
Leptin Improves Parameters of Brown Adipose Tissue Thermogenesis in Lipodystrophic Mice

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Temperature Measurements
2.3. Immunoblot Analysis
2.4. Histological Analysis
2.5. Quantitative Real-Time PCR Analysis
2.6. Immunoassays
2.7. Data Analysis and Statistics
3. Results
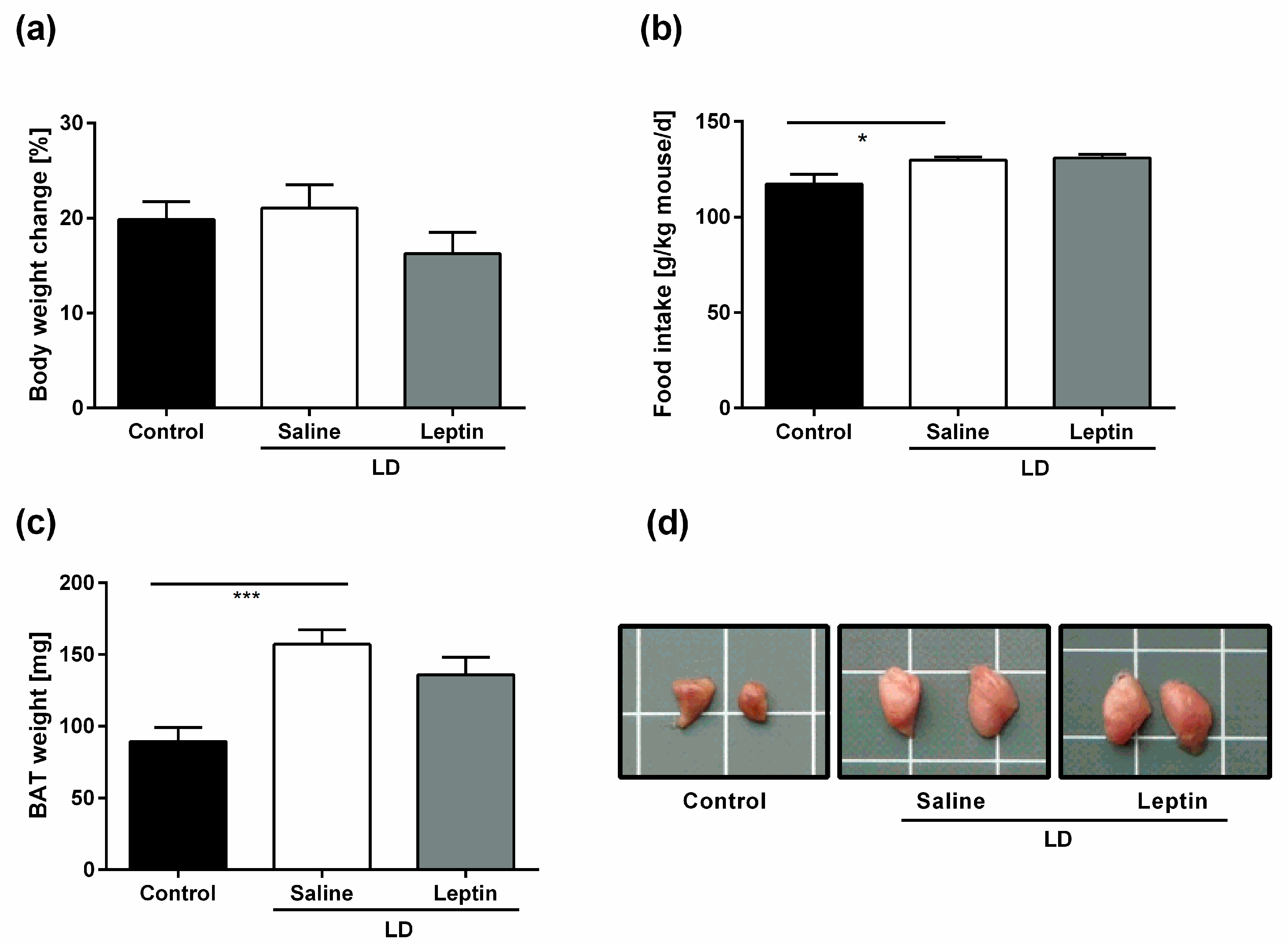
3.1. Impact of Chronic Leptin Treatment on Body Weight, Food Intake, and BAT Weight in LD Mice
3.2. Impact of Chronic Leptin Treatment on Body Temperature and BAT UCP1 Expression in LD Mice
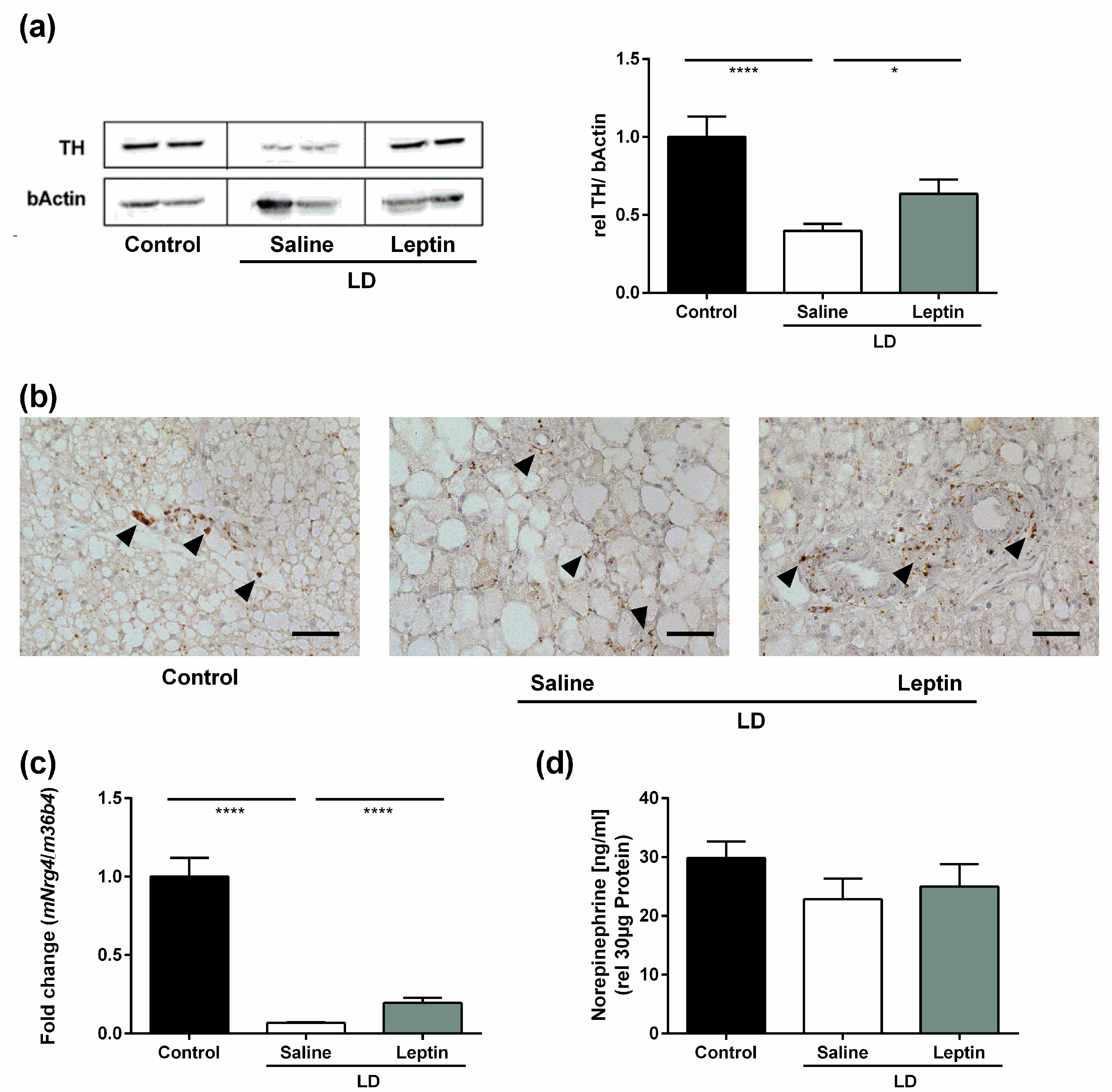
3.3. Impact of Chronic Leptin Treatment on BAT Sympathetic Markers in LD Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knebel, B.; Müller-Wieland, D.; Kotzka, J. Lipodystrophies—Disorders of the Fatty Tissue. Int. J. Mol. Sci. 2020, 21, 8778. [Google Scholar] [CrossRef] [PubMed]
- Haque, W.A.; Shimomura, I.; Matsuzawa, Y.; Garg, A. Serum Adiponectin and Leptin Levels in Patients with Lipodystrophies. J. Clin. Endocrinol. Metab. 2002, 87, 2395. [Google Scholar] [CrossRef]
- Miehle, K.; Ebert, T.; Kralisch, S.; Hoffmann, A.; Kratzsch, J.; Schlögl, H.; Stumvoll, M.; Fasshauer, M. Progranulin is increased in human and murine lipodystrophy. Diabetes Res. Clin. Pract. 2016, 120, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Miehle, K.; Ebert, T.; Kralisch, S.; Hoffmann, A.; Kratzsch, J.; Schlögl, H.; Stumvoll, M.; Fasshauer, M. Circulating serum chemerin levels are elevated in lipodystrophy. Clin. Endocrinol. 2015, 84, 932–938. [Google Scholar] [CrossRef]
- Oral, E.A.; Simha, V.; Ruiz, E.; Andewelt, A.; Premkumar, A.; Snell, P.; Wagner, A.J.; DePaoli, A.M.; Reitman, M.L.; Taylor, S.I.; et al. Leptin-Replacement Therapy for Lipodystrophy. N. Engl. J. Med. 2002, 346, 570–578. [Google Scholar] [CrossRef]
- Chan, J.L.; Lutz, K.; Cochran, E.; Huang, W.; Peters, Y.; Weyer, C.; Gorden, P. Clinical Effects of Long-Term Metreleptin Treatment in Patients with Lipodystrophy. Endocr. Pract. 2011, 17, 922–932. [Google Scholar] [CrossRef] [Green Version]
- Sinha, G. Leptin therapy gains FDA approval. Nat. Biotechnol. 2014, 32, 300–301. [Google Scholar] [CrossRef] [PubMed]
- EMA. Metreleptin for Treatment of Familial Partial Lipodystrophy (Orphan Decision Number: EU/3/12/1022); European Medicines Agency: London, UK, 2018. [Google Scholar]
- Eifler, L.; Hoffmann, A.; Wagner, I.V.; Klöting, N.; Sahlin, L.; Ebert, T.; Jessnitzer, B.; Lössner, U.; Stumvoll, M.; Söder, O.; et al. Leptin restores markers of female fertility in lipodystrophy. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3292–3297. [Google Scholar] [CrossRef]
- Liu, X.J.; Wu, X.Y.; Wang, H.; Wang, S.X.; Kong, W.; Zhang, L.; Liu, G.; Huang, W. Renal injury in Seipin-deficient lipodystrophic mice and its reversal by adipose tissue transplantation or leptin administration alone: Adipose tissue-kidney crosstalk. FASEB J. 2018, 32, 5550–5562. [Google Scholar] [CrossRef] [PubMed]
- Bruder-Nascimento, T.; Faulkner, J.L.; Haigh, S.; Kennard, S.; Antonova, G.; Patel, V.S.; Fulton, D.J.; Chen, W.; De Chantemèle, E.J.B. Leptin Restores Endothelial Function via Endothelial PPARγ-Nox1–Mediated Mechanisms in a Mouse Model of Congenital Generalized Lipodystrophy. Hypertension 2019, 74, 1399–1408. [Google Scholar] [CrossRef]
- Nedergaard, J.; Cannon, B. The Changed Metabolic World with Human Brown Adipose Tissue: Therapeutic Visions. Cell Metab. 2010, 11, 268–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, B.; Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- McNeill, B.T.; Morton, N.M.; Stimson, R.H. Substrate Utilization by Brown Adipose Tissue: What’s Hot and What’s Not? Front. Endocrinol. 2020, 11, 571659. [Google Scholar] [CrossRef]
- Rothwell, N.J.; Stock, M.J. Effects of denervating brown adipose tissue on the responses to cold, hyperphagia and noradrenaline treatment in the rat. J. Physiol. 1984, 355, 457–463. [Google Scholar] [CrossRef]
- Wang, P.; Loh, K.H.; Wu, M.; Morgan, D.A.; Schneeberger, M.; Yu, X.; Chi, J.; Kosse, C.; Kim, D.; Rahmouni, K.; et al. A leptin–BDNF pathway regulating sympathetic innervation of adipose tissue. Nat. Cell Biol. 2020, 583, 839–844. [Google Scholar] [CrossRef]
- Shimomura, I.; Hammer, R.E.; Richardson, J.A.; Ikemoto, S.; Bashmakov, Y.; Goldstein, J.L.; Brown, M.S. Insulin resistance and diabetes mellitus in transgenic mice expressing nuclear SREBP-1c in adipose tissue: Model for congenital generalized lipodystrophy. Genes Dev. 1998, 12, 3182–3194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimomura, I.; Hammer, R.E.; Ikemoto, S.; Brown, M.S.; Goldstein, J.L. Leptin reverses insulin resistance and diabetes mellitus in mice with congenital lipodystrophy. Nat. Cell Biol. 1999, 401, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, M.; Fujisaka, S.; Cai, W.; Winnay, J.N.; Konishi, M.; O’Neill, B.T.; Li, M.; García-Martín, R.; Takahashi, H.; Hu, J.; et al. Adipocyte Dynamics and Reversible Metabolic Syndrome in Mice with an Inducible Adipocyte-Specific Deletion of the Insulin Receptor. Cell Metab. 2017, 25, 448–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortés, V.A.; Cautivo, K.M.; Rong, S.; Garg, A.; Horton, J.D.; Agarwal, A.K. Leptin ameliorates insulin resistance and hepatic steatosis in Agpat2 lipodystrophic mice independent of hepatocyte leptin receptors. J. Lipid Res. 2014, 55, 276–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolze, F.; Bast, A.; Mocek, S.; Morath, V.; Yuan, D.; Rink, N.; Schlapschy, M.; Zimmermann, A.; Heikenwalder, M.; Skerra, A.; et al. Treatment of diet-induced lipodystrophic C57BL/6J mice with long-acting PASylated leptin normalises insulin sensitivity and hepatic steatosis by promoting lipid utilisation. Diabetologia 2016, 59, 2005–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucher, J.; Softic, S.; El Ouaamari, A.; Krumpoch, M.T.; Kleinridders, A.; Kulkarni, R.N.; O’Neill, B.T.; Kahn, C.R. Differential Roles of Insulin and IGF-1 Receptors in Adipose Tissue Development and Function. Diabetes 2016, 65, 2201–2213. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, L.; Ebihara, K.; Kusakabe, T.; Aotani, D.; Yamamoto-Kataoka, S.; Sakai, T.; Aizawa-Abe, M.; Yamamoto, Y.; Fujikura, J.; Hayashi, T.; et al. Leptin Activates Hepatic 5′-AMP-activated Protein Kinase through Sympathetic Nervous System and α1-Adrenergic Receptor: A potential mechanism for improvement of fatty liver in lipodystrophy by leptin. J. Biol. Chem. 2012, 287, 40441–40447. [Google Scholar] [CrossRef] [Green Version]
- Nagao, K.; Inoue, N.; Ujino, Y.; Higa, K.; Shirouchi, B.; Wang, Y.-M.; Yanagita, T. Effect of leptin infusion on insulin sensitivity and lipid metabolism in diet-induced lipodystrophy model mice. Lipids Health Dis. 2008, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavrilova, O.; Leon, L.R.; Marcus-Samuels, B.; Mason, M.M.; Castle, A.L.; Refetoff, S.; Vinson, C.; Reitman, M. Torpor in mice is induced by both leptin-dependent and -independent mechanisms. Proc. Natl. Acad. Sci. USA 1999, 96, 14623–14628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, A.; Ebert, T.; Klöting, N.; Dokas, J.; Jeromin, F.; Jessnitzer, B.; Burkhardt, R.; Fasshauer, M.; Kralisch, S. Leptin dose-dependently decreases atherosclerosis by attenuation of hypercholesterolemia and induction of adiponectin. Biochim. Biophys. Acta Mol. Basis Dis. 2016, 1862, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Weiner, J.; Kranz, M.; Klöting, N.; Kunath, A.; Steinhoff, K.; Rijntjes, E.; Köhrle, J.; Zeisig, V.; Hankir, M.; Gebhardt, C.; et al. Thyroid hormone status defines brown adipose tissue activity and browning of white adipose tissues in mice. Sci. Rep. 2016, 6, 38124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, A.W.; Hoefig, C.S.; Abreu-Vieira, G.; de Jong, J.M.; Petrovic, N.; Mittag, J.; Cannon, B.; Nedergaard, J. Leptin Raises Defended Body Temperature without Activating Thermogenesis. Cell Rep. 2016, 14, 1621–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Pellegrinelli, V.; Peirce, V.J.; Howard, L.; Virtue, S.; Türei, D.; Senzacqua, M.; Frontini, A.; Dalley, J.W.; Horton, A.R.; Bidault, G.; et al. Adipocyte-secreted BMP8b mediates adrenergic-induced remodeling of the neuro-vascular network in adipose tissue. Nat. Commun. 2018, 9, 1–18. [Google Scholar] [CrossRef]
- Weiner, J.; Hankir, M.; Heiker, J.T.; Fenske, W.; Krause, K. Thyroid hormones and browning of adipose tissue. Mol. Cell. Endocrinol. 2017, 458, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Deem, J.D.; Muta, K.; Ogimoto, K.; Nelson, J.T.; Velasco, K.R.; Kaiyala, K.J.; Morton, G.J. Leptin regulation of core body temperature involves mechanisms independent of the thyroid axis. Am. J. Physiol.-Endocrinol. Metab. 2018, 315, E552–E564. [Google Scholar] [CrossRef]
- Grover, A.; Quaye, E.; Brychta, R.J.; Christensen, J.; Startzell, M.S.; Meehan, C.A.; Valencia, A.; Marshall, B.; Chen, K.Y.; Brown, R.J. Leptin Decreases Energy Expenditure Despite Increased Thyroid Hormone in Patients with Lipodystrophy. J. Clin. Endocrinol. Metab. 2021. [Google Scholar] [CrossRef]
- Muzik, O.; Mangner, T.J.; Leonard, W.R.; Kumar, A.; Granneman, J.G. Sympathetic Innervation of Cold-Activated Brown and White Fat in Lean Young Adults. J. Nucl. Med. 2017, 58, 799–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chondronikola, M.; Volpi, E.; Børsheim, E.; Porter, C.; Annamalai, P.; Enerbäck, S.; Lidell, M.E.; Saraf, M.K.; Labbe, S.M.; Hurren, N.M.; et al. Brown Adipose Tissue Improves Whole-Body Glucose Homeostasis and Insulin Sensitivity in Humans. Diabetes 2014, 63, 4089–4099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chondronikola, M.; Volpi, E.; Børsheim, E.; Porter, C.; Saraf, M.K.; Annamalai, P.; Yfanti, C.; Chao, T.; Wong, D.; Shinoda, K.; et al. Brown Adipose Tissue Activation Is Linked to Distinct Systemic Effects on Lipid Metabolism in Humans. Cell Metab. 2016, 23, 1200–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berbée, J.F.; Boon, M.R.; Khedoe, P.P.S.; Bartelt, A.; Schlein, C.; Worthmann, A.; Kooijman, S.; Hoeke, G.; Mol, I.M.; John, C.; et al. Brown fat activation reduces hypercholesterolaemia and protects from atherosclerosis development. Nat. Commun. 2015, 6, 6356. [Google Scholar] [CrossRef] [Green Version]
- Bartelt, A.; Bruns, O.T.; Reimer, R.; Hohenberg, H.; Ittrich, H.; Peldschus, K.; Kaul, M.G.; Tromsdorf, U.I.; Weller, H.; Waurisch, C.; et al. Brown adipose tissue activity controls triglyceride clearance. Nat. Med. 2011, 17, 200–205. [Google Scholar] [CrossRef]
- Fischer, A.W.; Cannon, B.; Nedergaard, J. No insulating effect of obesity, neither in mice nor in humans. Am. J. Physiol.-Endocrinol. Metab. 2019, 317, E952–E953. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffmann, A.; Ebert, T.; Hankir, M.K.; Flehmig, G.; Klöting, N.; Jessnitzer, B.; Lössner, U.; Stumvoll, M.; Blüher, M.; Fasshauer, M.; et al. Leptin Improves Parameters of Brown Adipose Tissue Thermogenesis in Lipodystrophic Mice. Nutrients 2021, 13, 2499. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13082499
Hoffmann A, Ebert T, Hankir MK, Flehmig G, Klöting N, Jessnitzer B, Lössner U, Stumvoll M, Blüher M, Fasshauer M, et al. Leptin Improves Parameters of Brown Adipose Tissue Thermogenesis in Lipodystrophic Mice. Nutrients. 2021; 13(8):2499. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13082499
Chicago/Turabian StyleHoffmann, Annett, Thomas Ebert, Mohammed K. Hankir, Gesine Flehmig, Nora Klöting, Beate Jessnitzer, Ulrike Lössner, Michael Stumvoll, Matthias Blüher, Mathias Fasshauer, and et al. 2021. "Leptin Improves Parameters of Brown Adipose Tissue Thermogenesis in Lipodystrophic Mice" Nutrients 13, no. 8: 2499. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13082499