Obesity as an Emerging Risk Factor for Iron Deficiency

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Pathophysiology of Obesity
3. The Physiology of Iron Metabolism
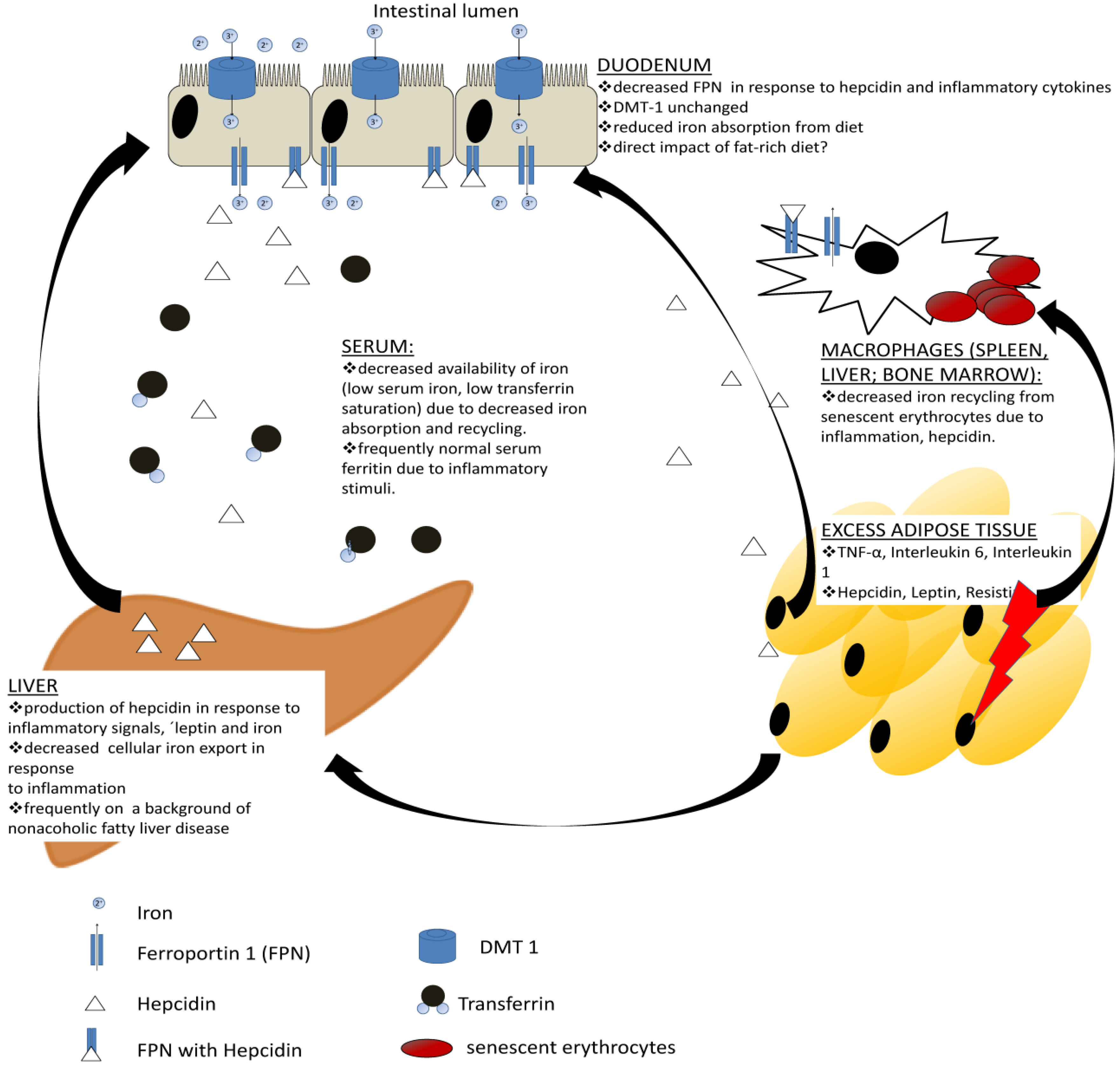
4. The Iron Phenotype of Obesity

5. Clinical Data on Obesity-Related Iron Deficiency—Adolescence
6. Adulthood
7. Response of Iron Parameters to Weight Reduction
8. Relevance of ID to the Course of Obesity
9. Bariatric Surgery and Iron Deficiency
10. Mechanisms Underlying Iron Deficiency in Obesity
11. Conclusions

Acknowledgements
Author Contributions
Conflict of Interest
References
- Low, S.; Chin, M.C.; Deurenberg-Yap, M. Review on epidemic of obesity. Ann. Acad. Med. Singapore 2009, 38, 57–59. [Google Scholar]
- Datz, C.; Felder, T.K.; Niederseer, D.; Aigner, E. Iron homeostasis in the metabolic syndrome. Eur. J. Clin. Investig. 2013, 43, 215–224. [Google Scholar]
- Lear, S.A.; Teo, K.; Gasevic, D.; Zhang, X.; Poirier, P.P.; Rangarajan, S.; Seron, P.; Kelishadi, R.; Tamil, A.M.; Kruger, A.; et al. The association between ownership of common household devices and obesity and diabetes in high, middle and low income countries. Can. Med. Assoc. J. 2014, 186, 258–266. [Google Scholar]
- Myint, P.K.; Kwok, C.S.; Luben, R.N.; Wareham, N.J.; Khaw, K.T. Body fat percentage, body mass index and waist-to-hip ratio as predictors of mortality and cardiovascular disease. Heart 2014. [Google Scholar] [CrossRef]
- Noto, H.; Goto, A.; Tsujimoto, T.; Osame, K.; Noda, M. Latest insights into the risk of cancer in diabetes. J. Diabetes Investig. 2013, 4, 225–232. [Google Scholar] [CrossRef]
- Dongiovanni, P.; Fracanzani, A.L.; Fargion, S.; Valenti, L. Iron in fatty liver and in the metabolic syndrome: A promising therapeutic target. J. Hepatol. 2011, 55, 920–932. [Google Scholar]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. 2006, 6, 772–783. [Google Scholar]
- Mather, K.J.; Goldberg, R.B. Clinical use of adiponectin as a marker of metabolic dysregulation. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 107–117. [Google Scholar] [CrossRef]
- DePaoli, A. Leptin in common obesity and associated disorders of metabolism. J. Endocrinol. 2014. [Google Scholar] [CrossRef]
- Flehmig, G.; Scholz, M.; Kloting, N.; Fasshauer, M.; Tonjes, A.; Stumvoll, M.; Youn, B.S.; Bluher, M. Identification of adipokine clusters related to parameters of fat mass, insulin sensitivity and inflammation. PLoS One 2014, 9, e99785. [Google Scholar]
- Wu, L.E.; Samocha-Bonet, D.; Whitworth, P.T.; Fazakerley, D.J.; Turner, N.; Biden, T.J.; James, D.E.; Cantley, J. Identification of fatty acid binding protein 4 as an adipokine that regulates insulin secretion during obesity. Mol. Metab. 2014, 3, 465–473. [Google Scholar]
- Cusi, K. The role of adipose tissue and lipotoxicity in the pathogenesis of type 2 diabetes. Curr. Diabetes Rep. 2010, 10, 306–315. [Google Scholar]
- Schwenger, K.J.; Allard, J.P. Clinical approaches to non-alcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 1712–1723. [Google Scholar]
- Hentze, M.W.; Muckenthaler, M.U.; Galy, B.; Camaschella, C. Two to tango: Regulation of mammalian iron metabolism. Cell 2010, 142, 24–38. [Google Scholar]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar]
- Abboud, S.; Haile, D.J. A novel mammalian iron-regulated protein involved in intracellular iron metabolism. J. Biol. Chem. 2000, 275, 19906–19912. [Google Scholar]
- Vulpe, C.D.; Kuo, Y.M.; Murphy, T.L.; Cowley, L.; Askwith, C.; Libina, N.; Gitschier, J.; Anderson, G.J. Hephaestin, a ceruloplasmin homologue implicated in intestinal iron transport, is defective in the sla mouse. Nat. Genet. 1999, 21, 195–199. [Google Scholar]
- Korolnek, T.; Hamza, I. Like iron in the blood of the people: The requirement for heme trafficking in iron metabolism. Front Pharmacol. 2014, 5, 126. [Google Scholar]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar]
- Bekri, S.; Gual, P.; Anty, R.; Luciani, N.; Dahman, M.; Ramesh, B.; Iannelli, A.; Staccini-Myx, A.; Casanova, D.; Ben Amor, I.; et al. Increased adipose tissue expression of hepcidin in severe obesity is independent from diabetes and nash. Gastroenterology 2006, 131, 788–796. [Google Scholar]
- Kulaksiz, H.; Fein, E.; Redecker, P.; Stremmel, W.; Adler, G.; Cetin, Y. Pancreatic beta-cells express hepcidin, an iron-uptake regulatory peptide. J. Endocrinol. 2008, 197, 241–249. [Google Scholar]
- Wenzel, B.J.; Stults, H.B.; Mayer, J. Hypoferraemia in obese adolescents. Lancet 1962, 2, 327–328. [Google Scholar]
- Seltzer, C.C.; Mayer, J. Serum iron and iron-binding capacity in adolescents. Ii. Comparison of obese and nonobese subjects. Am. J. Clin. Nutr. 1963, 13, 354–361. [Google Scholar]
- Manios, Y.; Moschonis, G.; Chrousos, G.P.; Lionis, C.; Mougios, V.; Kantilafti, M.; Tzotzola, V.; Skenderi, K.P.; Petridou, A.; Tsalis, G.; et al. The double burden of obesity and iron deficiency on children and adolescents in greece: The healthy growth study. J. Hum. Nutr. Diet. 2013, 26, 470–478. [Google Scholar]
- Nead, K.G.; Halterman, J.S.; Kaczorowski, J.M.; Auinger, P.; Weitzman, M. Overweight children and adolescents: A risk group for iron deficiency. Pediatrics 2004, 114, 104–108. [Google Scholar]
- Pinhas-Hamiel, O.; Newfield, R.S.; Koren, I.; Agmon, A.; Lilos, P.; Phillip, M. Greater prevalence of iron deficiency in overweight and obese children and adolescents. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 416–418. [Google Scholar]
- Moayeri, H.; Bidad, K.; Zadhoush, S.; Gholami, N.; Anari, S. Increasing prevalence of iron deficiency in overweight and obese children and adolescents (tehran adolescent obesity study). Eur. J. Pediatr. 2006, 165, 813–814. [Google Scholar]
- Shi, Z.; Lien, N.; Kumar, B.N.; Dalen, I.; Holmboe-Ottesen, G. The sociodemographic correlates of nutritional status of school adolescents in jiangsu province, china. J. Adolesc. Health 2005, 37, 313–322. [Google Scholar]
- Tussing-Humphreys, L.M.; Liang, H.; Nemeth, E.; Freels, S.; Braunschweig, C.A. Excess adiposity, inflammation, and iron-deficiency in female adolescents. J. Am. Diet. Assoc. 2009, 109, 297–302. [Google Scholar]
- Sanad, M.; Osman, M.; Gharib, A. Obesity modulate serum hepcidin and treatment outcome of iron deficiency anemia in children: A case control study. Ital. J. Pediatr. 2011, 37, 34. [Google Scholar]
- Aigner, E.; Hinz, C.; Steiner, K.; Rossmann, B.; Pfleger, J.; Hohla, F.; Steger, B.; Stadlmayr, A.; Patsch, W.; Datz, C. Iron stores, liver transaminase levels and metabolic risk in healthy teenagers. Eur. J. Clin. Investig. 2010, 40, 155–163. [Google Scholar]
- Micozzi, M.S.; Albanes, D.; Stevens, R.G. Relation of body size and composition to clinical biochemical and hematologic indices in us men and women. Am. J. Clin. Nutr. 1989, 50, 1276–1281. [Google Scholar]
- Ozata, M.; Mergen, M.; Oktenli, C.; Aydin, A.; Sanisoglu, S.Y.; Bolu, E.; Yilmaz, M.I.; Sayal, A.; Isimer, A.; Ozdemir, I.C. Increased oxidative stress and hypozincemia in male obesity. Clin. Biochem. 2002, 35, 627–631. [Google Scholar]
- Lefebvre, P.; Letois, F.; Sultan, A.; Nocca, D.; Mura, T.; Galtier, F. Nutrient deficiencies in patients with obesity considering bariatric surgery: A cross-sectional study. Surg. Obes. Relat. Dis. 2014, 10, 540–546. [Google Scholar]
- Cheng, H.L.; Bryant, C.; Cook, R.; O’Connor, H.; Rooney, K.; Steinbeck, K. The relationship between obesity and hypoferraemia in adults: A systematic review. Obes. Rev. 2012, 13, 150–161. [Google Scholar]
- Cepeda-Lopez, A.C.; Osendarp, S.J.; Melse-Boonstra, A.; Aeberli, I.; Gonzalez-Salazar, F.; Feskens, E.; Villalpando, S.; Zimmermann, M.B. Sharply higher rates of iron deficiency in obese mexican women and children are predicted by obesity-related inflammation rather than by differences in dietary iron intake. Am. J. Clin. Nutr. 2011, 93, 975–983. [Google Scholar]
- Tussing-Humphreys, L.M.; Nemeth, E.; Fantuzzi, G.; Freels, S.; Guzman, G.; Holterman, A.X.; Braunschweig, C. Elevated systemic hepcidin and iron depletion in obese premenopausal females. Obesity (Silver Spring) 2010, 18, 1449–1456. [Google Scholar] [CrossRef]
- Gong, L.; Yuan, F.; Teng, J.; Li, X.; Zheng, S.; Lin, L.; Deng, H.; Ma, G.; Sun, C.; Li, Y. Weight loss, inflammatory markers, and improvements of iron status in overweight and obese children. J. Pediatr. 2014, 164, 795–800.e2. [Google Scholar] [CrossRef]
- Amato, A.; Santoro, N.; Calabro, P.; Grandone, A.; Swinkels, D.W.; Perrone, L.; del Giudice, E.M. Effect of body mass index reduction on serum hepcidin levels and iron status in obese children. Int. J. Obes. 2010, 34, 1772–1774. [Google Scholar]
- Di Toro, A.; Marotta, A.; Todisco, N.; Ponticiello, E.; Collini, R.; Di Lascio, R.; Perrone, L. Unchanged iron and copper and increased zinc in the blood of obese children after two hypocaloric diets. Biol. Trace Elem. Res. 1997, 57, 97–104. [Google Scholar]
- Beard, J.; Borel, M.; Peterson, F.J. Changes in iron status during weight loss with very-low-energy diets. Am. J. Clin. Nutr. 1997, 66, 104–110. [Google Scholar]
- Tussing-Humphreys, L.M.; Nemeth, E.; Fantuzzi, G.; Freels, S.; Holterman, A.X.; Galvani, C.; Ayloo, S.; Vitello, J.; Braunschweig, C. Decreased serum hepcidin and improved functional iron status 6 months after restrictive bariatric surgery. Obesity (Silver Spring) 2010, 18, 2010–2016. [Google Scholar] [CrossRef]
- Munoz, M.; Botella-Romero, F.; Gomez-Ramirez, S.; Campos, A.; Garcia-Erce, J.A. Iron deficiency and anaemia in bariatric surgical patients: Causes, diagnosis and proper management. Nutr. Hosp. 2009, 24, 640–654. [Google Scholar]
- Aktas, G.; Alcelik, A.; Yalcin, A.; Karacay, S.; Kurt, S.; Akduman, M.; Savli, H. Treatment of iron deficiency anemia induces weight loss and improves metabolic parameters. Clin. Ter. 2014, 165, e87–e89. [Google Scholar]
- Fried, M.; Yumuk, V.; Oppert, J.M.; Scopinaro, N.; Torres, A.; Weiner, R.; Yashkov, Y.; Fruhbeck, G. Interdisciplinary european guidelines on metabolic and bariatric surgery. Obes. Surg. 2014, 24, 42–55. [Google Scholar]
- Baptista, V.; Wassef, W. Bariatric procedures: An update on techniques, outcomes and complications. Curr. Opin. Gastroenterol. 2013, 29, 684–693. [Google Scholar]
- Patel, S.R.; Hakim, D.; Mason, J.; Hakim, N. The duodenal-jejunal bypass sleeve (endobarrier gastrointestinal liner) for weight loss and treatment of type 2 diabetes. Surg. Obes. Relat. Dis. 2013, 9, 482–484. [Google Scholar]
- Flancbaum, L.; Belsley, S.; Drake, V.; Colarusso, T.; Tayler, E. Preoperative nutritional status of patients undergoing roux-en-y gastric bypass for morbid obesity. J. Gastrointest. Surg. 2006, 10, 1033–1037. [Google Scholar]
- Damms-Machado, A.; Friedrich, A.; Kramer, K.M.; Stingel, K.; Meile, T.; Kuper, M.A.; Konigsrainer, A.; Bischoff, S.C. Pre- and postoperative nutritional deficiencies in obese patients undergoing laparoscopic sleeve gastrectomy. Obes. Surg. 2012, 22, 881–889. [Google Scholar]
- Ernst, B.; Thurnheer, M.; Schmid, S.M.; Schultes, B. Evidence for the necessity to systematically assess micronutrient status prior to bariatric surgery. Obes. Surg. 2009, 19, 66–73. [Google Scholar]
- Harju, E. Empty iron stores as a significant risk factor in abdominal surgery. J. Parenter. Enteral Nutr. 1988, 12, 282–285. [Google Scholar]
- Hallet, J.; Hanif, A.; Callum, J.; Pronina, I.; Wallace, D.; Yohanathan, L.; McLeod, R.; Coburn, N. The impact of perioperative iron on the use of red blood cell transfusions in gastrointestinal surgery: A systematic review and meta-analysis. Transfus. Med. Rev. 2014. [Google Scholar] [CrossRef]
- Shander, A.; Van Aken, H.; Colomina, M.J.; Gombotz, H.; Hofmann, A.; Krauspe, R.; Lasocki, S.; Richards, T.; Slappendel, R.; Spahn, D.R. Patient blood management in europe. Br. J. Anaesth. 2012, 109, 55–68. [Google Scholar]
- Skroubis, G.; Kouri, N.; Mead, N.; Kalfarentzos, F. Long-term results of a prospective comparison of roux-en-y gastric bypass versus a variant of biliopancreatic diversion in a non-superobese population (bmi 35–50 kg/m2). Obes. Surg. 2014, 24, 197–204. [Google Scholar]
- Coupaye, M.; Puchaux, K.; Bogard, C.; Msika, S.; Jouet, P.; Clerici, C.; Larger, E.; Ledoux, S. Nutritional consequences of adjustable gastric banding and gastric bypass: A 1-year prospective study. Obes. Surg. 2009, 19, 56–65. [Google Scholar]
- Alexandrou, A.; Armeni, E.; Kouskouni, E.; Tsoka, E.; Diamantis, T.; Lambrinoudaki, I. Cross-sectional long-term micronutrient deficiencies after sleeve gastrectomy versus roux-en-y gastric bypass: A pilot study. Surg. Obes. Relat. Dis. 2014, 10, 262–268. [Google Scholar]
- Gehrer, S.; Kern, B.; Peters, T.; Christoffel-Courtin, C.; Peterli, R. Fewer nutrient deficiencies after laparoscopic sleeve gastrectomy (lsg) than after laparoscopic roux-y-gastric bypass (lrygb)-a prospective study. Obes. Surg. 2010, 20, 447–453. [Google Scholar]
- Anty, R.; Dahman, M.; Iannelli, A.; Gual, P.; Staccini-Myx, A.; Amor, I.B.; Luciani, N.; Saint-Paul, M.C.; Huet, P.M.; Sadoul, J.L.; et al. Bariatric surgery can correct iron depletion in morbidly obese women: A link with chronic inflammation. Obes. Surg. 2008, 18, 709–714. [Google Scholar]
- Ramalho, R.; Guimaraes, C.; Gil, C.; Neves, C.; Guimaraes, J.T.; Delgado, L. Morbid obesity and inflammation: A prospective study after adjustable gastric banding surgery. Obes. Surg. 2009, 19, 915–920. [Google Scholar]
- Mechanick, J.I.; Youdim, A.; Jones, D.B.; Garvey, W.T.; Hurley, D.L.; McMahon, M.M.; Heinberg, L.J.; Kushner, R.; Adams, T.D.; Shikora, S.; et al. Clinical practice guidelines for the perioperative nutritional, metabolic, and nonsurgical support of the bariatric surgery patient—2013 update: Cosponsored by american association of clinical endocrinologists, the obesity society, and american society for metabolic & bariatric surgery. Obesity (Silver Spring) 2013, 21 (Suppl. 1), S1–S27. [Google Scholar]
- Gesquiere, I.; Lannoo, M.; Augustijns, P.; Matthys, C.; Van der Schueren, B.; Foulon, V. Iron deficiency after roux-en-y gastric bypass: Insufficient iron absorption from oral iron supplements. Obes. Surg. 2014, 24, 56–61. [Google Scholar]
- Marambio, A.; Watkins, G.; Castro, F.; Riffo, A.; Zuniga, R.; Jans, J.; Villanueva, M.E.; Diaz, G. Changes in iron transporter divalent metal transporter 1 in proximal jejunum after gastric bypass. World J. Gastroenterol. 2014, 20, 6534–6540. [Google Scholar]
- Ruz, M.; Carrasco, F.; Rojas, P.; Codoceo, J.; Inostroza, J.; Basfi-Fer, K.; Valencia, A.; Csendes, A.; Papapietro, K.; Pizarro, F.; et al. Heme- and nonheme-iron absorption and iron status 12 mo after sleeve gastrectomy and roux-en-y gastric bypass in morbidly obese women. Am. J. Clin. Nutr. 2012, 96, 810–817. [Google Scholar]
- Santos, J.; Salgado, P.; Santos, C.; Mendes, P.; Saavedra, J.; Baldaque, P.; Monteiro, L.; Costa, E. Effect of bariatric surgery on weight loss, inflammation, iron metabolism, and lipid profile. Scand. J. Surg. 2014, 103, 21–25. [Google Scholar]
- Mujica-Coopman, M.F.; Brito, A.; Lopez de Romana, D.; Pizarro, F.; Olivares, M. Body mass index, iron absorption and iron status in childbearing age women. J. Trace Elem. Med. Biol. 2014. [Google Scholar] [CrossRef]
- Zimmermann, M.B.; Zeder, C.; Muthayya, S.; Winichagoon, P.; Chaouki, N.; Aeberli, I.; Hurrell, R.F. Adiposity in women and children from transition countries predicts decreased iron absorption, iron deficiency and a reduced response to iron fortification. Int. J. Obes. 2008, 32, 1098–1104. [Google Scholar]
- del Giudice, E.M.; Santoro, N.; Amato, A.; Brienza, C.; Calabro, P.; Wiegerinck, E.T.; Cirillo, G.; Tartaglione, N.; Grandone, A.; Swinkels, D.W.; et al. Hepcidin in obese children as a potential mediator of the association between obesity and iron deficiency. J. Clin. Endocrinol. Metab. 2009, 94, 5102–5107. [Google Scholar]
- Aeberli, I.; Hurrell, R.F.; Zimmermann, M.B. Overweight children have higher circulating hepcidin concentrations and lower iron status but have dietary iron intakes and bioavailability comparable with normal weight children. Int. J. Obes. 2009, 33, 1111–1117. [Google Scholar] [Green Version]
- Aigner, E.; Theurl, I.; Theurl, M.; Lederer, D.; Haufe, H.; Dietze, O.; Strasser, M.; Datz, C.; Weiss, G. Pathways underlying iron accumulation in human nonalcoholic fatty liver disease. Am. J. Clin. Nutr. 2008, 87, 1374–1383. [Google Scholar]
- Barisani, D.; Pelucchi, S.; Mariani, R.; Galimberti, S.; Trombini, P.; Fumagalli, D.; Meneveri, R.; Nemeth, E.; Ganz, T.; Piperno, A. Hepcidin and iron-related gene expression in subjects with dysmetabolic hepatic iron overload. J. Hepatol. 2008, 49, 123–133. [Google Scholar]
- Ruivard, M.; Laine, F.; Ganz, T.; Olbina, G.; Westerman, M.; Nemeth, E.; Rambeau, M.; Mazur, A.; Gerbaud, L.; Tournilhac, V.; et al. Iron absorption in dysmetabolic iron overload syndrome is decreased and correlates with increased plasma hepcidin. J. Hepatol. 2009, 50, 1219–1225. [Google Scholar]
- McClung, J.P.; Karl, J.P. Iron deficiency and obesity: The contribution of inflammation and diminished iron absorption. Nutr. Rev. 2009, 67, 100–104. [Google Scholar]
- Tussing-Humphreys, L.; Frayn, K.N.; Smith, S.R.; Westerman, M.; Dennis, A.L.; Nemeth, E.; Thomson, J.; Pusatcioglu, C. Subcutaneous adipose tissue from obese and lean adults does not release hepcidin in vivo. Sci. World J. 2011, 11, 2197–2206. [Google Scholar]
- Ganz, T.; Nemeth, E. Iron sequestration and anemia of inflammation. Semin. Hematol. 2009, 46, 387–393. [Google Scholar]
- Nairz, M.; Haschka, D.; Demetz, E.; Weiss, G. Iron at the interface of immunity and infection. Front Pharmacol. 2014, 5, 152. [Google Scholar]
- Sonnweber, T.; Ress, C.; Nairz, M.; Theurl, I.; Schroll, A.; Murphy, A.T.; Wroblewski, V.; Witcher, D.R.; Moser, P.; Ebenbichler, C.F.; et al. High-fat diet causes iron deficiency via hepcidin-independent reduction of duodenal iron absorption. J. Nutr. Biochem. 2012, 23, 1600–1608. [Google Scholar]
- Aigner, E.; Theurl, I.; Haufe, H.; Seifert, M.; Hohla, F.; Scharinger, L.; Stickel, F.; Mourlane, F.; Weiss, G.; Datz, C. Copper availability contributes to iron perturbations in human nonalcoholic fatty liver disease. Gastroenterology 2008, 135, 680–688. [Google Scholar]
- Matak, P.; Zumerle, S.; Mastrogiannaki, M.; El Balkhi, S.; Delga, S.; Mathieu, J.R.; Canonne-Hergaux, F.; Poupon, J.; Sharp, P.A.; Vaulont, S.; et al. Copper deficiency leads to anemia, duodenal hypoxia, upregulation of hif-2alpha and altered expression of iron absorption genes in mice. PLoS One 2013, 8, e59538. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Aigner, E.; Feldman, A.; Datz, C. Obesity as an Emerging Risk Factor for Iron Deficiency. Nutrients 2014, 6, 3587-3600. https://0-doi-org.brum.beds.ac.uk/10.3390/nu6093587
Aigner E, Feldman A, Datz C. Obesity as an Emerging Risk Factor for Iron Deficiency. Nutrients. 2014; 6(9):3587-3600. https://0-doi-org.brum.beds.ac.uk/10.3390/nu6093587
Chicago/Turabian StyleAigner, Elmar, Alexandra Feldman, and Christian Datz. 2014. "Obesity as an Emerging Risk Factor for Iron Deficiency" Nutrients 6, no. 9: 3587-3600. https://0-doi-org.brum.beds.ac.uk/10.3390/nu6093587