Antidiabetic Effects of Yam (Dioscorea batatas) and Its Active Constituent, Allantoin, in a Rat Model of Streptozotocin-Induced Diabetes


 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Plant Materials and Preparation of Water Extract
2.2. Experimental Animals
2.3. Induction of Experimental Diabetes Mellitus and Experimental Design
2.4. Measurement of Blood Glucose Levels
2.5. Biochemical Analysis
2.6. Serum Insulin, Nitrite/Nitrate, GLP-1, C-Peptide, and HbAlc Measurements
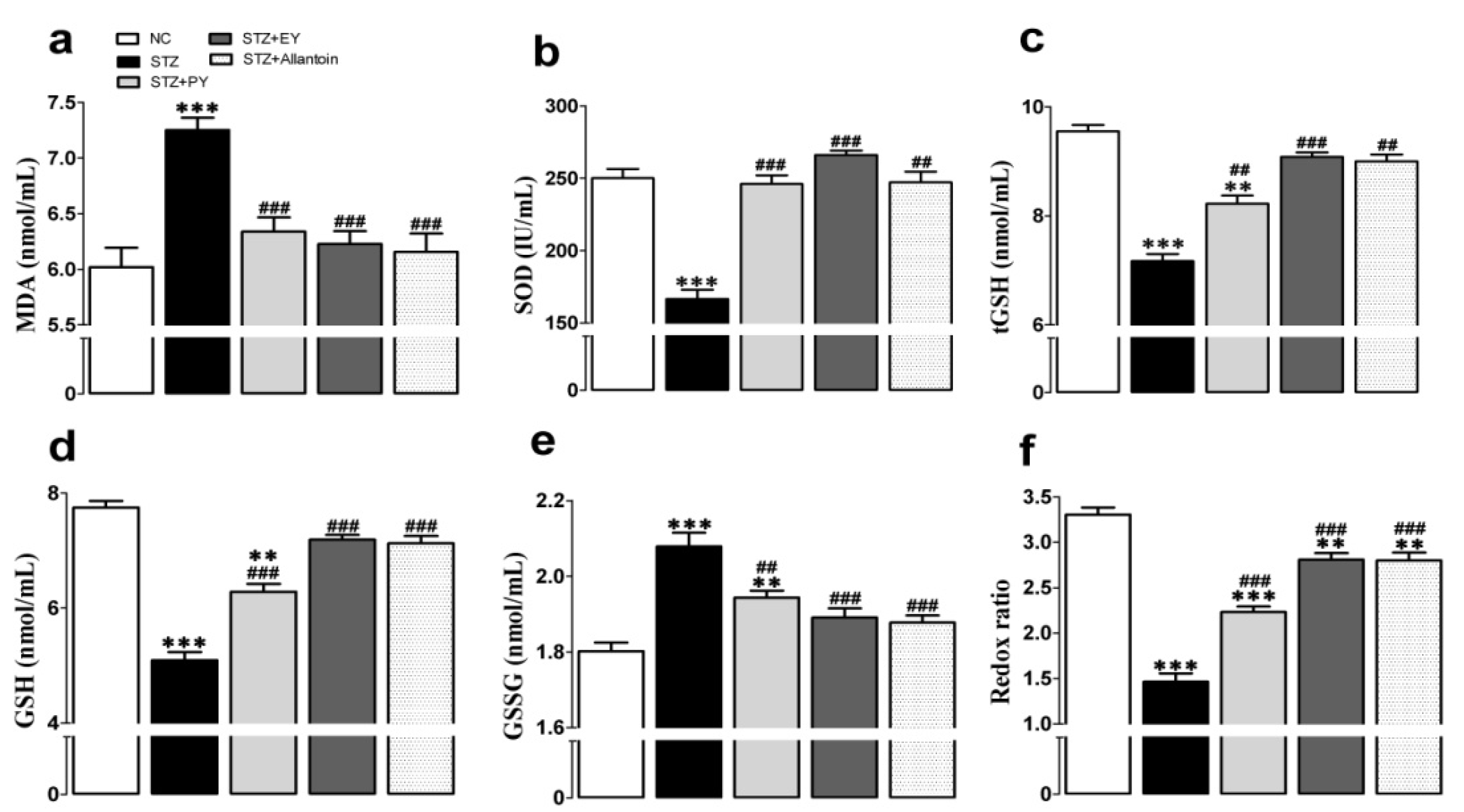
2.7. Measurement of Antioxidant Defense
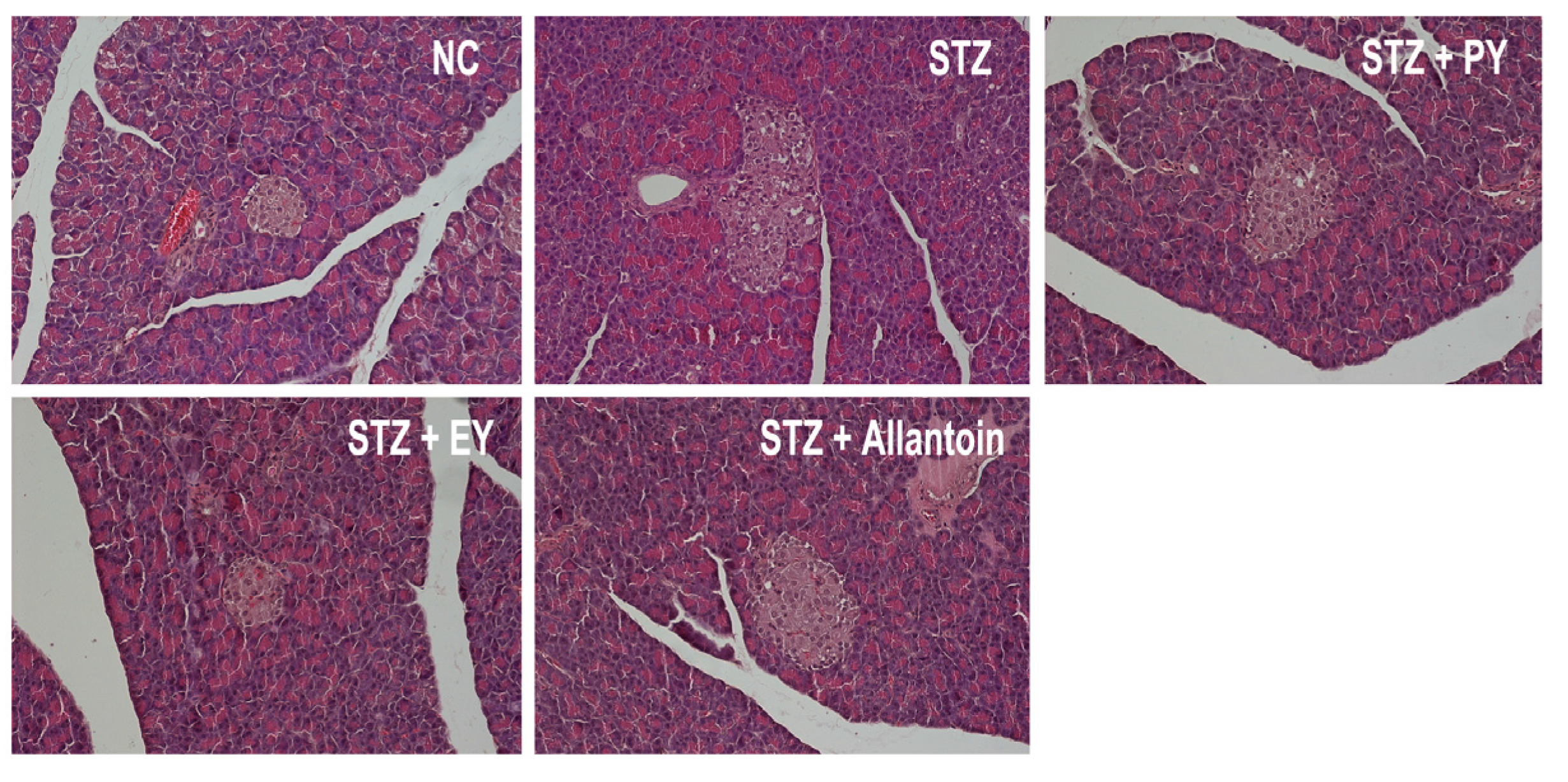
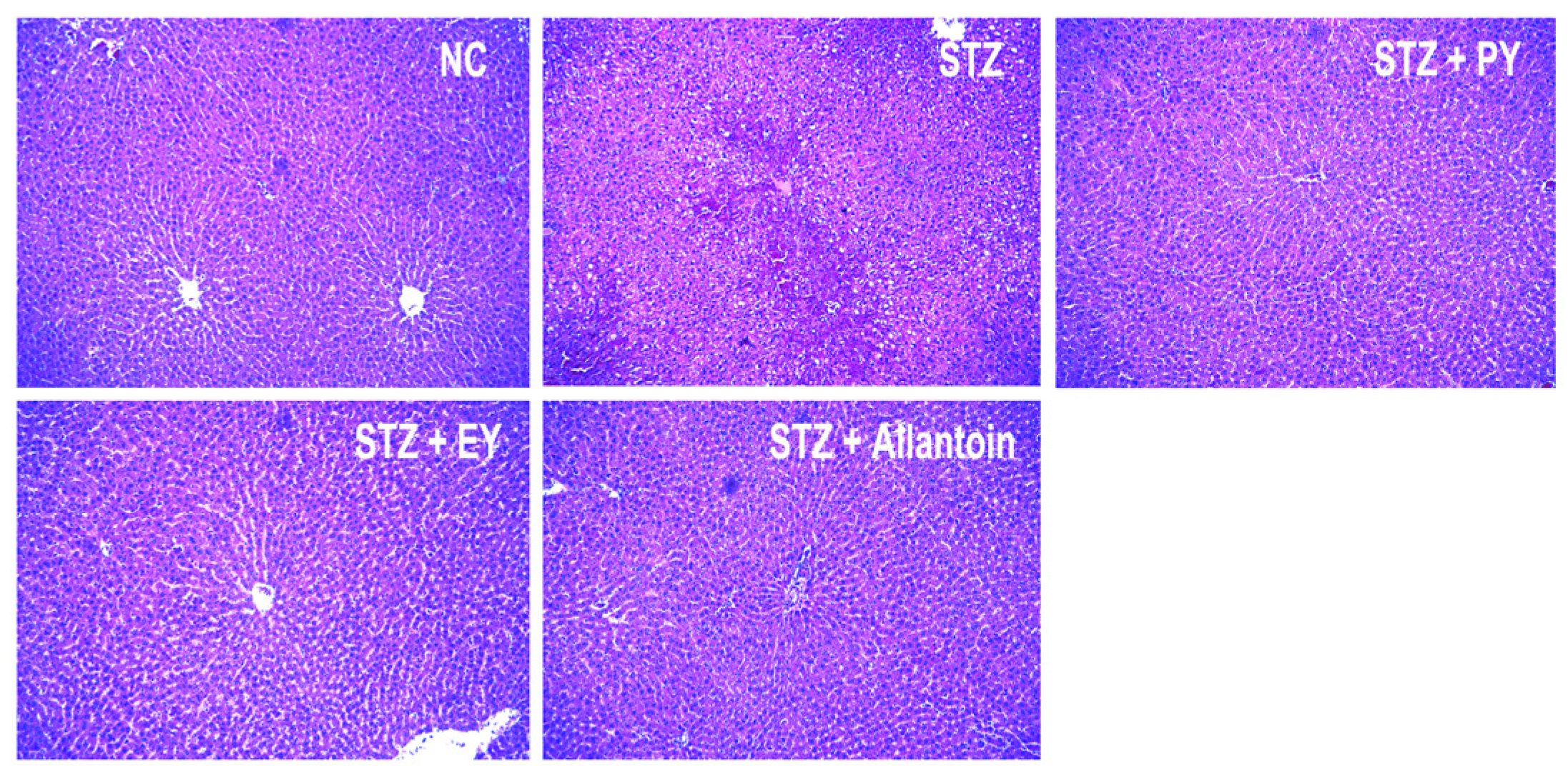
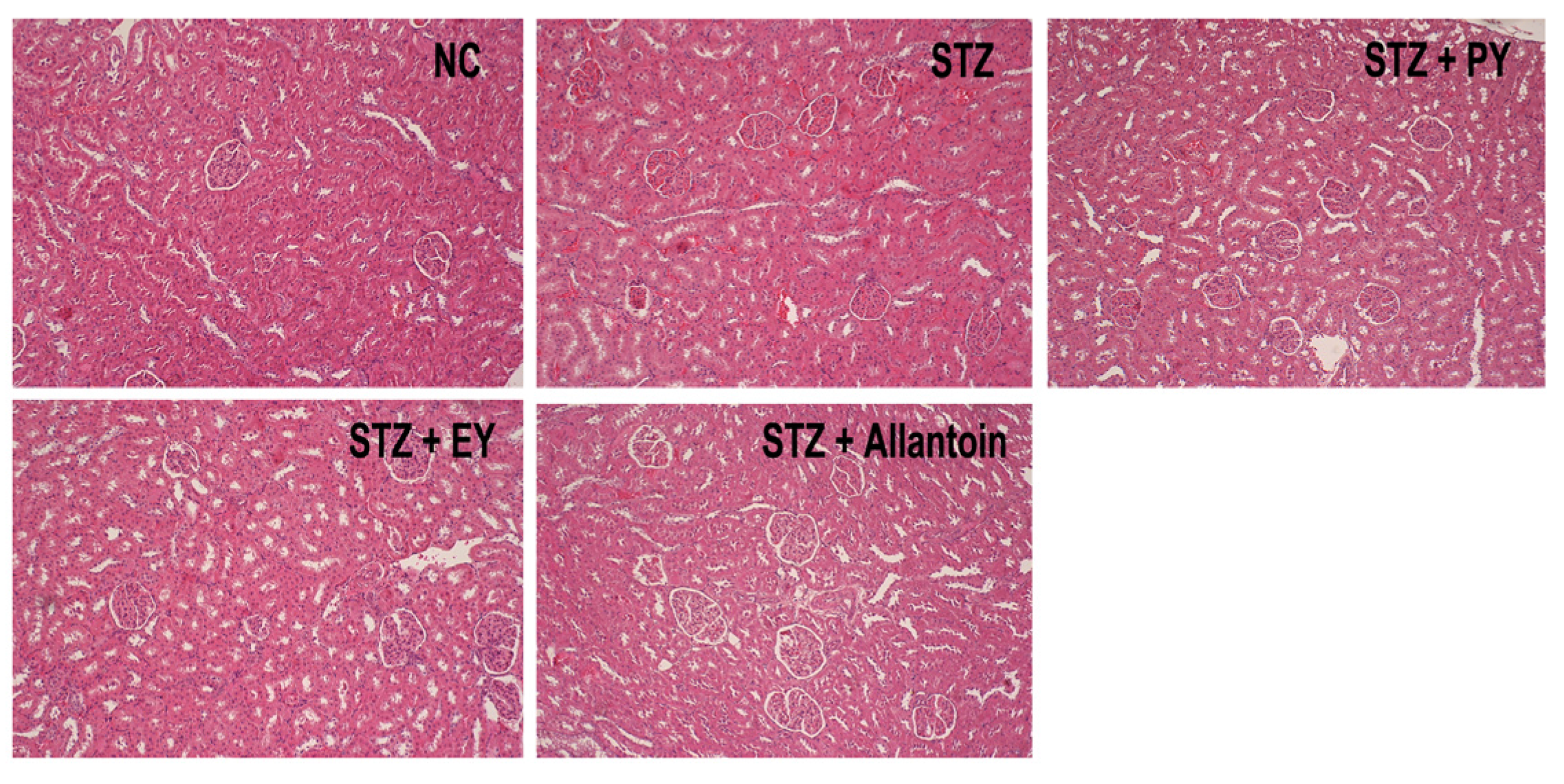
2.8. Histological Analysis
2.9. Statistical Analysis
3. Results
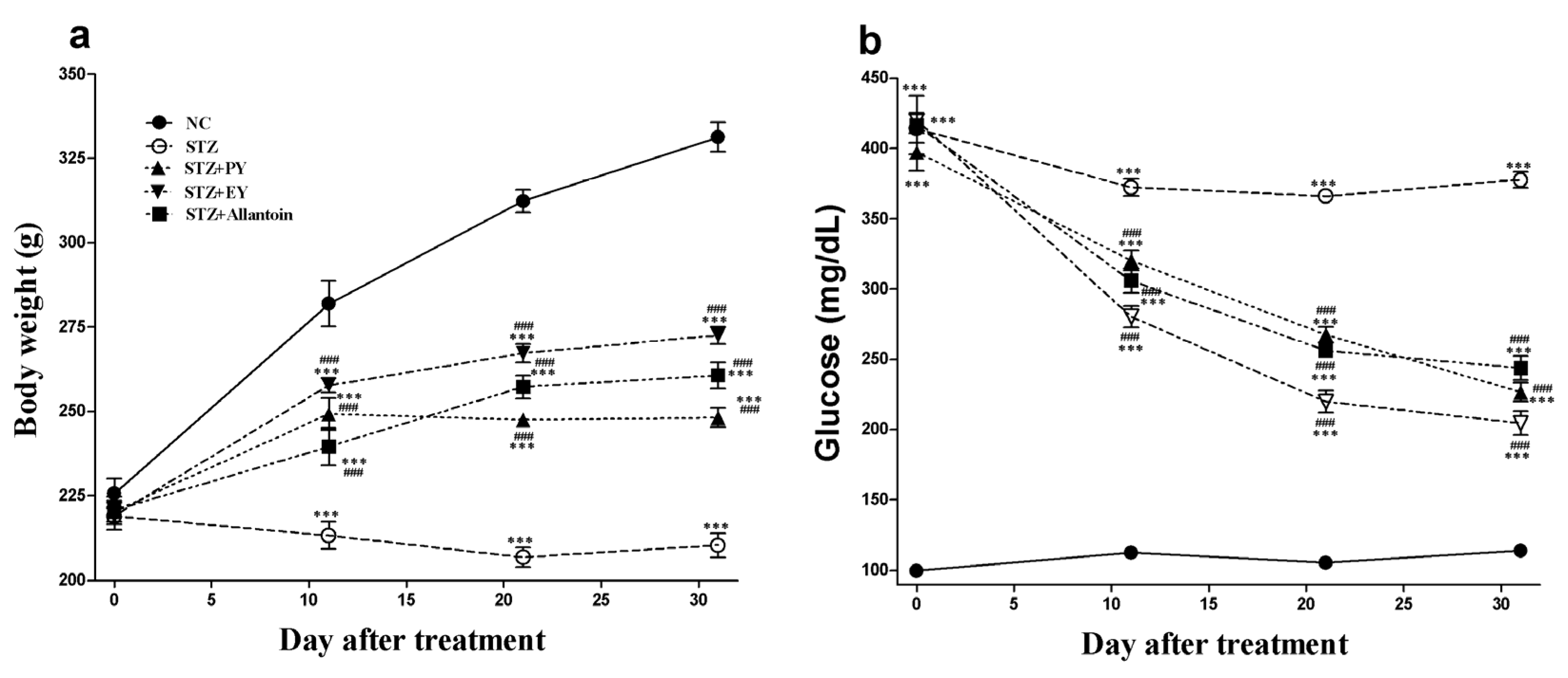
3.1. Effects of the Different Treatments on BW, Blood Glucose Levels, and Serum Insulin Levels
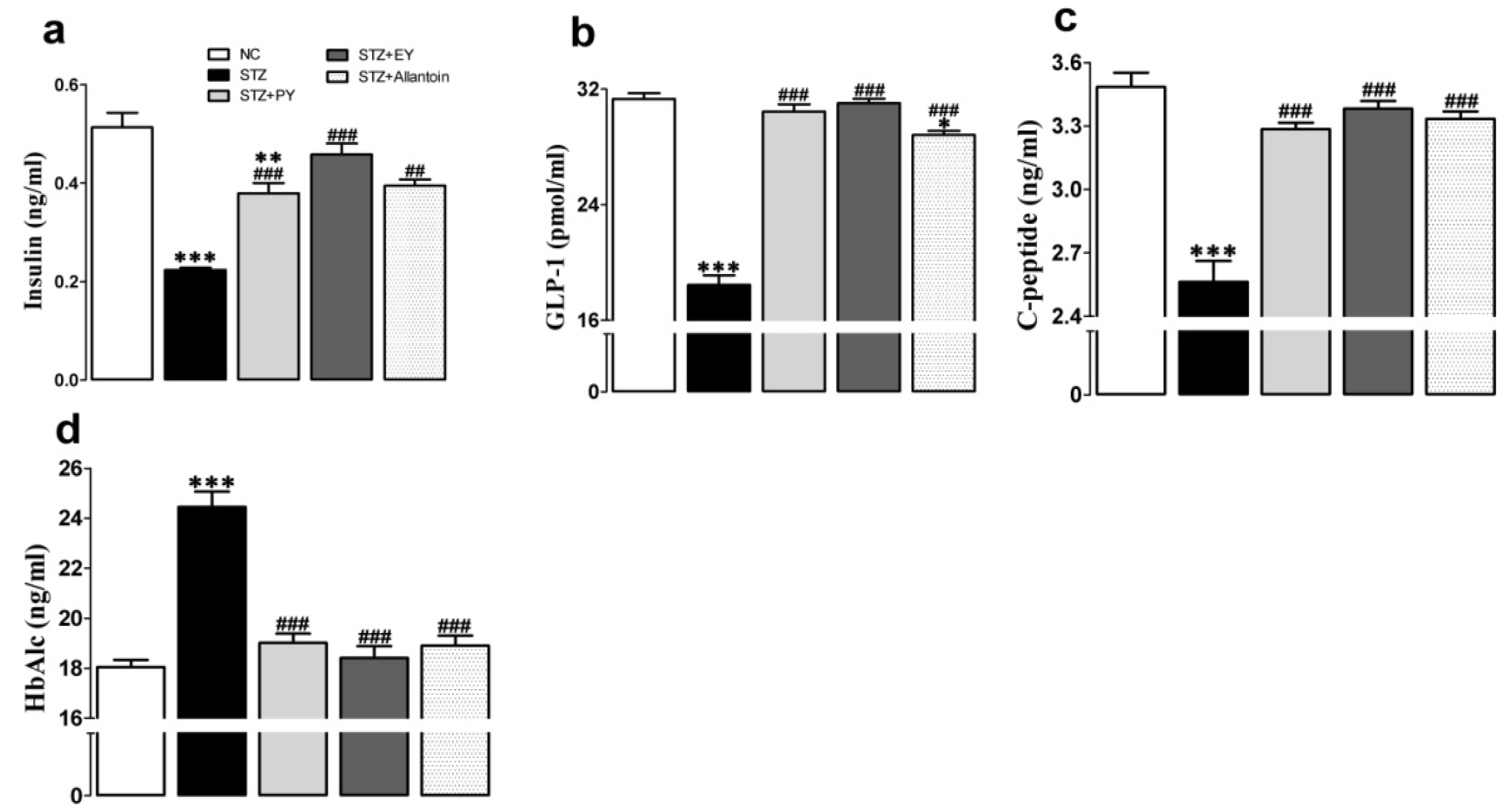
3.2. Effects of the Different Treatments on Serum Insulin, GLP-1, C-Peptide, HbAlc, and Nitric Oxide Levels


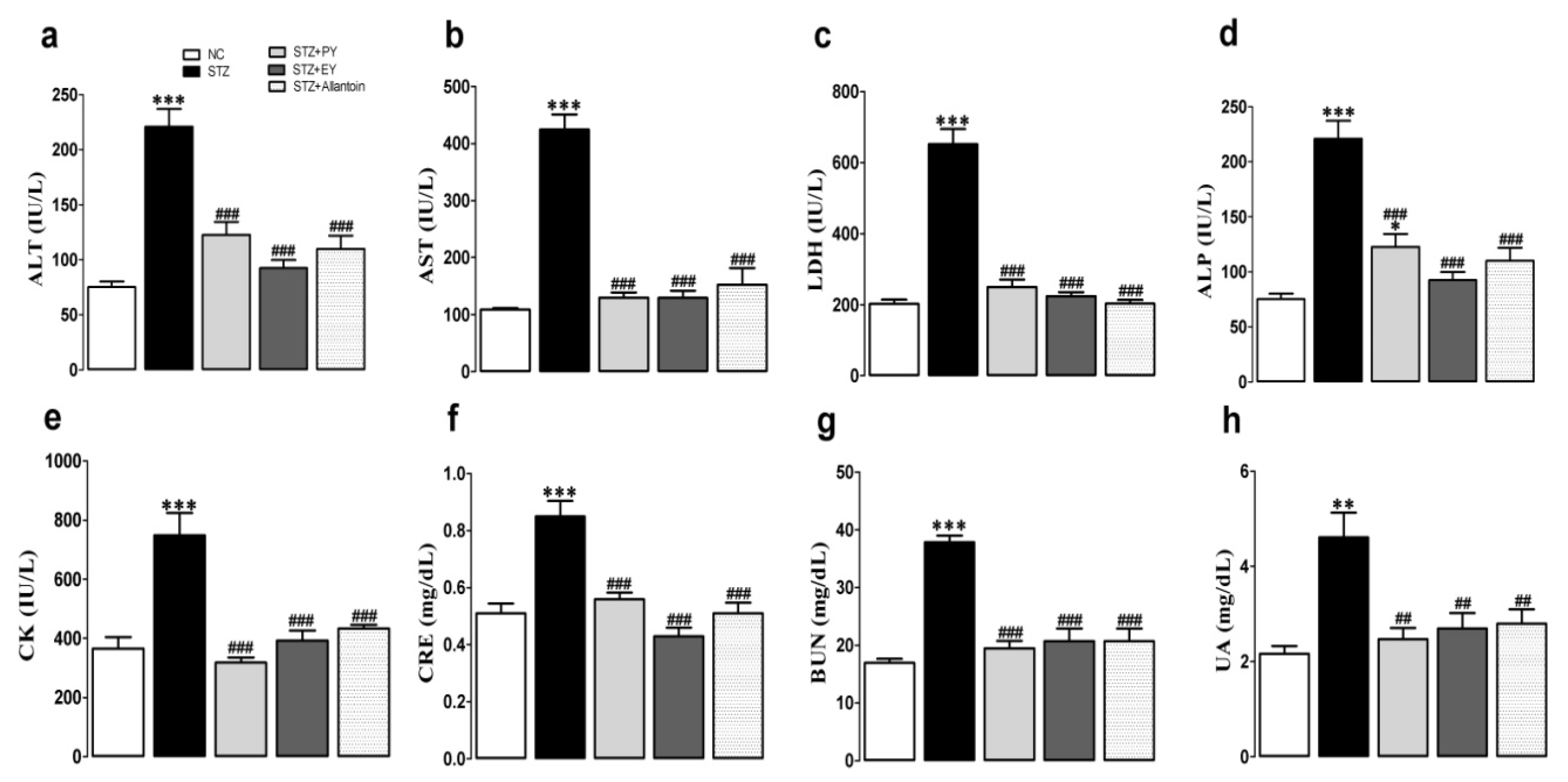
3.3. Effects of the Different Treatments on the Serum Levels of Metabolic Enzymes, BUN, CRE, and UA
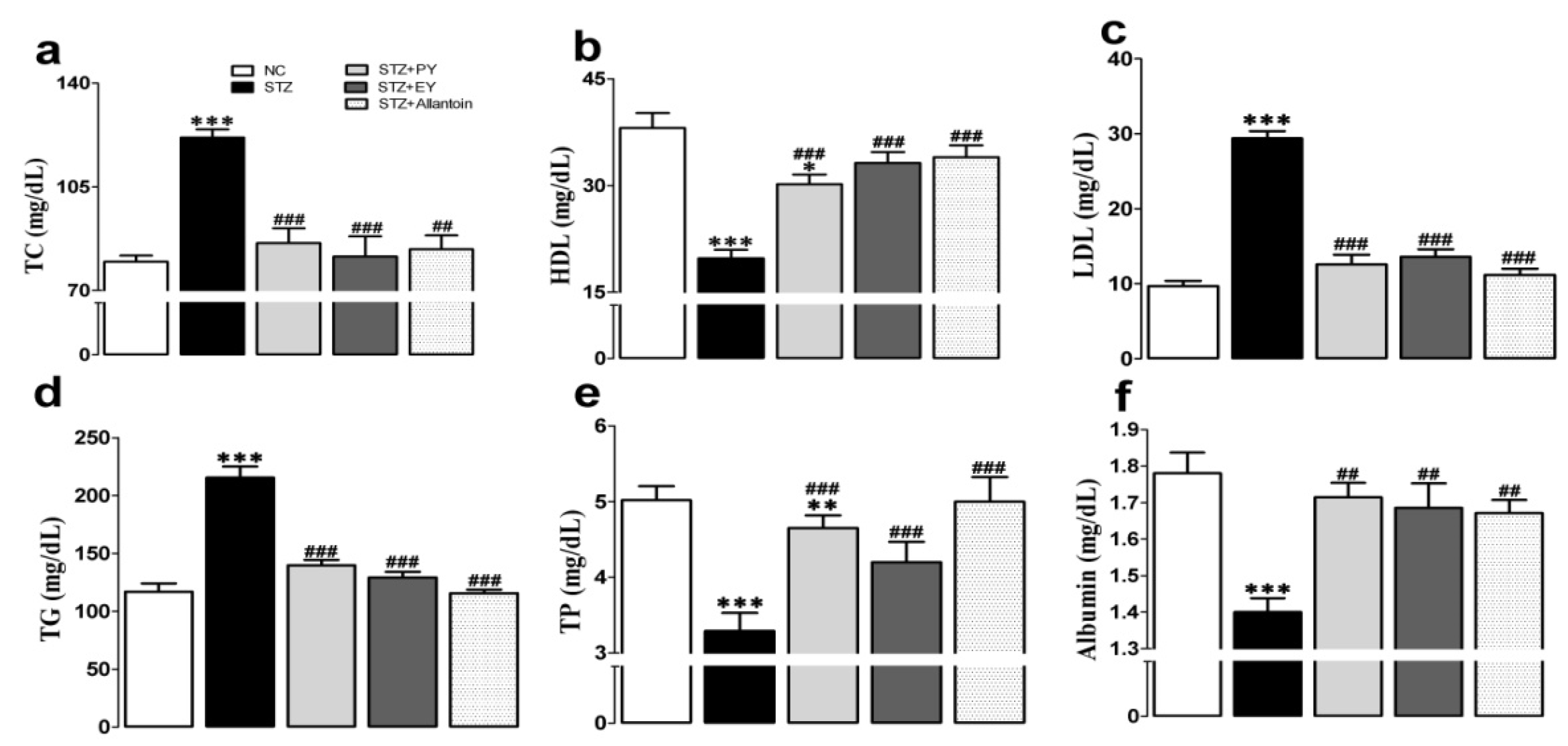
3.4. Effects of the Different Treatments on Serum Lipid and Protein Levels


3.5. Effects of the on Different Treatments Antioxidant Activities
3.6. Effects of the Different Treatments on Blood Ion and Metabolite Concentrations


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NC | STZ | STZ + PY | STZ + EY | STZ + Allatoin | |
|---|---|---|---|---|---|
| Hb (g/dL) | 13.4 ± 0.3 | 10.8 ± 0.3 | 13.2 ± 0.4 *** | 13.5 ± 0.2 * | 13.3 ± 0.1 *** |
| Hct (%) | 42 ± 1 | 36 ± 1 *** | 41 ± 1 ## | 42 ± 1 ### | 41 ± 1 ** |
| pH | 7.41 ± 0.01 | 7.33 ± 0.01 *** | 7.43 ± 0.01 ### | 7.43 ± 0.01 ### | 7.44 ± 0.01 ## |
| Lactate (mmol/L) | 3.9 ± 0.4 | 6.0 ± 0.3 ** | 4.2 ± 0.2 ### | 4.3 ± 0.3 ## | 4.4 ± 0.4 # |
| HCO3− (mmol/L) | 27.54 ± 0.58 | 23.63 ± 0.87 * | 26.86 ± 0.36 | 29.24 ± 0.40 ## | 28.97 ± 0.069 ## |
| Osm (mOsm/L) | 298 ± 1 | 324 ± 1 *** | 307 ± 3 *,## | 298 ± 1 ### | 302 ± 3 *,### |
| Angap (mmol/L) | 13.5 ± 0.7 | 14.7 ± 1.0 | 13.6 ± 0.8 | 12.8 ± 0.3 | 12.9 ± 1.1 |
| Na+ (mmol/L) | 149 ± 0 | 151 ± 0 * | 150 ± 1 | 148 ± 0 # | 150 ± 1 |
| Cl− (mmol/L) | 108 ± 1 | 113 ± 1 * | 112 ± 2 | 106 ± 0 ## | 110 ± 1 |
| Mg2+ (mmol/L) | 0.44 ± 0.11 | 0.41 ± 0.01 | 0.44 ± 0.01 ### | 0.43 ± 0.01 ### | 0.44 ± 0.02 ### |
| K+ (mmol/L) | 4.3 ± 0.1 | 5.0 ± 0.2 * | 4.5 ± 0.2 | 4.3 ± 0.1 # | 4.3 ± 0.1 # |
| Ca2+ (mmol/L) | 1.11 ± 0.01 | 1.02 ± 0.02 * | 1.14 ± 0.03 # | 1.15 ± 0.02 ## | 1.12 ± 0.01 # |
3.7. Histological Analysis



4. Discussion
5. Conclusions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mahendran, G.; Thamotharan, G.; Sengottuvelu, S.; Bai, V.N. Anti-diabetic activity of Swertia corymbosa (griseb.) Wight ex C.B. Clarke aerial parts extract in streptozotocin induced diabetic rats. J. Ethnopharmacol. 2014, 151, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Wild, S.; Roglic, G.; Green, A.; Sicree, R.; King, H. Global prevalence of diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care 2004, 27, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Giorgino, F.; Laviola, L.; Leonardini, A. Pathophysiology of type 2 diabetes: Rationale for different oral antidiabetic treatment strategies. Diabetes Res. Clin. Pract. 2005, 68, S22–S29. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.S.; Chen, W.; Wu, H.T.; Cheng, K.C.; Wen, Y.J.; Lin, K.C.; Cheng, J.T. Decrease of plasma glucose by allantoin, an active principle of yam (Dioscorea spp.), in streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2010, 58, 12031–12035. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Kim, M.J.; Moon, H.B. Occupational asthma caused by two herb materials, dioscorea batatas and pinellia ternata. Clin. Exp. Allergy 1994, 24, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, N.; Nagasawa, T.; Mabuchi, R.; Yasui, Y.; Wakabayashi, K.; Tanaka, T.; Ohshima, H. Chemoprevention of azoxymethane/dextran sodium sulfate-induced mouse colon carcinogenesis by freeze-dried yam sanyaku and its constituent diosgenin. Cancer Prev. Res. 2011, 4, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.F.; Tsai, J.T.; Chen, L.J.; Wu, T.P.; Yang, J.J.; Yin, L.T.; Yang, Y.L.; Chiang, T.A.; Lu, H.L.; Wu, M.C. Antihypertensive action of allantoin in animals. Biomed. Res. Int. 2014. [Google Scholar] [CrossRef] [PubMed]
- McAnuff, M.A.; Harding, W.W.; Omoruyi, F.O.; Jacobs, H.; Morrison, E.Y.; Asemota, H.N. Hypoglycemic effects of steroidal sapogenins isolated from jamaican bitter yam, dioscorea polygonoides. Food Chem. Toxicol. 2005, 43, 1667–1672. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Yang, F.L.; Lu, C.P.; Yang, Y.L.; Wen, C.L.; Hua, K.F.; Wu, S.H. Polysaccharides from dioscorea batatas induce tumor necrosis factor-alpha secretion via toll-like receptor 4-mediated protein kinase signaling pathways. J. Agric. Food. Chem. 2008, 56, 9892–9898. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, Y.; Chen, G. Simultaneous determination of allantoin, choline and l-arginine in Rhizoma Dioscoreae by capillary electrophoresis. J. Chromatogr. A 2004, 1043, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Son, I.S.; Kim, J.H.; Sohn, H.Y.; Son, K.H.; Kim, J.S.; Kwon, C.S. Antioxidative and hypolipidemic effects of diosgenin, a steroidal saponin of yam (Dioscorea spp.), on high-cholesterol fed rats. Biosci. Biotechnol. Biochem. 2007, 71, 3063–3071. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jwa, H.; Yanagawa, Y.; Park, T. Extract from dioscorea batatas ameliorates insulin resistance in mice fed a high-fat diet. J. Med. Food. 2012, 15, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.C.; Yeh, L.R.; Chen, L.J.; Wen, Y.J.; Cheng, K.C.; Cheng, J.T. Plasma glucose-lowering action of allantoin is induced by activation of imidazoline I-2 receptors in streptozotocin-induced diabetic rats. Horm. Metab. Res. 2012, 44, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Institute of Laboratory Animal Resources; Commission on Life Sciences; National Research Council; National Research Council. Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, DC, USA, 1996; pp. 1–124. [Google Scholar]
- Yoon, K.D.; Yang, M.H.; Chin, Y.W.; Park, J.H.; Kim, J.W. Determination of allantoin in Dioscorea Rhizoma by high performance liquid chromatography using cyano columns. Nat. Prod. Sci. 2008, 14, 254–259. [Google Scholar]
- Kwon, J.E.; Kwon, J.B.; Kwun, I.S.; Sohn, H.Y. Antimicrobial and antioxidant activity of the Dioscorea alata L. J. Microbiol. Biotechnol. 2010, 38, 283–288. [Google Scholar]
- Rahman, M.M.; Lee, S.J.; Mun, A.R.; Adam, G.O.; Park, R.M.; Kim, G.B.; Kang, H.S.; Kim, J.S.; Kim, S.J.; Kim, S.Z. Relationships between blood Mg2+ and energy metabolites/enzymes after acute exhaustive swimming exercise in rats. Biol. Trace Element Res. 2014, 161, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Behr, J.; Maier, K.; Degenkolb, B.; Krombach, F.; Vogelmeier, C. Antioxidative and clinical effects of high-dose N-acetylcysteine in fibrosing alveolitis—Adjunctive therapy to maintenance immunosuppression. Am. J. Respir. Crit. Care Med. 1997, 156, 1897–1901. [Google Scholar] [CrossRef] [PubMed]
- Manaer, T.; Yu, L.; Zhang, Y.; Xiao, X.J.; Nabi, X.H. Anti-diabetic effects of shubat in type 2 diabetic rats induced by combination of high-glucose-fat diet and low-dose streptozotocin. J. Ethnopharmacol. 2015, 169, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.; Wang, Y.; Mehendale, S.; Sun, S.; Wang, C.Z.; Xie, J.T.; Aung, H.H.; Yuan, C.S. Antioxidant protection by American ginseng in pancreatic β-cells. Am. J. Chin. Med. 2008, 36, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.J.; Lee, Y.C.; Liu, S.Y.; Chang, T.W. Chinese yam (Dioscorea alata cv. Tainung No. 2) feeding exhibited antioxidative effects in hyperhomocysteinemia rats. J. Agric. Food. Chem. 2004, 52, 1720–1725. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Gou, J.F.; Tian, J.H.; Yan, X.A.; Yang, L. Glucagon-like peptide-1 receptor agonists versus insulin glargine for type 2 diabetes mellitus: A systematic review and meta-analysis of randomized controlled trials. Curr. Ther. Res. Clin. Exp. 2010, 71, 211–238. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Lecourt, E.; Dewulf, E.M.; Sohet, F.M.; Pachikian, B.D.; Naslain, D.; de Backer, F.; Neyrinck, A.M.; Delzenne, N.M. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am. J. Clin. Nutr. 2009, 90, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.F.; Zhang, Y.Z.; Yin, Y.L.; Wu, G.Y.; Zhou, H.J.; Tan, Z.L.; Yang, F.; Bo, M.J.; Huang, R.L.; Li, T.J.; et al. Chinese yam polysaccharide enhances growth performance and cellular immune response in weanling rats. J. Sci. Food Agric. 2009, 89, 2039–2044. [Google Scholar] [CrossRef]
- Donath, M.Y.; Dalmas, E.; Sauter, N.S.; Boni-Schnetzler, M. Inflammation in obesity and diabetes: Islet dysfunction and therapeutic opportunity. Cell Metab. 2013, 17, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Gil, H.W.; Lee, E.Y.; Lee, J.H.; Kim, Y.S.; Lee, B.E.; Suk, J.W.; Song, H.Y. Dioscorea batatas extract attenuates high-fat diet-induced obesity in mice by decreasing expression of inflammatory cytokines. Med. Sci. Monitor 2015, 21, 489–495. [Google Scholar]
- Selvin, E.; Steffes, M.W.; Zhu, H.; Matsushita, K.; Wagenknecht, L.; Pankow, J.; Coresh, J.; Brancati, F.L. Glycated hemoglobin, diabetes, and cardiovascular risk in nondiabetic adults. N. Engl. J. Med. 2010, 362, 800–811. [Google Scholar] [CrossRef] [PubMed]
- VanBuecken, D.E.; Greenbaum, C.J. Residual C-peptide in type 1 diabetes: What do we really know? Pediatr. Diabetes 2014, 15, 84–90. [Google Scholar] [CrossRef] [PubMed]
- McKoy, M.L.; Grant, K.; Asemota, H.; Simon, O.; Omoruyi, F. Renal and hepatic function in hypercholesterolemic rats fed jamaican bitter yam (Dioscorea polygonoides). J. Diet. Suppl. 2015, 12, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhou, M.-N.; Lu, Q-J. Effects of yam polysaccharides on P-selectin expression and macrophage infiltration in diabetic nephropathy model rats. Clin. Exp. Med. Sci. 2015, 3, 11–21. [Google Scholar]
- Westerberg, D.P. Diabetic ketoacidosis: Evaluation and treatment. Am. Fam. Physician 2013, 87, 337–346. [Google Scholar] [PubMed]
- Jakobsen, J.; Knudsen, G.M.; Juhler, M. Cation permeability of the blood-brain barrier in streptozotocin-diabetic rats. Diabetologia 1987, 30, 409–413. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Go, H.-K.; Rahman, M.M.; Kim, G.-B.; Na, C.-S.; Song, C.-H.; Kim, J.-S.; Kim, S.-J.; Kang, H.-S. Antidiabetic Effects of Yam (Dioscorea batatas) and Its Active Constituent, Allantoin, in a Rat Model of Streptozotocin-Induced Diabetes. Nutrients 2015, 7, 8532-8544. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7105411
Go H-K, Rahman MM, Kim G-B, Na C-S, Song C-H, Kim J-S, Kim S-J, Kang H-S. Antidiabetic Effects of Yam (Dioscorea batatas) and Its Active Constituent, Allantoin, in a Rat Model of Streptozotocin-Induced Diabetes. Nutrients. 2015; 7(10):8532-8544. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7105411
Chicago/Turabian StyleGo, Hyeon-Kyu, Md. Mahbubur Rahman, Gi-Beum Kim, Chong-Sam Na, Choon-Ho Song, Jin-Shang Kim, Shang-Jin Kim, and Hyung-Sub Kang. 2015. "Antidiabetic Effects of Yam (Dioscorea batatas) and Its Active Constituent, Allantoin, in a Rat Model of Streptozotocin-Induced Diabetes" Nutrients 7, no. 10: 8532-8544. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7105411