Cocoa Flavonoid-Enriched Diet Modulates Systemic and Intestinal Immunoglobulin Synthesis in Adult Lewis Rats



 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals and Diets

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | REF (AIN-93M; g/kg) | PC0.2 (g/kg) | PC0.4 (g/kg) | PC0.8 (g/kg) |
|---|---|---|---|---|
| Cocoa powder | - | 100 | 8.7 | 17.4 |
| Nutrients provided by the basal mix: | ||||
| Casein | 121.8 | 97.1 | 120.7 | 119.5 |
| l-Cystine | 1.8 | 1.4 | 1.8 | 1.8 |
| Corn Starch | 419.1 | 421.0 | 418.3 | 417.5 |
| Maltodextrin | 147.3 | 116.8 | 147.3 | 147.3 |
| Sucrose | 117.0 | 108.7 | 117.0 | 117.0 |
| Soybean oil | 40.0 | 26.2 | 39.4 | 38.9 |
| Cellulose | 50.0 | 24.5 | 48.3 | 46.5 |
| Minerals | 27.0 | 27.8 | 27.0 | 27.0 |
| Vitamins | 2.0 | 7.2 | 2.0 | 2.0 |
| Choline bitartrate | 2.5 | 2.0 | 2.5 | 2.5 |
| tert-Butylhydroquine | 0.008 | 0.006 | 0.008 | 0.008 |
| Nutrients provided by cocoa powder: | ||||
| Protein | - | 22 | 1.13 | 2.26 |
| Carbohydrate | - | 16 | 0.82 | 1.64 |
| Lipid | - | 11 | 0.56 | 1.12 |
| Fiber (insoluble/soluble) | - | 34 (25.5/8.5) | 1.75 (1.31/0.44) | 3.5 (2.63/0.88) |
| Total polyphenols * | - | 2.12 | 4.44 | 8.88 |
| Polyphenols provided by cocoa powder **: | ||||
| Catechin | - | 0.073 | 0.048 | 0.097 |
| Epicatechin | - | 0.204 | 0.689 | 1.379 |
| Isoquercetin | - | 0.0053 | n.d. | n.d. |
| Quercetin | - | 0.0029 | n.d. | n.d. |
| Procyanidin B1 | - | n.d. | 0.127 | 0.254 |
| Procyanidin B2 | - | 0.167 | 0.356 | 0.713 |
| Total procyanidins | - | n.d. | 3.897 | 7.795 |
2.2. Sample Collection and Processing
2.3. Fecal pH Determination
2.4. Immunoglobulin Quantification in Serum and Feces (ELISA)
2.5. Fecal Bacteria and IgA-Coating Bacterial Analysis
2.6. Statistical Analysis
3. Results
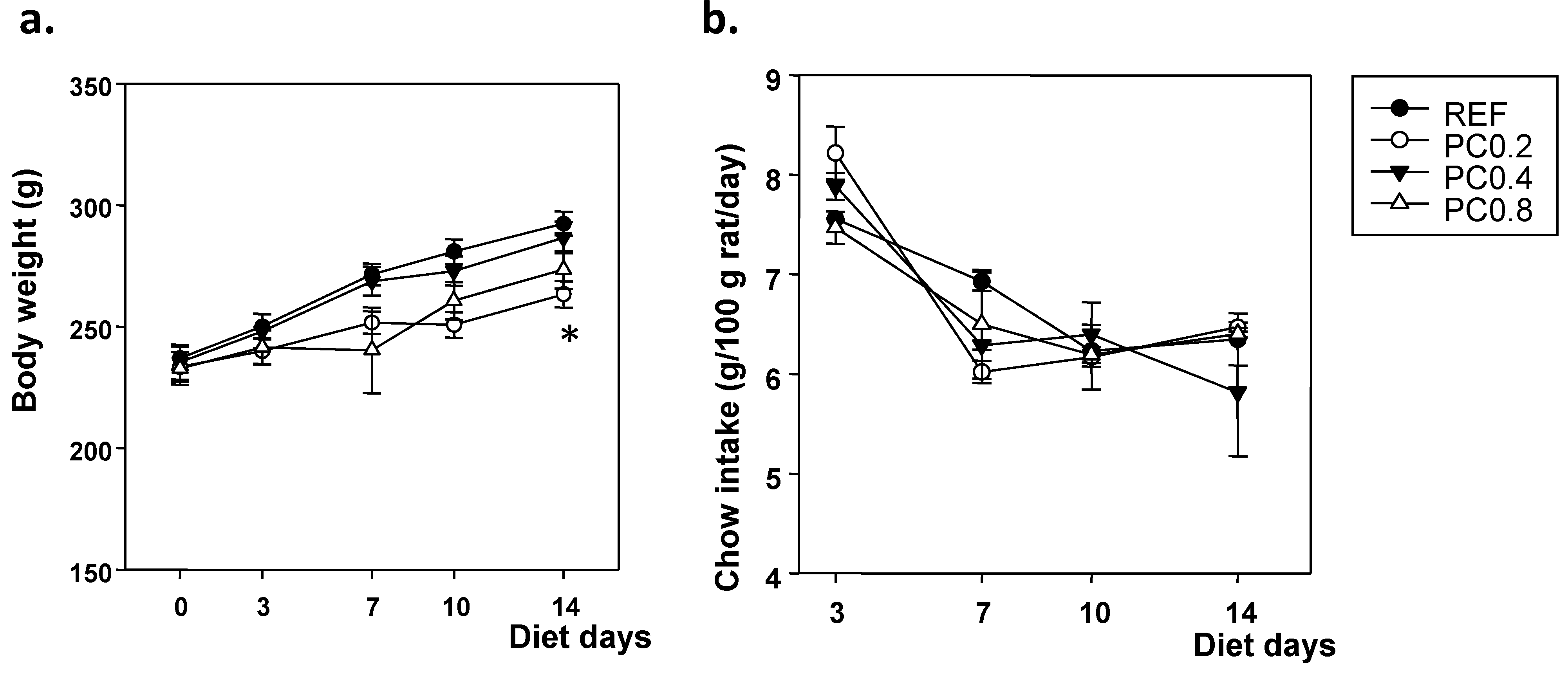
3.1. Body Weight and Chow Intake

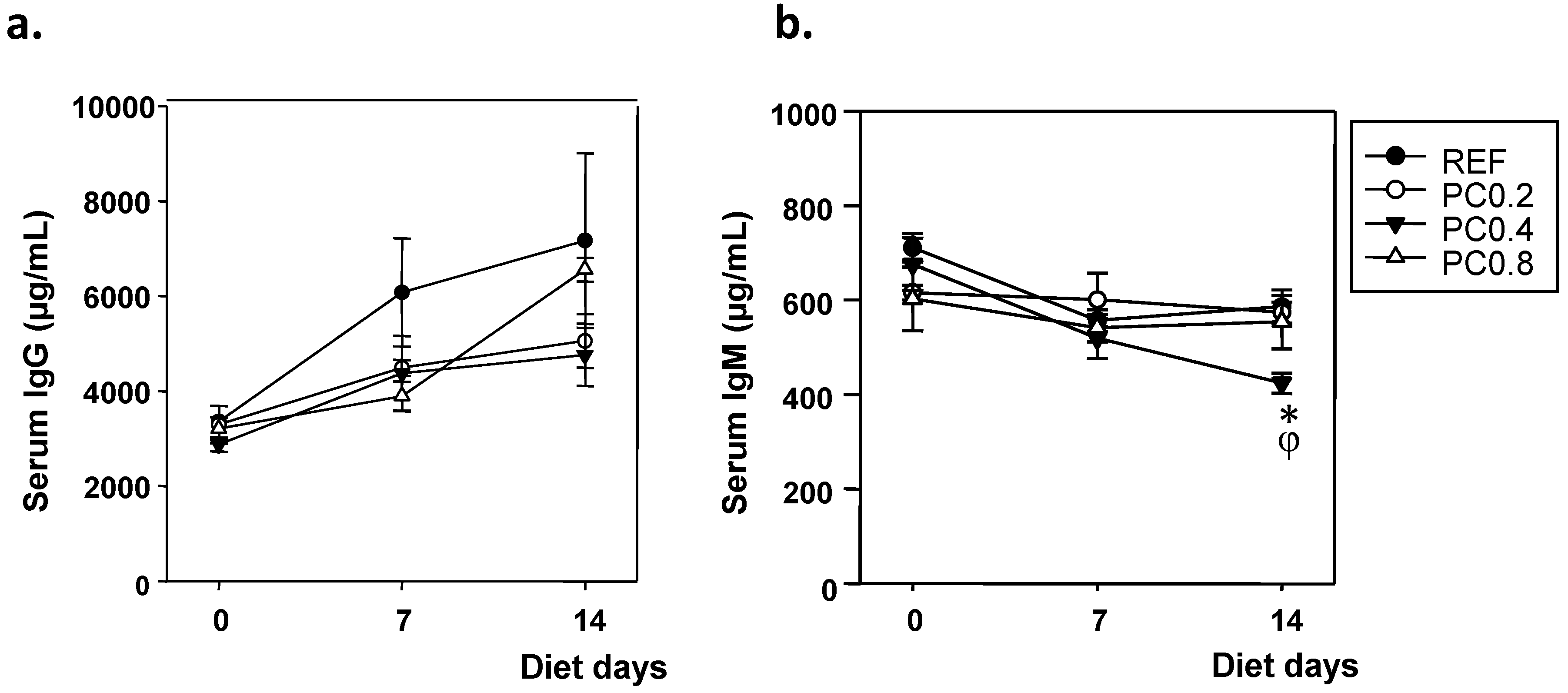
3.2. Serum IgG and IgM Concentrations
3.3. Fecal IgA Concentration

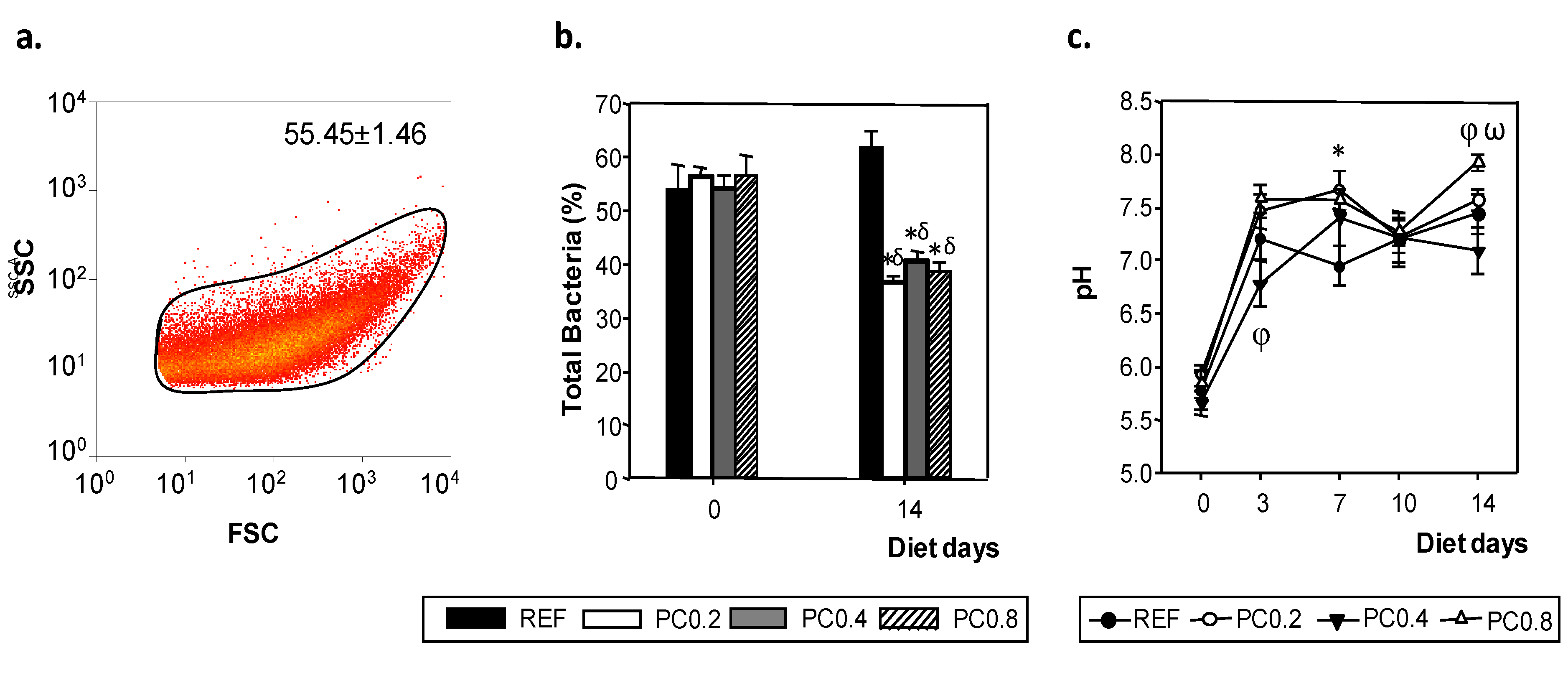
3.4. Total Bacteria Determination

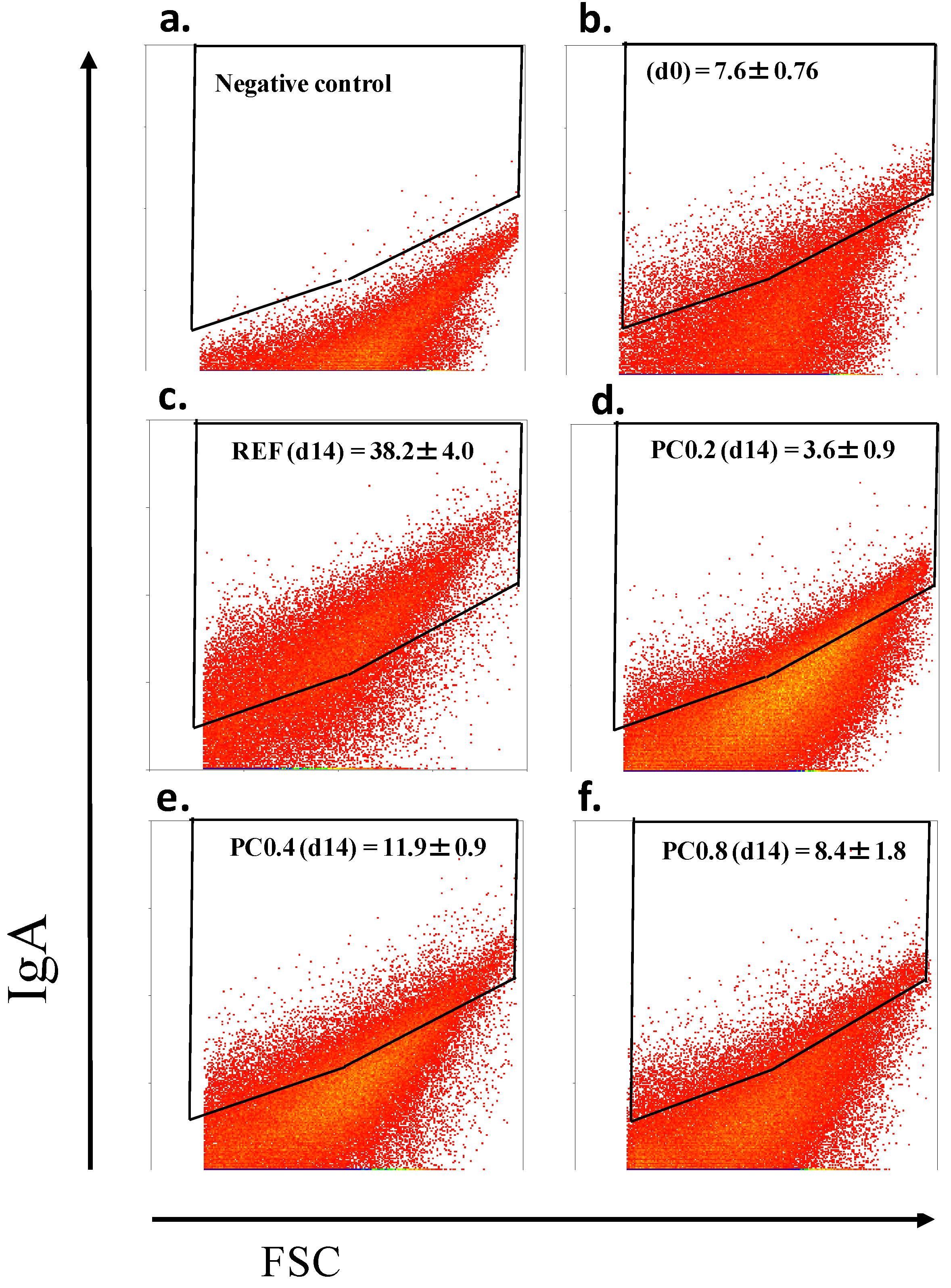
3.5. IgA-Coating Bacteria Analysis
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Henderson, J.S.; Joyce, R.A.; Hall, G.R.; Hurst, W.J.; McGovern, P.E. Chemical and archaeological evidence for the earliest cacao beverages. Proc. Natl. Acad. Sci. USA 2007, 104, 18937–18940. [Google Scholar]
- Dillinger, T.L.; Barriga, P.; Escárcega, S.; Jimenez, M.; Salazar, L.D.; Grivetti, L.E. Food of the Gods: Cure for humanity? A cultural history of the medicinal and ritual use of chocolate. J. Nutr. 2000, 130, S2057–S2072. [Google Scholar]
- Vinson, J.A.; Proch, J.; Zubik, L. Phenol antioxidant quantity and quality in foods: Cocoa, dark chocolate and milk chocolate. J. Agric. Food Chem. 1999, 47, 4821–4824. [Google Scholar] [CrossRef]
- Nishimoto, N.; Kishimoto, T.; Yoshizaki, K. Anti-interleukin 6 receptor antibody treatment in rheumatic disease. Ann. Rheum. Dis. 2000, 59, i21–i27. [Google Scholar] [CrossRef]
- Hammerstone, J.F.; Lazarus, S.A.; Mitchell, A.E.; Rucker, R.; Schmitz, H.H. Identification of procyanidins in cocoa (Theobroma cacao) and chocolate using high-performance liquid chromatography/mass spectrometry. J. Agric. Food Chem. 1999, 47, 490–496. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Lee, K.W.; Kim, Y.J.; Lee, H.J.; Lee, C.Y. Cocoa has more phenolic phytochemicals and a higher antioxidant capacity than teas and red wine. J. Agric. Food Chem. 2003, 51, 7292–7295. [Google Scholar] [CrossRef]
- Visioli, F.; Bernaert, H.; Corti, R.; Ferri, C.; Heptinstall, S.; Molinari, E.; Poli, A.; Serafini, M.; Smit, H.J.; Vinson, J.A.; et al. Chocolate, lifestyle, and health. Crit. Rev. Food Sci. Nutr. 2009, 49, 299–312. [Google Scholar] [CrossRef]
- Gu, L.; House, S.E.; Wu, X.; Ou, B.; Prior, R.L. Procyanidin and catechin contents and antioxidant capacity of cocoa and chocolate products. J. Agric. Food Chem. 2006, 54, 4057–4061. [Google Scholar]
- Andrés-Lacueva, C.; Monagas, M.; Khan, N.; Izquierdo-Pulido, M.; Urpi-Sarda, M.; Permanyer, J.; Lamuela-Raventós, R.M. Flavanol and flavonol contents of cocoa powder products: Influence of the manufacturing process. J. Agric. Food Chem. 2008, 56, 3111–3117. [Google Scholar] [CrossRef]
- Wollgast, J.; Anklam, E. Review on polyphenols in Theobroma cacao: Changes in composition during the manufacture of chocolate and methodology for identification and quantification. Food Res. Int. 2000, 33, 423–447. [Google Scholar] [CrossRef]
- Sánchez-Rabaneda, F.; Jáuregui, O.; Casals, I.; Andrés-Lacueva, C.; Izquierdo-Pulido, M.; Lamuela-Raventós, R.M. Liquid chromatographic/electrospray ionization tandem mass spectrometric study of the phenolic composition of cocoa (Theobroma cacao). J. Mass Spectrom. 2003, 38, 35–42. [Google Scholar] [CrossRef]
- Lazarus, S.A.; Hammerstone, J.F.; Schmitz, H.H. Chocolate contains additional flavonoids not found in tea. Lancet 1999, 354, 1825. [Google Scholar]
- Ramiro-Puig, E.; Casadesús, G.; Lee, H.G.; Zhu, X.; McShea, A.; Perry, G.; Pérez-Cano, F.J.; Smith, M.A.; Castell, M. Neuroprotective effect of cocoa flavonoids on in vitro oxidative stress. Eur. J. Nutr. 2009, 48, 54–61. [Google Scholar] [CrossRef]
- Martín, M.A.; Granado-Serrano, A.B.; Ramos, S.; Pulido, M.I.; Bravo, L.; Goya, L. Cocoa flavonoids up-regulate antioxidant enzyme activity via the ERK1/2 pathway to protect against oxidative stress-induced apoptosis in HepG2 cells. J. Nutr. Biochem. 2010, 21, 196–205. [Google Scholar] [CrossRef]
- Kenny, T.P.; Shu, S.A.; Moritoki, Y.; Keen, C.L.; Gershwin, M.E. Cocoa flavanols and procyanidins can modulate the lipopolysaccharide activation of polymorphonuclear cells in vitro. J. Med. Food 2009, 12, 1–7. [Google Scholar] [CrossRef]
- Ramiro-Puig, E.; Pérez-Cano, F.J.; Ramírez-Santana, C.; Castellote, C.; Izquierdo-Pulido, M.; Permanyer, J.; Franch, A.; Castell, M. Spleen lymphocyte function modulated by a cocoa-enriched diet. Clin. Exp. Immunol. 2007, 149, 535–542. [Google Scholar] [CrossRef]
- Pérez-Berezo, T.; Franch, A.; Castellote, C.; Castell, M.; Pérez-Cano, F.J. Mechanisms involved in down-regulation of intestinal IgA in rats by high cocoa. J. Nutr. Biochem. 2012, 23, 838–844. [Google Scholar] [CrossRef]
- Massot-Cladera, M.; Pérez-Berezo, T.; Franch, A.; Castell, M.; Pérez-Cano, F.J. Cocoa modulatory effect on rat fecal microbiota and colonic crosstalk. Arch. Biochem. Biophys. 2012, 527, 105–112. [Google Scholar] [CrossRef]
- Ramiro-Puig, E.; Pérez-Cano, F.J.; Ramos-Romero, S.; Pérez-Berezo, T.; Castellote, C.; Permanyer, J.; Franch, A.; Izquierdo-Pulido, M.; Castell, M. Intestinal immune system of young rats influenced by cocoa-enriched diet. J. Nutr. Biochem. 2008, 19, 555–565. [Google Scholar] [CrossRef]
- Pérez-Berezo, T.; Ramiro-Puig, E.; Pérez-Cano, F.J.; Castellote, C.; Permanyer, J.; Franch, A.; Castell, M. Influence of a cocoa-enriched diet on specific immune response in ovalbumin-sensitized rats. Mol. Nutr. Food Res. 2009, 53, 389–397. [Google Scholar] [CrossRef]
- Ramos-Romero, S.; Pérez-Cano, F.J.; Castellote, C.; Castell, M.; Franch, A. Effect of cocoa-enriched diets on lymphocytes involved in adjuvant arthritis in rats. Br. J. Nutr. 2012, 107, 378–387. [Google Scholar] [CrossRef]
- Niemenak, N.; Rohsius, C.; Elwers, S.; Ndoumou, D.O.; Lieberei, R. Comparative study of different cocoa (Theobroma cacao L.) clones in terms of their phenolics and anthocyanins contents. J. Food Compos. Anal. 2006, 19, 612–619. [Google Scholar] [CrossRef]
- Tomas-Barberán, F.A.; Cienfuegos-Jovellanos, E.; Marín, A.; Muguerza, B.; Gil-Izquierdo, A.; Cerda, B.; Zafrilla, P.; Morillas, J.; Mulero, J.; Ibarra, A.; et al. A new process to develop a cocoa poder with higher flavonoid monomer content and enhanced bioavailability in healthy humans. J. Agric. Food Chem. 2007, 55, 3926–3935. [Google Scholar] [CrossRef]
- Cienfuentes-Jovellanos, E.; Pasamar, M.A.; Fritz, J.; Arcos, J.; Ramón, D.; Castilla, Y. Method for Obtaining Polyphenol-Rich Cocoa Powder with a Low Fat Content and Cocoa thus Obtained. Patent Cooperation Treaty (PCT) WO 2007/096449A1, 2007. [Google Scholar]
- Serra Bonvehí, J.; Ventura Coll, F. Evaluation of bitterness and astringency of polyphenolic compounds in cocoa powder. Food Chem. 1997, 60, 365–370. [Google Scholar] [CrossRef]
- Cienfuegos-Jovellanos, E.; Quiñones Mdel, M.; Muguerza, B.; Moulay, L.; Miguel, M.; Aleixandre, A. Antihypertensive effect of a polyphenol-rich cocoa poder industrially processed to preserve the original flavonoids of the cocoa beans. J. Agric. Food Chem. 2009, 57, 6156–6162. [Google Scholar] [CrossRef]
- Pérez-Berezo, T.; Franch, A.; Ramos-Romero, S.; Castellote, C.; Pérez-Cano, F.J.; Castell, M. Cocoa-enriched diets modulate intestinal and systemic humoral immune response in young adult rats. Mol. Nutr. Food Res. 2011, 55, S56–S66. [Google Scholar] [CrossRef]
- Matsui, N.; Ito, R.; Nishimura, E.; Yoshikawa, M.; Kato, M.; Kamei, M.; Shibata, H.; Matsumoto, I.; Abe, K.; Hashizume, S. Ingested cocoa can prevent high-fat diet-induced obesity by regulating the expression of genes for fatty acid metabolism. Nutrition 2005, 21, 594–601. [Google Scholar] [CrossRef]
- Nogueira, L.; Ramirez-Sanchez, I.; Perkins, G.A.; Murphy, A.; Taub, P.R.; Ceballos, G.; Villarreal, F.J.; Hogan, M.C.; Malek, M.H. (–)-Epicatechin enhances fatigue resistance and oxidative capacity in mouse muscle. J. Physiol. 2011, 589, 4615–4631. [Google Scholar]
- Gu, Y.; Hurst, W.J.; Stuart, D.A.; Lambert, J.D. Inhibition of key digestive enzymes by cocoa extracts and procyanidins. J. Agric. Food Chem. 2011, 59, 5305–5311. [Google Scholar] [CrossRef]
- Gu, Y.; Yu, S.; Lambert, J.D. Dietary cocoa ameliorates obesity-related inflammation in high fat-fed mice. Eur. J. Nutr. 2013, 15. [Google Scholar] [CrossRef]
- Min, S.Y.; Yang, H.; Seo, S.G.; Shin, S.H.; Chung, M.Y.; Kim, J.; Lee, S.J.; Lee, K.W. Cocoa polyphenols suppress adipogenesis in vitro and obesity in vivo by targeting insulin receptor. Int. J. Obes. (Lond.) 2013, 37, 584–592. [Google Scholar] [CrossRef]
- Lin, J.K.; Lin-Shiau, S.Y. Mechanisms of hypolipidemic and anti-obesity effects of tea and tea polyphenols. Mol. Nutr. Food Res. 2006, 50, 211–217. [Google Scholar] [CrossRef]
- Cherniack, E.P. Polyphenols: Planting the seeds of treatment for the metabolic syndrome. Nutrition 2011, 27, 617–623. [Google Scholar] [CrossRef]
- Shin, S.K.; Ha, T.Y.; McGregor, R.A.; Choi, M.S. Long-term curcumin administration protects against atherosclerosis via hepatic regulation of lipoprotein cholesterol metabolism. Mol. Nutr. Food Res. 2011, 55, 1829–1840. [Google Scholar] [CrossRef]
- Ramos, S.; Moulay, L.; Granado-Serrano, A.B.; Vilanova, O.; Muguerza, B.; Goya, L.; Bravo, L. Hypolipidemic effect in cholesterol-fed rats of a soluble fiber-rich product obtained from cocoa husks. J. Agric. Food Chem. 2008, 56, 6985–6993. [Google Scholar]
- Tumbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef]
- Tagliabue, A.; Elli, M. The role of gut microbiota in human obesity: Recent findings and future perspectives. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 160–168. [Google Scholar] [CrossRef]
- Sanz, Y.; Rastmanesh, R.; Agostonic, C. Understanding the role of gut microbes and probiotics in obesity: How far are we? Pharmacol. Res. 2013, 69, 144–155. [Google Scholar] [CrossRef]
- Shen, J.; Obin, M.S.; Zhao, L. The gut microbiota, obesity and insulin resistance. Mol. Aspects Med. 2013, 34, 39–58. [Google Scholar] [CrossRef]
- Puupponen-Pimiä, R.; Nohynek, L.; Hartmann-Schmidlin, S.; Kähkönen, M.; Heinonen, M.; Määttä-Riihinen, K.; Oksman-Caldentey, K.M. Berry phenolics selectively inhibit the growth of intestinal pathogens. J. Appl. Microbiol. 2005, 98, 991–1000. [Google Scholar] [CrossRef]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolic and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Almajano, M.P.; Carbó, R.; Jiménez, J.A.L.; Gordon, M.H. Antioxidant and antimicrobial activities of tea infusions. Food Chem. 2008, 108, 55–63. [Google Scholar] [CrossRef]
- Dolara, P.; Luceri, C.; de Filippo, C.; Femia, A.P.; Giovannelli, L.; Caderni, G.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Cresci, A. Red wine polyphenols influence carcinogenesis, intestinal microflora, oxidative damage and gene expression profiles of colonic mucosa in F344 rats. Mutat. Res. 2005, 591, 237–246. [Google Scholar] [CrossRef]
- Molan, A.L.; Liu, Z.; Tiwari, R. The ability of green tea to positively modulate key markers of gastrointestinal function in rats. Phytother. Res. 2010, 24, 1614–1619. [Google Scholar] [CrossRef]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2011, 93, 62–72. [Google Scholar] [CrossRef]
- Fogliano, V.; Corollaro, M.L.; Vitaglione, P.; Napolitano, A.; Ferracane, R.; Travaglia, F.; Arlorio, M.; Costabile, A.; Klinder, A.; Gibson, G. In vitro bioaccessibility and gut biotransformation of polyphenols present in the water-insoluble cocoa fraction. Mol. Nutr. Food Res. 2011, 55, S44–S55. [Google Scholar] [CrossRef]
- Kogiso, M.; Sakai, T.; Mitsuya, K.; Komatsu, T.; Yamamoto, S. Genistein suppresses anti-specific immune responses through competition with 17β-estradiol for estrogen receptors in ovalbumin immunized BALB/c mice. Nutrition 2006, 22, 802–809. [Google Scholar] [CrossRef]
- Das, M.; Ram, A.; Ghosh, B. Luteolin alleviated bronchoconstriction and airway hyperreactivity in ovalbumin sensitized mice. Inflamm. Res. 2003, 52, 101–106. [Google Scholar] [CrossRef]
- Yano, S.; Umeda, D.; Yamashita, S.; Yamada, K.; Tachibana, H. Dietary apigenin attenuates the development of atopic dermatitis-like skin lesions in NC/Nga mice. J. Nutr. Biochem. 2009, 20, 876–881. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Massot-Cladera, M.; Franch, À.; Castellote, C.; Castell, M.; Pérez-Cano, F.J. Cocoa Flavonoid-Enriched Diet Modulates Systemic and Intestinal Immunoglobulin Synthesis in Adult Lewis Rats. Nutrients 2013, 5, 3272-3286. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5083272
Massot-Cladera M, Franch À, Castellote C, Castell M, Pérez-Cano FJ. Cocoa Flavonoid-Enriched Diet Modulates Systemic and Intestinal Immunoglobulin Synthesis in Adult Lewis Rats. Nutrients. 2013; 5(8):3272-3286. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5083272
Chicago/Turabian StyleMassot-Cladera, Malen, Àngels Franch, Cristina Castellote, Margarida Castell, and Francisco J. Pérez-Cano. 2013. "Cocoa Flavonoid-Enriched Diet Modulates Systemic and Intestinal Immunoglobulin Synthesis in Adult Lewis Rats" Nutrients 5, no. 8: 3272-3286. https://0-doi-org.brum.beds.ac.uk/10.3390/nu5083272