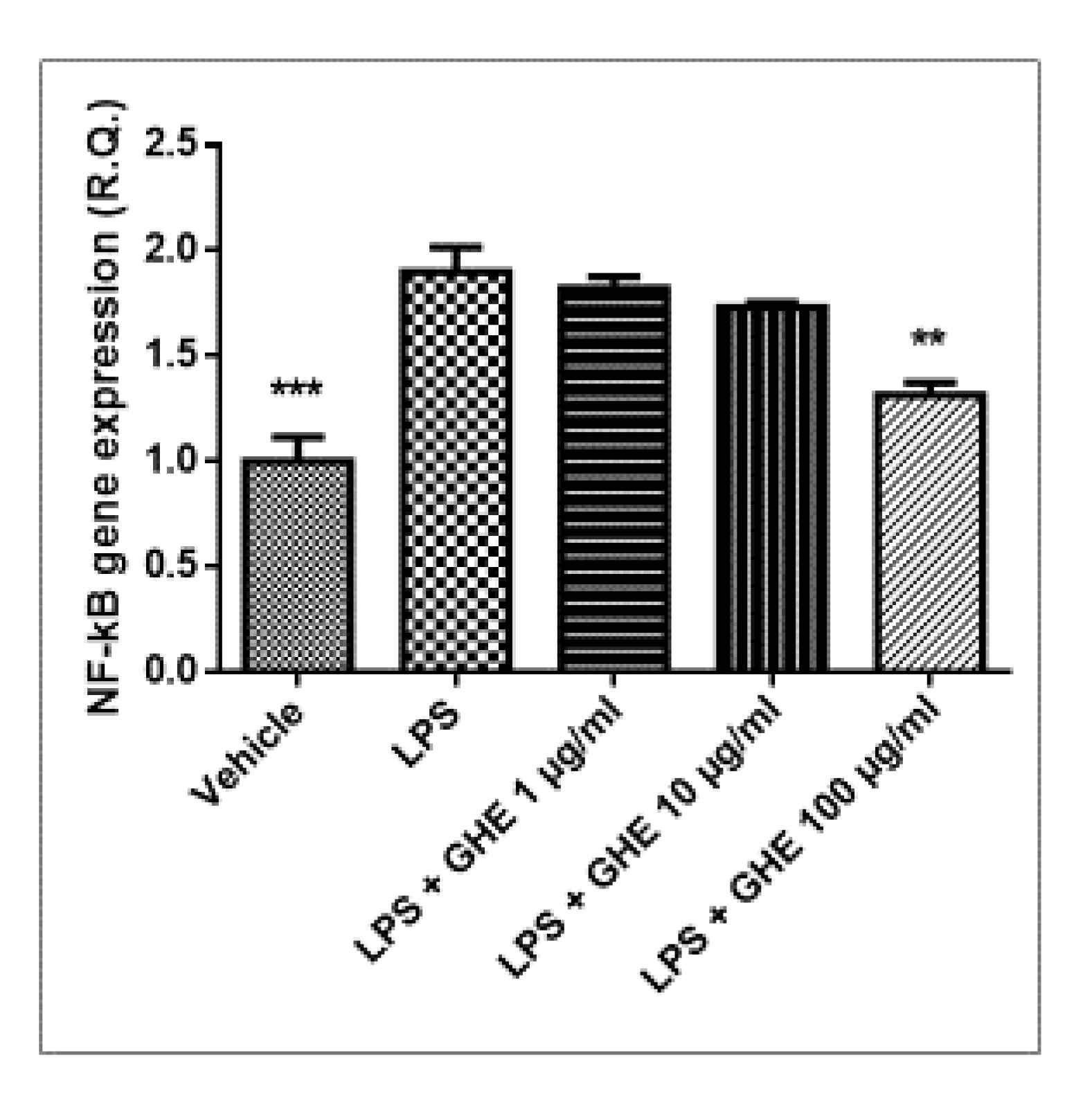
3.5. Toxicological and Pharmacological Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ansary, J.; Forbes-Hernández, T.Y.; Gil, E.; Cianciosi, D.; Zhang, J.; Elexpuru-Zabaleta, M.; Simal-Gandara, J.; Giampieri, F.; Battino, M. Potential Health Benefit of Garlic Based on Human Intervention Studies: A Brief Overview. Antioxidants 2020, 9, 619. [Google Scholar] [CrossRef]
- Leelarungrayub, N.; Rattanapanone, V.; Chanarat, N.; Gebicki, J.M. Quantitative evaluation of the antioxidant properties of garlic and shallot preparations. Nutrition 2006, 22, 266–274. [Google Scholar] [CrossRef]
- Durak, I.; Kavutcu, M.; Aytaç, B.; Avci, A.; Devrim, E.; Ozbek, H.; Oztürk, H.S. Effects of garlic extract consumption on blood lipid and oxidant/antioxidant parameters in humans with high blood cholesterol. J. Nutr. Biochem. 2004, 15, 373–377. [Google Scholar] [CrossRef]
- Lee, D.Y.; Li, H.; Lim, H.J.; Lee, H.J.; Jeon, R.; Ryu, J.H. Anti-inflammatory activity of sulfur-containing compounds from garlic. J. Med. Food 2012, 15, 992–999. [Google Scholar] [CrossRef] [Green Version]
- Percival, S.S. Aged Garlic Extract Modifies Human Immunity. J. Nutr. 2016, 146, 433S–436S. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Lim, W.C.; Lee, S.J.; Lee, S.H.; Lee, J.H.; Cho, H.Y. Antiobesity Effect of Garlic Extract Fermented by Lactobacillus plantarum BL2 in Diet-Induced Obese Mice. J. Med. Food 2016, 19, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.M.; Ban, J.O.; Park, K.R.; Lee, C.K.; Jeong, H.S.; Han, S.B.; Hong, J.T. Potential therapeutic effects of functionally active compounds isolated from garlic. Pharmacol. Ther. 2014, 142, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Neil, A.; Silagy, C. Garlic: Its cardio-protective properties. Curr. Opin. Lipidol. 1994, 5, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Qidwai, W.; Ashfaq, T. Role of garlic usage in cardiovascular disease prevention: An evidence-based approach. Evid. Based Complement. Altern. Med. 2013, 2013, 125649. [Google Scholar] [CrossRef] [PubMed]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef] [Green Version]
- Bradley, J.M.; Organ, C.L.; Lefer, D.J. Garlic-Derived Organic Polysulfides and Myocardial Protection. J. Nutr. 2016, 146, 403S–409S. [Google Scholar] [CrossRef]
- Dhawan, V.; Jain, S. Garlic supplementation prevents oxidative DNA damage in essential hypertension. Mol. Cell. Biochem. 2005, 275, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Borek, C. Garlic reduces dementia and heart-disease risk. J. Nutr. 2006, 136, 810S–812S. [Google Scholar] [CrossRef]
- Gu, C.; Howell, K.; Dunshea, F.R.; Suleria, H.A. Lc-esi-qtof/ms characterisation of phenolic acids and flavonoids in polyphenol-rich fruits and vegetables and their potential antioxidant activities. Antioxidants 2019, 8, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diretto, G.; Rubio-Moraga, A.; Argandoña, J.; Castillo, P.; Gómez-Gómez, L.; Ahrazem, O. Tissue-specific accumulation of sulfur compounds and saponins in different parts of garlic cloves from purple and white ecotypes. Molecules 2017, 22, 1359. [Google Scholar] [CrossRef]
- Wang, Y.; Guan, M.; Zhao, X.; Li, X. Effects of garlic polysaccharide on alcoholic liver fibrosis and intestinal microflora in mice. Pharm. Biol. 2018, 56, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.; Cao, S.Y.; Xu, X.Y.; Gan, R.Y.; Tang, G.Y.; Corke, H.; Mavumengwana, V.; Li, H.B. Bioactive Compounds and Biological Functions of Garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Batiha, G.E.; Beshbishy, A.M.; Wasef, L.G.; Elewa, Y.H.A.; Al-Sagan, A.A.; El-Hack, M.E.A.; Taha, A.E.; Abd-Elhakim, Y.M.; Devkota, H.P. Chemical Constituents and Pharmacological Activities of Garlic (Allium sativum L.): A Review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, H.; Suma, K.; Origuchi, K.; Seki, T.; Ariga, T. Thermostability of allicin determined by chemical and biological assays. Biosci. Biotechnol. Biochem. 2008, 72, 2877–2883. [Google Scholar] [CrossRef]
- Fujisawa, H.; Suma, K.; Origuchi, K.; Kumagai, H.; Seki, T.; Ariga, T. Biological and chemical stability of garlic-derived allicin. J. Agric. Food Chem. 2008, 56, 4229–4235. [Google Scholar] [CrossRef]
- Cairone, F.; Carradori, S.; Locatelli, M.; Casadei, M.A.; Cesa, S. Reflectance colorimetry: A mirror for food quality—A mini review. Eur. Food Res. Technol. 2020, 246, 259–272. [Google Scholar] [CrossRef]
- Simioni, C.; Cani, A.; Martelli, A.M.; Zauli, G.; Alameen, A.A.; Ultimo, S.; Tabellini, G.; McCubrey, J.A.; Capitani, S.; Neri, L.M. The novel dual PI3K/mTOR inhibitor NVP-BGT226 displays cytotoxic activity in both normoxic and hypoxic hepatocarcinoma cells. Oncotarget 2015, 6, 17147–17160. [Google Scholar] [CrossRef]
- Recinella, L.; Chiavaroli, A.; Orlando, G.; Menghini, L.; Ferrante, C.; Di Cesare Mannelli, L.; Ghelardini, C.; Brunetti, L.; Leone, S. Protective Effects Induced by Two Polyphenolic Liquid Complexes from Olive (Olea europaea, mainly Cultivar Coratina) Pressing Juice in Rat Isolated Tissues Challenged with LPS. Molecules 2019, 24, 3002. [Google Scholar] [CrossRef] [Green Version]
- Recinella, L.; Chiavaroli, A.; Orlando, G.; Ferrante, C.; Marconi, G.D.; Gesmundo, I.; Granata, R.; Cai, R.; Sha, W.; Schally, A.V.; et al. Antinflammatory, antioxidant, and behavioral effects induced by administration of growth hormone-releasing hormone analogs in mice. Sci. Rep. 2020, 10, 4850. [Google Scholar] [CrossRef] [Green Version]
- Chiavaroli, A.; Di Simone, S.C.; Sinan, K.I.; Ciferri, M.C.; Flores, G.A.; Zengin, G.; Etienne, O.K.; Ak, G.; Mahomoodally, M.F.; Jugreet, S.; et al. Pharmacological properties and chemical profiles of Passiflora foetida L. Extracts: Novel insights for pharmaceuticals and nutraceuticals. Processes 2020, 8, 1034. [Google Scholar] [CrossRef]
- Leone, S.; Chiavaroli, A.; Shohreh, R.; Ferrante, C.; Ricciuti, A.; Manippa, F.; Recinella, L.; Di Nisio, C.; Orlando, G.; Salvatori, R.; et al. Increased locomotor and thermogenic activity in mice with targeted ablation of the GHRH gene. Growth Horm. IGF Res. 2015, 25, 80–84. [Google Scholar] [CrossRef]
- Leone, S.; Recinella, L.; Chiavaroli, A.; Martinotti, S.; Ferrante, C.; Mollica, A.; Macedonio, G.; Stefanucci, A.; Dvorácskó, S.; Tömböly, C.; et al. Emotional disorders induced by Hemopressin and RVD-hemopressin(α) administration in rats. Pharmacol. Rep. 2017, 69, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Leone, S.; Shohreh, R.; Manippa, F.; Recinella, L.; Ferrante, C.; Orlando, G.; Salvatori, R.; Vacca, M.; Brunetti, L. Behavioural phenotyping of male growth hormone-releasing hormone (GHRH) knockout mice. Growth Horm. IGF Res. 2014, 24, 192–197. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Charan, J.; Kantharia, N.D. How to calculate sample size in animal studies? J. Pharmacol. Pharmacother. 2013, 4, 303–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trio, P.Z.; You, S.; He, X.; He, J.; Sakao, K.; Hou, D.X. Chemopreventive functions and molecular mechanisms of garlic organosulfur compounds. Food Funct. 2014, 5, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Kyung, K.H. Allyl alcohol is the sole antiyeast compound in heated garlic extract. J. Food Sci. 2005, 70, M305–M309. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kim, J.K.; Kim, H.H.; Lee, S.Y.; Park, N.I.; Park, S.U. Carotenoid accumulation and characterization of cDNAs encoding phytoene synthase and phytoene desaturase in garlic (Allium sativum). J. Agric. Food Chem. 2011, 59, 5412–5417. [Google Scholar] [CrossRef]
- Cesa, S.; Casadei, M.A.; Cerreto, F.; Paolicelli, P. Infant milk formulas: Effect of storage conditions on the stability of powdered products towards autoxidation. Foods 2015, 4, 487–500. [Google Scholar] [CrossRef]
- Maccelli, A.; Cesa, S.; Cairone, F.; Secci, D.; Menghini, L.; Chiavarino, B.; Fornarini, S.; Crestoni, M.E.; Locatelli, M. Metabolic profiling of different wild and cultivated Allium species based on high-resolution mass spectrometry, high-performance liquid chromatography-photodiode array detector, and color analysis. J. Mass Spectrom. 2020, 55, e4525. [Google Scholar] [CrossRef]
- Hajji, H.E.; Nkhili, E.; Tomao, V.; Dangles, O. Interactions of quercetin with iron and copper ions: Complexation and autoxidation. Free Radic. Res. 2006, 40, 303–320. [Google Scholar] [CrossRef]
- Kim, J.S.; Kang, O.J.; Gweon, O.C. Comparison of phenolic acids and flavonoids in black garlic at different thermal processing steps. J. Funct. Foods 2013, 5, 80–86. [Google Scholar] [CrossRef]
- Bozin, B.; Mimica-Dukic, N.; Samojlik, I.; Goran, A.; Igic, R. Phenolics as antioxidants in garlic (Allium sativum L., Alliaceae). Food Chem. 2008, 111, 925–929. [Google Scholar] [CrossRef]
- Dziri, S.; Hassen, I.; Fatnassi, S.; Mrabet, Y.; Casabianca, H.; Hanchi, B.; Hosni, K. Phenolic constituents, antioxidant and antimicrobial activities of rosy garlic (Allium roseum var. odoratissimum). J. Funct. Foods 2012, 4, 423–432. [Google Scholar] [CrossRef]
- Bloem, E.; Haneklaus, S.; Schnug, E. Influence of nitrogen and sulfur fertilization on the alliin content of onions and garlic. J. Plant Nutr. 2005, 27, 1827–1839. [Google Scholar] [CrossRef]
- Beato, V.M.; Orgaz, F.; Mansilla, F.; Montaño, A. Changes in phenolic compounds in garlic (Allium sativum L.) owing to the cultivar and location of growth. Plant Foods Hum. Nutr. 2011, 66, 218–223. [Google Scholar] [CrossRef]
- Recinella, L.; Chiavaroli, A.; Ronci, M.; Menghini, L.; Brunetti, L.; Leone, S.; Tirillini, B.; Angelini, P.; Covino, S.; Venanzoni, R.; et al. Multidirectional Pharma-Toxicological Study on Harpagophytum procumbens DC. ex Meisn.: An IBD-Focused Investigation. Antioxidants 2020, 9, 168. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity: Implications for metabolic syndrome, diabetes, hypertension, dyslipidemia, atherosclerosis, and cancer. Obes. Res. Clin. Pract. 2013, 7, e330–e341. [Google Scholar] [CrossRef]
- Ruparelia, N.; Chai, J.T.; Fisher, E.A.; Choudhury, R.P. Inflammatory processes in cardiovascular disease: A route to targeted therapies. Nat. Rev. Cardiol. 2017, 14, 314. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.H.; Efendy, J.L.; Smith, N.J.; Campbell, G.R. Molecular basis by which garlic suppresses atherosclerosis. J. Nutr. 2001, 131, 1006S–1009S. [Google Scholar] [CrossRef]
- El-Sabban, F.; Abouazra, H. Effect of garlic on atherosclerosis and its factors. East. Mediterr. Health J. 2008, 14, 195–205. [Google Scholar] [PubMed]
- Praticò, D.; Lee, V.M.Y.; Trojanoswki, J.Q.; Rokach, J.; FitzGerald, G.A. Increased F2-isoprostanes in Alzheimer’s disease: Evidence for enhanced lipid peroxidation in vivo. FASEB J. 1998, 12, 1777–1783. [Google Scholar] [CrossRef] [PubMed]
- Arreola, R.; Quintero-Fabián, S.; López-Roa, R.I.; Flores-Gutiérrez, E.O.; Reyes-Grajeda, J.P.; Carrera-Quintanar, L.; Ortuño-Sahagún, D. Immunomodulation and anti-inflammatory effects of garlic compounds. J. Immunol. Res. 2015, 2015, 401630. [Google Scholar] [CrossRef]
- Pârvu, M.; Moţ, C.A.; Pârvu, A.E.; Mircea, C.; Stoeber, L.; Roşca-Casian, O.; Ţigu, A.B. Allium sativum Extract Chemical Composition, Antioxidant Activity and Antifungal Effect against Meyerozyma guilliermondii and Rhodotorula mucilaginosa Causing Onychomycosis. Molecules 2019, 24, 3958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-H.; Park, Y.K.; Park, E. Antioxidative and Antigenotoxic Effects of Garlic (Allium sativum L.) Prepared by Different Processing Methods Plant. Foods Hum. Nutr. 2009, 64, 244–249. [Google Scholar] [CrossRef]
- Vlase, L.; Parvu, M.; Parvu, E.; Toiu, A. Chemical constituents of three Allium species from Romania. Molecules 2013, 18, 114–127. [Google Scholar] [CrossRef]
- Shi, G.F.; An, L.J.; Jiang, B.; Guan, S.; Bao, Y.M. Alpinia protocatechuic acid protects against oxidative damage in vitro and reduces oxidative stress in vivo. Neurosci. Lett. 2006, 403, 206–210. [Google Scholar] [CrossRef]
- Masodsai, K.; Lin, Y.Y.; Chaunchaiyakul, R.; Su, C.T.; Lee, S.D.; Yang, A.L. Twelve-Week Protocatechuic Acid Administration Improves Insulin-Induced and Insulin-Like Growth Factor-1-Induced Vasorelaxation and Antioxidant Activities in Aging Spontaneously Hypertensive Rats. Nutrients 2019, 11, 699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perazzoli, M.R.; Perondi, C.K.; Baratto, C.M.; Winter, E.; Creczynski-Pasa, T.B.; Locatelli, C. Gallic Acid and Dodecyl Gallate Prevents Carbon Tetrachloride-Induced Acute and Chronic Hepatotoxicity by Enhancing Hepatic Antioxidant Status and Increasing p53 Expression. Biol. Pharm. Bull. 2017, 40, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsimo, S.J.N.; de Dieu Tamokou, J.; Havyarimana, L.; Csupor, D.; Forgo, P.; Hohmann, J.; Kuiate, J.-R.; Tane, P. Antimicrobial and antioxidant activity of kaempferol rhamnoside derivatives from Bryophyllum pinnatum. BMC Res. Notes 2012, 5, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Ren, H.; Han, J.; Wang, W.; Zheng, Q.; Wang, D. Protective Effects of Kaempferol against Myocardial Ischemia/Reperfusion Injury in Isolated Rat Heart via Antioxidant Activity and Inhibition of Glycogen Synthase Kinase-3. Oxid. Med. Cell. Longev. 2015, 2015, 8. [Google Scholar] [CrossRef] [Green Version]
- Mojiminiyi, F.B.; Dikko, M.; Muhammad, B.Y.; Ojobor, P.D.; Ajagbonna, O.P.; Okolo, R.U.; Igbokwe, U.V.; Mojiminiyi, U.E.; Fagbemi, M.A.; Bello, S.O.; et al. Antihypertensive effect of an aqueous extract of the calyx of Hibiscus sabdariffa. Fitoterapia 2007, 78, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Sarr, M.; Ngom, S.; Kane, M.O.; Wele, A.; Diop, D.; Sarr, B.; Gueye, L.; Andriantsitohaina, R.; Diallo, A.S. In vitro vasorelaxation mechanisms of bioactive compounds extracted from Hibiscus sabdariffa on rat thoracic aorta. Nutr. Metab. 2009, 6, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, S.; Katare, P.B.; Elancheran, R.; Nizami, H.L.; Paramesha, B.; Arava, S.; Sarma, P.P.; Kumar, R.; Mahajan, D.; Kumar, Y.; et al. Musa balbisiana Fruit Rich in Polyphenols Attenuates Isoproterenol-Induced Cardiac Hypertrophy in Rats via Inhibition of Inflammation and Oxidative Stress. Oxid. Med. Cell. Longev. 2020, 2020, 7147498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakkar, S.; Bais, S. A review on protocatechuic acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.L.; Liu, J.X.; Dong, W.; Li, P.; Li, L.; Lin, C.R.; Zheng, Y.Q.; Cong, W.H.; Hou, J.C. Cardioprotective effect of protocatechuic acid on myocardial ischemia/reperfusion injury. J. Pharmacol. Sci. 2014, 125, 176–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safaeian, L.; Emami, R.; Hajhashemi, V.; Haghighatian, Z. Antihypertensive and antioxidant effects of protocatechuic acid in deoxycorticosterone acetate-salt hypertensive rats. Biomed. Pharmacother. 2018, 100, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Jadon, A.; Bhadauria, M.; Shukla, S. Protective effect of Terminalia belerica Roxb and gallic acid against carbon tetrachloride induced damage in albino rats. J. Ethnopharmacol. 2007, 109, 214–218. [Google Scholar] [CrossRef]
- Priscilla, D.H.; Prince, P.S. Cardioprotective effect of gallic acid on cardiac troponin-T, cardiac marker enzymes, lipid peroxidation products and antioxidants in experimentally induced myocardial infarction in Wistar rats. Chem. Biol. Interact. 2009, 179, 118–124. [Google Scholar] [CrossRef]
- Subbaramaiah, K.; Dannenberg, A.J. Cyclooxygenase 2: A molecular target for cancer prevention and treatment. Trends Pharmacol. Sci. 2003, 24, 96–102. [Google Scholar] [CrossRef]
- Lee, J.D.; Kato, K.; Tobias, P.S.; Kirkland, T.N.; Ulevitch, R.J. Transfection of CD14 into 70Z/3 cells dramatically enhances the sensitivity to complexes of lipopolysaccharide (LPS) and LPS binding protein. J. Exp. Med. 1992, 175, 1697–1705. [Google Scholar] [CrossRef] [Green Version]
- Yun, K.J.; Kim, J.Y.; Kim, J.B.; Lee, K.W.; Jeong, S.Y.; Park, H.J.; Jung, H.J.; Cho, Y.W.; Yun, K.; Lee, K.T. Inhibition of LPS-induced NO and PGE2 production by asiatic acid via NF-kappa B inactivation in RAW 264.7 macrophages: Possible involvement of the IKK and MAPK pathways. Int. Immunopharmacol. 2008, 8, 431–441. [Google Scholar] [CrossRef]
- Chenevard, R.; Hurlimann, D.; Bechir, M.; Enseleit, F.; Spieker, L.; Hermann, M.; Riesen, W.; Gay, S.; Gay, R.E.; Neidhart, M.; et al. Selective COX-2 inhibition improves endothelial function in coronary artery disease. Circulation 2003, 107, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Hodge, G.; Hodge, S.; Han, P. Allium sativum (garlic) suppresses leukocyte inflammatory cytokine production in vitro: Potential therapeutic use in the treatment of inflammatory bowel disease. Cytometry 2002, 48, 209–215. [Google Scholar] [CrossRef]
- Makris, A.; Thornton, C.E.; Xu, B.; Hennessy, A. Garlic increases IL-10 and inhibits TNFα and IL-6 production in endotoxin-stimulated human placental explants. Placenta 2005, 26, 828–834. [Google Scholar] [CrossRef]
- Keiss, H.P.; Dirsch, V.M.; Hartung, T.; Haffner, T.; Trueman, L.; Auger, J.; Kahane, R.; Vollmar, A.M. Garlic (Allium sativum L.) modulates cytokine expression in lipopolysaccharide-activated human blood thereby inhibiting NF-kappaB activity. J. Nutr. 2003, 133, 2171–2175. [Google Scholar] [CrossRef]
- Pan, M.H.; Lai, C.S.; Wang, Y.J.; Ho, C.T. Acacetin suppressed LPS-induced up-expression of iNOS and COX-2 in murine macrophages and TPA-induced tumor promotion in mice. Biochem. Pharmacol. 2006, 72, 1293–1303. [Google Scholar] [CrossRef]
- Li, Q.; Verma, I.M. NF-κB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef]
- Park, S.Y.; Seetharaman, R.; Ko, M.J.; Kim, D.Y.; Kim, T.H.; Yoon, M.K.; Kwak, J.H.; Lee, S.J.; Bae, Y.S.; Choi, Y.W. Ethyl linoleate from garlic attenuates lipopolysaccharide-induced pro-inflammatory cytokine production by inducing heme oxygenase-1 in RAW264.7 cells. Int. Immunopharmacol. 2014, 19, 253–261. [Google Scholar] [CrossRef]
- BenSaad, L.A.; Kim, K.H.; Quah, C.C.; Kim, W.R.; Shahimi, M. Anti-inflammatory potential of ellagic acid, gallic acid and punicalagin A & B isolated from Punica granatum. BMC Complement. Altern. Med. 2017, 17, 47. [Google Scholar]
- Adeyanju, A.A.; Asejeje, F.O.; Molehin, O.R.; Owoeye, O.; Olatoye, E.O.; Ekpo, E.N. Protective role of protocatechuic acid in carbon tetrachloride-induced oxidative stress via modulation of proinflammatory cytokines levels in brain and liver of Wistar rats. J. Basic Clin. Physiol. Pharmacol. 2021, accepted. [Google Scholar] [CrossRef]
- Kim, K.J.; Hwang, E.-S.; Kim, M.-J.; Park, J.-H.; Kim, D.-O. Antihypertensive Effects of Polyphenolic Extract from Korean Red Pine (Pinus densiflora Sieb. et Zucc.) Bark in Spontaneously Hypertensive Rats. Antioxidants 2020, 9, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Wang, X.Q.; Wang, H.Z.; Wu, Y.N. High performance liquid chromatographic determination of phenolic acids in fruits and vegetables. Biomed. Environ. Sci. 1993, 6, 389–398. [Google Scholar] [PubMed]
- El-Hussainy, M.; Hussein, A.M.; Abdel-Aziz, A.; El-Mehasseb, I. Effects of aluminum oxide (Al2O3) nanoparticles on ECG, myocardial inflammatory cytokines, redox state, and connexin 43 and lipid profile in rats: Possible cardioprotective effect of gallic acid. Can. J. Physiol. Pharmacol. 2016, 94, 868–878. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef]
- Mena, P.; Del Rio, D. Gold Standards for Realistic (Poly)phenol Research. J. Agric. Food Chem. 2018, 66, 8221–8223. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Gálvez, M.Á.; González-Sarrías, A.; Espín, J.C. In Vitro Research on Dietary Polyphenols and Health: A Call of Caution and a Guide on How to Proceed. J. Agric. Food Chem. 2018, 66, 7857–7858. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | 50 | 70 | 80 | 90 |
| Equilibration time (min) | 20 | |||
| 40 | ||||
| Sampling time (min) | 15 | |||
| 35 | ||||
| Compound | Class | Area% | RI | RIL a |
|---|---|---|---|---|
| Ethanol b | Alcohols | 0.10 | - | - |
| Allyl alcohol b | Alcohols | 3.07 | - | - |
| Trichloromethane b | Other | 0.12 | - | - |
| Hexanal | Other | 0.01 | 809 | 800 |
| Diallyl sulfide | SCC | 0.09 | 859 | 857 |
| Methyl allyl disulfide | SCC | 0.22 | 917 | 919 |
| Dimethyl trisulfide | SCC | 0.04 | 970 | 969 |
| 2-Thiophenecarboxaldehyde | SCC | 0.07 | 1011 | 1005 |
| Diallyl disulfide | SCC | 6.18 | 1081 | 1090 |
| Allyl methyl trisulfide | SCC | 2.08 | 1140 | 1137 |
| 2,3-Dihydro-3,5-dihydroxy-6-methyl-4H-pyran-4-one | Other | 0.78 | 1157 | 1162 |
| 3-Vinyl-1,2-dithiacyclohex-5-ene | SCC | 0.07 | 1190 | 1185 |
| 3-Vinyl-1,2-dithiacyclohex-4-ene | SCC | 0.24 | 1216 | 1214 |
| Diallyl trisulfide (allitridin) | SCC | 22.0 | 1305 | 1300 |
| Propenyl propyl trisulfide | SCC | 0.16 | 1316 | 1314 |
| Eugenol | Alcohols | 0.54 | 1366 | 1357 |
| Methyl 1-propenyl tetrasulfide | SCC | 0.93 | 1369 | 1368 |
| Allyl 1-methylthio propyl disulfide | SCC | 0.85 | 1386 | 1387 |
| Diallyl tetrasulfide | SCC | 6.80 | 1548 | 1539 |
| Tridecyl methyl ketone | Other | 0.15 | 1700 | 1697 |
| 2,4-dimethyl-5,6-dithia-2,7-nonadienal | SCC | 3.10 | 1789 | 1788 |
| Myristic acid | Other | 0.11 | 1795 | 1794 |
| FAE (unidentified) | FAE | 0.40 | 1823 | - |
| Phthalate diisobutyl | Other | 0.18 | 1876 | 1858 |
| Methyl palmitate | FAE | 1.26 | 1926 | 1925 |
| Phthalate dibutyl | FAE | 0.39 | 1969 | 1960 |
| Ethyl palmitoleate | FAE | 0.70 | 1972 | 1975 |
| Ethyl palmitate | FAE | 9.68 | 1995 | 1993 |
| Propyl palmitate | FAE | 0.31 | 2089 | 2077 |
| Methyl linoleate | FAE | 2.62 | 2096 | 2093 |
| Methyl elaidate | FAE | 0.56 | 2101 | 2107 |
| Ethyl linoleate | FAE | 17.8 | 2161 | 2164 |
| Ethyl oleate | FAE | 4.93 | 2165 | 2173 |
| Ethyl stearate | FAE | 0.16 | 2193 | 2198 |
| Methyl-9-12-heptadecadienoate b | FAE | 2.70 | 2242 | - |
| 29 unknown | 10.6 | |||
| Class | Area% | |||
| Alcohol | 3.7 | |||
| SCC | 42.8 | |||
| FAE | 41.5 | |||
| Others | 1.4 | |||
| Hydroalcoholic Extract | ||||
|---|---|---|---|---|
| Compound | Class | Area% | RI | RIL a |
| Diisobutyl phthalate | Other | 0.60 | 1883 | 1858 |
| Methyl palmitate | FAE | 2.07 | 1933 | 1925 |
| Ethyl palmitate | FAE | 2.14 | 2001 | 1993 |
| Methyl linoleate | FAE | 6.87 | 2103 | 2093 |
| FAE (unidentified) | FAE | 2.69 | 2107 | - |
| Ethyl linoleate | FAE | 4.76 | 2167 | 2164 |
| FAE (unidentified) | FAE | 3.63 | 2171 | - |
| Docosane | LA | 5.95 | 2202 | 2200 |
| Tricosane | LA | 11.98 | 2302 | 2300 |
| Tetracosane | LA | 16.48 | 2402 | 2400 |
| Pentacosane | LA | 12.75 | 2502 | 2500 |
| Hexacosane | LA | 6.31 | 2602 | 2600 |
| unknown | 23.76 | |||
| Class | Area% | |||
| FAE | 22.16 | |||
| LA | 53.47 | |||
| L* | a* | b* | C* | h° | ΔC*ab | Δhab | ΔE | |
|---|---|---|---|---|---|---|---|---|
| Powder t° | 90.15 | 0.47 | 16.02 | 16.03 | 88.32 | |||
| Powder t4m | 91.67 | 0.88 | 17.31 | 17.33 | 87.08 | 1.30 Brighter | −0.36 More red | 1.11 |
| Powder t8m | 93.77 | 0.45 | 13.77 | 13.78 | 88.13 | −2.25 More opaque | −0.05 More red | 1.98 |
| GHE | 61.04 | −0.79 | 6.81 | 6.86 | 96.65 | −9.17 More opaque | 1.52 More green | 12.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Recinella, L.; Chiavaroli, A.; Masciulli, F.; Fraschetti, C.; Filippi, A.; Cesa, S.; Cairone, F.; Gorica, E.; De Leo, M.; Braca, A.; et al. Protective Effects Induced by a Hydroalcoholic Allium sativum Extract in Isolated Mouse Heart. Nutrients 2021, 13, 2332. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072332
Recinella L, Chiavaroli A, Masciulli F, Fraschetti C, Filippi A, Cesa S, Cairone F, Gorica E, De Leo M, Braca A, et al. Protective Effects Induced by a Hydroalcoholic Allium sativum Extract in Isolated Mouse Heart. Nutrients. 2021; 13(7):2332. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072332
Chicago/Turabian StyleRecinella, Lucia, Annalisa Chiavaroli, Fabrizio Masciulli, Caterina Fraschetti, Antonello Filippi, Stefania Cesa, Francesco Cairone, Era Gorica, Marinella De Leo, Alessandra Braca, and et al. 2021. "Protective Effects Induced by a Hydroalcoholic Allium sativum Extract in Isolated Mouse Heart" Nutrients 13, no. 7: 2332. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072332