Variation in Fatty Acid Distribution of Different Acyl Lipids in Rice (Oryza sativa L.) Brans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Rice Seeds
2.2. Reagents and Standards
2.3. Extraction of Lipids
2.4. Lipid Analysis
2.5. Enzymatic Hydrolysis of Lipids
2.6. Statistical Analyses
3. Results and Discussion
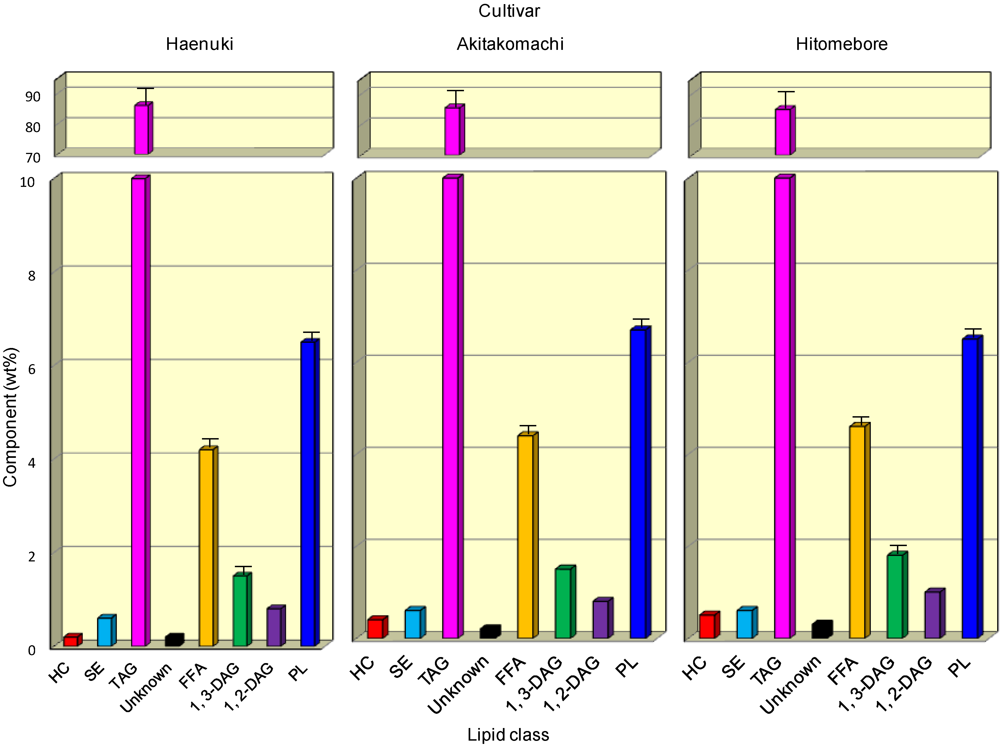
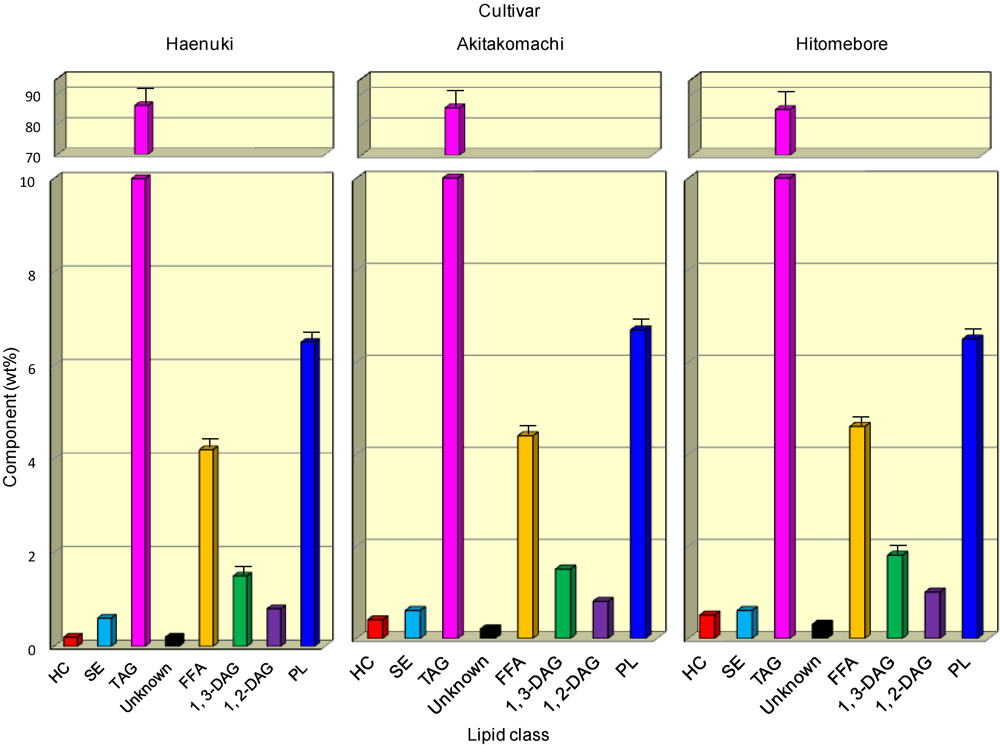
3.1. Lipid Components in the Rice Brans
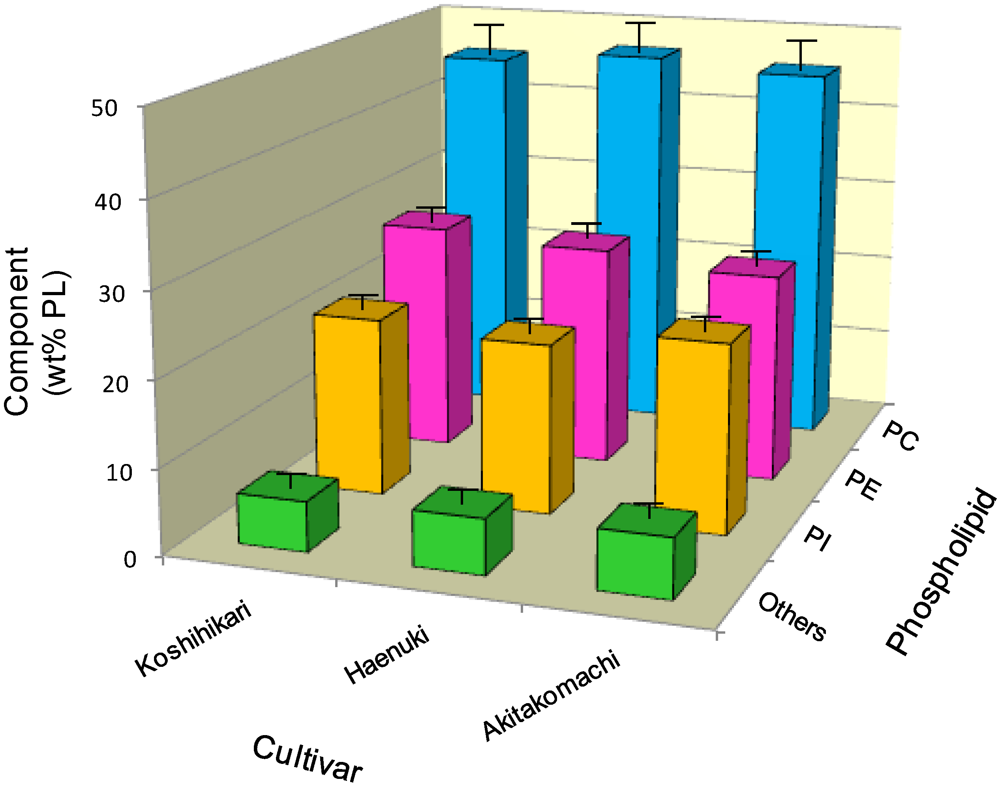
3.2. Lipid Components of Major Phospholipids in the Rice Brans

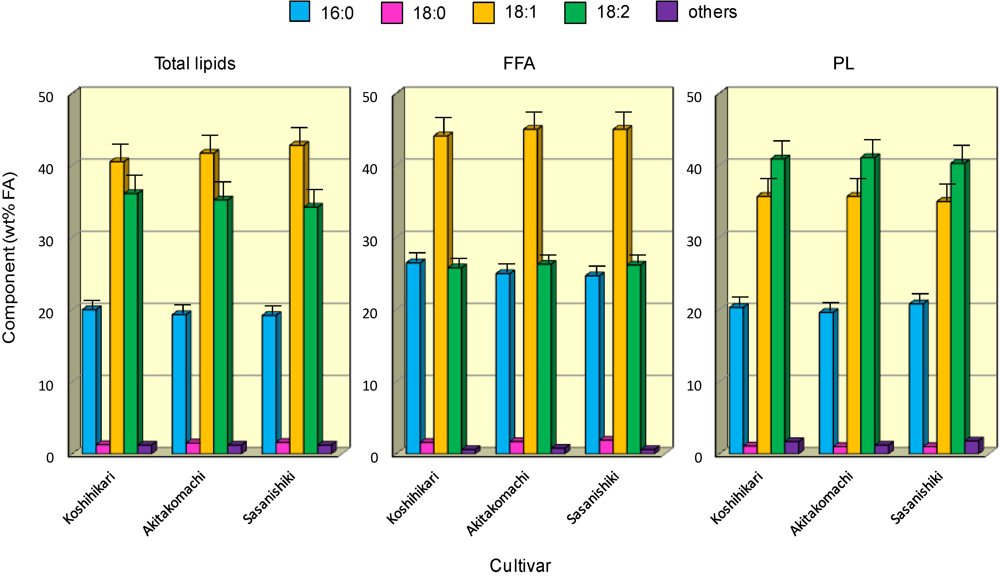
3.3. FA Composition of Major Lipids in the Rice Brans

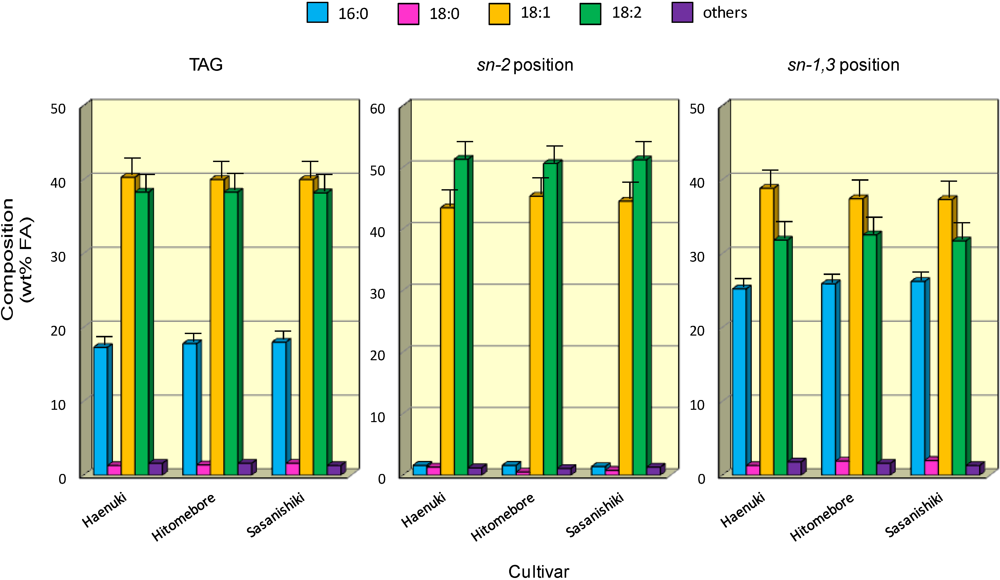
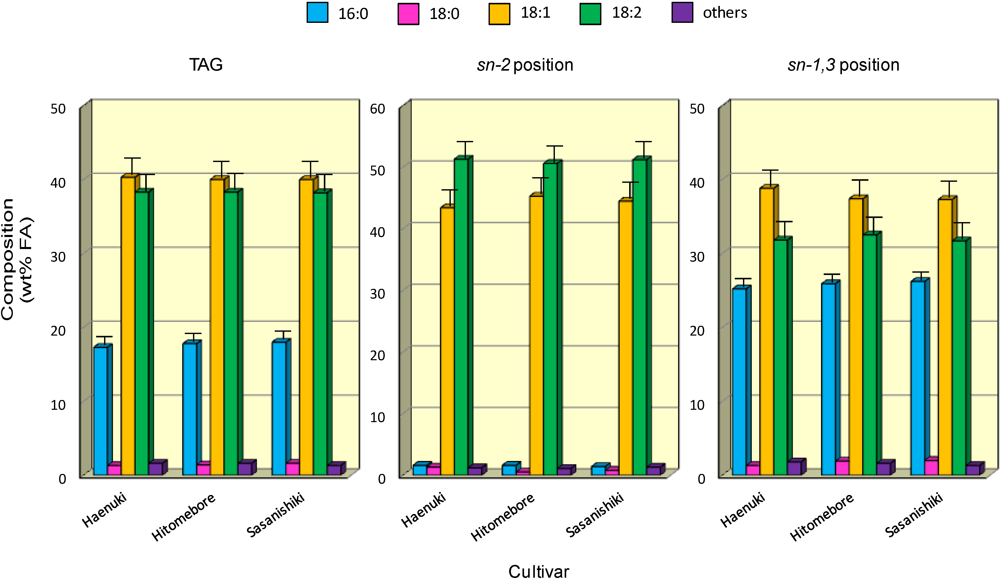
3.4. FA Positional Distribution of TAG in the Rice Brans
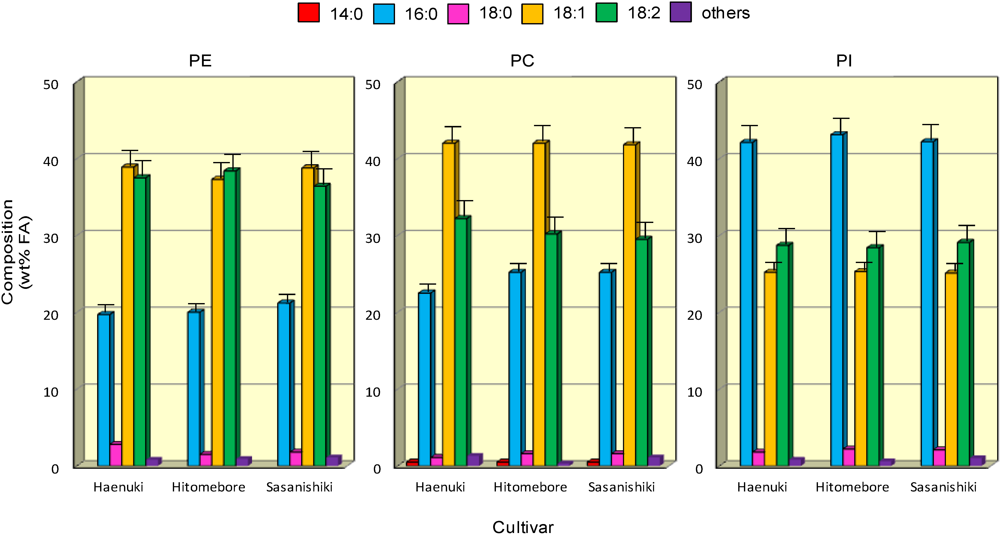
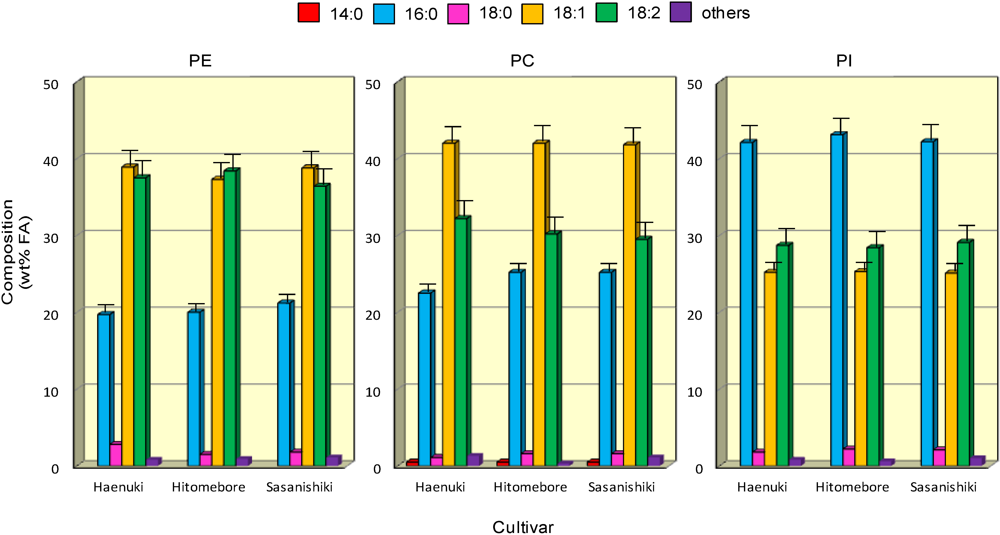
3.5. FA Compositions of Major Phospholipids in the Rice Brans
4. Conclusions
Acknowledgements
References
- Xue, Y.; Zhang, S.Q.; Yao, Q.H.; Peng, R.H.; Xiong, A.S.; Li, X.; Zhu, W.M.; Zhu, Y.Y.; Zha, D.S. Identification of quantitative trait loci for seed storability in rice (Oryza sativa L.). Euphytica 2008, 164, 739–744. [Google Scholar] [CrossRef]
- Zhou, Z.; Robards, K.; Helliwell, S.; Blanchand, C. Composition and functional properties of rice. Int. J. Food Sci. Technol. 2002, 37, 849–868. [Google Scholar]
- Siebenmorgen, T.; Matsler, A.L.; Earp, C.F. Milling characteristics of rice cultivars and hybrids. Cereal Chem. 2006, 83, 169–172. [Google Scholar]
- Juliano, B.C. Lipids in Rice and Rice Processing. In Lipids in Cereal Technology; Barenes, P.J., Ed.; Academic Press: New York, NY, USA, 1983; pp. 305–330. [Google Scholar]
- Tester, R.F.; Morrison, W.R. Swelling and gelatinization of cereal starches. I. Effects of amylopectin, amylpse, and lipids. Cereal Chem. 1990, 67, 551–557. [Google Scholar]
- Yoshino, G.; Kazumi, T.; Amano, M.; Tateiwa, M.; Yamasaki, T.; Takashima, S.; Iwai, M.; Hatanaka, H.; Baba, S. Effects of gamma-oryzanol on hyperlipidemic subjects. Curr. Ther. Res. Clin. Exp. 1989, 45, 543–552. [Google Scholar]
- Jenning, B.H.; Akoh, C.A. Effectiveness of natural versus synthetic antioxidants in a rice bran oil-based structured lipid. Food Chem. 2009, 114, 1456–1461. [Google Scholar]
- Wilson, T.A.; Nicolosi, R.J.; Woolfrey, B.; Kritchevsky, D. Rice bran oil and oryzanol reduce plasma lipid and lipoprotein cholesterol concentrations and aortic cholesterol ester accumulation to greater extent than ferulic acid in hypercholesterolemic hamsters. Nutr. Biochem. 2007, 18, 105–112. [Google Scholar]
- Zhou, Z.; Robards, K.; Helliwell, S.; Blanchard, C. Ageing of stored rice: Changes in chemical and physical attributes. J. Cereal Sci. 2002, 35, 65–78. [Google Scholar]
- Yoshida, H.; Alexander, J.C. Enzymatic hydrolysis in vitro of thermally oxidized sunflower oil. Lipids 1983, 18, 611–616. [Google Scholar]
- Folch, J.; Lee, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Yoshida, H.; Shougaki, Y.; Hirakawa, Y.; Tomiyama, Y.; Mizushina, Y. Lipid classes, fatty acid composition and triacylglycerol molecular species in the kernels of pumpkin (Cucurbita spp.) seeds. J. Sci. Food Agric. 2004, 84, 158–163. [Google Scholar] [CrossRef]
- Kitts, D.D.; Huynh, M.D.; Hu, C.; Trites, A.W. Sesason variation in nutrient composition of Alaskan walley pollock. Can. J. Zool. 2004, 82, 1408–1415. [Google Scholar]
- Yoshida, H.; Tomiyama, Y.; Yoshida, N.; Saiki, M.; Mizushina, Y. Lipid classes, fatty acid distributions and triacylglycerol molecular species of broad beans (Vicia faba). J. Am. Oil Chem. Soc. 2008, 87, 535–541. [Google Scholar]
- Yoshida, H.; Saiki, M.; Yoshida, N.; Tomiyama, Y.; Mizushina, Y. Fatty acid distribution in triacylglycerols and phospholipids of broad beans (Vicia faba). Food Chem. 2010, 112, 924–928. [Google Scholar]
- Steel, R.C.D.; Torrie, J.H.; Dickey, D. Principles and Procedures of Statistics-A Biomedical Approach,3rd ed.; McGraw-Hill: New York, NY, USA, 1995. [Google Scholar]
- Baker, R.J. Multiple comparison tests. Can. J. Plant Sci. 1980, 60, 325–327. [Google Scholar]
- Okunishi, T.; Ohtsubo, K. Lipid derivatives in brown rice for various storage and suppression methods. Nippon Shokuhi Kagaku Kogaku Kaish 2008, 55, 76–77. [Google Scholar] [CrossRef]
- Hirayama, O.; Matsuda, H. Lipid components and distribution in brown rice. Nippon Nogeikagaku Kaishi 1973, 7, 371–377. [Google Scholar]
- Alvarez-Ortega, R.; Cantisan, S.; Martinez-Force, E.; Garces, R. Characterization of polar and nonpolar seed lipid classes from highly saturated fatty acid sunflower mutants. Lipids 1997, 32, 833–837. [Google Scholar]
- Reske, J.; Siebrecht, J.; Hazebroek, J. Triacylglycerol composition and structure in genetically modified sunflower and soybean oil. J. Am. Oil Chem. Soc. 1997, 74, 989–998. [Google Scholar]
- Arcos, J.A.; Garci, H.S.; Hil, C.G., Jr. Regioselective analysis of the fatty acid composition of triacylglycerols with conventional high performance liquid chromatography. J. Am. Oil Chem. Soc. 2000, 77, 507–512. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yoshida, H.; Tanigawa, T.; Kuriyama, I.; Yoshida, N.; Tomiyama, Y.; Mizushina, Y. Variation in Fatty Acid Distribution of Different Acyl Lipids in Rice (Oryza sativa L.) Brans. Nutrients 2011, 3, 505-514. https://0-doi-org.brum.beds.ac.uk/10.3390/nu3040505
Yoshida H, Tanigawa T, Kuriyama I, Yoshida N, Tomiyama Y, Mizushina Y. Variation in Fatty Acid Distribution of Different Acyl Lipids in Rice (Oryza sativa L.) Brans. Nutrients. 2011; 3(4):505-514. https://0-doi-org.brum.beds.ac.uk/10.3390/nu3040505
Chicago/Turabian StyleYoshida, Hiromi, Takaaki Tanigawa, Isoko Kuriyama, Naoko Yoshida, Yuka Tomiyama, and Yoshiyuki Mizushina. 2011. "Variation in Fatty Acid Distribution of Different Acyl Lipids in Rice (Oryza sativa L.) Brans" Nutrients 3, no. 4: 505-514. https://0-doi-org.brum.beds.ac.uk/10.3390/nu3040505