
Phytochemical Combination (p-Synephrine, p-Octopamine Hydrochloride, and Hispidulin) for Improving Obesity in Obese Mice Induced by High-Fat Diet

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Adipogenic Differentiation
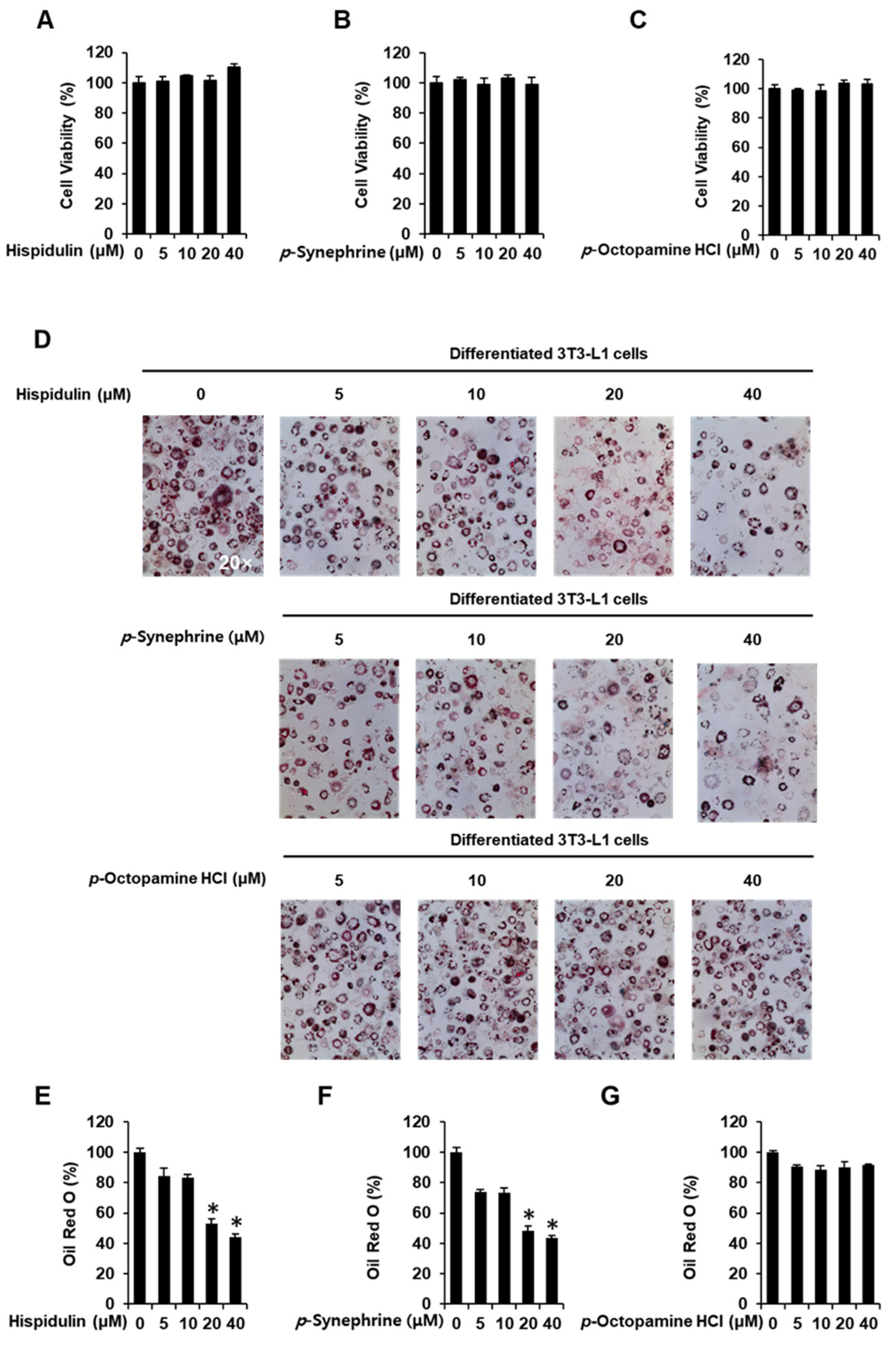
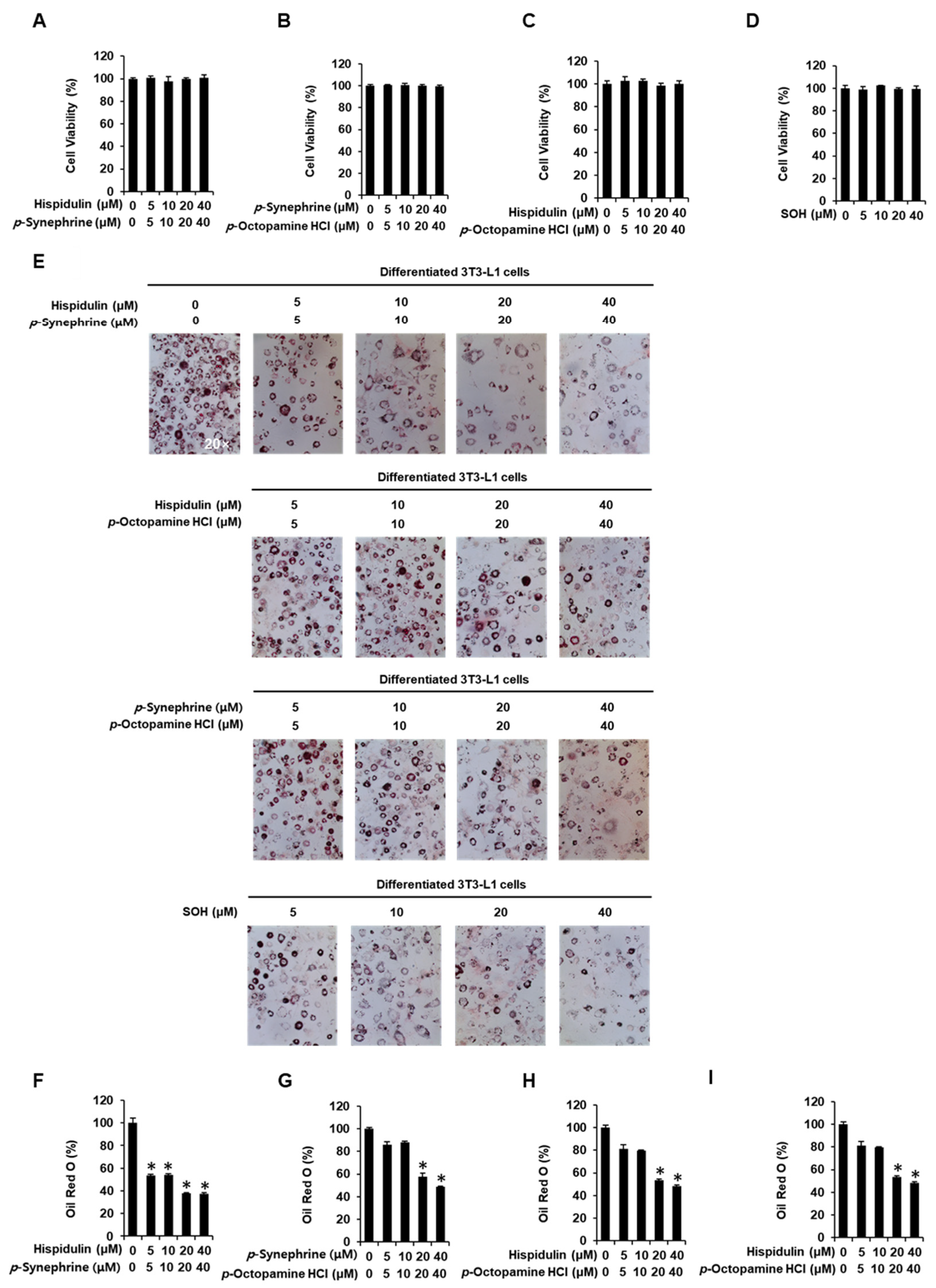
2.2. Cell Viability
2.3. Oil Red O Staining
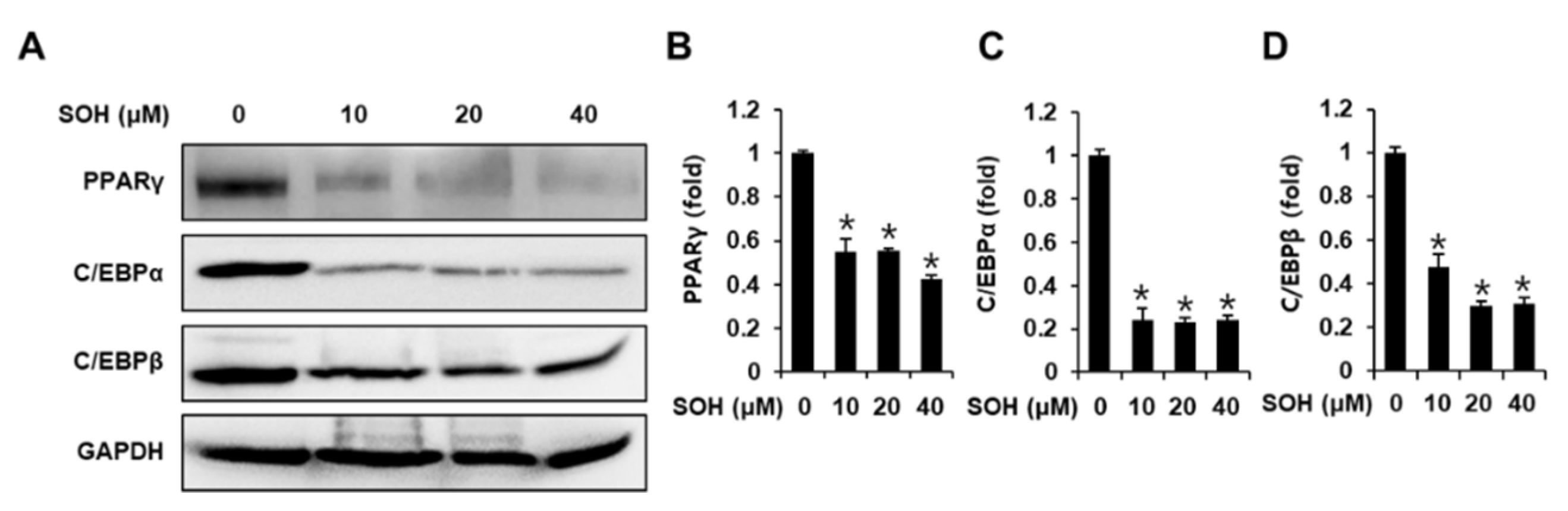
2.4. Western Blotting Analysis
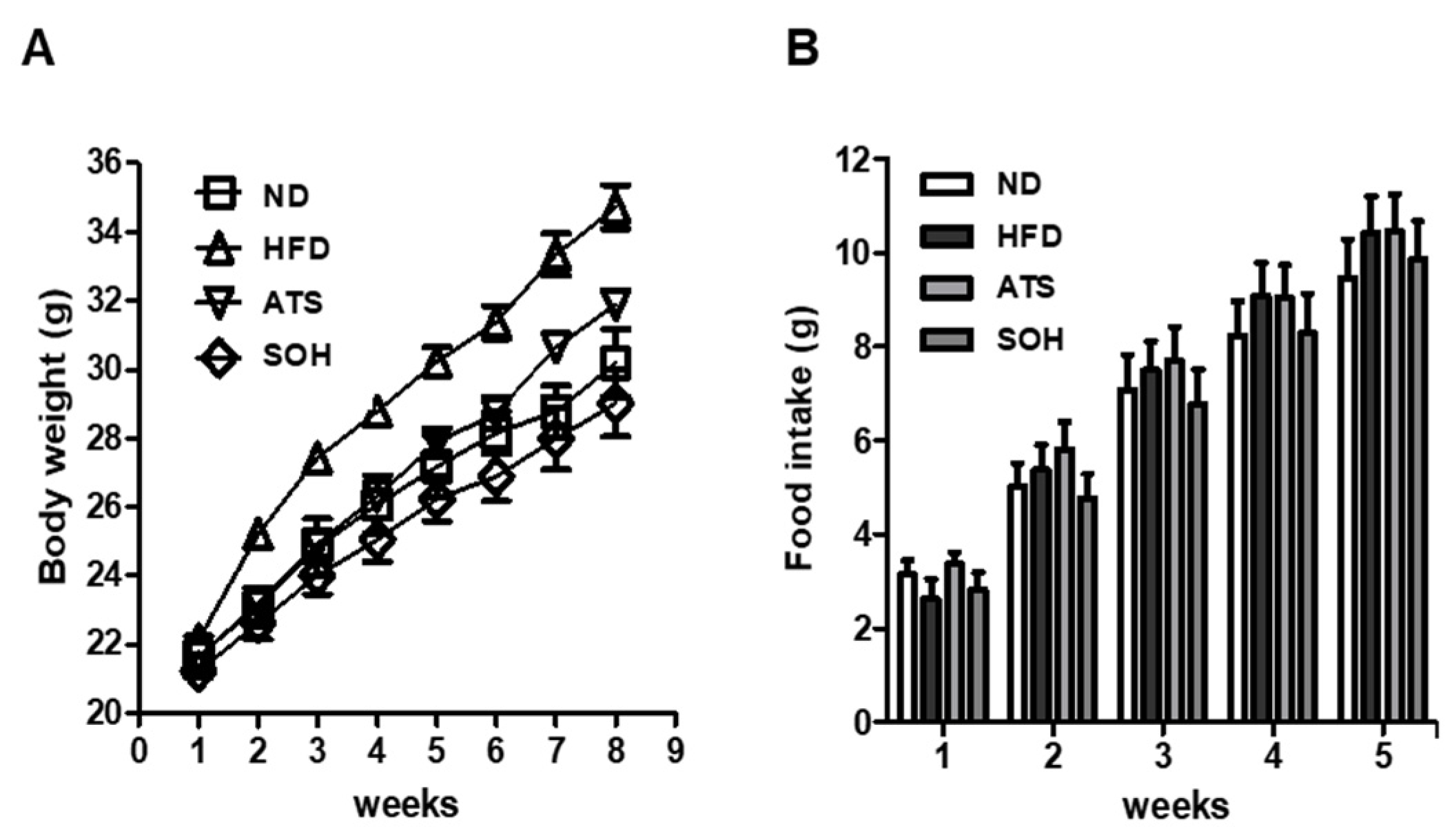
2.5. Study Animals and Experimental Design
2.6. Blood Biochemistry
2.7. Statistical Analyses
3. Results
3.1. Inhibitory Effects of Hispidulin, p-Synephrine, and p-Octopamine HCl on Adipogenesis in 3T3-L1 Preadipocytes
3.2. Inhibitory Effects of SOH on Adipogenesis in 3T3-L1 Preadipocytes
3.3. Inhibitory Effects of SOH on the Expression of Proteins Involved in Adipogenesis of Differentiated 3T3L-1 Cells
3.4. Effects of SOH on Body Weight and Food Intake in HFD-Induced Obese Mice
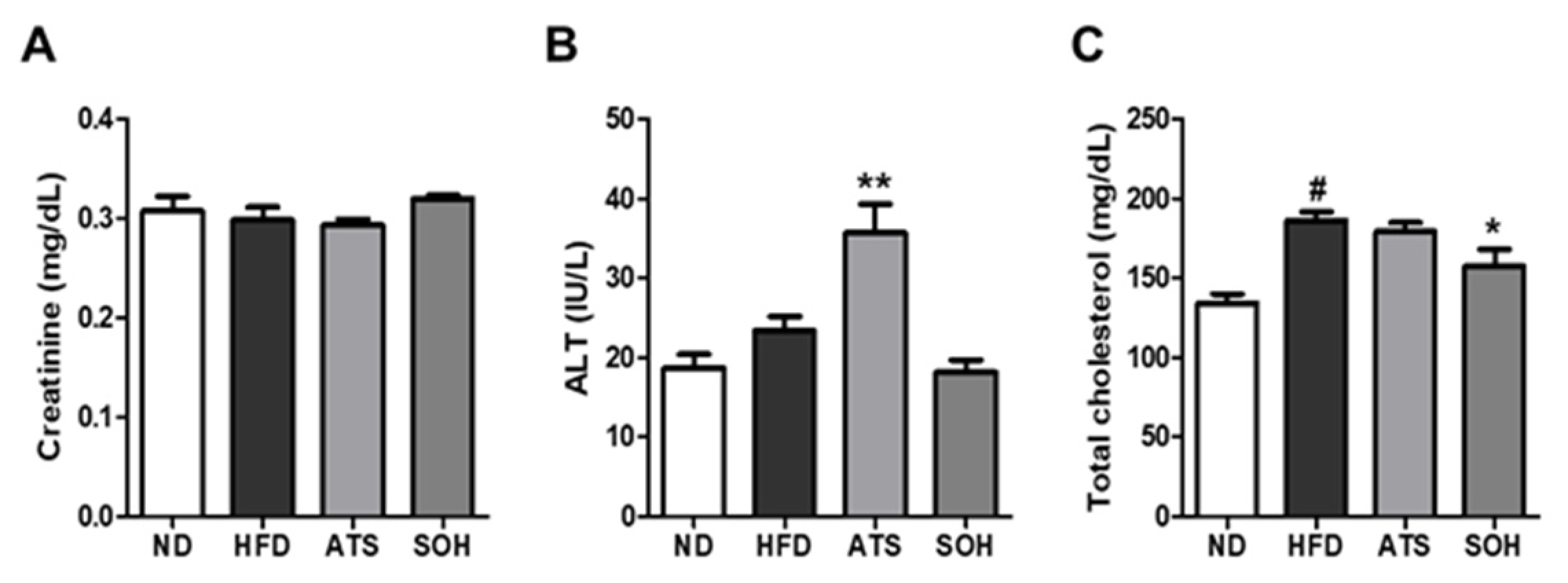
3.5. Effects of SOH on Blood Biochemistry in HFD-Induced Obese Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| C/EBPα | adenosine monophosphate |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| p-octopamine HCl | p-Octopamine hydrochloride |
| SOH | a mixture of p-synephrine, p-octopamine HCl, and hispidulin |
| P/S | penicillin/streptomycin |
| FBS | fetal bovine serum |
| ORO | Oil Red O solution |
| ND | normal diet control group |
| CMC | sodium carboxyl methyl cellulose |
| HFD | high-fat diet |
| ATS | atorvastatin |
| EDTA | ethylenediaminetetraacetic acid |
| ALT | alanine aminotransferase |
| SEM | standard error of the mean |
References
- Maric, C.; Hall, J.E. Obesity, metabolic syndrome and diabetic nephropathy. Contrib. Nephrol. 2011, 170, 28–35. [Google Scholar] [PubMed] [Green Version]
- Prentice, A.; Jebb, S. Energy intake/physical activity interactions in the homeostasis of body weight regulation. Nutr. Rev. 2004, 62, S98–S104. [Google Scholar] [CrossRef] [PubMed]
- Soria, A.; D’Alessandro, M.E.; Lombardo, Y.B. Duration of feeding on a sucrose-rich diet determines metabolic and morphological changes in rat adipocytes. J. Appl. Physiol. 2001, 91, 2109–2116. [Google Scholar] [CrossRef] [PubMed]
- Hausman, D.; DiGirolamo, M.; Bartness, T.; Hausman, G.; Martin, R. The biology of white adipocyte proliferation. Obes. Rev. 2001, 2, 239–254. [Google Scholar] [CrossRef]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBPα and PPARγ controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell 1999, 3, 151–158. [Google Scholar] [CrossRef]
- Koh, Y.-K.; Lee, M.-Y.; Kim, J.-W.; Kim, M.; Moon, J.-S.; Lee, Y.-J.; Ahn, Y.-H.; Kim, K.-S. Lipin1 is a key factor for the maturation and maintenance of adipocytes in the regulatory network with CCAAT/enhancer-binding protein α and peroxisome proliferator-activated receptor γ2. J. Biol. Chem. 2008, 283, 34896–34906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.H.; Gadde, K.M. Clinical utility of phentermine/topiramate (Qsymia™) combination for the treatment of obesity. Diabetes Metab. Syndr. Obes. Targets Ther. 2013, 6, 131–139. [Google Scholar]
- Vermaak, I.; Viljoen, A.M.; Hamman, J.H. Natural products in anti-obesity therapy. Nat. Prod. Rep. 2011, 28, 1493–1533. [Google Scholar] [CrossRef]
- Roh, C.; Jung, U.; Jo, S.-K. Screening of anti-obesity agent from herbal mixtures. Molecules 2012, 17, 3630–3638. [Google Scholar] [CrossRef]
- Stohs, S.J.; Preuss, H.G.; Keith, S.C.; Keith, P.L.; Miller, H.; Kaats, G.R. Effects of p-synephrine alone and in combination with selected bioflavonoids on resting metabolism, blood pressure, heart rate and self-reported mood changes. Int. J. Med. Sci. 2011, 8, 295. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Kwak, H.J.; Kim, B.H.; Kim, S.H.; Kim, D.-W.; Kang, K.S. Combined anti-adipogenic effects of hispidulin and p-synephrine on 3T3-L1 adipocytes. Biomolecules 2021, 11, 1764. [Google Scholar] [CrossRef] [PubMed]
- Ferrandiz, M.; Bustos, G.; Paya, M.; Gunasegaran, R.; Alcaraz, M. Hispidulin protection against hepatotoxicity induced by bromobenzene in mice. Life Sci. 1994, 55, PL145–PL150. [Google Scholar] [CrossRef]
- Kim, K.-W.; Kim, H.-D.; Jung, J.-S.; Woo, R.-S.; Kim, H.-S.; Suh, H.-W.; Kim, Y.-H.; Song, D.-K. Characterization of antidepressant-like effects of p-synephrine stereoisomers. Naunyn Schmiedebergs Arch. Pharmacol. 2001, 364, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.; Patel, D.K. Medicinal importance, pharmacological activities, and analytical aspects of hispidulin: A concise report. J. Tradit. Complement. Med. 2017, 7, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J. Physiological functions and pharmacological and toxicological effects of p-octopamine. Drug Chem. Toxicol. 2015, 38, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Preuss, H.G.; Shara, M. The safety of Citrus aurantium (bitter orange) and its primary protoalkaloid p-synephrine. Phytother. Res. 2011, 25, 1421–1428. [Google Scholar] [CrossRef]
- Bour, S.; Visentin, V.; Prévot, D.; Carpéné, C. Moderate weight-lowering effect of octopamine treatment in obese Zucker rats. J. Physiol. Biochem. 2003, 59, 175–182. [Google Scholar] [CrossRef]
- Mestrovic, N. Uber die behandlung hypotoner zustande bei chirugischen patienten. Z. Allg. 1972, 48, 1301–1303. [Google Scholar]
- Stucke, W. Die behandlung hypotoner kreislaufstorungen mit Norphen retard. Z. Allg. 1972, 48, 240–243. [Google Scholar]
- Madsen, L.; Petersen, R.K.; Kristiansen, K. Regulation of adipocyte differentiation and function by polyunsaturated fatty acids. Biochim. Biophys. Acta Mol. Basis Dis. 2005, 1740, 266–286. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Li, X.; Tang, Q.-Q. Transcriptional regulation of adipocyte differentiation: A central role for CCAAT/enhancer-binding protein (C/EBP) β. J. Biol. Chem. 2015, 290, 755–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Andrade, R.S.B.; de Carvalho França, L.F.; Pessoa, L.D.S.; de Almeida Arrais Landim, B.; Rodrigues, A.A.; Alves, E.H.P.; Lenardo, D.D.; Nascimento, H.M.S.; de Melo Sousa, F.B.; Barbosa, A.L.D.R. High-fat diet aggravates the liver disease caused by periodontitis in rats. J. Periodontol. 2019, 90, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the metabolic syndrome: A joint interim statement of the international diabetes federation task force on epidemiology and prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [PubMed] [Green Version]
- Kapoor, N.; Feingold, K.R.; Anawalt, B.; Boyce, A.; Chrousos, G.; de Herder, W.W.; Dhatariya, K.; Dungan, K.; Hershman, J.M.; Hofland, J.; et al. Thin Fat Obesity: The Tropical Phenotype of Obesity. In Endotext [Internet]; MDText.com, Inc.: South Dartmouth, MA, USA, 2021. [Google Scholar]
- Szczygielska, A.; Widomska, S.; Jaraszkiewicz, M.; Knera, P.; Muc, K. Blood lipids profile in obese or overweight patients. Ann. Univ. Mariae Curie Sklodowska Med. 2003, 58, 343–349. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Lee, J.H.; Kim, B.H.; Lee, S.; Kim, D.-W.; Kang, K.S. Phytochemical Combination (p-Synephrine, p-Octopamine Hydrochloride, and Hispidulin) for Improving Obesity in Obese Mice Induced by High-Fat Diet. Nutrients 2022, 14, 2164. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14102164
Lee D, Lee JH, Kim BH, Lee S, Kim D-W, Kang KS. Phytochemical Combination (p-Synephrine, p-Octopamine Hydrochloride, and Hispidulin) for Improving Obesity in Obese Mice Induced by High-Fat Diet. Nutrients. 2022; 14(10):2164. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14102164
Chicago/Turabian StyleLee, Dahae, Ji Hwan Lee, Byoung Ha Kim, Sanghyun Lee, Dong-Wook Kim, and Ki Sung Kang. 2022. "Phytochemical Combination (p-Synephrine, p-Octopamine Hydrochloride, and Hispidulin) for Improving Obesity in Obese Mice Induced by High-Fat Diet" Nutrients 14, no. 10: 2164. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14102164