Health Benefits of Methylxanthines in Cacao and Chocolate



Abstract
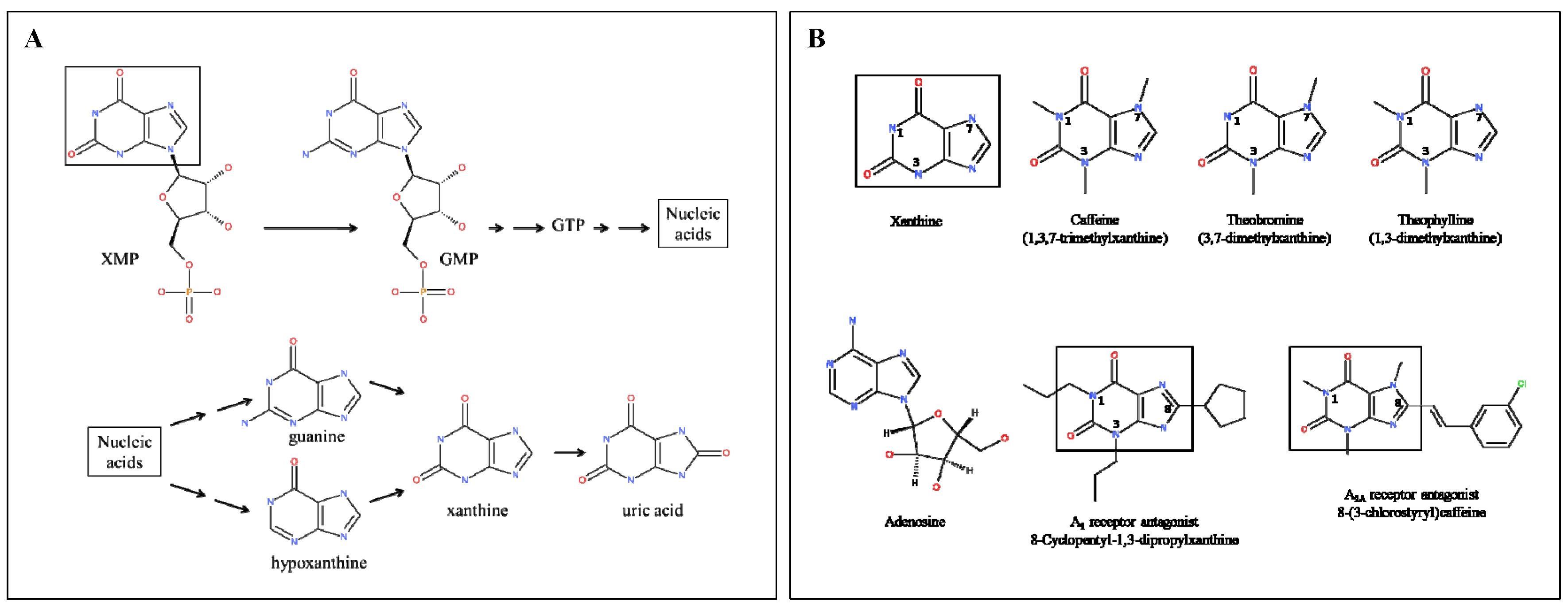
:1. Introduction

2. Flavonols and Methylxanthines in Cacao
3. Chocolate Appeal and Methylxanthine Reward
4. Methylxanthine Levels

{kind=link}
| Source | Theobromine | Caffeine | Theophylline |
|---|---|---|---|
| Raw ground paste (mg/kg) | 33,000 | 5600 | 200 |
| Roasted ground paste (mg/kg) | 36,000 | 330 | Below limit of detection |
| Cacao a (mg/kg) | 26,000 | 2400 | Not determined |
| Cacao butter a (mg/kg) | 140 | 400 | Not determined |
| Cacao a (mg/kg) | 4621 | 489 | Below limit of detection |
| Baking chocolate a (mg/kg of sample) | 10,040 | 1580 | Below limit of detection |
| Milka chocolate a (mg/kg of sample) | 1004 | 56 | Below limit of detection |
| Dark chocolate b (mg/kg) | 5000–7500 | 625–875 | Not determined |
5. Safety of Caffeine and Theobromine
6. Mechanism of Action of Methylxanthines
7. Methylxanthines as Adenosine Receptor Antagonists
8. Physiological and Health Benefits of Methylxanthines in Cacao
8.1. Theobromine in Oral Health
8.2. Methylxanthines in Respiratory Tract Diseases
8.3. Methylxanthines as Psycho-Stimulants
8.4. Methylxanthines and Sleep
8.5. Methylxanthines and Neurodegenerative Diseases
8.6. Methylxanthines in Hypertension and Cardiovascular Diseases
8.7. More Clinical Trials on Cacao Effects on Hypertension and Blood Vessel Status
8.8. Cacao in Insulin Resistance and Body Weight
9. Conclusions
Conflicts of Interest
References
- Andújar, I.; Recio, M.C.; Giner, R.M.; Ríos, J.L. Cocoa polyphenols and their potential benefits for human health. Oxid. Med. Cell. Longev. 2012, 2012. [Google Scholar] [CrossRef]
- Gu, Y.; Yu, S.; Lambert, J.D. Dietary cocoa ameliorates obesity-related inflammation in high fat-fed mice. Eur. J. Nutr. 2013. [Google Scholar] [CrossRef]
- Redovniković, I.R.; Delonga, K.; Mazor, S.; Dragović-Uzelac, V.; Caric, M.; Vorkapic- Furac, J. Polyphenolic content and composition, and antioxidative activity of different cocoa liquors. Czech J. Food Sci. 2009, 27, 330–337. [Google Scholar]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2011, 93, 62–72. [Google Scholar] [CrossRef]
- Hayek, N. Chocolate, gut microbiota, and human health. Front. Pharmacol. 2013, 4. [Google Scholar] [CrossRef]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Sotelo, A.; Soleri, D.; Wacher, C.; Sánchez-Chinchillas, A.; Argote, R.M. Chemical and nutritional composition of tejate, a traditional maize and cacao beverage from the Central Valleys of Oaxaca, Mexico. Plant Foods Hum. Nutr. 2012, 67, 148–155. [Google Scholar] [CrossRef]
- Crown, P.L.; Emerson, T.E.; Gu, J.; Hurst, W.J.; Pauketat, T.R.; Ward, T. Ritual black drink consumption at Cahokia. Proc. Natl. Acad. Sci. USA 2012, 109, 13944–13949. [Google Scholar]
- Nehlig, A. Is caffeine a cognitive enhancer? J. Alzheimer’s Dis. 2010, 2, S85–S94. [Google Scholar]
- Dórea, J.G.; da Costa, T.H. Is coffee a functional food? Br. J. Nutr. 2005, 93, 773–782. [Google Scholar] [CrossRef]
- Sullivan, R.J.; Hagen, E.H.; Hammerstein, P. Revealing the paradox of drug reward in human evolution. Proc. Biol. Sci. 2008, 275, 1231–1241. [Google Scholar] [CrossRef]
- Schiffman, S.S.; Gill, J.M.; Diaz, C. Methylxanthines enhance taste: Evidence for modulation of taste by adenosine receptor. Pharmacol. Biochem. Behav. 1985, 22, 195–203. [Google Scholar] [CrossRef]
- Wright, G.A.; Baker, D.D.; Palmer, M.J.; Stabler, D.; Mustard, J.A.; Power, E.F.; Borland, A.M.; Stevenson, P.C. Caffeine in floral nectar enhances a pollinator’s memory of reward. Science 2013, 339, 1202–1204. [Google Scholar]
- Trognitz, B.; Cros, E.; Assemat, S.; Davrieux, F.; Forestier-Chiron, N.; Ayestas, E.; Kuant, A.; Scheldeman, X.; Hermann, M. Diversity of cacao trees in Waslala, Nicaragua: Associations between genotype spectra, product quality and yield potential. PLoS One 2013, 8, e54079. [Google Scholar]
- Lo Coco, F.; Lanuzza, F.; Micali, G.; Cappellano, G. Determination of theobromine, theophylline, and caffeine in by-products of cupuacu and cacao seeds by high-performance liquid chromatography. J. Chromatogr. Sci. 2007, 45, 273–275. [Google Scholar] [CrossRef]
- Srdjenovic, B.; Djordjevic-Milic, V.; Grujic, N.; Injac, R.; Lepojevic, Z. Simultaneous HPLC determination of caffeine, theobromine, and theophylline in food, drinks, and herbal produts. J. Chromatogr. Sci. 2008, 46, 144–149. [Google Scholar] [CrossRef]
- Risner, C.H. Simultaneous determination of theobromine, (+)-catechin, caffeine, and (−)-epicatechin in standard reference material baking chocolate 2384, cocoa, cocoa beans, and cocoa butter. J. Chromatogr. Sci. 2008, 46, 892–899. [Google Scholar] [CrossRef]
- Bruinsma, K.; Taren, D.L. Chocolate: Food or drug? J. Am. Diet. Assoc. 1999, 99, 1249–1256. [Google Scholar] [CrossRef]
- Baselt, R.C. Disposition of Toxic Drugs and Chemicals in Man; Biomedical Publications: Seal Beach, CA, USA, 1982. [Google Scholar]
- Lelo, A.; Birkett, D.J.; Robson, R.A.; Miners, J.O. Comparative pharmacokinetics of caffeine and its primary demethylated metabolites paraxanthine, theobromine and theophylline in man. Br. J. Clin. Pharmacol. 1986, 22, 177–182. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Persson, C.G. Xanthine derivatives as adenosine receptor antagonists. Eur. J. Pharmacol. 1982, 81, 673–676. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Irenius, E.; Kull, B.; Schulte, G. Comparison of the potency of adenosine as an agonist at human adenosine receptors expressed in Chinese hamster ovary cells. Biochem. Pharmacol. 2001, 61, 443–448. [Google Scholar] [CrossRef]
- Johnson, I.M.; Prakash, H.; Prathiba, J.; Raghunathan, R.; Malathi, R. Spectral analysis of naturally occurring methylxanthines (theophylline, theobromine and caffeine) binding with DNA. PLoS One 2012, 7, e50019. [Google Scholar]
- Gans, J.H.; Korson, R.; Cater, M.R.; Ackerly, C.C. Effects of short-term and long-term theobromine administration to male dogs. Toxicol. Appl. Pharmacol. 1980, 53, 481–496. [Google Scholar] [CrossRef]
- Clark, N. Caffeine: A user’s guide. Phys. Sports Med. 1997, 25, 109–110. [Google Scholar]
- Higdon, J.V.; Frei, B. Coffee and health: A review of recent human research. Crit. Rev. Food Sci. Nutr. 2006, 46, 101–123. [Google Scholar] [CrossRef]
- Hornfeldt, C.S. Chocolate toxicity in dogs. Mod. Vet. Pract. 1987, 68, 552–554. [Google Scholar]
- Baggott, M.J.; Childs, E.; Hart, A.B.; de Bruin, E.; Palmer, A.A.; Wilkinson, J.E.; de Wit, H. Psychopharmacology of theobromine in healthy volunteers. Psychopharmacology 2013, 228, 109–118. [Google Scholar] [CrossRef]
- Pendleton, M.; Brown, S.; Thomas, C.; Odle, B. Potential toxicity of caffeine when used as a dietary supplement for weight loss. J. Diet. Suppl. 2012, 9, 293–298. [Google Scholar] [CrossRef]
- Sinchai, T.; Plasen, S.; Sanvarinda, Y.; Jaisin, Y.; Govitrapong, P.; Morales, N.P.; Ratanachamnong, P.; Plasen, D. Caffeine potentiates methamphetamine-induced toxicity both in vitro and in vivo. Neurosci. Lett. 2011, 502, 65–69. [Google Scholar] [CrossRef]
- Arria, A.M.; O’Brien, M.C. The “high” risk of energy drinks. JAMA 2011, 305, 600–601. [Google Scholar] [CrossRef]
- Chen, J.F.; Chern, Y. Impacts of methylxanthines and adenosine receptors on neurodegeneration: Human and experimental studies. Handb. Exp. Pharmacol. 2011, 200, 267–310. [Google Scholar] [CrossRef]
- McPherson, P.S.; Kim, Y.K.; Valdivia, H.; Knudson, C.M.; Takekura, H.; Franzini-Armstrong, C.; Coronado, R.; Campbell, K.P. The brain ryanodine receptor: A caffeine-sensitive calcium release channel. Neuron 1991, 7, 17–25. [Google Scholar] [CrossRef]
- Choi, O.H.; Shamim, M.T.; Padgett, W.L.; Daly, J.W. Caffeine and theophylline analogues: correlation of behavioral effects with activity as adenosine receptor antagonists and as phosphodiesterase inhibitors. Life Sci. 1988, 43, 387–398. [Google Scholar] [CrossRef]
- Marangos, P.J.; Paul, S.M.; Parma, A.M.; Goodwin, F.K.; Syapin, P.; Skolnick, P. Purinergic inhibition of diazepam binding to rat brain (in vitro). Life Sci. 1979, 24, 851–857. [Google Scholar] [CrossRef]
- Hove-Madsen, L.; Prat-Vidal, C.; Llach, A.; Ciruela, F.; Casadó, V.; Lluis, C.; Bayes-Genis, A.; Cinca, J.; Franco, R. Adenosine A2A receptors are expressed in human atrial myocytes and modulate spontaneous sarcoplasmic reticulum calcium release. Cardiovasc. Res. 2006, 72, 292–302. [Google Scholar] [CrossRef]
- Llach, A.; Molina, C.E.; Prat-Vidal, C.; Fernandes, J.; Casadó, V.; Ciruela, F.; Lluís, C.; Franco, R.; Cinca, J.; Hove-Madsen, L. Abnormal calcium handling in atrial fibrillation is linked to up-regulation of adenosine A2A receptors. Eur. Heart J. 2011, 32, 721–729. [Google Scholar] [CrossRef]
- Ciruela, F.; Casadó, V.; Rodrigues, R.J.; Luján, R.; Burgueño, J.; Canals, M.; Borycz, J.; Rebola, N.; Goldberg, S.R.; Mallol, J.; et al. Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2A receptor heteromers. J. Neurosci. 2006, 26, 2080–2087. [Google Scholar] [CrossRef]
- Orru, M.; Bakešová, J.; Brugarolas, M.; Quiroz, C.; Beaumont, V.; Goldberg, S.R.; Lluís, C.; Cortés, A.; Franco, R.; Casadó, V.; et al. Striatal pre- and postsynaptic profile of adenosine A2A receptor antagonists. PLoS One 2011, 6, e16088. [Google Scholar] [CrossRef]
- Lazarus, M.; Shen, H.Y.; Cherasse, Y.; Qu, W.M.; Huang, Z.L.; Bass, C.E.; Winsky-Sommerer, R.; Semba, K.; Fredholm, B.B.; Boison, D.; et al. Arousal effect of caffeine depends on adenosine A2A receptors in the shell of the nucleus accumbens. J. Neurosci. 2011, 31, 10067–10075. [Google Scholar] [CrossRef] [Green Version]
- Kargul, B.; Ozcan, M.; Peker, S.; Nakamoto, T.; Simmons, W.B.; Falster, A.U. Evaluation of human enamel surfaces treated with theobromine: A pilot study. Oral Health Prev. Dent. 2012, 10, 275–282. [Google Scholar]
- Usmani, O.S.; Belvisi, M.G.; Patel, H.J.; Crispino, N.; Birrell, M.A.; Korbonits, M.; Korbonits, D.; Barnes, P.J. Theobromine inhibits sensory nerve activation and cough. FASEB J. 2005, 19, 231–233. [Google Scholar]
- Coleman, R.A. Purine antagonists in the identification of adenosine-receptors in guinea-pig trachea and the role of purines in non-adrenergic inhibitory neurotransmission. Br. J. Pharmacol. 1980, 69, 359–366. [Google Scholar] [CrossRef]
- Halfdanarson, T.R.; Jatoi, A. Chocolate as a cough suppressant: Rationale and justification for an upcoming clinical trial. Support. Cancer Ther. 2007, 4, 119–122. [Google Scholar] [CrossRef]
- Bara, A.I.; Barley, E.A. Caffeine for asthma. Cochrane Database Syst. Rev. 2001. [Google Scholar] [CrossRef]
- Simons, F.E.; Becker, A.B.; Simons, K.J.; Gillespie, C.A. The bronchodilator effect and pharmacokinetics of theobromine in young patients with asthma. J. Allergy Clin. Immunol. 1985, 76, 703–707. [Google Scholar] [CrossRef]
- Pagano, R.; Negri, E.; Decarli, A.; La Vecchia, C. Coffee drinking and prevalence of bronchial asthma. Chest 1988, 94, 386–389. [Google Scholar] [CrossRef]
- Zhao, J.; Gonzalez, F.; Mu, D. Apnea of prematurity: From cause to treatment. Eur. J. Pediatr. 2011, 170, 1097–1105. [Google Scholar] [CrossRef]
- Aranda, J.V.; Beharry, K.; Valencia, G.B.; Natarajan, G.; Davis, J. Caffeine impact on neonatal morbidities. J. Matern. Fetal Neonatal Med. 2010, 23, 20–23. [Google Scholar] [CrossRef]
- Henderson-Smart, D.J.; Steer, P.A. Caffeine versus theophylline for apnea in preterm infants. Cochrane Database Syst. Rev. 2010. [Google Scholar] [CrossRef]
- Franco, R. Coffee and mental health. Aten. Primaria 2009, 41, 578–581. [Google Scholar] [CrossRef]
- Smit, H.J.; Gaffan, E.A.; Rogers, P.J. Methylxanthines are the psycho-pharmacologically active constituents of chocolate. Psychopharmacology 2004, 176, 412–419. [Google Scholar] [CrossRef]
- Grandner, M.A.; Jackson, N.; Gerstner, J.R.; Knutson, K.L. Dietary nutrients associated with short and long sleep duration. Data from a nationally representative sample. Appetite 2013, 64, 71–80. [Google Scholar] [CrossRef]
- Yang, A.; Palmer, A.A.; de Wit, H. Genetics of caffeine consumption and responses to caffeine. Psychopharmacology 2010, 211, 245–257. [Google Scholar] [CrossRef]
- Costa, J.; Lunet, N.; Santos, C.; Santos, J.; Vaz-Carneiro, A. Caffeine exposure and the risk of Parkinson’s disease: A systematic review and meta-analysis of observational studies. J. Alzheimer’s Dis. 2010, 20, S221–S238. [Google Scholar]
- Maia, L.; de Mendonca, A. Does caffeine intake protect from Alzheimer’s disease? Eur. J. Neurol. 2002, 9, 377–382. [Google Scholar] [CrossRef]
- Eskelinen, M.H.; Ngandu, T.; Tuomilehto, J.; Soininen, H.; Kivipelto, M. Midlife coffee and tea drinking and the risk of late-life dementia: A population-based CAIDE study. J. Alzheimer’sDis. 2009, 16, 85–91. [Google Scholar]
- Pelligrino, D.A.; Xu, H.L.; Vetri, F. Caffeine and the control of cerebral hemodynamics. J. Alzheimer’s Dis. 2010, 20, S51–S62. [Google Scholar]
- Klaassen, E.B.; de Groot, R.H.; Evers, E.A.; Snel, J.; Veerman, E.C.; Ligtenberg, A.J.; Jolles, J.; Veltman, D.J. The effect of caffeine on working memory load-related brain activation in middle-aged males. Neuropharmacology 2013, 64, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Koppelstaetter, F.; Poeppel, T.D.; Siedentopf, C.M.; Ischebeck, A.; Verius, M.; Haala, I.; Mottaghy, F.M.; Rhomberg, P.; Golaszewski, S.; Gotwald, T.; et al. Does caffeine modulate verbal working memory processes? An fMRI study. Neuroimage 2008, 39, 492–499. [Google Scholar] [CrossRef]
- Haller, S.; Rodriguez, C.; Moser, D.; Toma, S.; Hofmeister, J.; Sinanaj, I.; Van De Ville, D.; Giannakopoulos, P.; Lovblad, K.O. Acute caffeine administration impact on working memory-related brain activation and functional connectivity in the elderly: A BOLD and perfusion MRI study. Neuroscience 2013, 250, 364–371. [Google Scholar] [CrossRef]
- Han, M.E.; Kim, H.J.; Lee, Y.S.; Kim, D.H.; Choi, J.T.; Pan, C.S.; Yoon, S.; Baek, S.Y.; Kim, B.S.; Kim, J.B.; et al. Regulation of cerebrospinal fluid production by caffeine consumption. BMC Neuroscience 2009, 10, 110. [Google Scholar] [CrossRef]
- Wostyn, P.; van Dam, D.; Audenaert, K.; de Deyn, P.P. Increased cerebrospinal fluid production as a possible mechanism underlying Caffeine’s protective effect against Alzheimer’s disease. Int. J. Alzheimer’s Dis. 2011, 2011. [Google Scholar] [CrossRef]
- Bond, G.S. Effect of various agents on the blood flow through the coronary arteries and veins. J. Exp. Med. 1910, 12, 575–585. [Google Scholar] [CrossRef]
- Askanazy, S. Klinisches über Diuretin. In Deutsches Archiv für klinische Medicin; Vogel Editions: Leipzig, Germany, 1986; pp. 209–230. [Google Scholar]
- Dock, W. The use of theobromine for pain of arteriosclerotic origin. Calif. West. Med. 1926, 25, 636–638. [Google Scholar]
- McGovern, T.; McDevitt, E.; Wright, I.S. Theobromine sodium salicylate as a vasodilator. J. Clin. Investig. 1936, 15, 11–16. [Google Scholar] [CrossRef]
- Van den Bogaard, B.; Draijer, R.; Westerhof, B.E.; van den Meiracker, A.H.; van Montfrans, G.A.; van den Born, B.J. Effects on peripheral and central blood pressure of cocoa with natural or high-dose theobromine: A randomized, double-blind crossover trial. Hypertension 2010, 56, 839–846. [Google Scholar]
- Khan, N.; Monagas, M.; Andres-Lacueva, C.; Casas, R.; Urpí-Sardà, M.; Lamuela-Raventós, R.M.; Estruch, R. Regular consumption of cocoa powder with milk increases HDL cholesterol and reduces oxidized LDL levels in subjects at high-risk of cardiovascular disease. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 1046–1053. [Google Scholar] [CrossRef]
- Neufingerl, N.; Zebregs, Y.E.; Schuring, E.A.; Trautwein, E.A. Effect of cocoa and theobromine consumption on serum HDL-cholesterol concentrations: A randomized controlled trial. Am. J. Clin. Nutr. 2013, 97, 1201–1209. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Lindgren, E. The effect of alkylxanthines and other phosphodiesterase inhibitors on adenosine-receptor mediated decrease in lipolysis and cyclic AMP accumulation in rat fat cells. Acta Pharmacol. Toxicol. 1984, 54, 64–71. [Google Scholar] [CrossRef]
- Grassi, D.; Lippi, C.; Necozione, S.; Desideri, G.; Ferri, C. Short-term administration of dark chocolate is followed by a significant increase in insulin sensitivity and a decrease in blood pressure in healthy persons. Am. J. Clin. Nutr. 2005, 81, 611–614. [Google Scholar]
- Kelly, C.J. Effects of theobromine should be considered in future studies. Am. J. Clin. Nutr. 2005, 82, 486–487. [Google Scholar]
- Figler, R.A.; Wang, G.; Srinivasan, S.; Jung, D.Y.; Zhang, Z.; Pankow, J.S.; Ravid, K.; Fredholm, B.; Hedrick, C.C.; Rich, S.S.; et al. Links between insulin resistance, adenosine A2B receptors, and inflammatory markers in mice and humans. Diabetes 2011, 60, 669–679. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Franco, R.; Oñatibia-Astibia, A.; Martínez-Pinilla, E. Health Benefits of Methylxanthines in Cacao and Chocolate. Nutrients 2013, 5, 4159-4173. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5104159
Franco R, Oñatibia-Astibia A, Martínez-Pinilla E. Health Benefits of Methylxanthines in Cacao and Chocolate. Nutrients. 2013; 5(10):4159-4173. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5104159
Chicago/Turabian StyleFranco, Rafael, Ainhoa Oñatibia-Astibia, and Eva Martínez-Pinilla. 2013. "Health Benefits of Methylxanthines in Cacao and Chocolate" Nutrients 5, no. 10: 4159-4173. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5104159