
Receptor Mediated Effects of Advanced Glycation End Products (AGEs) on Innate and Adaptative Immunity: Relevance for Food Allergy

 , , , , and
, , , , and
Abstract
:1. Introduction
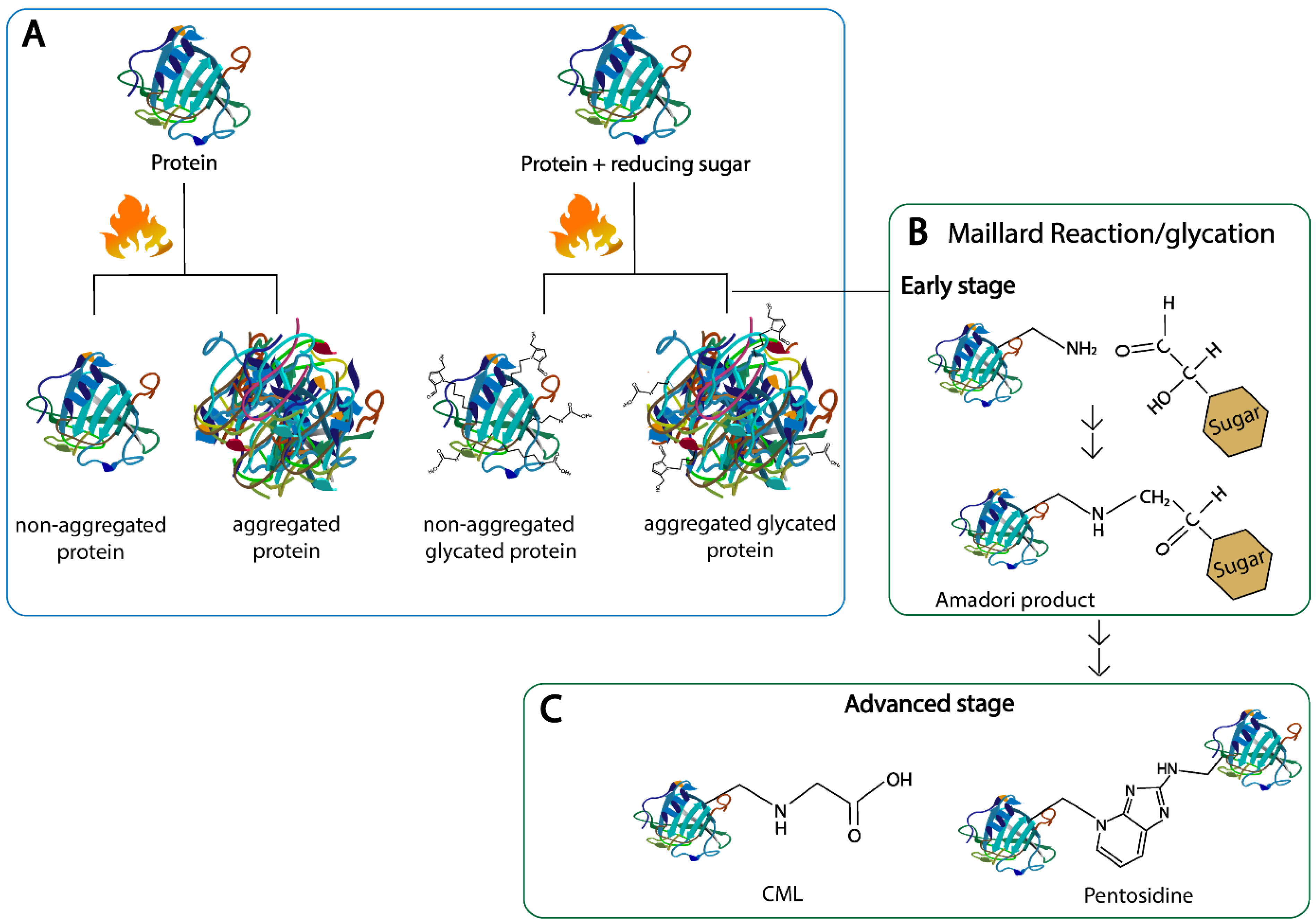
2. Maillard Reaction Products (MRPs)
2.1. Formation and Structural Changes in Proteins
2.2. Endogenous AGEs: Formation and Structure
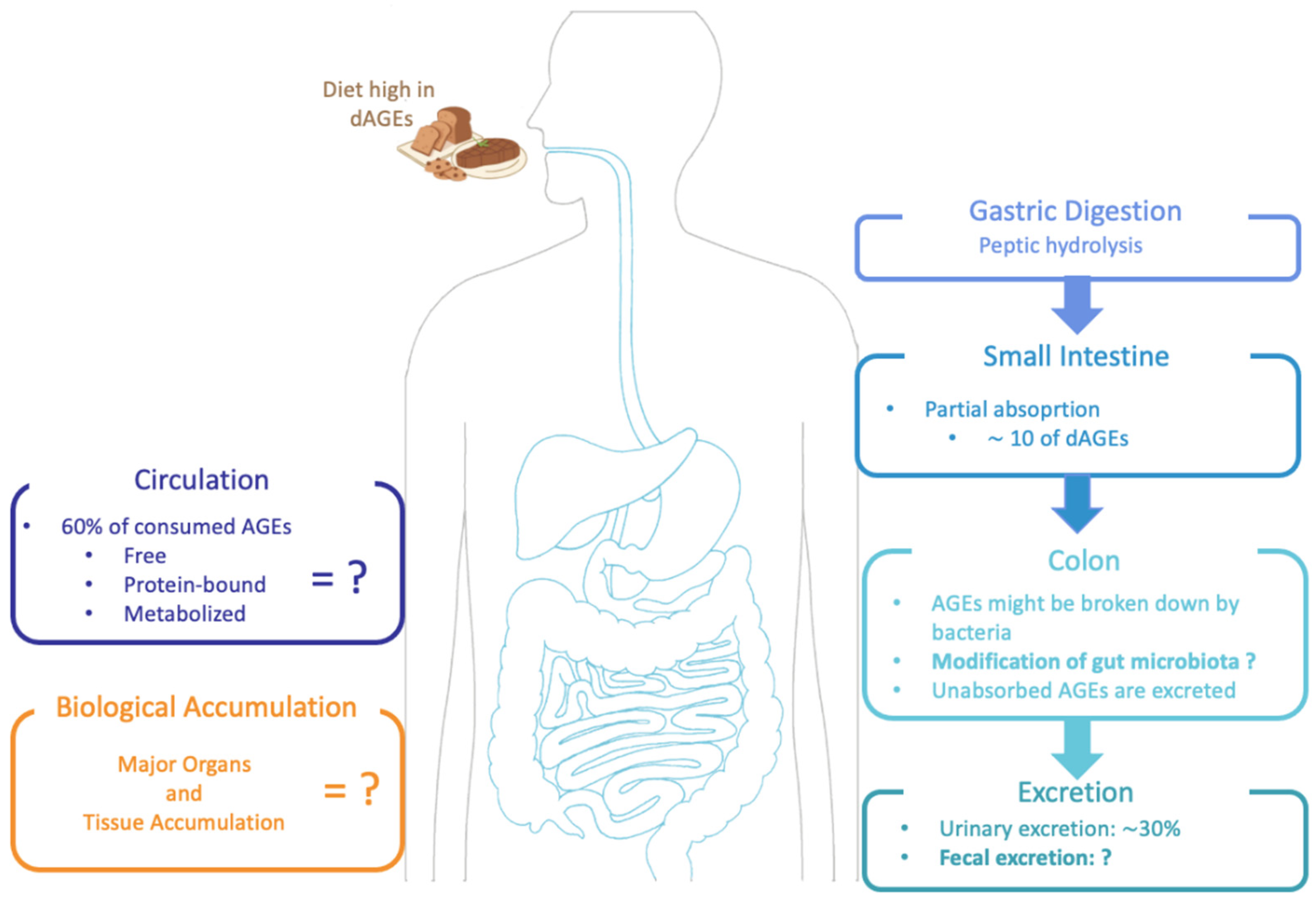
2.3. Bioavailability of Dietary AGES Induced by Food Processing
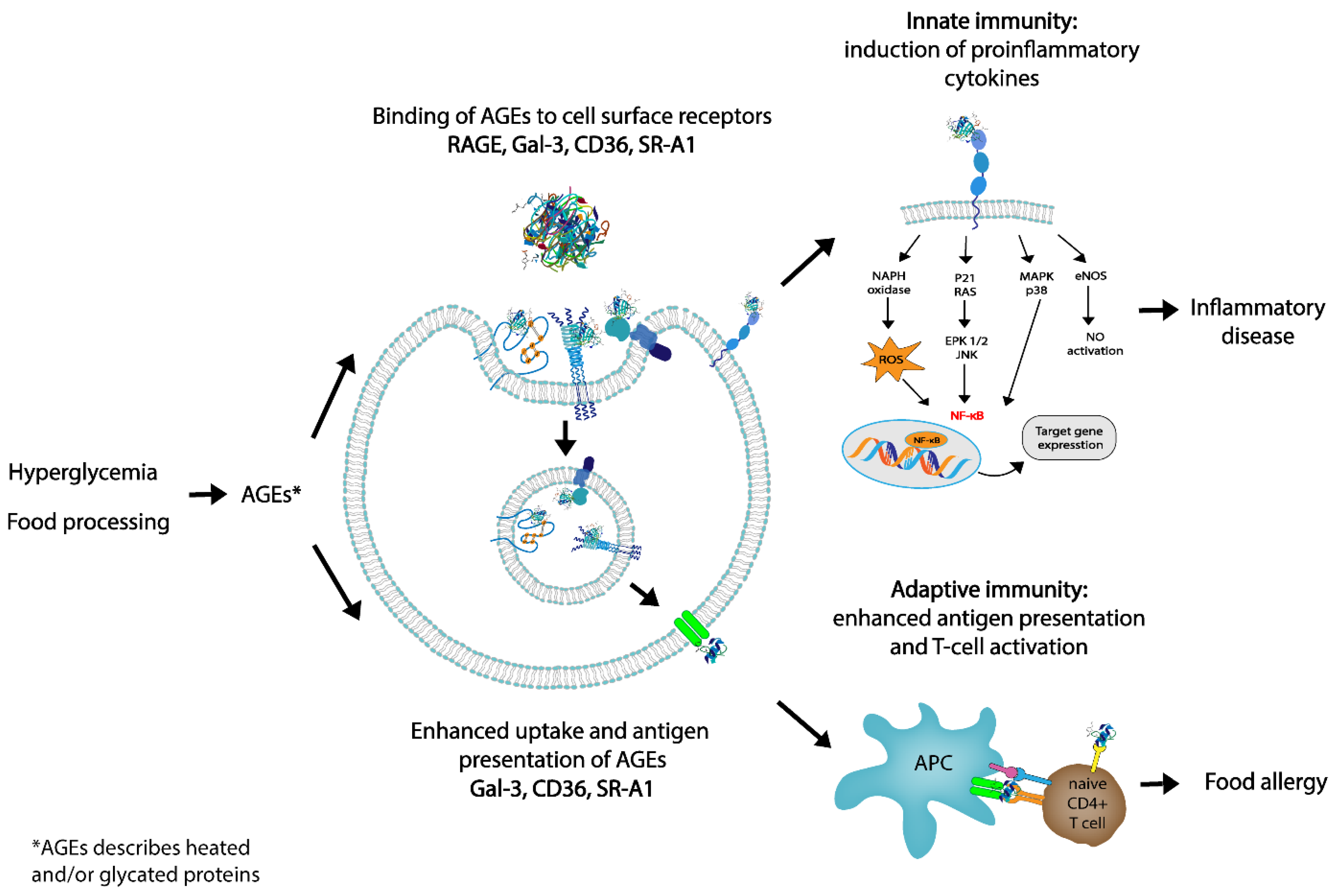
3. Interaction of AGEs with the Immune System
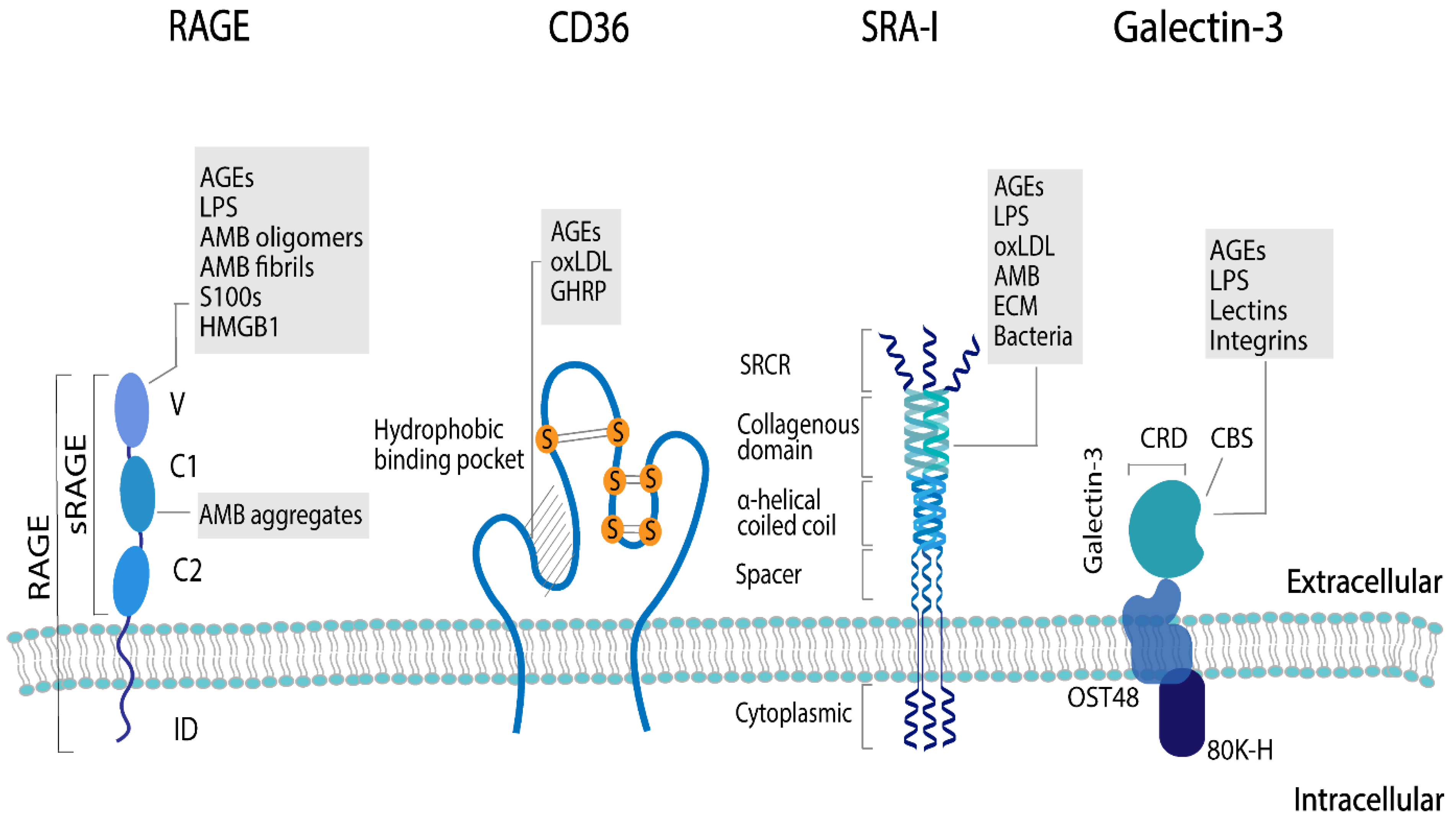
3.1. Critical Aspects of Binding of AGEs to AGE Receptors
3.2. Immunogenicity of AGEs
Influence of AGEs on the Innate and Adaptative Immune System
4. Association between dAGEs and Food Allergy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rico-Campa, A.; Martinez-Gonzalez, M.A.; Alvarez-Alvarez, I.; de Deus Mendoza, R.; de la Fuente-Arrillaga, C.; Gomez-Donoso, C.; Bes-Rastroll, M. Association between consumption of ultra-processed foods and all-cause mortality: SUN prospective cohort study. BMJ 2019, 365, 11949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.H.; Lin, X.; Bu, C.; Zhang, X. Role of advanced glycation end products in mobility and considerations in possible dietary and nutritional intervention strategies. Nutr. Metabol. 2018, 15, 72. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, Y.; Fu, L. Dietary advanced glycation end-products: Perspectives linking food processing with health implications. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2559–2587. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.I.F.S.; Jongen, W.M.F.; van Boekel, A.J.S. A review of Maillard reaction in food and implications to kinetic modelling. Trends Food Sci. Technol. 2001, 11, 364–373. [Google Scholar] [CrossRef]
- Tamanna, N.; Mahmood, N. Food Processing and Maillard Reactions Products: Effects on Human Health and Nutrition. Int. J. Food Sci. 2015, 2015, 526762. [Google Scholar] [CrossRef]
- Zhao, D.; Li, L.; Xu, D.; Sheng, B.; Chen, J.; Li, B.; Zhang, X. Heat induced amyloid-like aggregation of β-lactoglobulin regulated by glycation: A comparison of five kinds of reducing saccharides. Int. J. Biol. Macromol. 2018, 120, 302–309. [Google Scholar] [CrossRef]
- Cardoso, H.B.; Wierenga, P.A.; Gruppen, H.; Schols, H.A. Maillard induced aggregation of individual milk proteins and interactions involved. Food Chem. 2019, 276, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Vlassara, H.; Urribarri, J. Advanced glycation end products (AGE) and diabetes: Cause, effect or both? Curr. Diab. Rep. 2014, 14, 453. [Google Scholar] [CrossRef] [Green Version]
- Rungratanawanic, W.; Qu, W.; Wang, X.; Essa, M.M.; Song, B.J. Advanced glycation end products (AGEs) and other adducts in aging-related diseases and alcohol-mediated injury. Exp. Mol. Med. 2021, 53, 168–188. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Fogliano, V. Dietary Advanced Glycosylation End-Products (dAGEs) and Melanoidins Formed through the Maillard Reaction: Physiological Consequences of their Intake. Ann. Rev. Food Sci. Technol. 2018, 9, 271–291. [Google Scholar] [CrossRef]
- Angoorani, P.; Ejtahed, H.S.; Mirmiran, P.; Mirzaei, S.; Azizi, F. Dietary consumption of advanced glycation end products and risk of metabolic syndrome. Int. J. Food Sci. Nutr. 2016, 67, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Baye, E.; Kiriakova, V.; Uribarri, J.; Moran, L.J.; De Courten, B. Consumption of diets with low advanced glycation end products improves cardiometabolic parameters: Meta-analysis of randomized controlled trials. Sci. Rep. 2017, 7, 2266. [Google Scholar] [CrossRef] [PubMed]
- Di Pino, A.; Currenti, W.; Urbano, F.; Scicali, R.; Piro, S.; Purrello, F.; Rabuazzo, A.M. High intake of dietary advanced glycation end-products is associated with increased arterial stiffness and inflammation in subjects with type 2 diabetes. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Toda, M.; Heilmann, M.; Ilchmann, A.; Vieths, S. The Maillard reaction and food allergies: Is there a link? Clin. Chem. Lab. Med. 2014, 52, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Gupta, K.A.; Sharma Das, M.; Ansari, I.A.; Dwivedi, P.D. Maillard reaction in food allergy: Pros and cons. Crit. Rev. Food Sci. Nutr. 2018, 58, 208–226. [Google Scholar] [CrossRef] [PubMed]
- Teodorowicz, M.; Van Neerven, J.; Savelkoul, H. Food processing: The influence of the maillard reaction on immunogenicity and allergenicity of food proteins. Nutrients 2017, 9, 835. [Google Scholar] [CrossRef]
- Toda, M.; Hellwig, M.; Henle, T.; Vieths, S. Influence of the Maillard Reaction on the Allergenicity of Food Proteins and the Development of Allergic Inflammation. Curr. Allergy Asthma Rep. 2019, 19, 4. [Google Scholar] [CrossRef]
- Deng, Y.; Govers, C.; Bastian-Net, S.; Van der Hulst, N.; Hettinga, K.; Wichers, H.J. Hydrophobicity and aggregation, but not glycation, are the key determinants for uptake of thermally processed β-lactoglobulin by THP-1 macrophages. Food Res. Int. 2019, 120, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Zenker, H.E.; Ewaz, A.; Deng, Y.; Savelkoul, H.F.J.; Van Neerven, R.J.J.; De Jong, H.J.; Wichers, K.; Hettinga, A.; Teodorowicz, M. Differential effects of dry vs wet heating of β-lactoglobulin on formation of sRAGE binding ligands and sIgE epitope recognition. Nutrients 2019, 11, 1432. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, M.W.; Hedegaard, R.V.; Andersen, J.M.; De Courten, B.; Bugel, S.; Nielsen, J.; Skibsted, L.H.; Dragsted, L.O. Advanced glycation end-products in food and their effects on health. Food Chem. Toxicol. 2013, 60, 10–37. [Google Scholar] [CrossRef]
- Lund, M.N.; Ray, C.A. Control of Maillard reactions in foods: Strategies and chemical mechanisms. J. Agric. Food Chem. 2017, 65, 4537–4552. [Google Scholar] [CrossRef] [Green Version]
- Nowotny, K.; Schroter, D.; Schreiner, M.; Grune, T. Dietary advanced glycation end products and their relevance for human health. Ageing Res. Rev. 2018, 47, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Wijayanti, H.B.; Brodkorb, A.; Hogan, S.A.; Murphy, E.G. Chapter 6—Thermal Denaturation, Aggregation, and Methods of Prevention. Whey Proteins: From Milk to Medicine; Academic Press: Cambridge, MA, USA, 2019; pp. 185–247. [Google Scholar]
- Xiang, J.; Liu, F.; Wang, B.; Chen, L.; Liu, W.; Tan, S. A literature review on Maillard Reaction based on Milk proteins and Carbohydrates in Food and Pharmaceutical Products: Advantages, Disadvantages and Avoidance Strategies. Foods 2021, 10, 1998. [Google Scholar] [CrossRef]
- Arena, S.; Renzone, G.; D’Ambrosio, C.; Salzano, A.M.; Scaloni, A. Dairy products and the Maillard reaction: A promising future for extensive food characterization by integrated protemics studies. Food Chem. 2017, 219, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Van Lieshout, G.A.A.; Lambers, T.T.; Bragt, M.C.E.; Hettinga, K.A. How processing may affect milk protein digestion and overall physiological outcomes: A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 60, 2422–2445. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.; Mirshekar-Syahkal, B.; Kennish, L.; Karachalias, N.; Babaei-Jadidi, R.; Thornalley, P.J. Assay of advanced glycation end-products in selected beverages and food by liquid chromatography with tandem mass spectrometric detection. Mol. Nutr. Food Res. 2005, 49, 691–699. [Google Scholar] [CrossRef]
- Ruiz, H.H.; Ramasamy, R.; Schimdt, A.M. Advanced Glycation End Products: Building on the Concept of the “Common Soil” in Metabolic Disease. Endocrinology 2020, 161, bqz006. [Google Scholar] [CrossRef]
- Yan, S.F.; Ramasamy, R.; Schmidt, A.M. Mechanisms of disease: Advanced glycation end-products and their receptor in inflammation and diabetes complications. Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Reynaert, N.L.; Gopal, P.; Rutten, E.P.A.; Wouters, E.F.M.; Schalkwijk, C.G. Advanced glycation end products and their receptor in age-related, non-communicable chronic inflammatory diseases; Overview of clinical evidence and potential contributions to disease. Int. J. Biochem. Cell Biol. 2016, 81, 403–418. [Google Scholar] [CrossRef]
- Wetzels, S.; Wouters, K.; Schalkwijk, C.G.; Vanmierlo, T.; Hendricks, J.J.A. Methylglyoxal-Derived Advanced Glycation Endproducts in Multiple Sclerosis. Int. J. Mol. Sci. 2017, 18, 421. [Google Scholar] [CrossRef] [Green Version]
- Kellow, N.J.; Coughlan, M.T. Effect of diet-derived advanced glycation end products on inflammation. Nutr. Rev. 2015, 73, 737–759. [Google Scholar] [CrossRef] [PubMed]
- Snelson, M.; Coughlan, M.T. Dietary Advanced Glycation End Products: Digestion, Metabolism and Modulation of Gut Microbial Ecology. Nutrients 2019, 22, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henle, T. AGEs in food: Do they play a role in uremia? Kid Int. Suppl. 2003, 63, S145–S147. [Google Scholar] [CrossRef] [Green Version]
- Foster, D.; Spruill, L.; Walter, K.R.; Noguiera, L.M.; Fedarovich, H.; Turner, R.Y.; Ahmed, M.; Sadley, J.D.; Ford, M.E.; Findlay, V.J.; et al. AGE metabolites: A biomarker linked to cancer disparity? Cancer Epidemiol. Prev. Biomark. 2014, 23, 2186–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, R.; Mo, F.; Chang, B.; Zhang, Q.; Ran, H.; Yang, S.; Zhu, Z.; Hu, L.; Su, Q. Glucose-derived AGEs enhance human gastric cancer metastasis through RAGE/ERK/Spl/MMP2 cascade. Oncotarget 2017, 8, 104216–104226. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Wang, F.; Wang, J.; Liu, C.; Zhou, Y.; Xu, Z.; Zhang, C.; Sun, B.; Guan, Y. Pathological Mechanisms Linking Diabetes Mellitus and Alzheimer’s Disease: The receptor for Advanced Glycation End Products (RAGE). Front. Aging Neurosci. 2020, 22, 217. [Google Scholar] [CrossRef]
- Prasad, K.; Dhar, I.; Caspar-Bell, G. Role of Advanced Glycation End Products and Its Receptors in the Pathogenesis of Cigarette Smoke-Induced Cardiovascular Disease. Int. J. Angiol. 2015, 24, 75–80. [Google Scholar]
- Koschinsky, T.; He, C.J.; Mitsuhashi, T.; Bucala, R.; Liu, C.; Bueting, C.; Heitmann, K.; Vlassara, H. Orally absorbed reactive glycation products (glycotoxins): An environmental risk factor in diabetic nephropathy. Proc. Natl. Acad. Sci. USA 1997, 94, 6474–6479. [Google Scholar] [CrossRef] [Green Version]
- Forster, A.; Kuhne, Y.; Henle, T. Studies on absorption and elimination of dietary maillard reaction products. Ann. N. Y. Acad. Sci. 2005, 1043, 474–481. [Google Scholar] [CrossRef]
- Garay-Sevilla, M.E.; Rojas, A.; Portero-Otin, M.; Uribarri, J. Dietary AGEs as Exogenous Boosters on Inflammation. Nutrients 2021, 13, 2802. [Google Scholar] [CrossRef]
- Baig, M.H.; Jan, A.T.; Rabbani, G.; Ahmad, K.; Ashraf, J.M.; Kim, T.; Min, H.S.; Lee, Y.H.; Cho, W.K.; Ma, J.Y.; et al. Methylglyoxal and Advanced Glycation End products: Insight of the regulatory machinery affecting the myogenic program and of its modulation by natural compounds. Sci. Rep. 2017, 7, 5916. [Google Scholar] [CrossRef] [PubMed]
- Tessier, F.J.; Boulanger, E.; Howsam, M. Metabolic transit of dietary advanced glycation end-products: The case of Nε-carboxymethyl-lysine. Glycoconj. J. 2021, 38, 311–317. [Google Scholar] [CrossRef]
- Tessier, F.J.; Leridon, C.N.; Jacolot, P.; Jouquand, C.; Genin, M.; Schmidt, A.M.; Grossin, N.; Boulanger, E. Quantitative assessment of organ distribution of dietary protein-bound 13C-labeled Ne-carboxymethyllysine after a chronic oral exposure in mice. Mol. Nutr. Food Res. 2016, 60, 2446–2456. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.; Humpf, H.U.; Hengstler, J.; Mally, A.; Vieths, S.; Henle, T. Dietary glycation compounds- implications for human health? In Proceedings of the DFG-SKLM, Round Table Expert Meeting, Dortmund, Germany, 14 November 2019. [Google Scholar]
- Heilmann, M.; Wellner, A.; Gadermaier, G.; Ilchmann, A.; Briza, P.; Krause, M.; Nagai, R.; Burgdorf, S.; Scheurer, S.; Henle, T.; et al. Ovalbumin modified with pyrraline, a maillard reaction product shows enhanced T-cell immunogenicity. J. Biol. Chem. 2014, 289, 7919–7928. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yu, S.J. Review of pentosidine and pyrraline in food and chemical models: Formation, potential risk and determination. J. Sci. Food Agric. 2018, 98, 3225–3233. [Google Scholar] [CrossRef]
- Forster, A.; Kuhne, Y.; Henle, T. Dietary Intake and Urinary Excretion of Maillard Reaction Products (MRPs). Czec. J. Food Sci. 2004, 22, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Uribarri, J.; Stirban, A.; Sander, D.; Cai, W.; Negrean, M.; Buenting, C.E.; Koschinsky, T.; Vlassara, H. Single Oral Challenge by Advanced Glycation End Products Acutely Impairs Endothelial Function in Diabetic and Non-diabetic subjects. Diab. Care 2007, 30, 2579–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheijen, J.L.J.M.; Hanssen, N.M.J.; van Greevenbroek, M.M.; van der Kallen, C.J.; Feskens, E.J.; Stehouwer, C.D.A.; Schalkwijk, C.G. Dietary intake of advanced glycation end products in plasma and urine: The CODAM study. Clin. Nutr. 2018, 37, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.; Gensberger-Reigl, S.; Henle, T.; Pischetsrieder, M. Food-derived 1,2-dicarbonyl compounds and their role in diseases. Sem. Cancer Biol. 2018, 49, 1–8. [Google Scholar] [CrossRef]
- Uribarri, J.; del Castillo, M.D.; de la Maza, M.P.; Filip, R.; Gucliucci, A.; Luevano-Contreras, C.; Macias-Cervantes, M.H.; Bastos, D.H.M.; Mendrano, A.; Menini, T.; et al. Dietary Advanced Glycation end products and their Role in Health and Disease. Adv. Nutr. 2015, 6, 461–473. [Google Scholar] [CrossRef]
- Teodorowicz, M.; Hendriks, W.H.; Wichers, H.J.; Savelkoul, H.F.J. Immunomodulation by Processed Animal Feed: The role of Maillard Reaction Products and Advanced Glycation End-Products (AGEs). Front. Immunol. 2018, 9, 2088. [Google Scholar] [CrossRef] [Green Version]
- Marcowicz Bastos, D.H.; Gugliucci, A. Contemporary and controversial aspects of the Maillard reaction products. Curr. Opin. Food Sci. 2015, 1, 13–20. [Google Scholar] [CrossRef]
- Ott, C.; Jacobs, K.; Haucke, E.; Navarrete Santos, A.; Grune, T.; Simm, A. Role of advanced glycation end products in cellular signaling. Redox Biol. 2014, 2, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.M.; Hoffmann, M.; Tauchi, A.; Du, S.; Stern, D.M. RAGE: A multiligand receptor contributing to the cellular response in diabetic vasculopathy and inflammation. Sem. Thromb. Hemost. 2000, 26, 485–493. [Google Scholar] [CrossRef]
- Vlassara, H.; Li, Y.M.; Imani, F.; Wojciechowicz, D.; Yang, Z.; Liu, F.T.; Cerami, A. Identification of galectin-3 as a high-affinity binding protein for advanced glycation end products (AGE): A new member of the AGE-receptor complex. Mol. Med. 1995, 1, 634–646. [Google Scholar] [CrossRef] [Green Version]
- Pricci, F.; Leto, G.; Amadio, L.; Lacobini, C.; Romeo, G.; Cordone, S.; Gradini, R.; Barsotti, P.; Liu, F.T.; Mario, U.D.; et al. Role of galectin-3 as a receptor for advanced glycosylation end products. Kidney Int. 2000, 58, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Kurihara, Y.; Takeya, M.; Kamada, N.; Kataoka, M.; Jishage, K.; Ueda, O.; Sakaguchi, H.; Higashi, T.; Susuki, T.; et al. A role for macrophage scavenger receptors in atherosclerosis and susceptibility to infection. Nature 1997, 386, 292–296. [Google Scholar] [CrossRef]
- Ohgami, N.; Nagai, R.; Miyazaki, A.; Ikemoto, M.; Arai, H.; Horiuchi, S.; Nakayama, H. Scavenger Receptor Class B Type I-mediated Reverse Cholesterol Transport Is Inhibited by Advanced Glycation End Products. J. Biol. Chem. 2001, 276, 3348–3355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohgami, N.; Nagai, R.; Ikemoto, M.; Arai, H.; Kuniyasu, A.; Horiuchi, S.; Nakayama, H. CD36, a Member of the Class B Scavenger Receptor Family, as a Receptor for Advanced Glycation End Products. J. Biol. Chem. 2001, 276, 3195–3202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Zoelen, M.A.D.; Yang, H.; Florquin, S.; Meijers, J.C.; Akira, S.; Arnold, B.; Naworth, P.P.; Bierhaus, A.; Tracey, K.J.; van der Poll, T. Role of toll-like receptors 2 and 4, and the receptor for advanced glycation end products in high-mobility group box 1-induced inflammation in vivo. Shock 2009, 31, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Park, J.H. Receptor for Advanced Glycation End-products (RAGE), Its Ligands, and Soluble RAGE: Potential Biomarkers for Diagnosis and Therapeutic Targets for Human Renal Diseases. Genom. Inform. 2013, 11, 224–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.M.; Yan, S.D.; Yan, S.F.; Stern, D.M. The multiligand receptor RAGE as a progression factor amplifying immune and inflammatory responses. J. Clin. Investig. 2001, 108, 949–955. [Google Scholar] [CrossRef]
- Neyen, C.; Pluddemann, A.; Rovers, P.; Thomas, B.; Cai, L.; van der Westhuyzen, D.R.; Sim, R.B.; Gordon, S. Macrophage Scavenger Receptor a Mediates Adhesion to Apolipoproteins A-I and E. Biochemistry 2009, 48, 11858–11871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Govers, C.; Teodorowicz, M.; Liobyte, I.; de Simone, I.; Hettings, K.A.; Wichers, H.J. Hydrophobicity drives receptor-mediated uptake of heat-processed proteins by THP-1 macrophages and dendritic cells, but not cytokine responses. PLoS ONE 2020, 15, e0236212. [Google Scholar] [CrossRef] [PubMed]
- Platt, N.; Haworth, R.; Darley, L.; Gordon, S. The many roles of the Class a Macrophage Scavenger Receptor. Int. Rev. Cytol. 2002, 212, 1e–42e. [Google Scholar]
- Hilmenyuk, T.; Bellinghausen, I.; Heydenreich, B.; Ilchmann, A.; Toda, M.; Grabbe, S.; Saloga, J. Effects of glycation of the model food allergen ovalbumin on antigen uptake and presentation by human dendritic cells. Immunology 2010, 129, 437–445. [Google Scholar] [CrossRef]
- Van der Lugt, T.; Weseler, A.R.; Gebbink, W.A.; Vrolijk, M.F.; Opperhuizen, A.; Bast, A. Dietary advanced glycation end-products induce an inflammatory response in human macrophages in vitro. Nutrients 2018, 10, 1868. [Google Scholar] [CrossRef] [Green Version]
- Zenker, H.E.T.; Ewaz, A.; van Nerveen, R.J.J.; Savelkoul, H.F.J.; De Jong, N.W.; Wichers, H.J.; Hettings, K.A. Binding of CML-Modified as Well as Heat-Glycated β-lactoglobulin to Receptors for AGEs is Determined by Charge and Hydrophobicity. Int. J. Mol. Sci. 2020, 21, 4567. [Google Scholar] [CrossRef]
- Xue, J.; Rai, V.; Singer, D.; Chabierski, S.; Xie, J.; Reverdatto, S.; Burz, D.S.; Schmidt, A.M.; Hoffmann, R.; Shekhtman, A. Advanced glycation end product recognition by the receptor for AGEs. Structure 2011, 19, 722–732. [Google Scholar] [CrossRef] [Green Version]
- Teodorowicz, M.; Zenker, H.E.T.; Ewaz, A.; Tsallis, T.; Mauser, A.; Gensberger-Reigl, S.; De Jong, N.W.; Hettings, K.A.; Wichers, H.J.; van Nerveen, R.J.J.; et al. Enhanced Uptake of Processed Bovine β-lactoglobulin by Antigen Presenting Cells: Identification of Receptors and Implications for Allergenicity. Mol. Nutr. Food Res. 2021, 65, 2000834. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Teodorowicz, M.; Wichers, H.J.; Van Boekel, M.A.J.S.; Hettinga, K.A. Generation of Soluble Advanced Glycation End Products Receptor (sRAGE)-Binding Ligands during Extensive Heat Treatment of Whey Protein/Lactose Mixtures Is Dependent on Glycation and Aggregation. J. Agric. Food Chem. 2016, 64, 6477–6486. [Google Scholar] [CrossRef] [PubMed]
- Malec, L.S.; Pereyra Gonzales, A.S.; Naranjo, G.B.; Vigo, M.S. Influence of water activity and storage temperature on lysine availability of a milk like system. Food Res. Int. 2002, 35, 849–853. [Google Scholar] [CrossRef]
- Pereyra Gonzales, A.S.; Naranjo, G.B.; Leiva, G.E.; Malec, L.S. Maillard reaction kinetics in milk powder: Effect of water activity at mild temperatures. Int. Dairy J. 2010, 20, 40–45. [Google Scholar] [CrossRef]
- Naranjo, G.B.; Pereyra Gonzales, A.S.; Leiva, G.E.; Malec, L.S. The kinetics of Maillard reaction in lactose-hydrolysed milk powder and related systems containing carbohydrate mixtures. Food Chem. 2013, 141, 3790–3795. [Google Scholar] [CrossRef]
- Pischetsrieder, M.; Henle, T. Glycation products in infant formulas: Chemical, analytical and physiological aspects. Amino Acids 2012, 42, 1111–1118. [Google Scholar] [CrossRef]
- Kislinger, T.; Fu, C.; Huber, B.; Qu, W.; Taguchi, A.; Yan, S.D.; Hofmann, M.; Yan, S.F.; Pischetsrieder, M.; Stern, D.; et al. N(ε)-(carboxymethyl)lysine adducts of proteins are ligands for receptor for advanced glycation end products that activate cell signaling pathways and modulate gene expression. J. Biol. Chem. 1999, 274, 31740–31749. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Ray, R.; Singer, D.; Bohme, D.; Burz, D.S.; Rai, V.; Hoffmann, R.; Shekthtman, A. The receptor for advanced glycation end products (RAGE) specifically recognizes methylglyoxal-derived AGEs. Biochemistry 2014, 53, 3327–3335. [Google Scholar] [CrossRef] [PubMed]
- Nogueira Silva Lima, M.T.; Howsam, M.; Anton, P.M.; Delayre-Orthez, C.; Tessier, F.J. Effect of Advanced Glycation End-Products and Excessive Calorie Intake on Diet-Induced Chronic Low-Grade Inflammation Biomarkers in Murine Models. Nutrients 2021, 13, 3091. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, X.M.; Xiao, H.; Nowak-Wegrzyn, A.; Zhou, P. Insight into the allergenicity of shrimp trompyosin glycated by functional oligosaccharides containing advanced glycation end products. Food Chem. 2020, 302, 125348. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K. Do advanced glycation end-products cause food allergy? Curr. Opin. Allergy Clin. Immunol. 2017, 17, 325–331. [Google Scholar] [CrossRef]
- Reed, J.C.; Preston-Hulburt, P.; Philbrick, W.; Betancur, G.; Korah, M.; Lucas, C.; Herold, K.C. The receptor for advanced glycation end products (RAGE) modulates T cell signaling. PLoS ONE 2020, 15, e0236921. [Google Scholar] [CrossRef] [PubMed]
- Oczypok, E.A.; Milutinovic, P.S.; Alcorn, J.F.; Khare, A.; Crum, L.T.; Manini, M.L.; Epperly, M.W.; Pawluk, A.M.; Ray, A.; Oury, T.D. Pulmonary receptor for advanced glycation end-products promotes pathogenesis through IL-33 and accumulation of group 2 innate lymphoid cells. J. Allergy Clin. Immunol. 2015, 136, 747–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oczypok, E.A.; Perkins, T.N.; Oury, T.D. All the “RAGE” in lung disease: The receptor for advanced glycation endproducts (RAGE) is a major mediator of pulmonary inflammatory responses. Paediatr. Respir. Rev. 2017, 23, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Ichmann, A.; Burgdorf, S.; Scheurer, S.; Waibler, Z.; Nagai, Z.; Wellner, A.; Yamamoto, Y.; Yamamato, H.; Henle, T.; Kurts, C.; et al. Glycation of a food allergen by the Maillard reaction enhances its T-cell immunogenicity: Role of macrophage scavenger receptor class A type I and II. J. Allergy Clin. Immunol. 2010, 125, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.E.; Hillson, W.R.; Noti, M.; Gartlan, K.H.; Johnson, S.; Thomas, B.; Artis, D.; Sattentau, Q.J. Dry roasting enhances peanut-induced allergic sensitization across mucosal and cutaneous routes in mice. J. Allergy Clin. Immunol. 2014, 134, 1453–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbezier, N.; Tessier, F.J.; Chango, A. Receptor of advanced glycation end-products RAGE/AGE: An integrative view for clinical applications. Ann. Biol. Clin. 2014, 72, 669–680. [Google Scholar]
- Gaens, K.H.; Stehouwer, C.D.; Schalkwijk, C.G. Advanced glycation end-products and its receptor for advanced glycation end-products in obesity. Curr. Opin. Lipidol. 2013, 24, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Van Beijnum, J.R.; Buurman, W.A.; Griffioen, A.W. Convergence and amplification of toll-like receptor (TLR) and receptor for advanced glycation end products (RAGE) signaling pathways via high mobility group B1 (HMGB1). Angiogenesis 2008, 11, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.; Figueroa, H.; Morales, E. Fueling inflammation at tumor microenvironment: The role of multiligand/rage axis. Carcinogenesis 2010, 31, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Toyomura, T.; Wake, H.; Liu, K.; Teshigawara, K.; Takahashi, H.; Nishibori, M.; Mori, S. Differential contribution of possible pattern-recognition receptors to advanced glycation end product–induced cellular responses in macrophage-like RAW264.7 cells. Biotechnol. Appl. Biochem. 2020, 67, 265–272. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, Y.; Ciu, Q.; Xu, J.; Tang, Z.; Wang, Y.; He, C.; Wang, X. Toll-like receptor 4 plays a key role in advanced glycation end products induced M1 macrophage polarization. Biochem. Biophys. Res. Comm. 2020, 531, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. Macrophage Polarization. Ann. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef]
- Thomas, D.; Apovian, C. Macrophage functions in lean and obese adipose tissue. Metabolism 2017, 72, 120–143. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Hu, Q.; Xu, X.; Niu, Y.; Chen, Y.; Lu, Y.; Su, Q.; Qin, L. Advanced glycation end products enhance M1 macrophage polarization by activating the MAPK pathway. Biochem. Biophys. Res. Comm. 2020, 525, 334–340. [Google Scholar] [CrossRef]
- Han, X.; Ma, W.; Zhu, Y.; Sun, X.; Liu, N. Advanced glycation end products enhance macrophage polarization to the M1 phenotype via the HIF-1α/PDK4 pathway. Mol. Cell Endocr. 2020, 514, 110878. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Y.R.; Li, P.C.; Feng, B. Advanced glycation end products increase lipids accumulation in macrophages through upregulation of receptor of advanced glycation end products: Increasing uptake, esterification and decreasing efflux of cholesterol. Lipids Health Dis. 2016, 15, 161. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, S.; Dai, J.; Singh, S.; Mwangi, D.; Rivera, A.; Fitzgerald-Bocarsly, P. Human pDCs express the C-type Lectin receptor Dectin-1 and uptake and kill Aspergillus fumigatus spores in vitro (MPF4P.734). J. Immunol. 2015, 194, 136. [Google Scholar]
- Ge, J.; Jia, Q.; Liang, C.; Luo, Y.; Huang, D.; Sun, A.; Wang, K.; Zou, Y.; Chen, H. Advanced glycosylation end products might promote atherosclerosis through inducing the immune maturation of dendritic cells. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2157–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, C.L.; Sharp, P.S.; North, M.E.; Rainbow, S.J.; Knight, S.C. Advanced glycation end products modulate the maturation and function of peripheral blood dendritic cells. Diabetes 2004, 53, 1452–1458. [Google Scholar] [CrossRef] [Green Version]
- Perusko, M.; van Roest, M.; Stanic-Vucinic, D.; Simons, P.J.; Pieters, R.H.H.; Velickovic, T.C.; Smit, J.J. Glycation of the Major Milk Allergen β-Lactoglobulin Changes Its Allergenicity by Alterations in Cellular Uptake and Degradation. Mol. Nutr. Food Res. 2018, 62, e1800341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, M.; Jurets, A.; Wallner, M.; Briza, P.; Ruzek, S.; Hainzl, S.; Pichler, U.; Kitzmüller, C.; Bohle, B.; Huber, C.G.; et al. Assessing protein immunogenicity with a dendritic cell line-derived endolysosomal degradome. PLoS ONE 2011, 6, e17278. [Google Scholar] [CrossRef] [Green Version]
- Imran, S.; Neeland, M.R.; Shepherd, R.; Messina, N.; Perrett, K.P.; Netea, M.G.; Curtis, N.; Saffery, R.; Novakovic, B. A potential role for Epigenetically Mediated Trained Immunity in Food Allergy. iScience 2020, 23, 101171. [Google Scholar] [CrossRef] [PubMed]
- Van Esch, B.C.M.; Porbahaie, M.; Abbring, S.; Garssen, J.; Potaczek, D.P.; Savelkoul, H.F.J.; Neerven, R.J.J. The Impact of Milk and its Components on Epigenetic Programing of Immune Function in Early Life and Beyond: Implications for Allergy and Asthma. Front. Immunol. 2020, 11, 2141. [Google Scholar] [CrossRef] [PubMed]
- Mueller, G.A.; Maleki, S.J.; Johnson, K.; Hulburt, B.K.; Cheng, H.; Ruan, S.; Nesbit, J.B.; Pomes, A.; Edwards, L.L.; Schorzman, A.; et al. Identification of Maillard reaction products on peanut allergens that influence binding to the receptor for advanced glycation end products. Allergy 2013, 68, 1546–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teodorowicz, M.; Jansen, A.P.H.; Roovers, M.H.W.M.; Ruinemans-Koerts, J.; Wichers, H.J.; Savelkoul, H.F.J. Maillard-type neoallergens present in processed soy extract may cause an allergic reaction in soy allergic patients. Clin. Transl. Allergy 2015, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.K.; Masilamani, M.; Li, X.M.; Sampson, H.A. The false alarm hypothesis: Food allergy is associated with high dietary advanced glycation end-products and proglycating dietary sugars that mimic alarmins. J. Allergy Clin. Immunol. 2017, 139, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Rider, P.; Vornov, E.; Dinarello, C.A.; Apte, R.N.; Cohen, I. Alarmins: Feel the Stress. J. Immunol. 2017, 198, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- Del Castillo, M.D.; Iriondo-DeHond, A.; Iriondo-DeHond, M.; Gonzalez, I.; Medrano, A.; Filip, R.; Uribarri, J. Healthy eating recommendations: Good reducing dietary contribution to the body’s advanced glycation/lipoxidation end products pool? Nutr. Res. Rev. 2021, 34, 48–63. [Google Scholar] [CrossRef]
- Li, Z.; Luo, Y.; Feng, L.; Liao, P. Effect of Maillard reaction conditions on antigenicity of B-lactoglobulin and the properties of glycated whey protein during simulated gastric digestion. Food Agric. Immunol. 2013, 24, 433–443. [Google Scholar] [CrossRef]
- Uribarri, J.; Woodruff, S.; Goodmann, S.; Weijing, C.A.I.; Chen, X.; Pyzik, R.; Yong, A.; Striker, G.E.; Vlassara, H. Advanced Glycation End Products in Foods and a Practical Guide to their Reduction in the Diet. J. Am. Diet. Assoc. 2010, 110, 911–916.e12. [Google Scholar] [CrossRef] [Green Version]
- Tessier, T.; Boulanger, E. The receptor for advanced glycation end-products (RAGE) is an important pattern recognition receptor (PRR) for inflammaging. Biogerontology 2019, 20, 279–301. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
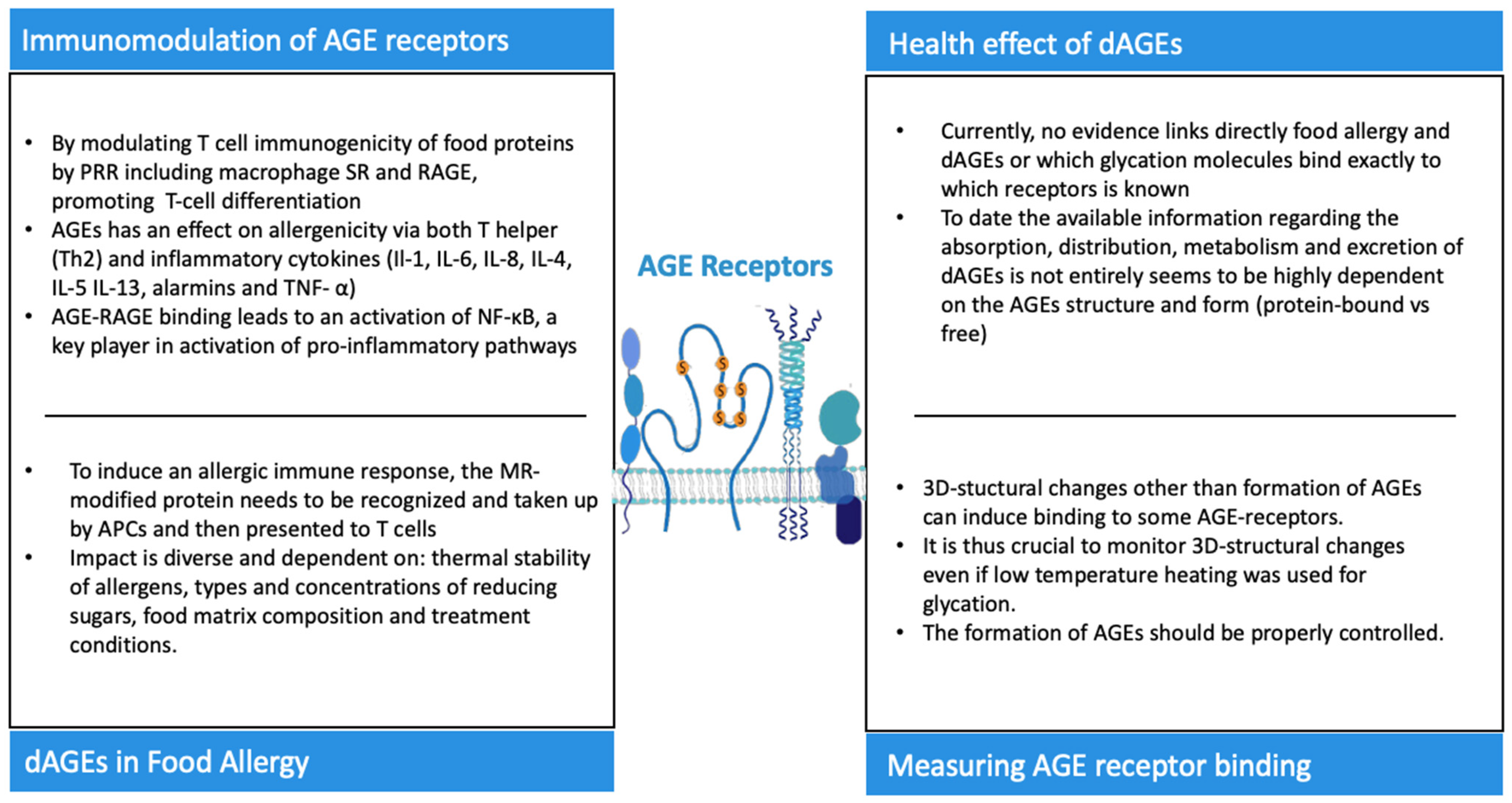{kind=link}
| Receptor | Structure | AGE Binding Sides | Forces for AGE Interaction |
|---|---|---|---|
| RAGE | EC: one V-type domain, two C-types domains and a short transmembrane domain | V-type domain | Electrostatic |
| IC: cytoplasmic tail | |||
| Galectin-3 | Component of AGE-R complex has a carbohydrate recognition domain (CRD) and a carbohydrate binding side (CBS) | CBS | Hydrophilic interactions via hydrogen bonds, and hydrophobic interactions, specifically the CH-π interaction explains binding to lectins and lipopolysaccharides. For specific AGEs unknown. |
| SR-A | EC: scavenger receptor cysteine-rich structure (SRCR), collagenous domain, α-helical coiled coil, and spacer as well as an intracellular cytoplasmic | Collagenous domain | For specific AGEs unknown.All ligands are macromolecular and polyanionic. For apo-A and apo- E amphipathic α-helix suggested as a potential recognition motif. Dual cation-binding site proposed as main domain for ligand binding via SR-A, hence electrostatic interactions. |
| CD36 | Two transmembrane domains, an EC loop with glycosylation sites and two short IC tails | Hydrophobic binding pocket located at the highly glycosylated sites | Electrostatic, via a positively charged moiety that binds to negatively charged ligands, based on studies with diacylglycerol and oxLDL as ligands. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briceno Noriega, D.; Zenker, H.E.; Croes, C.-A.; Ewaz, A.; Ruinemans-Koerts, J.; Savelkoul, H.F.J.; van Neerven, R.J.J.; Teodorowicz, M. Receptor Mediated Effects of Advanced Glycation End Products (AGEs) on Innate and Adaptative Immunity: Relevance for Food Allergy. Nutrients 2022, 14, 371. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14020371
Briceno Noriega D, Zenker HE, Croes C-A, Ewaz A, Ruinemans-Koerts J, Savelkoul HFJ, van Neerven RJJ, Teodorowicz M. Receptor Mediated Effects of Advanced Glycation End Products (AGEs) on Innate and Adaptative Immunity: Relevance for Food Allergy. Nutrients. 2022; 14(2):371. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14020371
Chicago/Turabian StyleBriceno Noriega, Daniela, Hannah E. Zenker, Cresci-Anne Croes, Arifa Ewaz, Janneke Ruinemans-Koerts, Huub F. J. Savelkoul, R. J. Joost van Neerven, and Malgorzata Teodorowicz. 2022. "Receptor Mediated Effects of Advanced Glycation End Products (AGEs) on Innate and Adaptative Immunity: Relevance for Food Allergy" Nutrients 14, no. 2: 371. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14020371