
Effect of Polyphenols Intake on Obesity-Induced Maternal Programming
by
 , , and
, , and
Isabela Monique Fortunato
1 ,
,
Tanila Wood dos Santos
1,
Lucio Fábio Caldas Ferraz
1,
Juliana Carvalho Santos
2,* and
Marcelo Lima Ribeiro
1,* 1
Post Graduate Program in Health Science, Universidade São Francisco (USF), Bragança Paulista 12916-900, SP, Brazil
2
Lymphoma Translational Group, Josep Carreras Leukemia Research Institute (IJC), 08916 Badalona, Spain
*
Authors to whom correspondence should be addressed.
Nutrients 2021, 13(7), 2390; https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072390
Submission received: 25 May 2021
/
Revised: 30 June 2021
/
Accepted: 3 July 2021
/
Published: 13 July 2021
(This article belongs to the Special Issue Plant Polyphenols and Human Health: Novel Findings for Future Therapeutic Developments)
{kind=link}
{kind=link}
Abstract
:Excess caloric intake and body fat accumulation lead to obesity, a complex chronic disease that represents a significant public health problem due to the health-related risk factors. There is growing evidence showing that maternal obesity can program the offspring, which influences neonatal phenotype and predispose offspring to metabolic disorders such as obesity. This increased risk may also be epigenetically transmitted across generations. Thus, there is an imperative need to find effective reprogramming approaches in order to resume normal fetal development. Polyphenols are bioactive compounds found in vegetables and fruits that exert its anti-obesity effect through its powerful anti-oxidant and anti-inflammatory activities. Polyphenol supplementation has been proven to counteract the prejudicial effects of maternal obesity programming on progeny. Indeed, some polyphenols can cross the placenta and protect the fetal predisposition against obesity. The present review summarizes the effects of dietary polyphenols on obesity-induced maternal reprogramming as an offspring anti-obesity approach.
1. Introduction
Excess caloric intake and body fat accumulation lead to obesity. Obesity is considered as one of the main public health problems worldwide, and affects not only industrialized nations, but also developing countries. Epidemiological data presented by the World Health Organization have found that since 1975, obesity has almost tripled and at least 2.8 million people die annually due to complications from obesity or overweight. Despite the current public awareness of the consequences of obesity, its incidence continues to rise, and is distributed in almost all ethnicities and in both sexes, mainly affecting the population aged 25 to 44 years [1].
Obesity has a multifactorial etiology and involves an interaction between genetic and environmental factors [2]. It is characterized as white adipose tissue (WAT) expansion, and in general results from an energy imbalance as a result of an increase in caloric intake coupled with a decrease in daily energy spending [3,4]. The hyperplasia or hypertrophy of adipocytes caused by an accumulation of triacylglycerols leads to an expansion of body fat deposits and an increase in the concentrations of free circulating fatty acids, peptides, inflammatory cytokines, and adipokines, resulting in metabolic disorders such as hepatic steatosis, diabetes, metabolic syndrome, atherosclerosis, and dyslipidemia, conditions that also contribute to the cardiometabolic risk [5,6].
Maternal obesity also has different clinical implications, and is associated with significant health risks to the mother and the newborn. Of note, in animal models, high fat diet (HFD) intake during the pregnancy may predispose offspring to postnatal metabolic inflammatory disorders such as obesity. In this sense, the well-known anti-inflammatory activity of polyphenols could be considered as a reprogramming strategy against maternal obesity-induced adversities. Thus, the present review summarizes the effects of dietary polyphenols on obesity-induced maternal reprogramming as an offspring anti-obesity approach.
2. Obesity and Maternal Programming
It has been described that nutritional, hormonal, and environmental changes during pregnancy and lactation are strongly associated with the appearance of adulthood diseases. The physiological adaptations developed by the organism, as a survival strategy, could potentially become detrimental to the individual’s health from the moment when adverse conditions are restored to normal levels. Although several studies have shown that these disorders can have origins even before birth, the exact mechanisms by which these alterations occur are still poorly understood. This biological phenomenon was initially called “metabolic programming” [7,8,9] and later as “ontogenetic plasticity”, as it is a more probabilistic than a deterministic event [10].
Indeed, the phenomenon of programming or ontogenetic plasticity has been extensively studied for decades by several research groups around the world. Adequate nutrition is known to be essential during crucial periods of development as it can act as an imprinting or priming factor, leading to physiological changes programming future diseases. The association between the observed changes in critical periods and disorders in adulthood gave rise to the “Barker hypothesis”, then the Fetal Origin of Adult Disease, and is now the Developmental Origins of Health and Disease (DOHaD) [11].
Several factors involved in disease-programming have already been described [12,13,14,15]. Among them, we highlight maternal obesity, which has been widely linked to the birth of newborns who are more susceptible to overweight and obesity development [16,17]. Although the prevalence of obesity grows alarmingly and affects all age groups, it should be noted that approximately 39.7% of women between 20 and 39 years of age are obese in the U.S. (NCHS Data Brief No. 360, February 2020). This dataset shows that obesity is a seriously concerning vicious cycle that requires intensive research.
Maternal obesity has been associated with abdominal fat expansion and higher cardiometabolic risk (increased blood pressure, increased serum concentrations of triglycerides, LDL-C and C-Reactive Protein, and low levels of HDL-C) [18]. In animal studies, maternal obesity is correlated with increased adiposity, hyperphagia, increased cholesterol and triglyceride concentrations and increased lipogenesis and a decrease in beta oxidation, leading to the development of hepatic steatosis on offspring [15,19]. Furthermore, it has already been demonstrated that the progeny of HFD-fed rats showed changes such as weight gain, increased adiposity, hyperleptinemia, leptin resistance, and hyperglycemia, even immediately after weaning (21 days) [20].
The deleterious consequence of maternal obesity occurs during specific periods of fetal and offspring development, where epigenetic memory can be switched [21]. Murine models have shown that adipogenesis is active, especially during the last week of gestation, and accelerates in early postnatal life until the puppies are weaned. In humans, adipogenesis and adipose tissue growth occur mainly before birth [9]. In both, the expansion of adipose tissue involves hyperplasia and hypertrophy. In adult mice, the adipocyte reserve remains quite stable, ranging from 10 to 20% of adipocyte renewal per month [22]. The number of fat cells in humans is established in adulthood, with a renewal of 10% per year, regardless of body weight, which means that obese persons renew twice as many fat cells annually. Furthermore, when obese persons lose a substantial amount of weight, they maintain their high number of adipocytes, indicating that the adipose tissue has a numerical adipocyte memory, which is defined at the beginning of development [9]. Thus, it is suggested that multiple epigenetic factors are involved in “memorizing” the number of adipocytes.
3. Adipose Tissue Programming
Human adipose tissue development is an uninterrupted process that starts early during embryogenesis [23]. The transcriptional cascade promoting adipogenesis is very complex and has been well-studied. Mechanically, during the adipose tissue development, the stem cells are sensitive to pro-adipogenic signals (such as metabolites and hormones). These signals stimulate several epigenomic changes in transcriptionally accessible regions of adipogenic genes. The epigenomic profiles established during adipogenesis take place in two steps: (1) differentiation of multipotent mesenchymal stem cells (MSCs) into preadipocytes, and (2) terminal adipocyte differentiation [24].
Stem cells have active transcription of pluripotency genes, and repression of adipogenic genes through the presence of epigenetic marks such as DNA methylation (H3K27me3 and H3K9me3) [9,24]. During the pre-adipogenic process, pro-adipogenic genes are silenced by the presence of bivalent histones H3K27me3 and H3K4me3 [25]. The bivalent state is converted to an active state during the adipocyte terminal differentiation, which is associated with the recruitment of the first wave of transcription factors (such as CCAAT Enhancer Binding Protein (C/EBP)β, glucocorticoid receptor, Sterol Regulatory Element Binding Transcription Factor 1 (SREBP-1C), and Zinc Finger Protein 423 (ZFP423)) [25,26,27,28]. These transcription factors, in turn, stimulate the open state of chromatin at regions containing genes involved in the second wave of transcription factors, among which Peroxisome Proliferator Activated Receptor (PPAR)γ and CEBPα stand out. The second wave of transcription factors leads to the transcription of pro-adipogenic genes (such as Solute Carrier Family 2 Member 4 (SLC2A4), Adiponectin (ADIPOQ), and Leptin (LEP)) [25]. The pregnancy and lactation period involve a strong adipogenesis commitment. The adipose tissue, in turn, originates in both embryonic and postnatal development, especially during early childhood and puberty. In fact, in rodents and in humans, there is a gradual gain in adipocyte number until puberty under physiological conditions [29,30]. The number and size of adipocytes remain stable after puberty [29,31].
Several molecular evidences suggest that transmission of epigenetic marks such as DNA methylation and changes in histone code may be important in the children’s phenotypes. Recent studies suggest that the emergence of chronic diseases such as obesity and diabetes may have an epigenetic inheritance [32,33,34]. Accordingly, it has been shown in rodent models that maternal obesity modulates the expression of pro-adipogenic genes such as C/EBPβ, ZFP423, and PPARγ in the offspring, thus reprogramming adipogenesis. In a maternal obesity-programming model, it was observed that the offspring exhibited an increase in the levels of ZFP423 expression, which was associated with a reduced methylation in its promoter. At weaning, it was observed that the high activity ZFP423 led to faster adipogenesis, increased adipocyte differentiation, and higher adiposity [35,36,37]. In addition, DNA methylation and histone modifications may regulate PPARγ expression in adipose tissue. Thus, maternal obesity inhibits PPARγ2 expression through epigenetic mechanisms in the WAT of the offspring. Although it seems paradoxical, it is believed that the repression of this gene in adipose tissue may be an adaptive mechanism to avoid further accumulation of fat [38,39]. However, the persistent repression of PPARγ2 is associated with an increase in the expression of genes involved with fat accumulation (TNF receptor superfamily member 6 (Fas), diacylglycerol O-acyltransferase 2 (Dgat2), and lipoprotein lipase (Lpl)), suggesting that, at least in part, such regulation should occur by additional signaling pathways independent of PPARγ [15]. In summary, maternal obesity may be related to important epigenetic modifications that explain the emergence of obesity and other morbidities presented by the offspring.
4. Polyphenol Effect on Obesity
Beneficial effects of some natural diet components are attributed to their anti-inflammatory and anti-oxidant properties, and have been commonly used to treat and/or prevent diseases. In this sense, several studies highlight a broad range of beneficial effects to polyphenols [40,41].
Polyphenols are bioactive compounds abundantly found in vegetables and fruits involved in plant defense against oxidative stress and ultraviolet radiation, or attracting pollinators [42]. Since chronic inflammation is known to be a leading cause of different disorders such as obesity, diabetes type 2, cancer, arthritis, neurodegenerative, and cardiovascular diseases, many studies have studied the beneficial role of polyphenols in reducing inflammation to treat chronic disorders [43].
These natural compounds contribute to the regulation of inflammatory signaling through the modulation of several pro-inflammatory genes such as cytokines, lipoxygenases, nitric oxide synthases cyclooxygenase, and through their anti-oxidant activity, which support progress toward decreased metabolic disorders [44,45].
Polyphenols belong to a broad group of chemical metabolites, and can be categorized into flavonoids, allied phenolic, and polyphenolic compounds [46]. Some polyphenols such as anthocyanins are absorbed through the gut barrier, while the unabsorbed polyphenols must be enzymatically hydrolyzed to be uptaken by epithelial cells [47]. The anti-obesity effect ascribed to polyphenols are achieved by their ability to interact, directly or indirectly, with adipose tissues and activate 5′ adenosine monophosphate-activated protein kinase (AMPK), which results in the reduction of cholesterol, fatty acid synthesis, and triglyceride formation. Moreover, polyphenols can repress genes that regulate adipocyte differentiation and triglyceride accumulation [48].
Epigallocatechin gallate (EGCG) is the most active flavonoid compound present in green tea. Strong emerging evidence has shown the anti-obesity potential of EGCG. For instance, in vitro studies have demonstrated that EGCG inhibits preadipocyte differentiation, decreases adipocyte proliferation, suppresses lipogenesis, induces adipocyte apoptosis, and promotes lipolysis and fatty acid β-oxidation [49,50,51]. Additionally, EGCG decreased obesity and epididymal WAT weight in mice partially via activating the AMPK pathway [52]. Adipocyte differentiation may be suppressed by EGCG through the inhibition of the PI3K/AKT and MEK/ERK pathways, which may lead to downregulation of PPARγ and C/EBPα, the main adipogenesis regulators [48].
Quercetin is one of the primary flavonoid compounds and widely exists in vegetables, tea, and fruits. It has been reported that quercetin suppresses lipid accumulation, body weight, and insulin resistance in mice [53]. In HFD-induced rodents, treatment with quercetin attenuated both obesity and insulin resistance, inhibited hepatic lipid accumulation by inducing the expression of beta-oxidation related genes and decreasing inflammation [54,55]. In human adipocyte models, quercetin significantly downregulated adipokines (i.e., Angiopoietin Like 4 (ANGPTL4) and Serpin Family E Member 1(SERPINE1, previously known as PAI-1)) and glycolysis-associated enzymes (i.e., Enolase 2, gamma neuronal (ENO2) and 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4 (PFKFB4)), which are closely related to obesity [56]. Mechanistically, in vitro and in vivo analysis has shown that quercetin is able to revert unfavorable epigenomic profiles associated with adipogenesis through the induction of chromatin remodeling and histone modifications, which lead to a decrease in C/EBPα and PPARγ gene expression [55].
In a HFD-fed rat model, kaempferol suppressed the visceral fat accumulation and improved hyperlipidemia through the downregulation of SREBPs and upregulation of hepatic PPARα expression [57]. In obese HFD-induced mice, kaempferol protected against obesity and ameliorated hyperlipidemia partly through maintaining microbial diversity and modulating microbial communities as well as downregulating PPARγ and SREBP-1C [58,59]. In another study, kaempferol modulated 3T3-L1 adipocyte differentiation through regulation of C/EBPα and PPARα [60,61].
Curcumin, the primary natural polyphenolic compound of the spice turmeric, also has anti-insulin sensitivity and anti-obesity activity. It has been shown that curcumin can suppress weight gain, improve insulin sensitivity, and prevent the development of diabetes in rodents and in prediabetic subjects [62]. Curcumin promoted beige adipogenesis and induced pre-adipocyte apoptosis in white adipocytes possibly mediated by AMPK, PPARγ, and C/EBPα [63,64]. In pro-adipogenic conditions of rat MSCs, curcumin inhibited the expression of PPARγ and C/EBPα, which in turn suppressed adipocyte differentiation [65].
Resveratrol, one of the most studied bioactive polyphenolic compounds found in grapes, red wine, and some berries, also modulates several events involved in the obesity process. Due its powerful antioxidant and anti-inflammatory activity, resveratrol has been used as dietary supplements, and as a functional food ingredient to achieve different health benefits. Several in vitro studies have demonstrated that resveratrol has an anti-obesity effect by negatively regulating white adipogenesis via inhibiting PPARγ and C/EBPα [66], activating Sirtuin 1 (SIRT1) and (PPARγ Coactivator 1) PGC1α [67], attenuating white adipogenesis and lipid accumulation. Furthermore, their anti-obesity effect is carried out by inhibiting the transcriptional activities of PPARγ, suppressing adipocyte differentiation, proliferation, and lipogenesis, and promoting adipocyte apoptosis, lipolysis, and fatty acid oxidation [68,69,70].
Several studies have shown that chlorogenic acid (CGA), the major polyphenol present in the coffee, has beneficial effects on obesity. It has been demonstrated, in an HFD-induced mice obese model, that CGA supplementation significantly decreased body weight, visceral fat mass, and leptin and insulin plasma levels [71,72]. Mechanistically, the CGA beneficial anti-obesity effects might be attributed to its ability to decrease C/EBP, PPARγ, and SREBP expression [72]. Furthermore, an interesting study conducted in mice that become identifiably obese due to homozygous diabetes spontaneous mutation (Leprdb) showed that CGA treatment inhibited G6Pase expression, improved lipid metabolism, insulin sensitivity, and glucose tolerance via AMPK activation [73].
Increasing clinical evidence has also shown that coffee exerts anti-obesity effects in humans [74]. Nordestgaard et al. [75] conducted a Mendelian randomization study including 93,179 subjects, and showed that the consumption of up to four cups of coffee per day was associated with a lower risk of obesity than non-coffee drinkers. Similarly, Koyama et al. [76] reported that the daily coffee intake was associated with lower levels of visceral obesity and metabolic syndrome in a population of 3539 Japanese. Another clinical study indicated that 30 obese women who consumed 180 mg of CGA for eight weeks presented significant reductions in body weight, BMI, and fat mass indices as well as lower serum LDL, TC, leptin, and plasma free FA levels when compared with 34 obese women from the placebo group [77].
Salvianolic acid A (SA) is another natural polyphenolic compound found in Radix Salvia miltiorrhiza, with well reported anti-oxidant and anti-inflammatory properties [78]. It has been shown that SA can also reverse the HFD-induced obesity [79]. Indeed, a study conducted with HFD-induced obese mice showed that SA supplementation exerts its anti-obesity effects by activation of WAT browning via the AMPK-SIRT1 pathway [80]. Moreover, SA has a cardioprotective role against lipotoxicity by suppressing the expression of TLR4 target genes, both in vitro and in vivo [81].
Ferulic acid (FA), a polyphenol abundantly found in whole grains, also has the ability to activate AMPK signaling and exerts its anti HFD-induced obesity effects by inhibiting oxidative stress, inflammation, and circulating LDL levels along with increasing adiponectin expression and circulating HDL levels [82]. Additionally, using a 3T3-L1 pre-adipocytes in vitro model, Kuppusamy et al. [83] demonstrated that FA may prevent obesity through downregulation of key transcriptional factors PPARγ and C/EBPα and consequently suppressing adipocyte differentiation and lipid accumulation.
Recent data have shown that TOTUM-63 (T63), a novel plant-based polyphenol extracted from olive leaves, bilberry, artichoke, chrysanthellum, and black pepper, may also have anti-obesity properties. Indeed, a translational study indicated that T63 can reduce body weight, control glucose homeostasis, and may protect against type 2 diabetes [84]. Accordantly, recent studies based on obese rodent models have shown that T63 supplementation improves glycemic levels by inducing skeletal muscle oxidative capacity [85], decreases inflammation, and improves insulin sensitivity [86], suggesting that T63 might be a promising novel nutritional supplement against obesity. Figure 1 summarizes the molecular mechanisms underlying the anti-obesity effects of the main polyphenols.
5. Beneficial Impact of Polyphenols on Obesity-Induced Maternal Programming
It has been shown that maternal obesity functionally modulates adipogenesis from fetal development [87] to adulthood [88], presenting larger adipocytes and WAT mass accumulation [89]. Moreover, maternal obesity leads to increased cytokine production and placental-mediated inflammation, which could affect fetal development and may predispose progeny to subsequent obesity [90,91]. Since maternal obesity-induced metabolic programming has a profound impact on offspring, there is a compelling need to find effective reprogramming approaches in order to resume normal development. In this sense, supplementing with natural compounds such as polyphenols could be helpful in the reprogramming of maternal adversities associated with obesity.
A study conducted by Kataoka, Norikura, and Sato (2018) [92] showed that the intake of EGCG–rich green tea extract during lactation of Wistar rats had a protective effect on the kidney of HFD-fed adult offspring through the suppression of epigenetic modulators such as DNA methyltransferase 1 (DNMT1), ubiquitin like with PHD and ring finger domains 1 (UHRF1), and euchromatic histone lysine methyltransferase 2 (EHMT2), highlighting that the offspring phenotype can be programmed by maternal polyphenol intake. In addition, EGCG treatment of rat embryos blocks Forkhead Box O3 (FOXO3A) activation and reverses AKT inhibition, preventing hyperglycemia-induced embryopathy [93]. Similarly, it has been shown that an isoflavone-rich soy-based diet during rat pregnancy may afford cardiovascular protection in the offspring through the stimulation of antioxidant and redox-sensitive gene expression [94].
Since efficiency and safety of EGCG supplementation has rarely been evaluated during pregnancy in humans, a well-conducted randomized, placebo-controlled, double-blind clinical study evaluated the effects of EGCG in gestational diabetes mellitus-affected women. In general, the authors described that daily administration of 500 mg of EGCG, starting from the beginning of their third trimester, was well tolerated and improved both circulating glucose and insulin response. Additionally, EGCG also ameliorates some neonatal complications such as low birth weight and hypoglycemia [95].
Interestingly, Tain Y et al. [96] highlighted the beneficial effects of resveratrol supplementation as an important reprogramming approach against the metabolic syndrome-related disorders. The long-term effects of both maternal and postnatal HFD intake lead to metabolic disruption characterized by body weight gain, high levels of serum HDL, ALT, triglycerides, leptin, cholesterol, and angiotensin I and II, which could be ameliorated by resveratrol therapy [97]. It has been suggested that resveratrol could cross the placenta and exert its anti-inflammatory [98] and anti-oxidant function [99]. Animal studies have shown the fat browning activator action of resveratrol involving the secretion of several myokines and adipokines and suggest that maternal resveratrol supplementation may have a protective role against HFD-induced obesity [100,101]. Similarly, Zou et al. found that the resveratrol supplementation during pregnancy led to a thermogenic program in BAT and WAT, and stimulated beige adipocyte development of WAT in male progeny [102]. Hsu et al. [103] assessed the effect of maternal resveratrol intake of obese rats and found that resveratrol treatment restored adiponectin, AKT phosphorylation, and brain-derived neurotrophic factor (BDNF) in male fetal brain, increased blood pressure, and reduced increased body weight and peripheral insulin resistance in adult male offspring, further demonstrating that intervention with this polyphenol may protect offspring against HFD-induced obesity. Another study using the HFD-obese rat model [104] highlighted that maternal resveratrol supplementation had a reprogramming role for progeny through lipid metabolic modulation. The authors showed the anti-obesity effect of resveratrol treatment through the suppression of lipogenesis and SIRT1 protein expression, attenuation of leptin resistance, and induction of lipolysis for offspring. Additionally, it has been shown that resveratrol supplementation prevents maternal glucose intolerance and lower blood glucose levels by insulin secretion stimulation [105]. Figure 2 summarizes obesity-induced maternal programming and how polyphenol intake could reprogram the epigenetic memory of adult offspring.
Although there is growing evidence suggesting the therapeutic potential of resveratrol in obesity-induced maternal reprogramming, the safety and long-term risk of in utero exposure in humans are unexplored. Thus, the effects of resveratrol supplementation was evaluated in a Western-style diet-fed pregnant nonhuman primates [106]. In this interesting study, the authors showed that resveratrol supplementation in pregnancy has beneficial effects such as a reduction in maternal weight, an improvement on insulin secretion, restore uteroplacental blood flow, decreased placental inflammation, and an improvement in lipid deposition in the fetal liver. However, an increase in fetal pancreas mass was observed. Altogether, these data highlight that the use of resveratrol should be carefully evaluated in pregnant women.
6. Conclusions
The current evidence highlights the relevance of maternal obesity for the metabolic health of the progeny. This review shows that there is an increasing amount of experimental data pointing to the potential effects of polyphenols as a strategy to counteract the deleterious effects induced by maternal obesity. In general, the data reviewed here demonstrated that supplementing pregnant and lactating obese animals with polyphenols including resveratrol, genistein, EGCG, and anthocyanins led to metabolic health reprogramming that ultimately decreased adiposity in the offspring. Whether these observations translate to the human condition remains to be determined. Further examination of obesity-induced maternal programming, especially in humans, is urgently needed and may help to develop polyphenol-based strategies to decrease the propagation of obesity across generations.
Author Contributions
All authors made a substantial contribution to all aspects of the preparation of this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
MLR acknowledges support from the Brazilian National Council for Scientific and Technological Development (CNPq, 305402/2019-6). I.M.F. was supported by a scholarship from the Coordination for the Improvement of Higher Education Personnel (CAPES, Ministry of Education of Brazil). J.C.S. holds a Sara Borrell research contract from Instituto de Salud Carlos III (CD19/00228).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bentham, J.; Di Cesare, M.; Bilano, V.; Bixby, H.; Zhou, B.; Stevens, G.A.; Riley, L.M.; Taddei, C.; Hajifathalian, K.; Lu, Y.; et al. Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: A pooled analysis of 2416 population-based measurement studies in 128.9 million children, adolescents, and adults. Lancet 2017, 390, 2627–2642. [Google Scholar]
- Hruby, A.; Hu, F.B. The Epidemiology of Obesity: A Big Picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Finer, N. Medical consequences of obesity. Medicine 2015, 43, 88–93. [Google Scholar] [CrossRef]
- De Ferranti, S.; Mozaffarian, D. The perfect storm: Obesity, adipocyte dysfunction, and metabolic consequences. Clin. Chem. 2008, 54, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neeland, I.J.; Ayers, C.R.; Rohatgi, A.K.; Turer, A.T.; Berry, J.D.; Das, S.R.; Vega, G.L.; Khera, A.; McGuire, D.K.; Grundy, S.M.; et al. Associations of visceral and abdominal subcutaneous adipose tissue with markers of cardiac and metabolic risk in obese adults. Obesity 2013, 21, E439–E447. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.J.P. The Developmental Origins of Adult Disease. J. Am. Coll. Nutr. 2004, 23, 588S–595S. [Google Scholar] [CrossRef]
- De Moura, E.G.; Passos, M.C.F. Neonatal programming of body weight regulation and energetic metabolism. Biosci. Rep. 2005, 25, 251–264. [Google Scholar] [CrossRef]
- Lecoutre, S.; Petrus, P.; Rydén, M.; Breton, C. Transgenerational Epigenetic Mechanisms in Adipose Tissue Development. Trends Endocrinol. Metab. 2018, 29, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Hanson, M.A. Developmental plasticity and human disease: Research directions. J. Intern. Med. 2007, 261, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.P.; Hales, C.N.; Fall, C.H.D.; Osmond, C.; Phipps, K.; Clark, P.M.S. Type 2 (non-insulin-dependent) diabetes mellitus, hypertension and hyperlipidaemia (syndrome X): Relation to reduced fetal growth. Diabetologia 1993, 36, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Lima, N.; Gaspar De Moura, E.; Cottini Fonseca Passos, M.; Firmino Nogueira Neto, J.; Reis, A.M.; De Oliveira, E.; Lisboa, P.C. Early weaning causes undernutrition for a short period and programmes some metabolic syndrome components and leptin resistance in adult rat offspring. Br. J. Nutr. 2011, 105, 1405–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, K.; Desai, N.; Gupta, M.; Xue, X.; Chatterjee, P.K.; Rochelson, B.; Metz, C.N. The effects of prenatal metformin on obesogenic diet-induced alterations in maternal and fetal fatty acid metabolism. Nutr. Metab. 2016, 13, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veiga-Lopez, A.; Moeller, J.; Sreedharan, R.; Singer, K.; Lumeng, C.; Ye, W.; Pease, A.; Padmanabhan, V. Developmental programming: Interaction between prenatal bpa exposure and postnatal adiposity on metabolic variables in female sheep. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E238–E247. [Google Scholar] [CrossRef] [Green Version]
- Lecoutre, S.; Pourpe, C.; Butruille, L.; Marousez, L.; Laborie, C.; Guinez, C.; Lesage, J.; Vieau, D.; Eeckhoute, J.; Gabory, A.; et al. Reduced PPARγ2 expression in adipose tissue of male rat offspring from obese dams is associated with epigenetic modifications. FASEB J. 2018, 32, 2768–3778. [Google Scholar] [CrossRef] [Green Version]
- Catalano, P.M. Obesity and pregnancy—The propagation of a viscous cycle? J. Clin. Endocrinol. Metab. 2003, 88, 3505–3506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Mendez, E.; Quintero-Fabian, S.; Fernandez-Mejia, C.; Lazo-De-La-Vega-Monroy, M.L. Early-life programming of adipose tissue. Nutr. Res. Rev. 2020, 33, 244–259. [Google Scholar] [CrossRef]
- Tan, H.C.; Roberts, J.; Catov, J.; Krishnamurthy, R.; Shypailo, R.; Bacha, F. Mother’s pre-pregnancy BMI is an important determinant of adverse cardiometabolic risk in childhood. Pediatr. Diabetes 2015, 16, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Ornellas, F.; Souza-Mello, V.; Mandarim-de-Lacerda, C.A.; Aguila, M.B. Programming of obesity and comorbidities in the progeny: Lessons from a model of diet-induced obese parents. PLoS ONE 2015, 10, e0124737. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.G.; Fernandes, T.P.; Rocha, C.P.D.; Calviño, C.; Pazos-Moura, C.C.; Lisboa, P.C.; Moura, E.G.; Trevenzoli, I.H. Maternal high-fat diet induces obesity and adrenal and thyroid dysfunction in male rat offspring at weaning. J. Physiol. 2012, 590, 5503–5518. [Google Scholar] [CrossRef]
- Desai, M.; Jellyman, J.K.; Han, G.; Beall, M.; Lane, R.H.; Ross, M.G. Maternal obesity and high-fat diet program offspring metabolic syndrome. Am. J. Obstet. Gynecol. 2014, 211, 237.e1–237.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birsoy, K.; Berry, R.; Wang, T.; Ceyhan, O.; Tavazoie, S.; Friedman, J.M.; Rodeheffer, M.S. Analysis of gene networks in white adipose tissue development reveals a role for ETS2 in adipogenesis. Development 2011, 138, 4709–4719. [Google Scholar] [CrossRef] [Green Version]
- Poissonnet, C.M.; Burdi, A.R.; Garn, S.M. The chronology of adipose tissue appearance and distribution in the human fetus. Early Hum. Dev. 1984, 10, 1–11. [Google Scholar] [CrossRef]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From stem cell to adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef] [Green Version]
- Steger, D.J.; Grant, G.R.; Schupp, M.; Tomaru, T.; Lefterova, M.I.; Schug, J.; Manduchi, E.; Stoeckert, C.J.; Lazar, M.A. Propagation of adipogenic signals through an epigenomic transition state. Genes Dev. 2010, 24, 1035–1044. [Google Scholar] [CrossRef] [Green Version]
- Lefterova, M.I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARγ and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.K.; Arany, Z.; Seale, P.; Mepani, R.J.; Ye, L.; Conroe, H.M.; Roby, Y.A.; Kulaga, H.; Reed, R.R.; Spiegelman, B.M. Transcriptional control of preadipocyte determination by Zfp. Nature 2010, 7188, 619–623. [Google Scholar] [CrossRef] [Green Version]
- Musri, M.M.; Párrizas, M. Epigenetic regulation of adipogenesis. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 342–349. [Google Scholar] [CrossRef]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Näslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Holtrup, B.; Church, C.D.; Berry, R.; Colman, L.; Jeffery, E.; Bober, J.; Rodeheffer, M.S. Puberty is an important developmental period for the establishment of adipose tissue mass and metabolic homeostasis. Adipocyte 2017, 6, 224–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, D.C.; Jiang, Y.; Graff, J.M. Emerging Roles of Adipose Progenitor Cells in Tissue Development, Homeostasis, Expansion and Thermogenesis. Trends Endocrinol. Metab. 2016, 27, 574–585. [Google Scholar] [CrossRef]
- Radford, E.J.; Ito, M.; Shi, H.; Corish, J.A.; Yamazawa, K.; Isganaitis, E.; Seisenberger, S.; Hore, T.A.; Reik, W.; Erkek, S.; et al. In utero undernourishment perturbs the adult sperm methylome and intergenerational metabolism. Science 2014, 345, 1255903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Yang, C.R.; Wei, Y.P.; Zhao, Z.A.; Hou, Y.; Schatten, H.; Sun, Q.Y. Paternally induced transgenerational inheritance of susceptibility to diabetes in mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 1873–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Kang, S. Functional implications of DNA methylation in adipose biology. Diabetes 2019, 68, 871–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borengasser, S.J.; Zhong, Y.; Kang, P.; Lindsey, F.; Ronis, M.J.J.; Badger, T.M.; Gomez-Acevedo, H.; Shankar, K. Maternal obesity enhances white adipose tissue differentiation and alters genome-scale DNA methylation in male rat offspring. Endocrinology 2013, 154, 4113–4125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.Y.; Liang, J.F.; Rogers, C.J.; Zhao, J.X.; Zhu, M.J.; Du, M. Maternal obesity induces epigenetic modifications to facilitate Zfp423 expression and enhance adipogenic differentiation in fetal mice. Diabetes 2013, 62, 3727–3735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Yang, Q.; Fu, X.; Rogers, C.J.; Wang, B.; Pan, H.; Zhu, M.J.; Nathanielsz, P.W.; Du, M. Maternal obesity epigenetically alters visceral fat progenitor cell properties in male offspring mice. J. Physiol. 2016, 594, 4453–4466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiki, K.; Kano, F.; Shiota, K.; Murata, M. Expression of the peroxisome proliferator activated receptor γ gene is repressed by DNA methylation in visceral adipose tissue of mouse models of diabetes. BMC Biol. 2009, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Zwamborn, R.A.J.; Slieker, R.C.; Mulder, P.C.A.; Zoetemelk, I.; Verschuren, L.; Suchiman, H.E.D.; Toet, K.H.; Droog, S.; Slagboom, P.E.; Kooistra, T.; et al. Prolonged high-fat diet induces gradual and fat depot-specific DNA methylation changes in adult mice. Sci. Rep. 2017, 7, 43261. [Google Scholar] [CrossRef] [Green Version]
- Eberhardt, M.V.; Lee, C.Y.; Liu, R.H. Antioxidant activity of fresh apples. Nature 2000, 405, 903–904. [Google Scholar] [CrossRef]
- Andriantsitohaina, R.; Auger, C.; Chataigneau, T.; Étienne-Selloum, N.; Li, H.; Martínez, M.C.; Schini-Kerth, V.B.; Laher, I. Molecular mechanisms of the cardiovascular protective effects of polyphenols. Br. J. Nutr. 2012, 108, 1532–1549. [Google Scholar] [CrossRef] [Green Version]
- Vuolo, M.M.; Lima, V.S.; Maróstica Junior, M.R. Phenolic Compounds: Structure, Classification, and Antioxidant Power. In Bioactive Compounds: Health Benefits and Potential Applications; Woodhead Publishing: Sawston, UK, 2018; ISBN 9780128147757. [Google Scholar]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, C.; Varì, R.; Scazzocchio, B.; Di Benedetto, R.; Filesi, C.; Masella, R. Polyphenols, intracellular signalling and inflammation. Ann. Ist. Super. Sanita 2007, 43, 394. [Google Scholar]
- Kalupahana, N.S.; Moustaid-Moussa, N. The adipose tissue renin-angiotensin system and metabolic disorders: A review of molecular mechanisms. Crit. Rev. Biochem. Mol. Biol. 2012, 47, 379–390. [Google Scholar] [CrossRef]
- Dragan, S.; Andrica, F.; Serban, M.-C.; Timar, R. Polyphenols-Rich Natural Products for Treatment of Diabetes. Curr. Med. Chem. 2014, 22, 14–22. [Google Scholar] [CrossRef]
- Mosele, J.I.; Macià, A.; Romero, M.P.; Motilva, M.J.; Rubió, L. Application of in vitro gastrointestinal digestion and colonic fermentation models to pomegranate products (juice, pulp and peel extract) to study the stability and catabolism of phenolic compounds. J. Funct. Foods 2015, 14, 529–540. [Google Scholar] [CrossRef]
- Meydani, M.; Hasan, S.T. Dietary polyphenols and obesity. Nutrients 2010, 2, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.Y.; Wei, L.; Castro-Muñozledo, F.; Koo, W.L. (−)-Epigallocatechin-3-gallate blocks 3T3-L1 adipose conversion by inhibition of cell proliferation and suppression of adipose phenotype expression. Life Sci. 2011, 89, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.T.; Park, I.J.; Shin, J.I.; Lee, Y.K.; Lee, S.K.; Baik, H.W.; Ha, J.; Park, O.J. Genistein, EGCG, and capsaicin inhibit adipocyte differentiation process via activating AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2005, 338, 694–699. [Google Scholar] [CrossRef]
- Moon, H.S.; Chung, C.S.; Lee, H.G.; Kim, T.G.; Choi, Y.J.; Cho, C.S. Inhibitory effect of (−)-epigallocatechin-3-gallate on lipid accumulation of 3T3-L1 cells. Obesity 2007, 15, 2571–2582. [Google Scholar] [CrossRef]
- Li, F.; Gao, C.; Yan, P.; Zhang, M.; Wang, Y.; Hu, Y.; Wu, X.; Wang, X.; Sheng, J. EGCG reduces obesity and white adipose tissue gain partly through AMPK activation in mice. Front. Pharmacol. 2018, 9, 1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.J.; Lee, Y.J.; Hwang, J.H.; Kim, K.J.; Lee, B.Y. The inhibitory effects of quercetin on obesity and obesity-induced inflammation by regulation of MAPK signaling. J. Nutr. Biochem. 2015, 26, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Forney, L.A.; Lenard, N.R.; Stewart, L.K.; Henagan, T.M. Dietary quercetin attenuates adipose tissue expansion and inflammation and alters adipocyte morphology in a tissue-specific manner. Int. J. Mol. Sci. 2018, 19, 895. [Google Scholar] [CrossRef] [Green Version]
- Nettore, I.C.; Rocca, C.; Mancino, G.; Albano, L.; Amelio, D.; Grande, F.; Puoci, F.; Pasqua, T.; Desiderio, S.; Mazza, R.; et al. Quercetin and its derivative Q2 modulate chromatin dynamics in adipogenesis and Q2 prevents obesity and metabolic disorders in rats. J. Nutr. Biochem. 2019, 69, 151–162. [Google Scholar] [CrossRef]
- Leiherer, A.; Stoemmer, K.; Muendlein, A.; Saely, C.H.; Kinz, E.; Brandtner, E.M.; Fraunberger, P.; Drexel, H. Quercetin impacts expression of metabolism-and obesity-associated genes in SGBS adipocytes. Nutrients 2016, 8, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.J.; Tzeng, T.F.; Liou, S.S.; Chang, Y.S.; Liu, I.M. Kaempferol regulates the lipid-profile in high-fat diet-fed rats through an increase in hepatic PPAR levels. Planta Med. 2011, 77, 1876–1882. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wu, Q.; Zhao, T. Preventive Effects of Kaempferol on High-Fat Diet-Induced Obesity Complications in C57BL/6 Mice. Biomed. Res. Int. 2020, 2020, 4532482. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Zhang, L.; Igarashi, K.; Yu, C. The anti-obesity and anti-diabetic effects of kaempferol glycosides from unripe soybean leaves in high-fat-diet mice. Food Funct. 2015, 6, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Kwon, M.; Choi, J.S.; Jeong, H.O.; Chung, H.Y.; Kim, H.R. Kaempferol Isolated from Nelumbo nucifera Inhibits Lipid Accumulation and Increases Fatty Acid Oxidation Signaling in Adipocytes. J. Med. Food 2015, 18, 1363–1370. [Google Scholar] [CrossRef] [PubMed]
- Torres-Villarreal, D.; Camacho, A.; Castro, H.; Ortiz-Lopez, R.; de la Garza, A.L. Anti-obesity effects of kaempferol by inhibiting adipogenesis and increasing lipolysis in 3T3-L1 cells. J. Physiol. Biochem. 2019, 75, 83–88. [Google Scholar] [CrossRef]
- Jin, T.; Song, Z.; Weng, J.; Fantus, I.G. Curcumin and other dietary polyphenols: Potential mechanisms of metabolic actions and therapy for diabetes and obesity. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E201–E205. [Google Scholar] [CrossRef] [PubMed]
- Lone, J.; Choi, J.H.; Kim, S.W.; Yun, J.W. Curcumin induces brown fat-like phenotype in 3T3-L1 and primary white adipocytes. J. Nutr. Biochem. 2016, 27, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.Y.; Chen, C.W.; Chen, L.K.; Chou, H.Y.; Chang, C.L.; Juan, C.C. Curcumin attenuates adipogenesis by inducing preadipocyte apoptosis and inhibiting adipocyte differentiation. Nutrients 2019, 11, 2307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Q.; Cai, Y.; Huang, C.; Shi, Q.; Yang, H. Curcumin increases rat mesenchymal stem cell osteoblast differentiation but inhibits adipocyte differentiation. Pharmacogn. Mag. 2012, 8, 202–208. [Google Scholar]
- Aguirre, L.; Fernández-Quintela, A.; Arias, N.; Portillo, M.P. Resveratrol: Anti-obesity mechanisms of action. Molecules 2014, 19, 18632–18655. [Google Scholar] [CrossRef] [Green Version]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol Improves Mitochondrial Function and Protects against Metabolic Disease by Activating SIRT1 and PGC-1alpha. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Chen, S.; Li, Z.; Li, W.; Shan, Z.; Zhu, W. Resveratrol inhibits cell differentiation in 3T3-L1 adipocytes via activation of AMPK. Can. J. Physiol. Pharmacol. 2011, 89, 793–799. [Google Scholar] [PubMed]
- Chen, S.; Xiao, X.; Feng, X.; Li, W.; Zhou, N.; Zheng, L.; Sun, Y.; Zhang, Z.; Zhu, W. Resveratrol induces Sirt1-dependent apoptosis in 3T3-L1 preadipocytes by activating AMPK and suppressing AKT activity and survivin expression. J. Nutr. Biochem. 2012, 23, 1100–1112. [Google Scholar] [CrossRef]
- Lasa, A.; Schweiger, M.; Kotzbeck, P.; Churruca, I.; Simón, E.; Zechner, R.; del Puy Portillo, M. Resveratrol regulates lipolysis via adipose triglyceride lipase. J. Nutr. Biochem. 2012, 23, 379–384. [Google Scholar] [CrossRef]
- Cho, A.S.; Jeon, S.M.; Kim, M.J.; Yeo, J.; Seo, K., II; Choi, M.S.; Lee, M.K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef]
- Wang, Z.; Lam, K.L.; Hu, J.; Ge, S.; Zhou, A.; Zheng, B.; Zeng, S.; Lin, S. Chlorogenic acid alleviates obesity and modulates gut microbiota in high-fat-fed mice. Food Sci. Nutr. 2019, 7, 579–588. [Google Scholar] [CrossRef]
- Ong, K.W.; Hsu, A.; Tan, B.K.H. Anti-diabetic and anti-lipidemic effects of chlorogenic acid are mediated by ampk activation. Biochem. Pharmacol. 2013, 85, 1341–1351. [Google Scholar] [CrossRef]
- Santos, R.M.M.; Lima, D.R.A. Coffee consumption, obesity and type 2 diabetes: A mini-review. Eur. J. Nutr. 2016, 55, 1345–1358. [Google Scholar] [CrossRef]
- Nordestgaard, A.T.; Thomsen, M.; Nordestgaard, B.G. Coffee intake and risk of obesity, metabolic syndrome and type 2 diabetes: A Mendelian randomization study. Int. J. Epidemiol. 2015, 44, 551–565. [Google Scholar] [CrossRef]
- Koyama, T.; Maekawa, M.; Ozaki, E.; Kuriyama, N.; Uehara, R. Daily consumption of coffee and eating bread at breakfast time is associated with lower visceral adipose tissue and with lower prevalence of both visceral obesity and metabolic syndrome in Japanese populations: A cross-sectional study. Nutrients 2020, 12, 3090. [Google Scholar] [CrossRef] [PubMed]
- Haidari, F.; Samadi, M.; Mohammadshahi, M.; Jalali, M.T.; Engali, K.A. Energy restriction combined with green coffee bean extract affects serum adipocytokines and the body composition in obese women. Asia Pac. J. Clin. Nutr. 2017, 26, 1048–1054. [Google Scholar]
- Zhang, H.F.; Wang, Y.L.; Gao, C.; Gu, Y.T.; Huang, J.; Wang, J.H.; Wang, J.H.; Zhang, Z. Salvianolic acid A attenuates kidney injury and inflammation by inhibiting NF-κB and p38 MAPK signaling pathways in 5/6 nephrectomized rats. Acta Pharmacol. Sin. 2018, 39, 1855–1864. [Google Scholar] [CrossRef] [Green Version]
- Ding, C.; Zhao, Y.; Shi, X.; Zhang, N.; Zu, G.; Li, Z.; Zhou, J.; Gao, D.; Lv, L.; Tian, X.; et al. New insights into salvianolic acid A action: Regulation of the TXNIP/NLRP3 and TXNIP/ChREBP pathways ameliorates HFD-induced NAFLD in rats. Sci. Rep. 2016, 6, 28743. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Qian, Q.; Ding, Q.; Zhou, L.; Fu, A.; Du, Z.; Wang, C.; Song, Z.; Li, S.; Dou, X. Activation of AMP-Activated Protein Kinase-Sirtuin 1 Pathway Contributes to Salvianolic Acid A-Induced Browning of White Adipose Tissue in High-Fat Diet Fed Male Mice. Front. Pharmacol. 2021, 12, 1361. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chen, Y.; Yan, Z.; Xu, T.T.; Wu, X.; Pi, A.; Liu, Q.; Chai, H.; Li, S.; Dou, X. Inhibition of TLR4/MAPKs Pathway Contributes to the Protection of Salvianolic Acid A Against Lipotoxicity-Induced Myocardial Damage in Cardiomyocytes and Obese Mice. Front. Pharmacol. 2021, 12, 76. [Google Scholar]
- Luna-Vital, D.; Luzardo-Ocampo, I.; Cuellar-Nuñez, M.L.; Loarca-Piña, G.; Gonzalez de Mejia, E. Maize extract rich in ferulic acid and anthocyanins prevents high-fat-induced obesity in mice by modulating SIRT1, AMPK and IL-6 associated metabolic and inflammatory pathways. J. Nutr. Biochem. 2020, 12, 1361. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, P.; Ilavenil, S.; Hwang, I.H.; Kim, D.; Choi, K.C. Ferulic acid stimulates adipocyte-specific secretory proteins to regulate adipose homeostasis in 3T3-L1 adipocytes. Molecules 2021, 26, 1984. [Google Scholar] [CrossRef] [PubMed]
- Chavanelle, V.; Otero, Y.F.; Le Joubioux, F.; Ripoche, D.; Bargetto, M.; Vluggens, A.; Montaurier, C.; Pickering, G.; Ducheix, G.; Dubray, C.; et al. Effects of Totum-63 on glucose homeostasis and postprandial glycemia: A translational study. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E1119–E1137. [Google Scholar] [CrossRef] [PubMed]
- Dupuit, M.; Chavanelle, V.; Chassaing, B.; Perriere, F.; Etienne, M.; Plissonneau, C.; Boscaro, A.; Barnich, N.; Pialoux, V.; Maugard, T.; et al. The totum-63 supplement and high-intensity interval training combination limits weight gain, improves glycemic control, and influences the composition of gut mucosa-associated bacteria in rats on a high fat diet. Nutrients 2021, 13, 1569. [Google Scholar] [CrossRef]
- Van der Zande, H.J.P.; Lambooij, J.M.; Chavanelle, V.; Zawistowska-Deniziak, A.; Otero, Y.; Otto, F.; Lantier, L.; McGuinness, O.P.; Le Joubioux, F.; Giera, M.; et al. Effects of a novel polyphenol-rich plant extract on body composition, inflammation, insulin sensitivity, and glucose homeostasis in obese mice. Int. J. Obes. 2021. [Google Scholar] [CrossRef]
- Muhlhausler, B.; Smith, S.R. Early-life origins of metabolic dysfunction: Role of the adipocyte. Trends Endocrinol. Metab. 2009, 20, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Murabayashi, N.; Sugiyama, T.; Zhang, L.; Kamimoto, Y.; Umekawa, T.; Ma, N.; Sagawa, N. Maternal high-fat diets cause insulin resistance through inflammatory changes in fetal adipose tissue. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 169, 39–44. [Google Scholar] [CrossRef]
- Lecoutre, S.; Breton, C. The cellularity of offspring’s adipose tissue is programmed by maternal nutritional manipulations. Adipocyte 2014, 3, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Challier, J.C.; Basu, S.; Bintein, T.; Minium, J.; Hotmire, K.; Catalano, P.M.; Hauguel-de Mouzon, S. Obesity in Pregnancy Stimulates Macrophage Accumulation and Inflammation in the Placenta. Placenta 2008, 29, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Denison, F.C.; Roberts, K.A.; Barr, S.M.; Norman, J.E. Obesity, pregnancy, inflammation, and vascular function. Reproduction 2010, 140, 373–385. [Google Scholar] [CrossRef]
- Kataoka, S.; Norikura, T.; Sato, S. Maternal green tea polyphenol intake during lactation attenuates kidney injury in high-fat-diet-fed male offspring programmed by maternal protein restriction in rats. J. Nutr. Biochem. 2018, 56, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Li, H. Epigallocatechin-3-gallate ameliorates hyperglycemia-induced embryonic vasculopathy and malformation by inhibition of Foxo3a activation. Am. J. Obstet. Gynecol. 2010, 203, 75.e1–75.e6. [Google Scholar] [CrossRef] [Green Version]
- Bonacasa, B.; Siow, R.C.M.; Mann, G.E. Impact of Dietary Soy Isoflavones in Pregnancy on Fetal Programming of Endothelial Function in Offspring. Microcirculation 2011, 18, 270–285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Su, S.; Yu, X.; Li, Y. Dietary epigallocatechin 3-gallate supplement improves maternal and neonatal treatment outcome of gestational diabetes mellitus: A double-blind randomised controlled trial. J. Hum. Nutr. Diet. 2017, 30, 753–758. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Developmental programming of the metabolic syndrome: Can we reprogram with resveratrol? Int. J. Mol. Sci. 2018, 19, 2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheen, J.M.; Yu, H.R.; Tain, Y.L.; Tsai, W.L.; Tiao, M.M.; Lin, I.C.; Tsai, C.C.; Lin, Y.J.; Huang, L.T. Combined maternal and postnatal high-fat diet leads to metabolic syndrome and is effectively reversed by resveratrol: A multiple-organ study. Sci. Rep. 2018, 8, 5607. [Google Scholar] [CrossRef]
- De Brito Oliveira, A.L.; Monteiro, V.V.S.; Navegantes-Lima, K.C.; Reis, J.F.; de Souza Gomes, R.; Rodrigues, D.V.S.; de França Gaspar, S.L.; Monteiro, M.C. Resveratrol role in autoimmune disease—A mini-review. Nutrients 2017, 9, 1306. [Google Scholar] [CrossRef] [Green Version]
- Truong, V.L.; Jun, M.; Jeong, W.S. Role of resveratrol in regulation of cellular defense systems against oxidative stress. BioFactors 2018, 44, 36–49. [Google Scholar] [CrossRef]
- Kim, O.Y.; Chung, J.Y.; Song, J. Effect of resveratrol on adipokines and myokines involved in fat browning: Perspectives in healthy weight against obesity. Pharmacol. Res. 2019, 148, 104411. [Google Scholar] [CrossRef]
- Jeon, B.T.; Jeong, E.A.; Shin, H.J.; Lee, Y.; Lee, D.H.; Kim, H.J.; Kang, S.S.; Cho, G.J.; Choi, W.S.; Roh, G.S. Resveratrol attenuates obesity-associated peripheral and central inflammation and improves memory deficit in mice fed a high-fat diet. Diabetes 2012, 61, 1444–1454. [Google Scholar] [CrossRef] [Green Version]
- Zou, T.; Chen, D.; Yang, Q.; Wang, B.; Zhu, M.J.; Nathanielsz, P.W.; Du, M. Resveratrol supplementation of high-fat diet-fed pregnant mice promotes brown and beige adipocyte development and prevents obesity in male offspring. J. Physiol. 2017, 595, 1547–1562. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.H.; Sheen, J.M.; Lin, I.C.; Yu, H.R.; Tiao, M.M.; Tain, Y.L.; Huang, L.T. Effects of maternal resveratrol on maternal high-fat diet/obesity with or without postnatal high-fat diet. Int. J. Mol. Sci. 2020, 21, 3428. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.Y.; Yu, H.R.; Tsai, C.C.; Huang, L.T.; Chen, C.C.; Sheen, J.M.; Tiao, M.M.; Tain, Y.L.; Lin, I.C.; Lai, Y.J.; et al. Resveratrol intake during pregnancy and lactation re-programs adiposity and ameliorates leptin resistance in male progeny induced by maternal high-fat/high sucrose plus postnatal high-fat/high sucrose diets via fat metabolism regulation. Lipids Health Dis. 2020, 19, 174. [Google Scholar] [CrossRef] [PubMed]
- Brawerman, G.M.; Kereliuk, S.M.; Brar, N.; Cole, L.K.; Seshadri, N.; Pereira, T.J.; Xiang, B.; Hunt, K.L.; Fonseca, M.A.; Hatch, G.M.; et al. Maternal resveratrol administration protects against gestational diabetes-induced glucose intolerance and islet dysfunction in the rat offspring. J. Physiol. 2019, 597, 4175–4192. [Google Scholar] [CrossRef]
- Roberts, V.H.J.; Pound, L.D.; Thorn, S.R.; Gillingham, M.B.; Thornburg, K.L.; Friedman, J.E.; Frias, A.E.; Grove, K.L. Beneficial and cautionary outcomes of resveratrol supplementation in pregnant nonhuman primates. FASEB J. 2014, 28, 2466–2477. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic representation of molecular mechanisms underlying the anti-obesity effects of the main polyphenols.
Figure 1.
Schematic representation of molecular mechanisms underlying the anti-obesity effects of the main polyphenols.

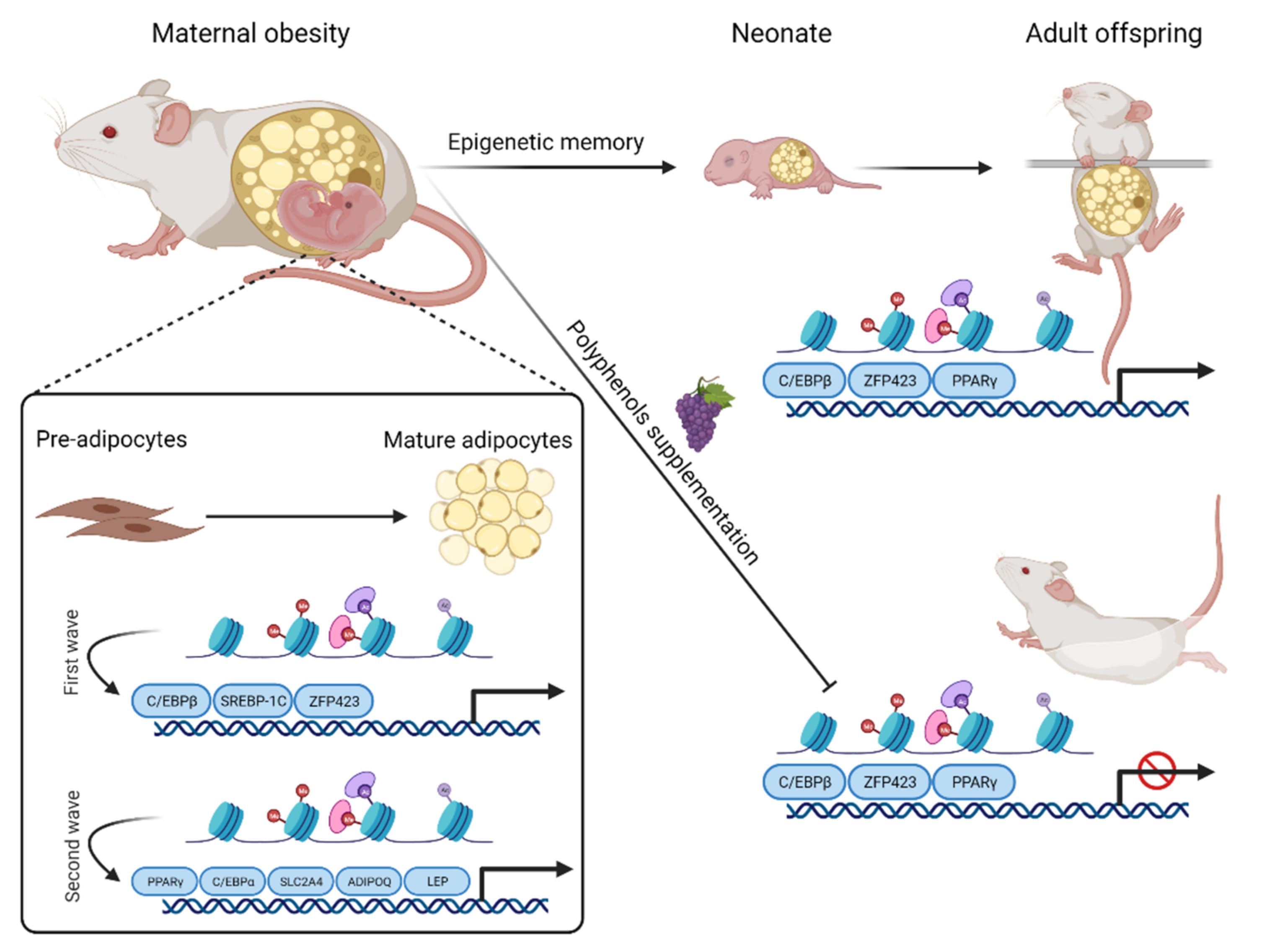
Figure 2.
Schematic representation of polyphenol effect on obesity-induced maternal programming. The recruitment of the first wave of transcription factors (C/EBPβ, SREBP-1C, and ZFP423) take place during adipocyte differentiation, leading to the conversion of the histone bivalent state to an active state. These transcription factors, in turn, promote an open state of chromatin in regions containing genes involved in the second wave of transcription factors, among which PPARγ and CEBPα stand out. The second wave of transcription factors induces the expression of pro-adipogenic genes such as LEP, SLC2A4, and ADIPOQ. Maternal obesity is also involved in the regulation of pro-adipogenic transcription factors such as ZFP423, C/EBPβ, and PPARγ during adipogenesis in the perinatal period and affects the offspring. Polyphenol supplementation could counteract the detrimental effects of maternal obesity programming on the progeny by negatively regulating adipogenesis via inhibiting PPARγ, ZFP423, and C/EBPα.
Figure 2.
Schematic representation of polyphenol effect on obesity-induced maternal programming. The recruitment of the first wave of transcription factors (C/EBPβ, SREBP-1C, and ZFP423) take place during adipocyte differentiation, leading to the conversion of the histone bivalent state to an active state. These transcription factors, in turn, promote an open state of chromatin in regions containing genes involved in the second wave of transcription factors, among which PPARγ and CEBPα stand out. The second wave of transcription factors induces the expression of pro-adipogenic genes such as LEP, SLC2A4, and ADIPOQ. Maternal obesity is also involved in the regulation of pro-adipogenic transcription factors such as ZFP423, C/EBPβ, and PPARγ during adipogenesis in the perinatal period and affects the offspring. Polyphenol supplementation could counteract the detrimental effects of maternal obesity programming on the progeny by negatively regulating adipogenesis via inhibiting PPARγ, ZFP423, and C/EBPα.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fortunato, I.M.; dos Santos, T.W.; Ferraz, L.F.C.; Santos, J.C.; Ribeiro, M.L. Effect of Polyphenols Intake on Obesity-Induced Maternal Programming. Nutrients 2021, 13, 2390. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072390
AMA Style
Fortunato IM, dos Santos TW, Ferraz LFC, Santos JC, Ribeiro ML. Effect of Polyphenols Intake on Obesity-Induced Maternal Programming. Nutrients. 2021; 13(7):2390. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072390
Chicago/Turabian StyleFortunato, Isabela Monique, Tanila Wood dos Santos, Lucio Fábio Caldas Ferraz, Juliana Carvalho Santos, and Marcelo Lima Ribeiro. 2021. "Effect of Polyphenols Intake on Obesity-Induced Maternal Programming" Nutrients 13, no. 7: 2390. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072390
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.