Vitamin B12 and Folic Acid Imbalance Modifies NK Cytotoxicity, Lymphocytes B and Lymphoprolipheration in Aged Rats


Abstract
:1. Introduction
2. Material and Methods
2.1. Animals and Diets
2.2. Dietary Treatment
2.3. Sacrifice and Tissue Handling
2.4. Serum Folate
2.5. Hepatic Folate
2.6. Serum Vitamin B12
2.7. Isolation of Rat Blood Neutrophils and Lymphocytes
2.8. Lymphoproliferation
2.9. Cytotoxicity Assay
2.10. Interleukins (IL), Granulate-Macrophage Stimulating Factor Precursor (GM-CSF), Interferon Gamma (IFN-γ), Tumor Necrosis Factor Alpha (TNF-α) Levels
2.11. Statistical Analysis
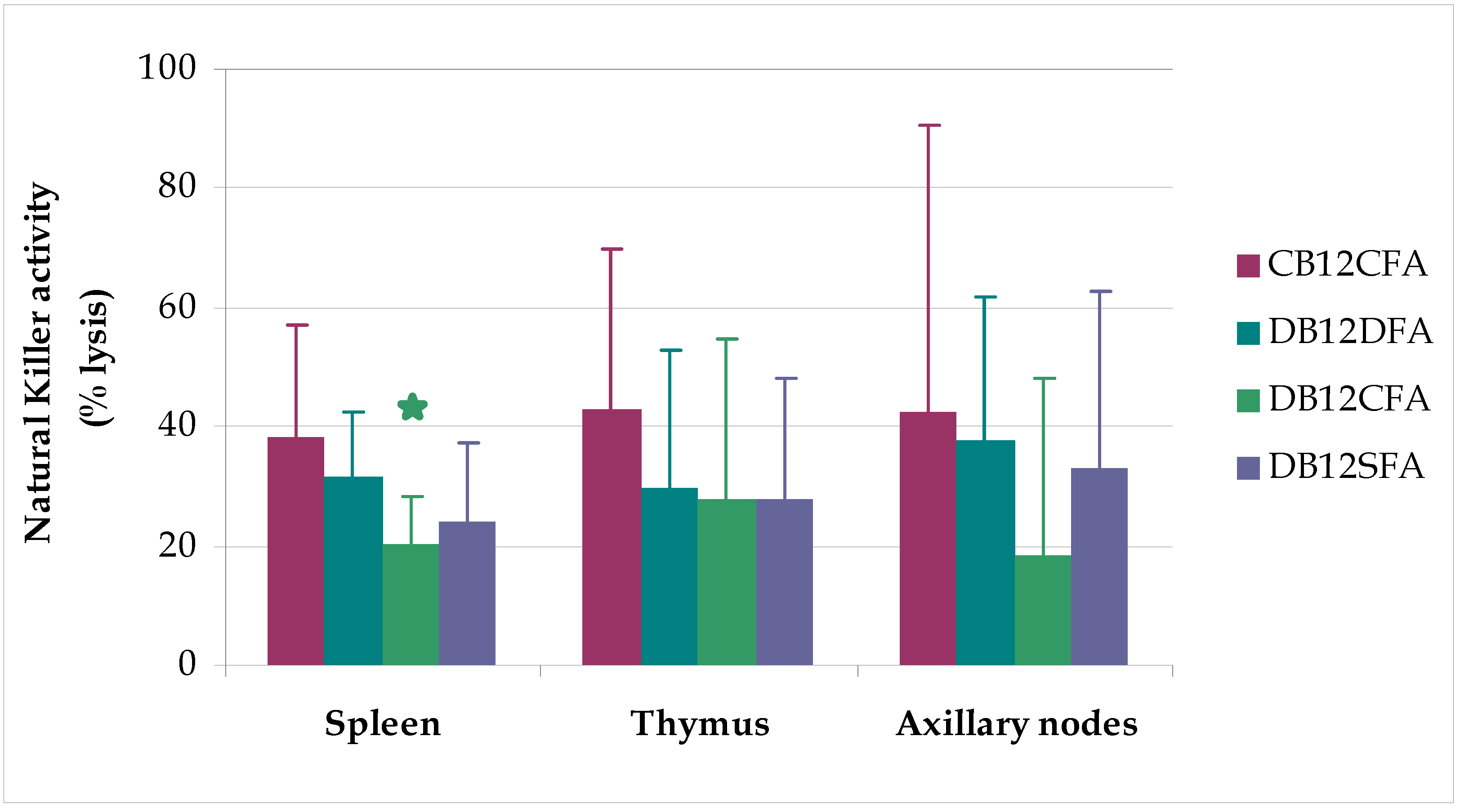
3. Results

{kind=link}
| Group | Serum Folate (ng/mL) | Liver Folate (μg/g liver) | Serum Vitamin B12 (ng/L) |
|---|---|---|---|
| CB12CFA | 38.6 | 18.3 | 1203.4 |
| n = 8 | (34.1 to 43.1) | (14.7 to 22.0) | (1036.6 to 1370.3) |
| DB12DFA | 4.6 ***,### | 7.6 **,# | 744.4 *** |
| n = 9 | (4.0 to 5.2) | (4.9 to 10.2) | (663.0 to 825.7) |
| DB12CFA | 62.5 * | 16.1 | 906.8 *** |
| n = 9 | (54.2 to 70.8) | (11.0 to 21.2) | (845.7 to 967.9) |
| DB12SFA | 114.5 ***,### | 29.6 **,## | 807.6 *** |
| n = 9 | (92.8 to 136.3) | (23.6 to 35.7) | (738.1 to 877.0) |

| Group | Helper Tcells | Cytotoxic-Supressor T cells | CD4/CD8 | Mature Tcells | B lymphocytes | Natural Killer Cells |
|---|---|---|---|---|---|---|
| CB12CFA | 24.6 | 9.6 | 2.7 | 35.5 | 18.2 | 15.6 |
| n = 7 | (18.1 to 31.1) | (6.7 to 12.5) | (2.0 to 3.3) | (26.9 to 44.1) | (12.3 to 24.0) | (9.82 to 21.4) |
| DB12DFA | 20.6 | 10.0 | 2.2 | 32.2 | 12.5 | 15.7 |
| n = 8 | (13.7 to 27.4) | (6.6 to 13.3) | (1.5 to 2.8) | (22.8 to 41.6) | (6.2 to 18.9) | (4.8 to 26.5) |
| DB12CFA | 26.5 | 10.4 | 2.6 | 38.4 | 7.3 * | 5.0 |
| n = 7 | (20.1 to 32.8) | (7.5 to 13.4) | (2.1 to 3.1) | (29.2 to 47.5) | (2.1 to 12.5) | (1.7 to 8.4) |
| DB12SFA | 21.6 | 7.5 | 3.5 | 30.4 | 8.9 * | 28.0 |
| n = 8 | (18.3 to 24.9) | (4.5 to 10.6) | (2.0 to 5.0) | (24.3 to 36.4) | (4.0 to 13.7) | (11.9 to 44.1) |
| Organ | Mitogen | CB12CFA | DB12DFA | DB12CFA | DB12SFA |
| n = 8 | n = 9 | n = 9 | n = 9 | ||
| Spleen | Con A | 138.1 | 168.0 | 185.4 | 146.3 |
| (87.4 to 188.8) | (111.1 to 224.9) | (173.6 to 197.2) | (110.7 to 181.9) | ||
| LPS | 140.0 | 178.1 | 223.0 | 207.4 * | |
| (98.3 to 181.7) | (129.1 to 227.2) | (142.5 to 303.5) | (133.3 to 281.6) | ||
| PHA | 177.1 | 164.1 | 270.8 | 180.6 | |
| (90.1 to 264.2) | (130.9 to 197.4) | (156.8 to 384.8) | (97.0 to 264.1) | ||
| Organ | Mitogen | CB12CFA | DB12DFA | DB12CFA | DB12SFA |
| n = 8 | n = 9 | n = 8 | n = 9 | ||
| Thymus | Con A | 111.3 | 161.1 | 151.3 | 125.2 |
| (58.9 to 163.7) | (126.7 to 195.5) | (102.5 to 200.1) | (116.9 to 133.5) | ||
| LPS | 152.9 | 148.6 | 141.0 | 157.8 | |
| (59.2 to 246.6) | (108.8 to 188.3) | (92.7 to 189.3) | (131.8 to 183.8) | ||
| PHA | 161.3 | 149.9 | 204.9 | 154.8 | |
| (53.3 to 269.3) | (124.7 to 175.0) | (132.9 to 276.8) | (121.2 to 188.4) | ||
| Organ | Mitogen | CB12CFA | DB12DFA | DB12CFA | DB12SFA |
| n = 6 | n = 8 | n = 7 | n = 7 | ||
| Axillary nodes | Con A | 115.6 | 152.0 | 171.0 | 131.5 |
| (102.2 to 129.0) | (74.1 to 229.9) | (42.0 to 300.0) | (108.9 to 154.1) | ||
| LPS | 110.2 | 191.2 | 211.7 | 139.0 | |
| (63.8 to 156.6) | (109.8 to 272.5) | (27.7 to 395.6) | (105.9 to 172.1) | ||
| PHA | 145.2 | 190.2 | 181.0 | 149.5 | |
| (125.0 to 165.4) | (148.3 to 232.0) | (43.6 to 318.4) | (81.0 to 218.0) |
| Cytokines | Mitogen | CB12CFAn = 5 | DB12DFAn = 5 | DB12CFAn = 5 | DB12SFAn = 5 |
|---|---|---|---|---|---|
| n = 5 | n = 5 | n = 5 | n = 5 | ||
| IL-1a (pg/mL) | Basal | 54.9 (53.9 –55.9) | 54.6 (46.7 –62.6) | 53.8 (49.6 –57.9) | 53.5 (50.7 –56.3) |
| Con A | 56.0 (43.3 –68.7) | 54.3 (44.7 –63.8) | 53.3 (51.9 –54.8) | 58.0 (46.2 –69.8) | |
| LPS | 64.3 (54.3 –74.4) | 58.0 (55.5 –60.5) | 62.0 (48.9 –75.1) | 62.6 (56.8 –68.5) | |
| PHA | 58.0 (54.6 –61.4) | 59.1 (53.8 –64.5) | 55.8 (50.7 –60.9) | 54.3 (52.8 –55.8) | |
| IL-1b (pg/mL) | Basal | 13.8 (3.0 –24.7) | 15.0 (6.9 –23.1) | 18.3 (8.8 –27.7) | 13.1 (10.7 –15.5) |
| Con A | 9.7 (2.1 –17.3) | 9.1 (−6.1 –24.3) | 14.2 (5.3 –23.1) | 12.1 (8.5 –15.6) | |
| LPS | 20.5 (17.1–23.9) | 17.3 (17.3–17.3) | 26.6 (9.6–43.6) | 18.5 (11.3–25.8) | |
| PHA | 18.0 (3.05–33.0) | 18.1 (6.4–29.8) | 18.5 (12.0–25.1) | 12.1 (8.9–15.4) | |
| IL-4 (pg/mL) | Basal | 4.39 (4.35–4.42) | 4.40 (4.34–4.45) | 4.39 (4.35–4.42) | 4.38 (4.36–4.41) |
| Con A | 4.42 (4.18–4.67) | 4.39 (4.32–4.45) | 4.38 (4.33–4.43) | 4.39 (4.35–4.42) | |
| LPS | 4.39 (4.34–4.45) | 4.39 (4.34–4.45) | 4.39 (4.34–4.45) | 4.38 (4.35–4.41) | |
| PHA | 4.43 (4.41–4.45) | 4.41 (4.41–4.41) | 4.41 (4.41–4.41) | 4.31 (4.13–4.49) | |
| IL-12 (pg/mL) | Basal | 31.5 (−2.8–65.7) | 24.8 (9.3–40.3) | 27.4 (9.5–45.2) | 35.5 (27.3–43.7) |
| Con A | 18.9 (−67.5–105.3) | 28.2 (18.1–38.3) | 29.1 (14.6–43.6) | 18.7 (27.3–36.7) | |
| LPS | 54.1 (33.0–75.2) | 30.8 (18.3–43.4) | 61.1 (−4.5–126.7) | 54.3 (27.2–81.4) | |
| PHA | 29.4 (−8.2–66.9) | 29.1 (14.6–43.6) | 41.2 (12.4–69.9) | 32.5 (22.8–42.2) | |
| GM-CSF (pg/mL) | Basal | 12.9 (11.9–14.0) | 13.6 (12.6–14.5) | 13.3 (13.3–13.3) | 12.5 (11.6–13.4) |
| Con A | 13.9 (6.1–21.7) | 13.5 (12.4–14.6) | 12.4 (10.5–14.3) | 12.9 (11.9–14.0) | |
| LPS | 13.7 (11.9–15.4) | 13.7 (11.9–15.4) | 12.8 (11.0–14.7) | 13.6 (12.6–14.5) | |
| PHA | 13.7 (11.9–15.4) | 13.7 (11.9–15.4) | 13.3 (13.3–13.3) | 13.3 (13.3–13.3) | |
| IFN-γ (pg/mL) | Basal | 3.0 (1.3–4.7) | 2.0 (−0.2–4.1) | 2.7 (0.7–4.6) | 3.0 (0.7–5.2) |
| Con A | 2.3 (−6.0–10.8) | 1.7 (1.7–1.7) | 1.2 (−1.0–3.3) | 2.98 (0.6–5.4) | |
| LPS | 3.4 (−0.3–7.1) | 2.1 (0.2–4.0) | 3.0 (−0.2–6.2) | 4.2 (−0.0–8.4) | |
| PHA | 2.8 (1.9–3.7) | 2.1 (0.2–4.03) | 3.0 (1.4–4.6) | 2.7 (2.0–3.5) | |
| TNF-α (pg/mL) | Basal | 13.6 (−1.0–28.2) | 13.9 (0.1–27.7) | 11.9 (5.7–18.2) | 17.6 (11.6–23.5) |
| Con A | 13.9 (1.5–29.1) | 9.9 (3.5–16.3) | 11.0 (0.2–21.9) | 15.5 (5.7–25.3) | |
| LPS | 80.9 (66.1–95.7) | 56.0 (−23.8–135.9) | 67.3 (−11.8–146.3) | 64.4 (41.4–87.3) | |
| PHA | 27.0 (−11.8–65.9) | 17.03 (0.4–33.6) | 44.4 (0.7–88.1) | 34.9 (14.2–55.6) | |
| TNF-α/IL-4 | Basal | 3.1 (−0.2–6.4) | 3.2 (−0.0–6.4) | 2.7 (1.3–4.2) | 4.0 (2.7–5.3) |
| Con A | 3.1 (−1.2–7.5) | 2.1 (1.2–2.9) | 2.7 (1.2–4.3) | 4.2 (1.8–6.7) | |
| LPS | 19.7 (15.3–24.1) | 13.2 (3.7–22.8) | 13.2 (1.6–24.7) | 14.7 (9.5–19.9) | |
| PHA | 8.4 (−0.2–16.9) | 3.9 (0.1–7.6) | 10.1 (0.2–20.0) | 8.1 (3.3–13.0) |
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- US Food and Drug Administration. Food standards: Amendment of standars of identity for enriched grain products to require addition of folic acid. In Fed. Regist.; 1996; 61, pp. 8781–8797. [Google Scholar]
- CDC. Trends in Spina Bifida and Anencephalus in the United States, 1991–2006. Available online: http://www.cdc.gov/nchs/products/pubs/pubd/hestats/spine_anen.htm (accessed on 7 May 2013).
- Honein, M.A.; Paulozzi, L.J.; Mathews, T.J.; Erickson, J.D.; Wong, L.Y. Impact of folic acid fortification of the US food supply on the ocurrence of neural tube defects. JAMA 2001, 285, 2981–2986. [Google Scholar] [CrossRef]
- Mathews, T.J.; Honein, M.A.; Erickson, J.D. Spina bifida and anencephaly prevalence: United States, 1991–2001. MMWR Recomm. Rep. 2002, 51, 9–11. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Spina bifida and anencephaly before and after folic acid mandate—United States, 1995–1996 and 1999–2000. Morb. Mortal. Wkly. Rep. 2004, 53, 362–365. [Google Scholar]
- Samaniego Vaesken, M.L.; Alonso-Aperte, E.; Varela-Moreiras, G. Elaboración de una base de datos del contenido en folatos de los alismentos: Fortificación voluntaria con ácido fólico en España. Nutr. Hosp. 2009, 24, 459–466. [Google Scholar]
- Samaniego-Vaesken, M.L.; Alonso-Aperte, E.; Varela-Moreiras, G. Analysis and evaluation of voluntary folic acid fortification of breakfast cereals in the Spanish market. J. Food Compos. Anal. 2010, 23, 419–423. [Google Scholar] [CrossRef]
- Mills, J.L. Fortification of foods with folic acid: How much is enough? N. Engl. J. Med. 2000, 342, 1442–1445. [Google Scholar] [CrossRef]
- Morris, M.S.; Jacques, P.F.; Rosenberg, I.H.; Selhub, J. Folate and vitamin B-12 status in relation to anemia, macrocytosis, and cognitive impairment in older Americans in the age of folic acid fortification. Am. J. Clin. Nutr. 2007, 85, 193–200. [Google Scholar]
- Field, C.J.; Johnson, I.R.; Schley, P.D. Nutrients and their role in host resistance to infection. J. Leukoc. Biol. 2002, 71, 16–32. [Google Scholar]
- Dhur, A.; Galan, P.; Hercberg, S. Folate status and the immune system. Prog. Food Nutr. Sci. 1991, 15, 43–60. [Google Scholar]
- Courtemanche, C.; Elson-Schwab, I.; Mashiyama, S.T.; Kerry, N.; Ames, B.N. Folate deficiency inhibits the proliferation of primary human CD8+ T lymphocites in vitro. J. Immunol. 2004, 173, 3186–3192. [Google Scholar]
- Field, C.J.; van Aerde, A.; Drager, K.L.; Goruk, S.; Basu, T. Dietary folate improves age-related decreases in lymphocyte function. J. Nutr. Biochem. 2005, 17, 37–44. [Google Scholar]
- Gomez, C.R.; Nomellini, V.; Faunce, D.E.; Kovacs, E.J. Innate immunity and aging. Exp. Gerontol. 2008, 43, 718–728. [Google Scholar] [CrossRef]
- Dorshkind, K.; Montecino-Rodriguez, E.; Signer, R.A. The ageing immune system: Is it ever too old to become young again? Nat. Rev. Immunol. 2009, 9, 57–62. [Google Scholar] [CrossRef]
- Weng, N.P. Aging of the immune system: How much can the adaptive immune system adapt? Immunity 2006, 24, 495–499. [Google Scholar] [CrossRef]
- Troen, A.M.; Mitchell, B.; Sorensen, B.; Wener, M.H.; Johnston, A.; Wood, B.; Selhub, J.; McTiernan, A.; Yasui, Y.; Oral, E.; et al. Unmetabolized folic acid in plasma is associated with reduced natural killer cell cytotoxicity among postmenopausal women. J. Nutr. 2006, 136, 189–194. [Google Scholar]
- Bailey, R.L.; Mills, J.L.; Yetley, E.A.; Gahche, J.J.; Pfeiffer, C.M.; Dwyer, J.T.; Dodd, K.W.; Sempos, C.T.; Betz, J.M.; Picciano, M.F. Unmetabolized serum folic acid and its relation to folic acid intake from diet and supplements in a nationally representative sample of adults aged > or = 60 year in the United States. Am. J. Clin. Nutr. 2010, 92, 383–389. [Google Scholar] [CrossRef]
- Tamura, J.; Kubota, K.; Murakami, H.; Sawamura, M.; Matsushima, T.; Tamura, T.; Saitoh, T.; Kurabayshi, H.; Naruse, T. Immunomodulation by vitamin B12: Augmentation of CD8+ T lymphocytes and natural killer (NK) cell activity in vitamin B12-deficient patients by methyl-B12 treatment. Clin. Exp. Immunol. 1999, 116, 28–32. [Google Scholar] [CrossRef]
- Walzem, R.L.; Clifford, A.J. Folate deficiency in rats fed diets containing free amino acids or intact proteins. J. Nutr. 1988, 118, 1089–1096. [Google Scholar]
- Tamura, T. Microbiological Assays of Folates. In Folic Acid Metabolism in Health and Disease; Wiley-Liss Inc.: Wilmington, DE, USA, 1990; pp. 121–137. [Google Scholar]
- Horne, D.W.; Krumdieck, C.L.; Wagner, C. Properties of folic acid gamma-glutamyl hydrolase (conjugase) in rat bile and plasma. J. Nutr. 1981, 111, 442–449. [Google Scholar]
- Hyun, T.H.; Tamura, T. Trienzyme extraction in combination with microbiologic assay in food folate analysis: An updated review. Exp. Biol. Med. 2005, 230, 444–454. [Google Scholar]
- De la Fuente, M.; Victor, V.M. Anti-oxidants as modulators of immune function. Immunol. Cell Biol. 2000, 78, 49–54. [Google Scholar] [CrossRef]
- Baker, J.W. An innovate lymphocyte preparation system for flow cytometry. Am. Clin. Lab. 1988, 120, 320–324. [Google Scholar]
- Alvarado, C.; Alvarez, P.; Jiménez, L.; De la Fuente, M. Improvement of leukocyte functions in young prematurely aging mice after a 5-week ingestion of a diet supplemented with biscuits enriched in antioxidants. Antioxid. Redox Signal. 2005, 7, 1203–1210. [Google Scholar] [CrossRef]
- De la Fuente, M.; Hernanz, A.; Guayerbas, N.; Victor, V.M.; Arnalich, F. Vitamin E ingestion improves several immune functions in elderly men and women. Free Radic. Res. 2008, 42, 272–280. [Google Scholar] [CrossRef]
- Carrasco, M.; Hernanz, A.; De La Fuente, M. Effect of cholecystokinin and gastrin on human peripheral blood lymphocyte functions, implication of cyclic AMP and interleukin 2. Regul. Pept. 1997, 70, 135–142. [Google Scholar] [CrossRef]
- Ravaglia, G.; Forti, P.; Maioli, F.; Bastagli, L.; Facchini, A.; Mariani, E.; Savarino, L.; Sassi, S.; Cucinotta, D.; Lenaz, G. Effect of micronutrient status on natural killer cell inmune function in health free-living subjects aged >90. Am. J. Clin. Nutr. 2000, 71, 590–598. [Google Scholar]
- Kim, Y.I.; Hayek, M.; Mason, J.B.; Meydani, S.N. Severe folate deficiency impairs natural killer cell-mediated cytotoxicity in rats. J. Nutr. 2002, 132, 1361–1367. [Google Scholar]
- De la Fuente, M. Effects of antioxidants on immune system ageing. Eur. J. Clin. Nutr. 2002, 56, S5–S8. [Google Scholar] [CrossRef]
- De la Fuente, M.; Miquel, J. An update of the oxidation-inflammation theory of aging: The involvement of the immune system in oxi-inflamm-aging. Curr. Pharm. Des. 2009, 15, 3003–3026. [Google Scholar] [CrossRef]
- Gibson, K.L.; Wu, Y.C.; Barnett, Y.; Duggan, O.; Vaughan, R.; Kondeatis, E.; Nilsson, B.O.; Wikby, A.; Kipling, D.; Dunn-Walters, D.K. B-cell diversity decreases in old age and is correlated with poor health status. Aging Cell 2009, 8, 18–25. [Google Scholar] [CrossRef]
- Crawford, A.; Macleod, M.; Schumacher, T.; Corlett, L.; Gray, D. Primary T cell expansion and differentiation in vivo requires antigen presentation by B cells. J. Immunol. 2006, 176, 3498–3506. [Google Scholar]
- Fillatreau, S.; Gray, D.; Anderton, S.M. Not always the bad guys: B cells as regulators of autoimmune pathology. Nat. Rev. Immunol. 2008, 8, 391–397. [Google Scholar] [CrossRef]
- Funada, U.; Wada, M.; Kawata, T.; Mori, K.; Tamai, H.; Kawanishi, T.; Kunou, A.; Tanaka, N.; Tadokoro, T.; Maekawa, A. Changes in CD4+CD8−/CD4−CD8+ ratio and humoral immune functions in vitamin B12-deficient rats. Int. J. Vitam. Nutr. Res. 2000, 70, 167–171. [Google Scholar] [CrossRef]
- Funada, U.; Wada, M.; Kawata, T.; Mori, K.; Tamai, H.; Isshiki, T.; Onoda, J.; Tanaka, N.; Tadokoro, T.; Maekawa, A. Vitamin B-12-deficiency affects immunoglobulin production and cytokine levels in mice. Int. J. Vitam. Nutr. Res. 2001, 71, 60–65. [Google Scholar] [CrossRef]
- Grimble, R.F. Effect of antioxidative vitamins on immune function with clinical applications. Int. J. Vitam. Nutr. Res. 1997, 67, 312–320. [Google Scholar]
- Gebhardt, B.M.; Newberne, P.M. Nutrition and immunological responsiveness. T-cell function in the offspring of lipotrope and protein-deficient rats. Immunology 1974, 26, 489–495. [Google Scholar]
- Grimble, R.F. Malnutrition and the immune response. 2. Impact of nutrients on cytokine biology in infection. Trans. R. Soc. Trop. Med. Hyg. 1994, 88, 615–619. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Partearroyo, T.; Úbeda, N.; Montero, A.; Achón, M.; Varela-Moreiras, G. Vitamin B12 and Folic Acid Imbalance Modifies NK Cytotoxicity, Lymphocytes B and Lymphoprolipheration in Aged Rats. Nutrients 2013, 5, 4836-4848. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5124836
Partearroyo T, Úbeda N, Montero A, Achón M, Varela-Moreiras G. Vitamin B12 and Folic Acid Imbalance Modifies NK Cytotoxicity, Lymphocytes B and Lymphoprolipheration in Aged Rats. Nutrients. 2013; 5(12):4836-4848. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5124836
Chicago/Turabian StylePartearroyo, Teresa, Natalia Úbeda, Ana Montero, María Achón, and Gregorio Varela-Moreiras. 2013. "Vitamin B12 and Folic Acid Imbalance Modifies NK Cytotoxicity, Lymphocytes B and Lymphoprolipheration in Aged Rats" Nutrients 5, no. 12: 4836-4848. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5124836