Effects of Dietary Carbohydrate Replaced with Wild Rice (Zizania latifolia (Griseb) Turcz) on Insulin Resistance in Rats Fed with a High-Fat/Cholesterol Diet

Abstract
:1. Introduction
2. Experimental Section
2.1. Animals and Diets
2.2. Tissue Collection
2.3. Biochemical Analysis
2.4. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis

{kind=link}
{kind=link}
{kind=link}
| Primer | Sense (5′–3′) | Antisense (5′–3′) | Size (bp) |
|---|---|---|---|
| β-actin | GAAATCGTGCGTGACATTAAG | GCTAGAAGCATTTGCGGTGGA | 511 |
| AdipoR2 | TGCGCACACGTTTTCAGTCTCCT | TTCTATGATCCCCAAAAGTGTGC | 150 |
| PPAR-α | AGGCTATCCCAGGCTTTGC | GCGTCTGACTCGGTCTTCTTG | 487 |
| PPAR-γ | TCCGTGATGGAAGACCACTC | CCCTTGCATCCTTCACAAGC | 532 |
| Leptin | CCCATTCTGAGTTTGTCCA | GCATTCAGGGCTAAGGTC | 301 |
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
3.1. Food Intake, Weight Gain, Relative Liver Weights and Liver Lipids
| LF | HFC | CD | WR | F | p | |
|---|---|---|---|---|---|---|
| Food intake (g/day) | 13.63 ± 0.07 | 13.63 ± 0.10 | 13.63 ± 0.09 | 13.62 ± 0.10 | 0.11 | 0.954 |
| Weight gain (g/8 weeks) | 205.71 ± 19.98 a | 240.54 ± 15.93 b | 232.33 ± 20.15 b | 214.05 ± 20.39 a | 5.80 | 0.002 |
| Relative liver weight (g/100 g) | 2.56 ± 0.29 a | 4.48 ± 0.42 b | 4.27 ± 0.50 b | 2.89 ± 0.65 a | 39.27 | <0.001 |
| Liver lipids contents | ||||||
| Triglyceride (μmol/gprot) | 0.66 ± 0.28 a | 1.22 ± 0.19 b | 1.19 ± 0.13 b | 0.73 ± 0.19 a | 21.57 | <0.001 |
| Free fatty acid (μmol/gprot) | 40.75 ± 4.90 a | 77.70 ± 10.25 c | 69.28 ± 9.83 b | 41.35 ± 7.44 a | 51.63 | <0.001 |
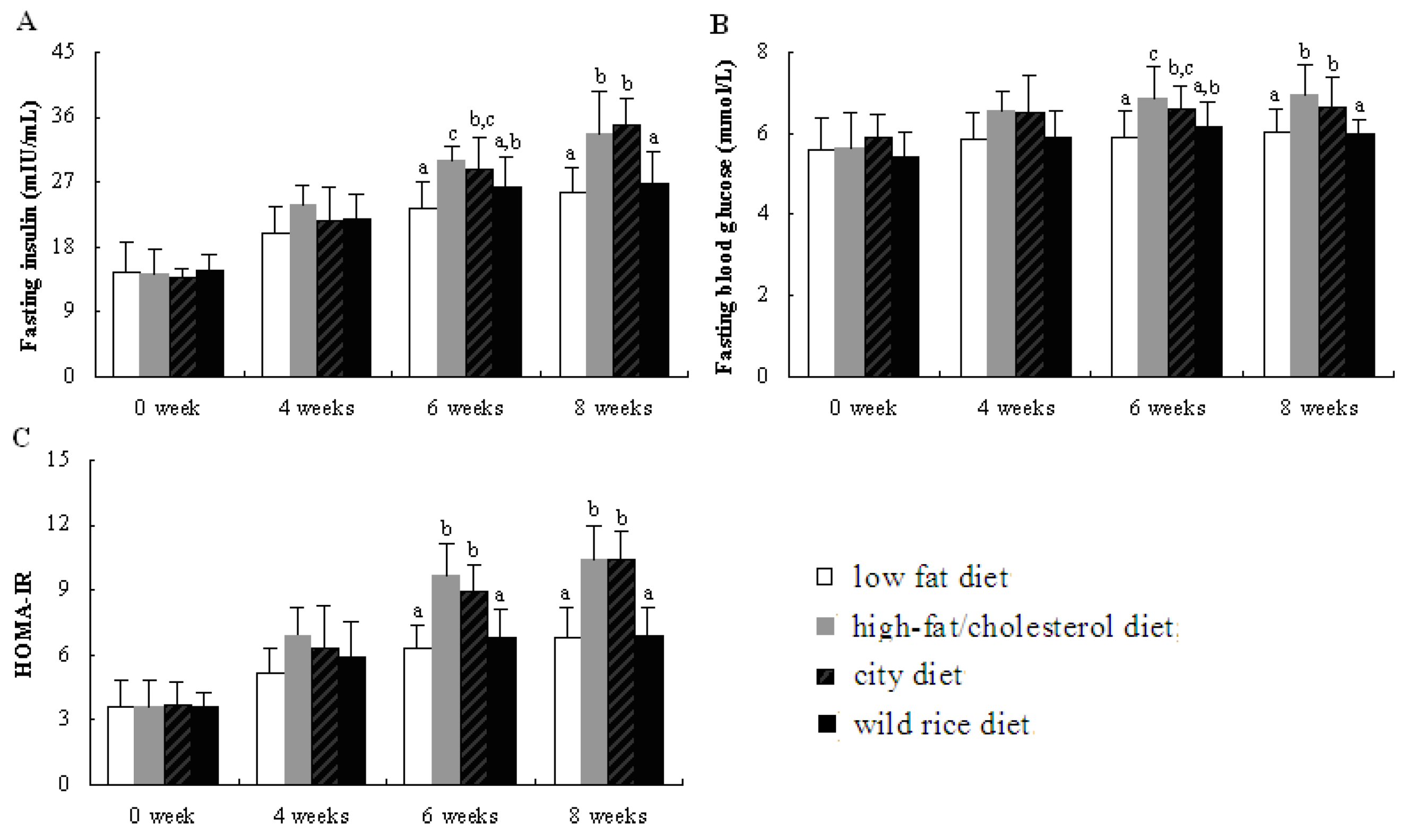
3.2. Fasting Blood Glucose and Insulin Levels, HOMA-IR Index

3.3. Serum Adiponectin, Lipocalin-2 and Visfatin Levels
| LF | HFC | CD | WR | F | p | |
|---|---|---|---|---|---|---|
| Adiponectin (ng/mL) | 5.44 ± 0.82 b | 2.92 ± 0.65 a | 3.14 ± 0.74 a | 5.05 ± 0.61 b | 32.74 | <0.001 |
| Lipocalin-2 (pg/mL) | 75.09 ± 10.98 a | 96.95 ± 13.38 b | 92.83 ± 9.98 b | 80.70 ± 8.49 a | 8.85 | <0.001 |
| Visfatin (ng/L) | 32.42 ± 4.24 a | 54.23 ± 5.14 c | 49.97 ± 6.19 c | 38.92 ± 4.25 b | 39.70 | <0.001 |
3.4. Gene Expression of AdipoR2, PPAR-а, PPAR-γ and Leptin in Liver or Epididymal Tissues

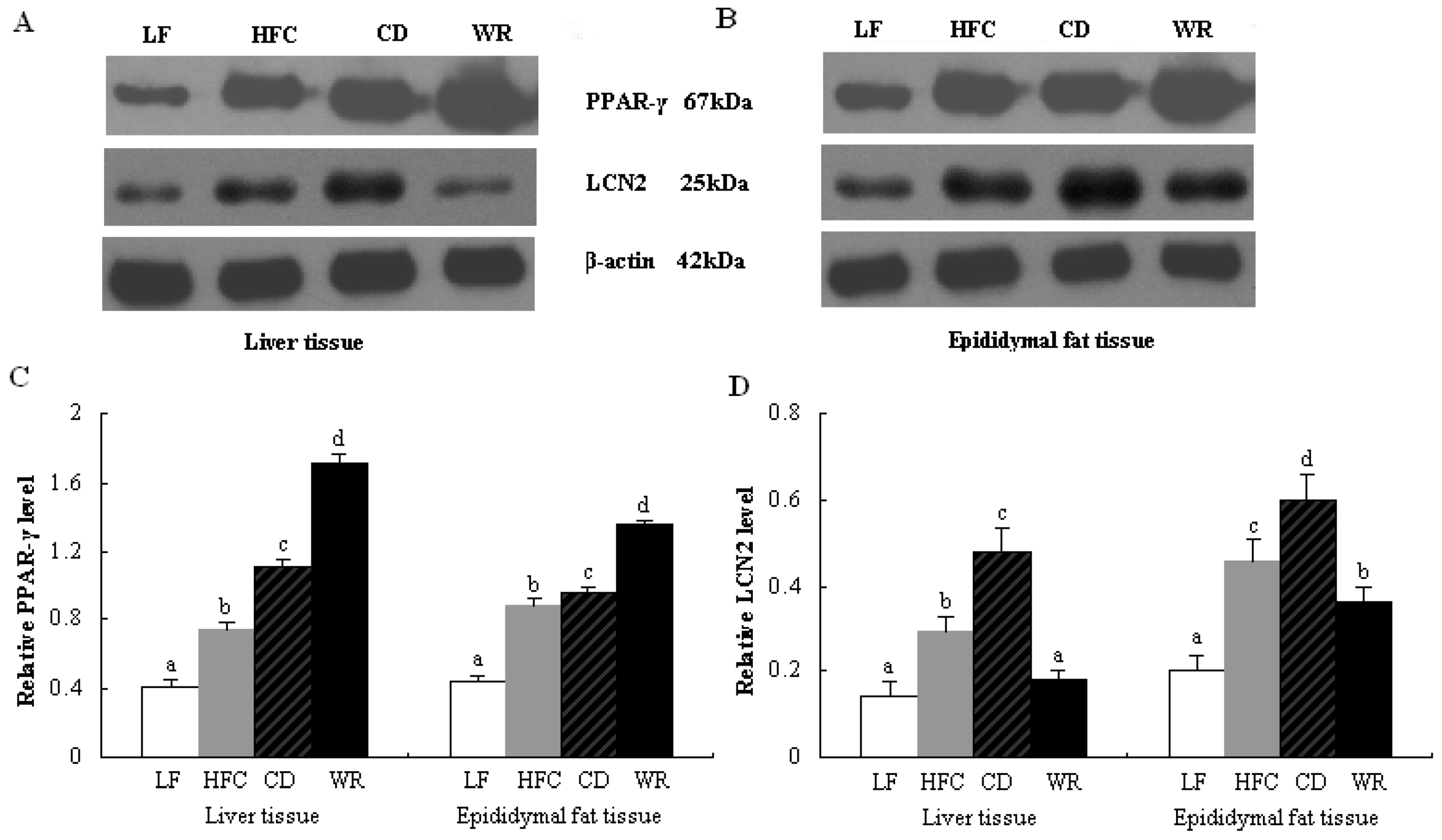
3.5. Protein Expression of PPAR-γ and LCN2 in Liver and Epididymal Tissues

4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Lü, J.; Li, L.M. Diet and nutritional status related to cardiovascular diseases risks in contemporary China. CVD Prev. Control. 2009, 4, 51–59. [Google Scholar]
- Popkin, B.M.; Adair, L.S.; Ng, S.W. Global nutrition transition and the pandemic of obesity in developing countries. Nutr. Rev. 2012, 70, 3–21. [Google Scholar] [CrossRef]
- Alberti, K.G.; Zimmet, P.; Shaw, J. Metabolic syndrome—a new world-wide definition. A consensus statement from the international diabetes federation. Diabet. Med. 2006, 23, 469–480. [Google Scholar] [CrossRef]
- Chavez, J.A.; Summers, S.A. Lipid oversupply, selective insulin resistance, and lipotoxicity. Biochim. Biophys.Acta 2010, 1801, 252–265. [Google Scholar] [CrossRef]
- Giacco, R.; Pepa, G.D.; Luongo, D.; Riccardi, G. Whole grain intake in relation to body weight: From epidemiological evidence to clinical trials. Nutr. Metab. Cardiovas. 2011, 21, 901–908. [Google Scholar] [CrossRef]
- Lutsey, P.L.; Jacobs, D.R., Jr.; Kori, S.; Mayer-Davis, E.; Shea, S.; Steffen, L.M.; Szklo, M.; Trazcy, R. Whole grain intake and its cross-sectional association with obesity, insulin resistance, inflammation, diabetes and subclinical CVD: The MESA Study. Br. J. Nutr. 2007, 98, 397–405. [Google Scholar] [CrossRef]
- Harris, K.A.; Kris-Etherton, P.M. Effects of whole grain on coronary heart disease risk. Curr. Atheroscler. Rep. 2010, 12, 368–376. [Google Scholar] [CrossRef]
- Liu, R.H. Whole grain phytochemicals and health. J. Cereal. Sci. 2007, 46, 207–219. [Google Scholar] [CrossRef]
- Fardet, A. New hypothesis for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef]
- Food and Drug Administration. Available online: http://www.fda.gov/Food/GuidanceComplianceregulatoryInformation/GuidanceDocuments/FoodLabelingNutrition/UCM059088 (accessed on 10 January 2010).
- Geng, Y.L. Diagram of Chinese Staple Plants; Science Publishing House: Beijing, China, 1965; pp. 534–635. [Google Scholar]
- Kim, J.M.; Lorenz, K. Enzymatic activities in wild rice (Zizania aquatica). Food Sci. Technol. 1981, 14, 23–27. [Google Scholar]
- Zhai, C.K.; Lu, C.M.; Zhang, X.Q.; Sun, G.J.; Lorenz, K.J. Comparative study on nutritional value of Chinese and North American wild rice. J. Food Compos. Anal. 2001, 4, 371–382. [Google Scholar]
- Zhai, C.K.; Tang, W.L.; Jiang, X.L. Protein and amino acid composition of Chinese and North American wild rice. J. Food Sci. 1994, 27, 380–383. [Google Scholar]
- Oelke, E.A. Amino and content in wild rice (Zizania palustris L.) grain. Agron. J. 1976, 68, 146–148. [Google Scholar]
- Zhang, H.; Cao, P.; Agellon, L.B.; Zhai, C.K. Wild rice (Zizania latifolia (Griseb) Turcz) improves the serum lipid profile and antioxidant status of rats fed with a high fat/cholesterol diet. Br. J. Nutr. 2009, 102, 1723–1727. [Google Scholar] [CrossRef]
- Azgar, S. Peroxisome proliferator-activated receptors, metabolic syndrome and cardiovascular disease. Future Cardiol. 2010, 6, 657–691. [Google Scholar] [CrossRef]
- Pittas, A.G.; Joseph, N.A.; Greenberg, A.S. Adipocytokines and insulin resistance. J. Clin. Endocrinol. Metab. 2004, 89, 447–452. [Google Scholar] [CrossRef]
- Han, S.F.; Zhang, H.; Zhai, C.K. Protective potentials of wild rice (Zizania latifolia (Griseb) Turcz) against obesity and lipotoxicity induced by a high-fat/cholesterol diet in rats. Food Chem. Toxicol. 2012, 50, 2263–2269. [Google Scholar] [CrossRef]
- Song, G.Y.; Gao, Y.; Wang, C.; Hu, S.G.; Wang, J.; Qu, D.M.; Ma, H.J. Rosiglitazone reduces fatty acid translocase and increases AMPK in skeletal muscle in aged rats: A possible mechanism to prevent high-fat-induced insulin resistance. Chin. Med. J. 2010, 123, 2384–2391. [Google Scholar]
- Zhang, W.; Liu, C.Q.; Wang, P.W.; Sun, S.Y.; Su, W.J.; Zhang, H.J.; Li, X.J.; Yang, S.Y. Puerarin improves insulin resistance and modulates adipokine expression in rats fed a high-fat diet. Eur. J. Pharmacol. 2010, 649, 398–402. [Google Scholar] [CrossRef]
- He, Y.L.; Zhai, F.Y.; Wang, H.J.; Hu, Y.S.; Yang, X.G. The status and trend for dietary pattern of energy, protein and fat in Chinese residents. Acta Nutr. Sin. 2005, 27, 358–365. [Google Scholar]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- de Kok, T.M.; van Breda, S.G.; Manson, M.M. Mechanisms of combined action of different chemopreventive dietary compounds: A review. Eur. J. Nutr. 2008, 47, S51–S59. [Google Scholar] [CrossRef]
- Truswell, A.S. Cereal grains and coronary heart disease. Eur. J. Clin. Nutr. 2002, 56, 1–14. [Google Scholar] [CrossRef]
- Liu, S. Intake of refined carbohydrates and whole grain foods in relation to risk of type 2 diabetes mellitus and coronary heart disease. J. Am. Coll. Nutr. 2002, 21, 298–306. [Google Scholar] [CrossRef]
- Slavin, J.L. Dietary fiber and body weight. Nutrition 2005, 21, 411–418. [Google Scholar] [CrossRef]
- McCarty, M.F. Magnesium may mediate the favorable impact of whole grains on insulin sensitivity by acting as a mild calcium antagonist. Med. Hypotheses. 2005, 64, 619–627. [Google Scholar] [CrossRef]
- Dong, J.Y.; He, K.; Xun, P.C.; Qin, L.Q. Magnesium intake and risk of type 2 diabetes. Diabetes Care 2011, 34, 2116–2122. [Google Scholar] [CrossRef]
- Przybylski, P.; Klensporf-Pawlik, D.; Anwar, F.; Rudzinska, M. Lipid components of North American wild rice. J. Am. Oil. Chem. Soc. 2009, 86, 553–559. [Google Scholar] [CrossRef]
- Qiu, Y.; Liu, Q.; Beta, T. Antioxidant properties of commercial wild rice and analysis of soluble and insoluble phenolic acids. Food Chem. 2010, 121, 140–147. [Google Scholar]
- Odegaard, J.I.; Ricardo-Gonzalez, R.R.; Goforth, M.H.; Morel, C.R.; Subramanian, V.; Mukundan, L.; Eagle, A.R.; Vats, D.; Brombacher, F.; Ferrante, A.W.; Chawla, A. Macrophage-specific PPAR gamma controls alternative activation and improves insulin resistance. Nature 2007, 447, 1116–1120. [Google Scholar]
- Szanto, A.; Nagy, L. The many faces of PPARγ: Anti-inflammatory by any means. Immunobiology 2008, 213, 789–803. [Google Scholar] [CrossRef]
- Chou, C.J.; Haluzik, M.; Gregory, C.; Dietz, K.R.; Vinson, C.; Gavrilova, O.; Reitman, M.L. WY14,643, a peroxisome proliferator-activated receptor α (PPARα) agonist, improves hepatic and muscle steatosis and reverses insulin resistance in lipoatrophic A-ZIP/F-1 mice. J. Biol. Chem. 2002, 277, 24484–24489. [Google Scholar]
- Inoue, M.; Ohtake, T.; Motomura, W.; Takahashi, N.; Hosoki, Y.; Miyoshi, S.; Suzuki, Y.; Saito, H.; Kohgo, Y.; Okumura, T. Increased expression of PPARγ in high fat diet-induced liver steatosis in mice. Biochem. Biophys. Res. Commun. 2005, 336, 215–222. [Google Scholar] [CrossRef]
- Lehrke, M.; Lazar, M.A. The many faces of PPARγ. Cell 2005, 123, 993–999. [Google Scholar] [CrossRef]
- Haluzík, M.M.; Haluzík, M. PPAR-alpha and insulin sensitivity. Physiol. Res. 2006, 55, 115–122. [Google Scholar]
- Civitarese, A.E.; Jenkinson, C.P.; Richardson, D.; Bajaj, M.; Cusi, K.; Kashyap, S.; Berria, R.; Belfort, R.; DeFronzo, R.A.; Mandarino, L.J.; Ravussin, E. Adiponectin receptors gene expression and insulin sensitivity in non-diabetic Mexican Americans with or without a family history of Type 2 diabetes. Diabetologia 2004, 47, 816–820. [Google Scholar] [CrossRef]
- Wang, Y.; Lam, K.; Kraegen, E.W.; Sweeney, G.; Zhang, J.; Tso, A.; Chow, W.S.; Wat, N.; Xu, J.Y.; Hoo, R.; Xu, A. Lipocalin-2 is an inflammatory maker closely associated with obesity, insulin resistance, and hyperglycemia in humans. Clin. Chem. 2007, 53, 34–41. [Google Scholar]
- Zou, C.H.; Shao, J.H. Role of adipocytokines in obesity-associated insulin resistance. J. Nutr. Biochem. 2008, 19, 277–286. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Han, S.; Zhang, H.; Qin, L.; Zhai, C. Effects of Dietary Carbohydrate Replaced with Wild Rice (Zizania latifolia (Griseb) Turcz) on Insulin Resistance in Rats Fed with a High-Fat/Cholesterol Diet. Nutrients 2013, 5, 552-564. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5020552
Han S, Zhang H, Qin L, Zhai C. Effects of Dietary Carbohydrate Replaced with Wild Rice (Zizania latifolia (Griseb) Turcz) on Insulin Resistance in Rats Fed with a High-Fat/Cholesterol Diet. Nutrients. 2013; 5(2):552-564. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5020552
Chicago/Turabian StyleHan, Shufen, Hong Zhang, Liqiang Qin, and Chengkai Zhai. 2013. "Effects of Dietary Carbohydrate Replaced with Wild Rice (Zizania latifolia (Griseb) Turcz) on Insulin Resistance in Rats Fed with a High-Fat/Cholesterol Diet" Nutrients 5, no. 2: 552-564. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5020552