Hepatoprotective Effect of Herb Formula KIOM2012H against Nonalcoholic Fatty Liver Disease

Abstract
:1. Introduction
2. Experimental Section
2.1. Herb Materials and Preparation of KIOM2012H

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Family Name | Latin Name | Source | Amount (g) |
|---|---|---|---|---|
| Arctium lappa Linne | Compositae | Arctii Semen | China | 9.0 |
| Glycyrrhiza uralensis Fischer | Leguminosae | Glycyrrhizae Radix | China | 7.5 |
| Magnolia officinalis Rehder & Wilson | Magnoliaceae | Magnoliae Cortex | China | 6.0 |
| Zingiber officinale Roscoe | Zingiberaceae | Zingiberis Rhizoma | Korea | 7.5 |
| Total | 30.0 | |||
2.2. Cell Culture
2.3. High Performance Liquid Chromatography (HPLC) Analysis of KIOM2012H
2.4. Cell Viability Assay
2.5. FFA-Induced Lipid Accumulation and Measurement of Intracellular Triglyceride Content
2.6. Nile Red and Oil Red O Staining
2.7. RNA Isolation and qRT-PCR
2.8. Measurement of Fatty Acid Uptake
2.9. In Vivo Animal Experiment
2.10. Statistical Analysis
3. Results
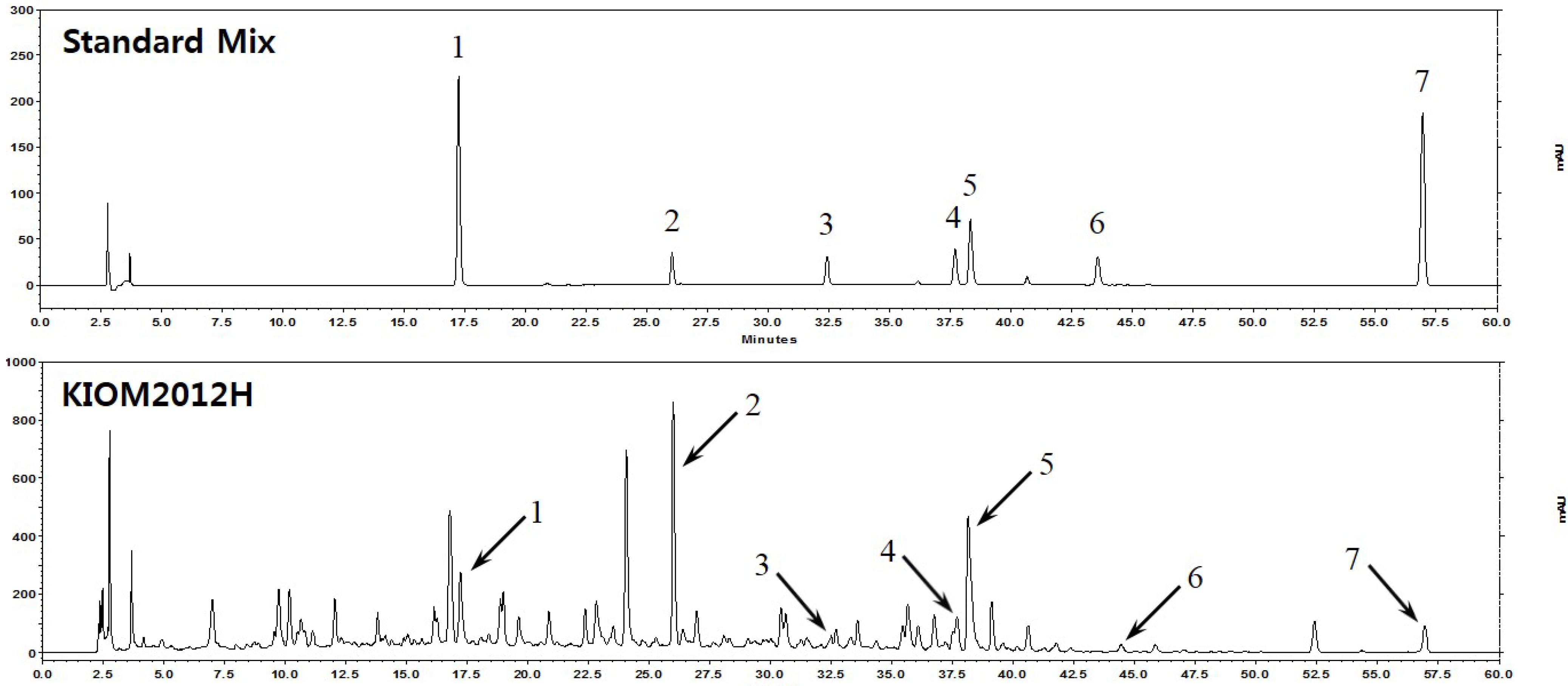
3.1. HPLC Fingerprint of KIOM2012H

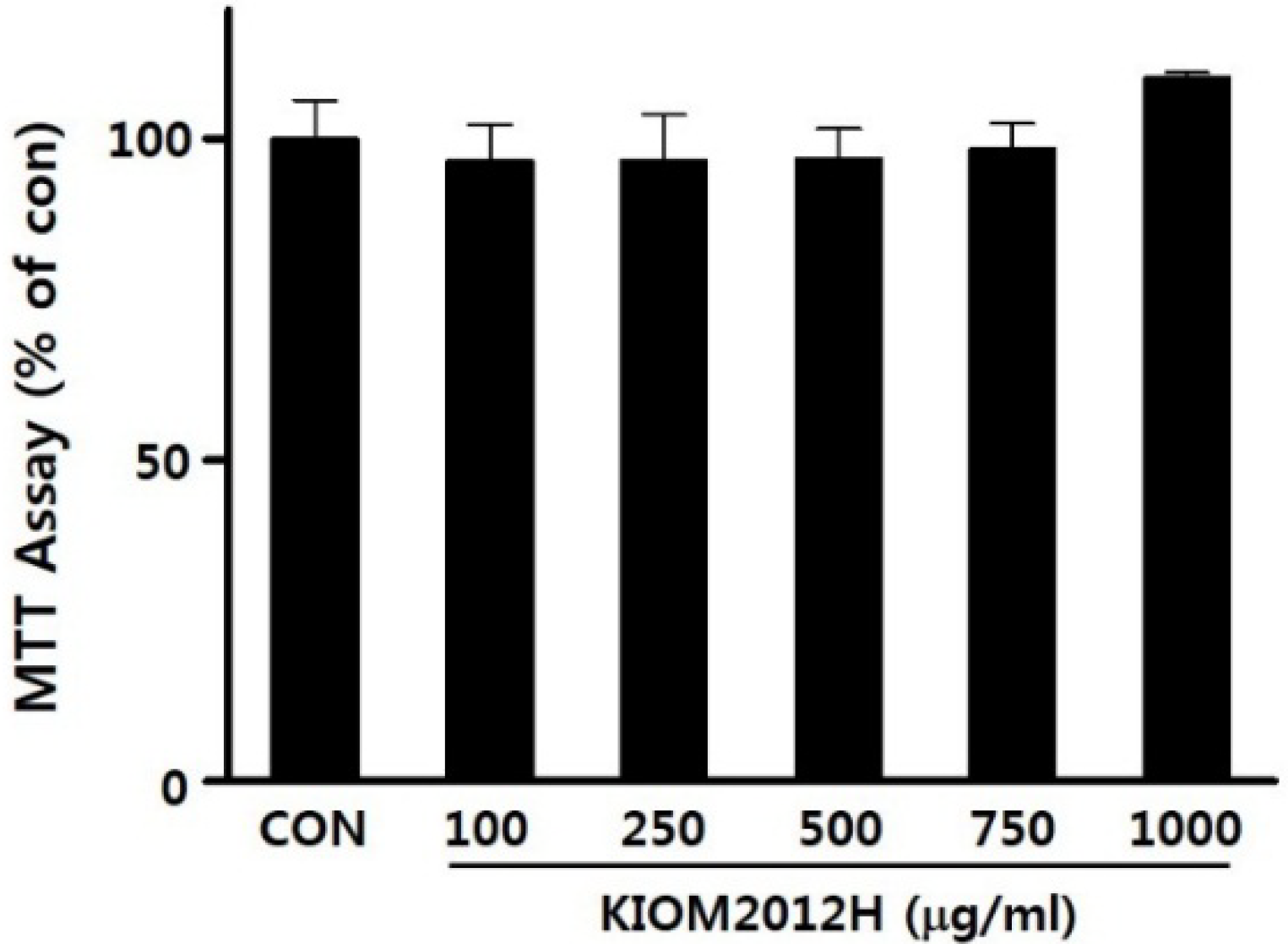
3.2. Cytotoxicity of KIOM2012H

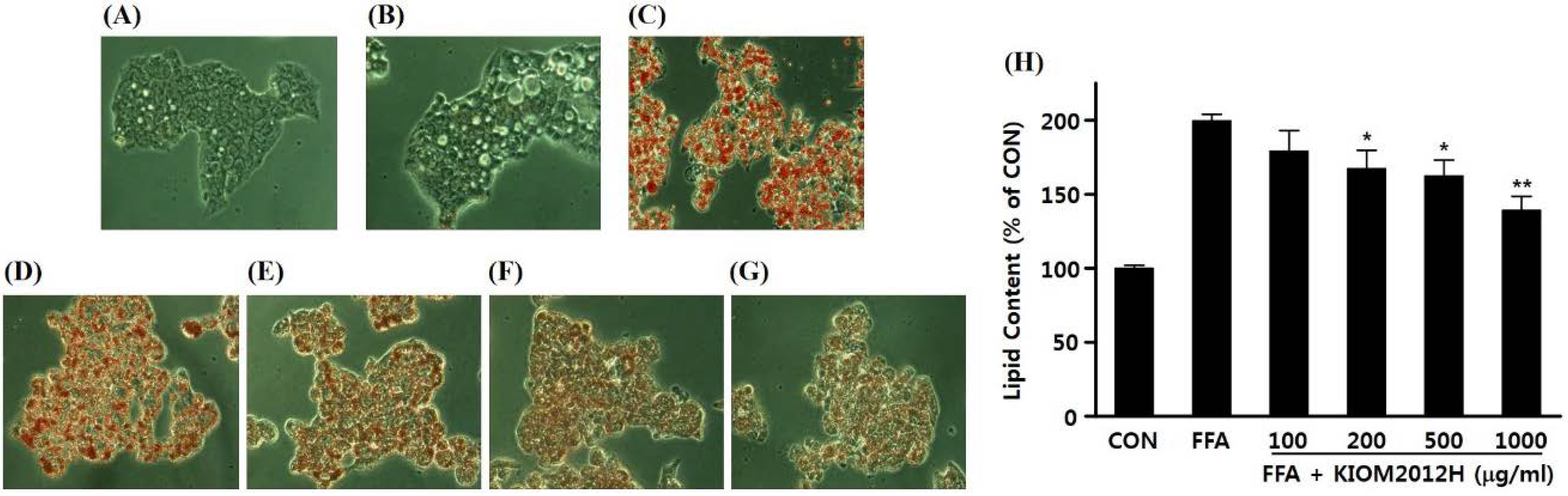
3.3. Effect of KIOM2012H on FFA-Induced Lipid Accumulation in HepG2 Cells


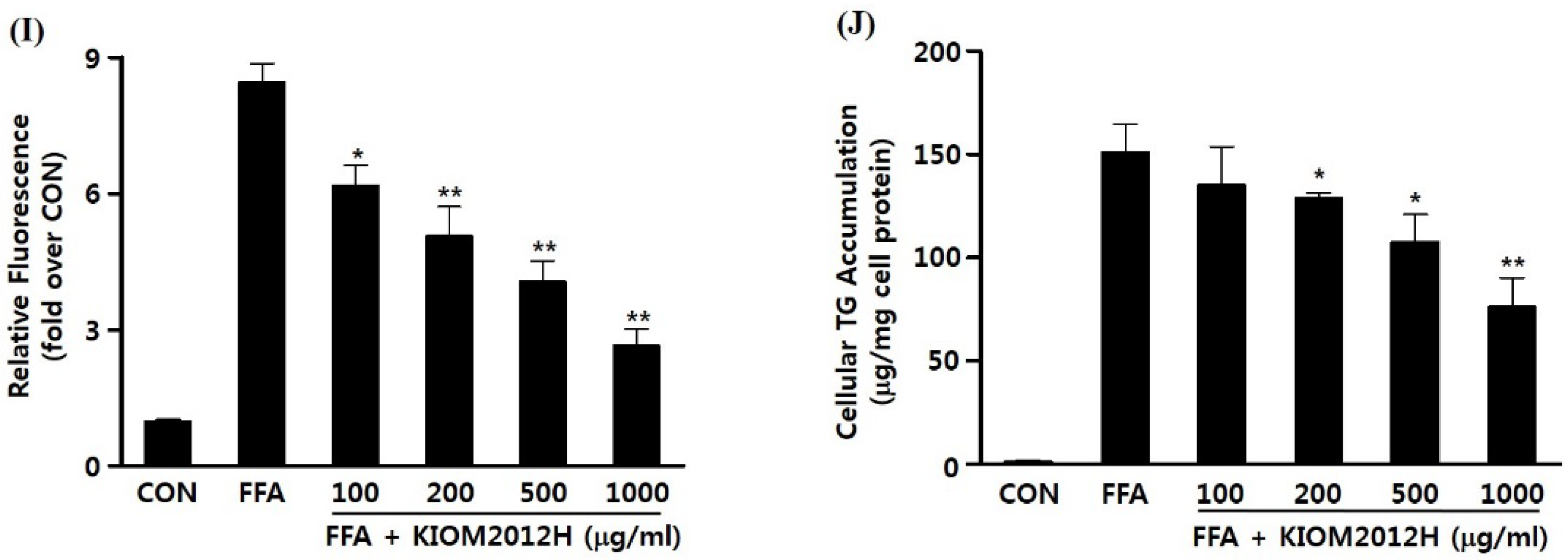
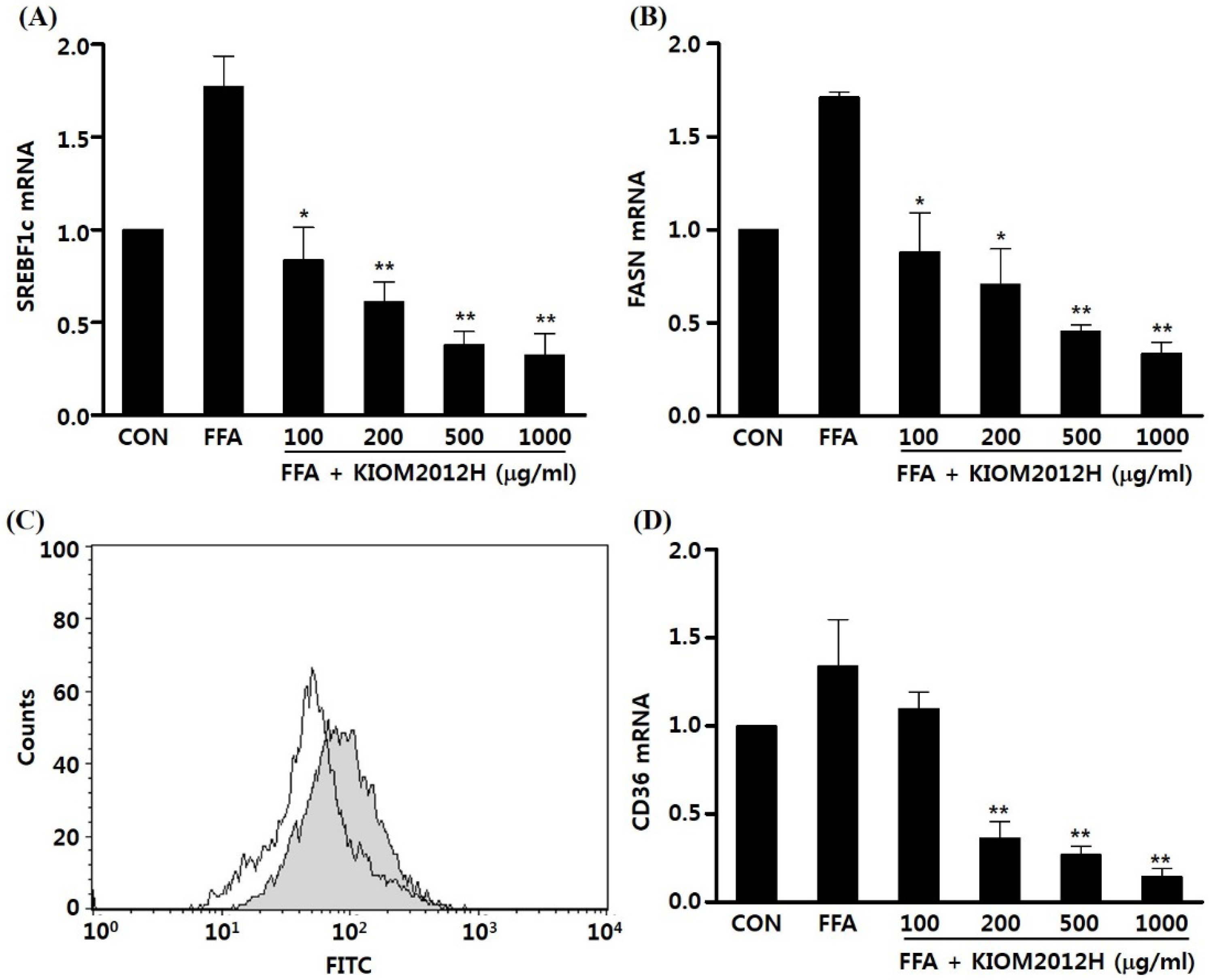
3.4. KIOM2012H Inhibited Gene Expression and Lipid Uptake

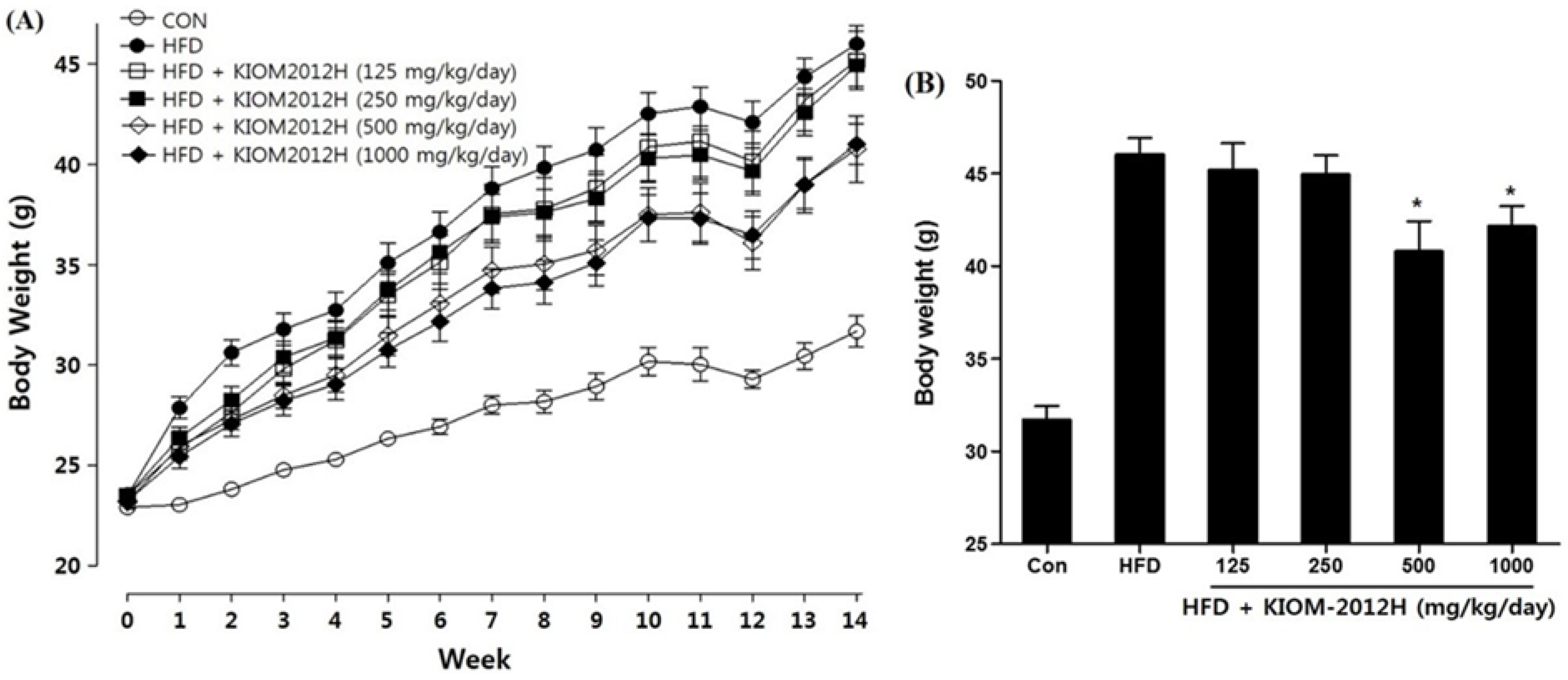
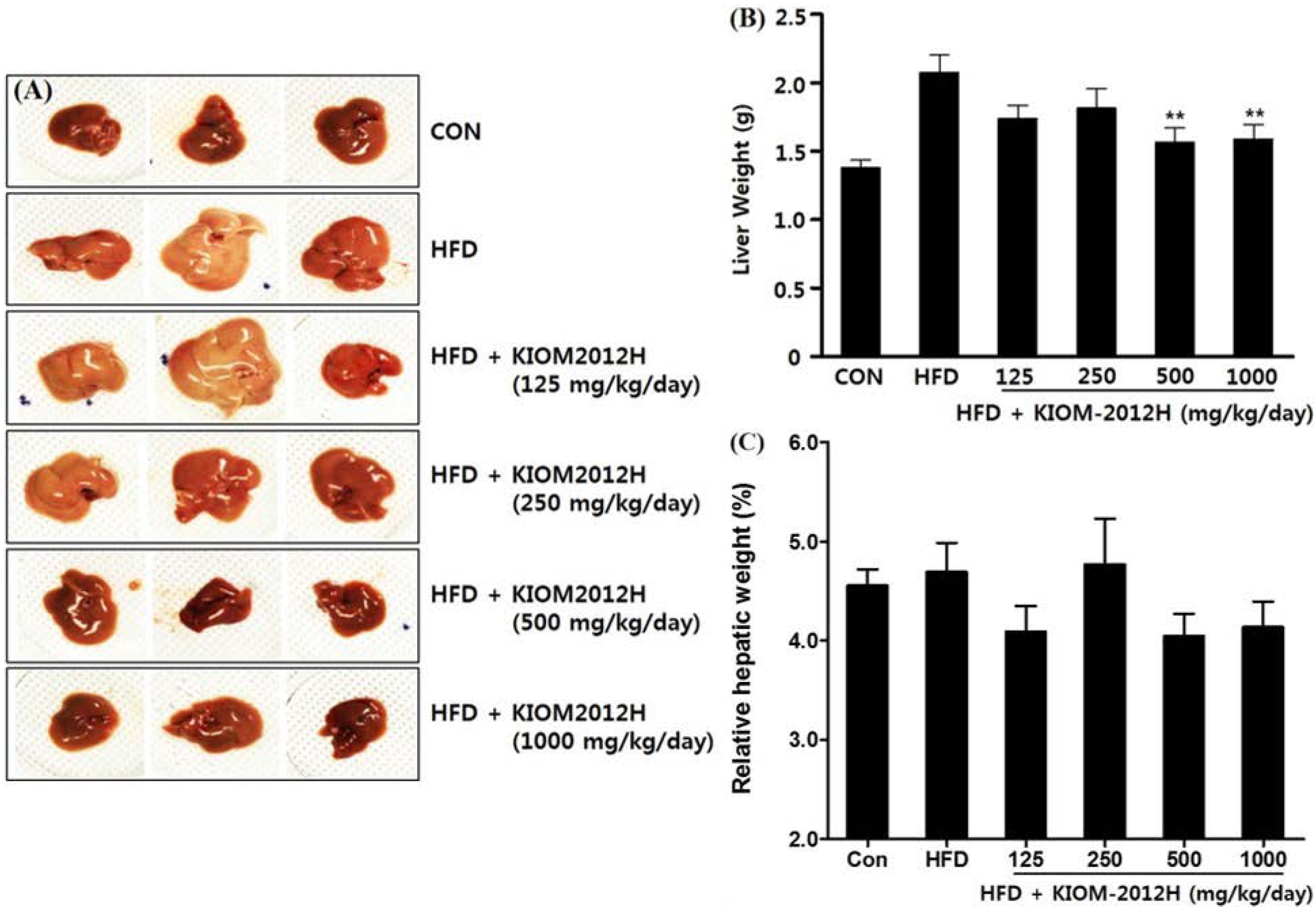
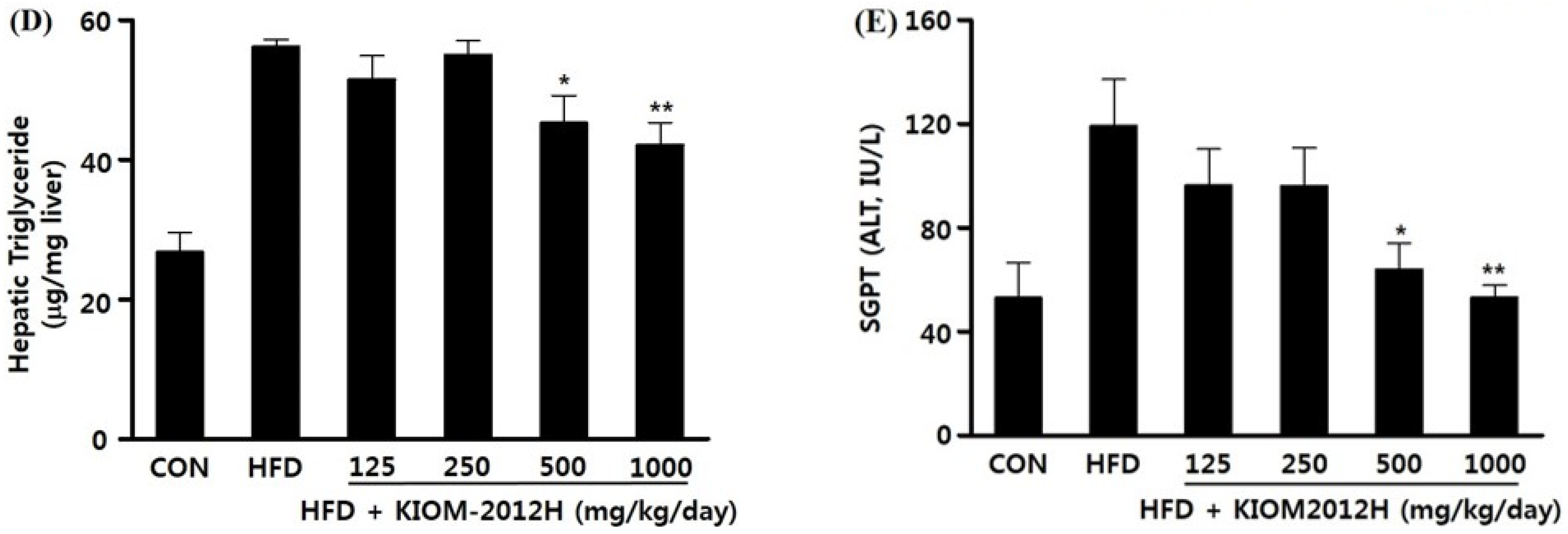
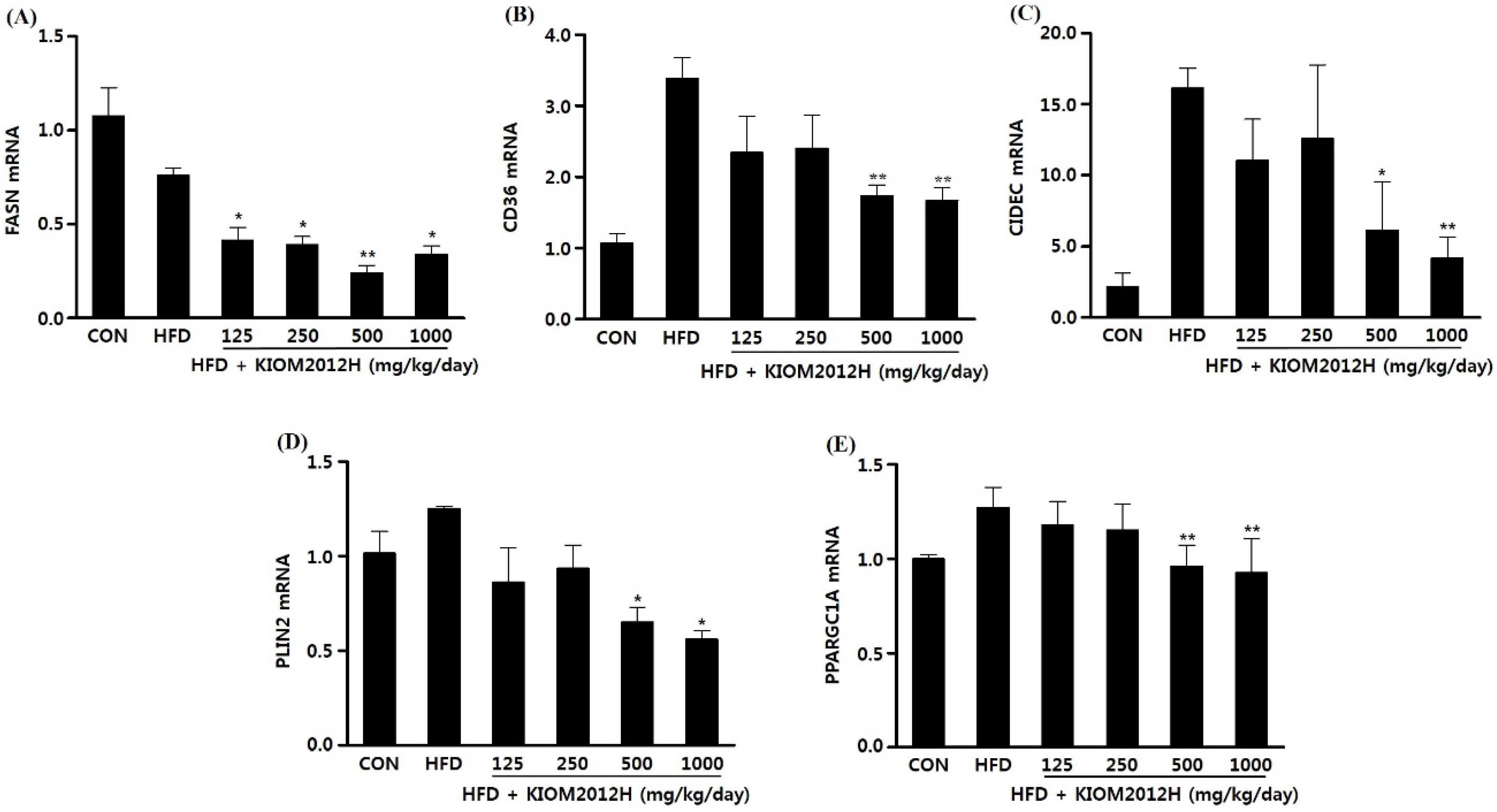
3.5. KIOM2012H Effects in HFD-Fed Mice




4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Adams, L.A.; Angulo, P.; Lindor, K.D. Nonalcoholic fatty liver disease. Can. Med. Assoc. J. 2005, 172, 899–905. [Google Scholar] [CrossRef]
- Bedogni, G.; Nobili, V.; Tiribelli, C. Epidemiology of fatty liver: An update. World J. Gastroenterol. 2014, 20, 9050–9054. [Google Scholar] [PubMed]
- Younossi, Z.M.; Stepanova, M.; Afendy, M.; Fang, Y.; Younossi, Y.; Mir, H.; Srishord, M. Changes in the prevalence of the most common causes of chronic liver diseases in the United States from 1988 to 2008. Clin. Gastroenterol. Hepatol. 2011, 9, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Schwimmer, J.B.; Deutsch, R.; Kahen, T.; Lavine, J.E.; Stanley, C.; Behling, C. Prevalence of fatty liver in children and adolescents. Pediatrics 2006, 118, 1388–1393. [Google Scholar] [CrossRef] [PubMed]
- Than, N.N.; Newsome, P.N. A concise review of non-alcoholic fatty liver disease. Atherosclerosis 2015, 239, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, K.L.; Smith, C.I.; Schwarzenberg, S.J.; Jessurun, J.; Boldt, M.D.; Parks, E.J. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J. Clin. Investig. 2005, 115, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, G.; Scopacasa, F.; Colao, A.; Capone, D.; Tarantino, M.; Grimaldi, E.; Savastano, S. Serum Bcl-2 concentrations in overweight-obese subjects with nonalcoholic fatty liver disease. World J. Gastroenterol. 2011, 17, 5280–5288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begriche, K.; Igoudjil, A.; Pessayre, D.; Fromenty, B. Mitochondrial dysfunction in NASH: Causes, consequences and possible means to prevent it. Mitochondrion 2006, 6, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, G.; Colao, A.; Capone, D.; Conca, P.; Tarantino, M.; Grimaldi, E.; Chianese, D.; Finelli, C.; Contaldo, F.; Scopacasa, F.; et al. Circulating levels of cytochrome C, gamma-glutamyl transferase, triglycerides and unconjugated bilirubin in overweight/obese patients with non-alcoholic fatty liver disease. J. Biol. Regul. Homeost. Agents 2011, 25, 47–56. [Google Scholar] [PubMed]
- Tamura, S.; Shimomura, I. Contribution of adipose tissue and de novo lipogenesis to nonalcoholic fatty liver disease. J. Clin. Investig. 2005, 115, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Awale, S.; Lu, J.; Kalauni, S.K.; Kurashima, Y.; Tezuka, Y.; Kadota, S.; Esumi, H. Identification of arctigenin as an antitumor agent having the ability to eliminate the tolerance of cancer cells to nutrient starvation. Cancer Res. 2006, 66, 1751–1757. [Google Scholar] [CrossRef] [PubMed]
- Hyam, S.R.; Lee, I.A.; Gu, W.; Kim, K.A.; Jeong, J.J.; Jang, S.E.; Han, M.J.; Kim, D.H. Arctigenin ameliorates inflammation in vitro and in vivo by inhibiting the PI3K/AKT pathway and polarizing M1 macrophages to M2-like macrophages. Eur. J. Pharmacol. 2013, 708, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Guo, L.P.; Hu, X.L.; Huang, J.; Fan, Y.H.; Ren, T.S.; Zhao, Q.C. Protective effects of Arctium lappa L. roots against hydrogen peroxide-induced cell injury and potential mechanisms in SH-SY5Y cells. Cell. Mol. Neurobiol. 2014. [Google Scholar] [CrossRef]
- Shin, Y.W.; Bae, E.A.; Lee, B.; Lee, S.H.; Kim, J.A.; Kim, Y.S.; Kim, D.H. In vitro and in vivo antiallergic effects of Glycyrrhiza glabra and its components. Planta Med. 2007, 73, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Seon, M.R.; Park, S.Y.; Kwon, S.J.; Lim, S.S.; Choi, H.J.; Park, H.; Lim, D.Y.; Kim, J.S.; Lee, C.H.; Kim, J.; et al. Hexane/ethanol extract of Glycyrrhiza uralensis and its active compound isoangustone A induce G1 cycle arrest in DU145 human prostate and 4T1 murine mammary cancer cells. J. Nutr. Biochem. 2012, 23, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Adianti, M.; Aoki, C.; Komoto, M.; Deng, L.; Shoji, I.; Wahyuni, T.S.; Lusida, M.I.; Soetjipto; Fuchino, H.; Kawahara, N.; et al. Anti-hepatitis C virus compounds obtained from Glycyrrhiza uralensis and other Glycyrrhiza species. Microbiol. Immunol. 2014, 58, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Chen, J.; Li, Y.J.; Zheng, Y.F.; Li, P. Antioxidant and anti-inflammatory activities of six flavonoids separated from licorice. Food Chem. 2013, 141, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Liu, Y.; Huang, Y.; Sun, J.; Wu, Z.; Zhang, X.; Ping, Q. Glycyrrhizin surface-modified chitosan nanoparticles for hepatocyte-targeted delivery. Int. J. Pharm. 2008, 359, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.S.; Hong, S.W.; Yeon, S.H.; Kim, Y.M.; Um, K.A.; Kim, J.H.; Kim, H.J.; Chang, K.C.; Park, S.W. Magnolia officinalis attenuates free fatty acid-induced lipogenesis via AMPK phosphorylation in hepatocytes. J. Ethnopharmacol. 2014, 157, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.Q.; Je, Y.T.; Kim, Y.C.; Shin, Y.K.; Sung, S.; Lee, K.; Jeong, G.S.; Kim, Y.C.; Lee, B.H. Magnolia officinalis reverses alcoholic fatty liver by inhibiting the maturation of sterol regulatory element-binding protein-1c. J. Pharmacol. Sci. 2009, 109, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Liu, Y.; Wang, X.; Jiao, R.; Ma, K.Y.; Li, Y.M.; Wang, L.; Man, S.W.; Sang, S.; Huang, Y.; et al. Plasma cholesterol-lowering activity of gingerol- and shogaol-enriched extract is mediated by increasing sterol excretion. J. Agric. Food Chem. 2014, 62, 10515–10521. [Google Scholar] [CrossRef] [PubMed]
- Nammi, S.; Sreemantula, S.; Roufogalis, B.D. Protective effects of ethanolic extract of Zingiber officinale rhizome on the development of metabolic syndrome in high-fat diet-fed rats. Basic Clin. Pharmacol. Toxicol. 2009, 104, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, B.; Rosenblat, M.; Hayek, T.; Coleman, R.; Aviram, M. Ginger extract consumption reduces plasma cholesterol, inhibits LDL oxidation and attenuates development of atherosclerosis in atherosclerotic, apolipoprotein E-deficient mice. J. Nutr. 2000, 130, 1124–1131. [Google Scholar] [PubMed]
- Yim, N.H.; Cho, W.K.; Lee, J.H.; Jung, Y.P.; Yang, H.J.; Ma, J.Y. HRT, Herbal formula, induces G(2)/M cell cycle arrest and apoptosis via suppressing Akt signaling pathway in human colon cancer cells. Evid. Based Complement. Altern. Med. 2012, 2012, 871893. [Google Scholar] [CrossRef]
- Greenspan, P.; Fowler, S.D. Spectrofluorometric studies of the lipid probe, Nile Red. J. Lipid Res. 1985, 26, 781–789. [Google Scholar] [PubMed]
- McMillian, M.K.; Grant, E.R.; Zhong, Z.; Parker, J.B.; Li, L.; Zivin, R.A.; Burczynski, M.E.; Johnson, M.D. Nile Red binding to HepG2 cells: An improved assay for in vitro studies of hepatosteatosis. In Vitro Mol. Toxicol. 2001, 14, 177–190. [Google Scholar] [CrossRef]
- Song, K.H.; Chiang, J.Y. Glucagon and cAMP inhibit cholesterol 7α-hydroxylase (CYP7A1) gene expression in human hepatocytes: Discordant regulation of bile acid synthesis and gluconeogenesis. Hepatology 2006, 43, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Erickson, S.K. Nonalcoholic fatty liver disease. J. Lipid Res. 2009, 50, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Grefhorst, A.; Elzinga, B.M.; Voshol, P.J.; Plösch, T.; Kok, T.; Bloks, V.W.; van der Sluijs, F.H.; Havekes, L.M.; Romijn, J.A.; Verkade, H.J.; et al. Stimulation of lipogenesis by pharmacological activation of the liver X receptor leads to production of large, triglyceride-rich very low density lipoprotein particles. J. Biol. Chem. 2002, 277, 34182–34190. [Google Scholar] [CrossRef] [PubMed]
- Greco, D.; Kotronen, A.; Westerbacka, J.; Puig, O.; Arkkila, P.; Kiviluoto, T.; Laitinen, S.; Kolak, M.; Fisher, R.M.; Hamsten, A.; et al. Gene expression in human NAFLD. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1281–G1287. [Google Scholar] [CrossRef] [PubMed]
- Miquilena-Colina, M.E.; Lima-Cabello, E.; Sánchez-Campos, S.; García-Mediavilla, M.V.; Fernández-Bermejo, M.; Lozano-Rodríguez, T.; Vargas-Castrillón, J.; Buqué, X.; Ochoa, B.; Aspichueta, P.; et al. Hepatic fatty acid translocase CD36 upregulation is associated with insulin resistance, hyperinsulinaemia and increased steatosis in non-alcoholic steatohepatitis and chronic hepatitis C. Gut 2011, 60, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Febbraio, M.; Wada, T.; Zhai, Y.; Kuruba, R.; He, J.; Lee, J.H.; Khadem, S.; Ren, S.; Li, S.; et al. Hepatic fatty acid transporter Cd36 is a common target of LXR, PXR, and PPARγ in promoting steatosis. Gastroenterology 2008, 134, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Zhao, M.; Xu, Z.; Yokoyama, K.K.; Li, T. Molecular cloning and characterization of CIDE-3, a novel member of the cell-death-inducing DNA-fragmentation-factor (DF45)-like effector family. Biochem. J. 2003, 370, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Keller, P.; Petrie, J.T.; de Rose, P.; Gerin, I.; Wright, W.S.; Chiang, S.H.; Nielsen, A.R.; Fischer, C.P.; Pedersen, B.K.; MacDougald, O.A. Fat-specific protein 27 regulates storage of triacylglycerol. J. Biol. Chem. 2008, 283, 14355–14365. [Google Scholar] [CrossRef] [PubMed]
- Puri, V.; Konda, S.; Ranjit, S.; Aouadi, M.; Chawla, A.; Chouinard, M.; Chakladar, A.; Czech, M.P. Fat-specific protein 27, a novel lipid droplet protein that enhances triglyceride storage. J. Biol. Chem. 2007, 282, 34213–34218. [Google Scholar] [CrossRef] [PubMed]
- Van Rossum, T.G.; Vulto, A.G.; de Man, R.A.; Brouwer, J.T.; Schalm, S.W. Review article: Glycyrrhizin as a potential treatment for chronic hepatitis C. Aliment. Pharmacol. Ther. 1998, 12, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Irii, H.; Tahara, Y.; Ishii, H.; Hirao, A.; Udagawa, H.; Hiramoto, M.; Yasuda, K.; Takanishi, A.; Shibata, S.; et al. Synthesis of a new [6]-gingerol analogue and its protective effect with respect to the development of metabolic syndrome in mice fed a high-fat diet. J. Med. Chem. 2011, 54, 6295–6304. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.; Hwang, Y.-H.; Kim, D.-G.; Jeon, J.; Ma, J.Y. Hepatoprotective Effect of Herb Formula KIOM2012H against Nonalcoholic Fatty Liver Disease. Nutrients 2015, 7, 2440-2455. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7042440
Park H, Hwang Y-H, Kim D-G, Jeon J, Ma JY. Hepatoprotective Effect of Herb Formula KIOM2012H against Nonalcoholic Fatty Liver Disease. Nutrients. 2015; 7(4):2440-2455. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7042440
Chicago/Turabian StylePark, Hwayong, Youn-Hwan Hwang, Dong-Gun Kim, Jongwook Jeon, and Jin Yeul Ma. 2015. "Hepatoprotective Effect of Herb Formula KIOM2012H against Nonalcoholic Fatty Liver Disease" Nutrients 7, no. 4: 2440-2455. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7042440