Proanthocyanidins Prevent High Glucose-Induced Eye Malformation by Restoring Pax6 Expression in Chick Embryo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Animals and Treatments
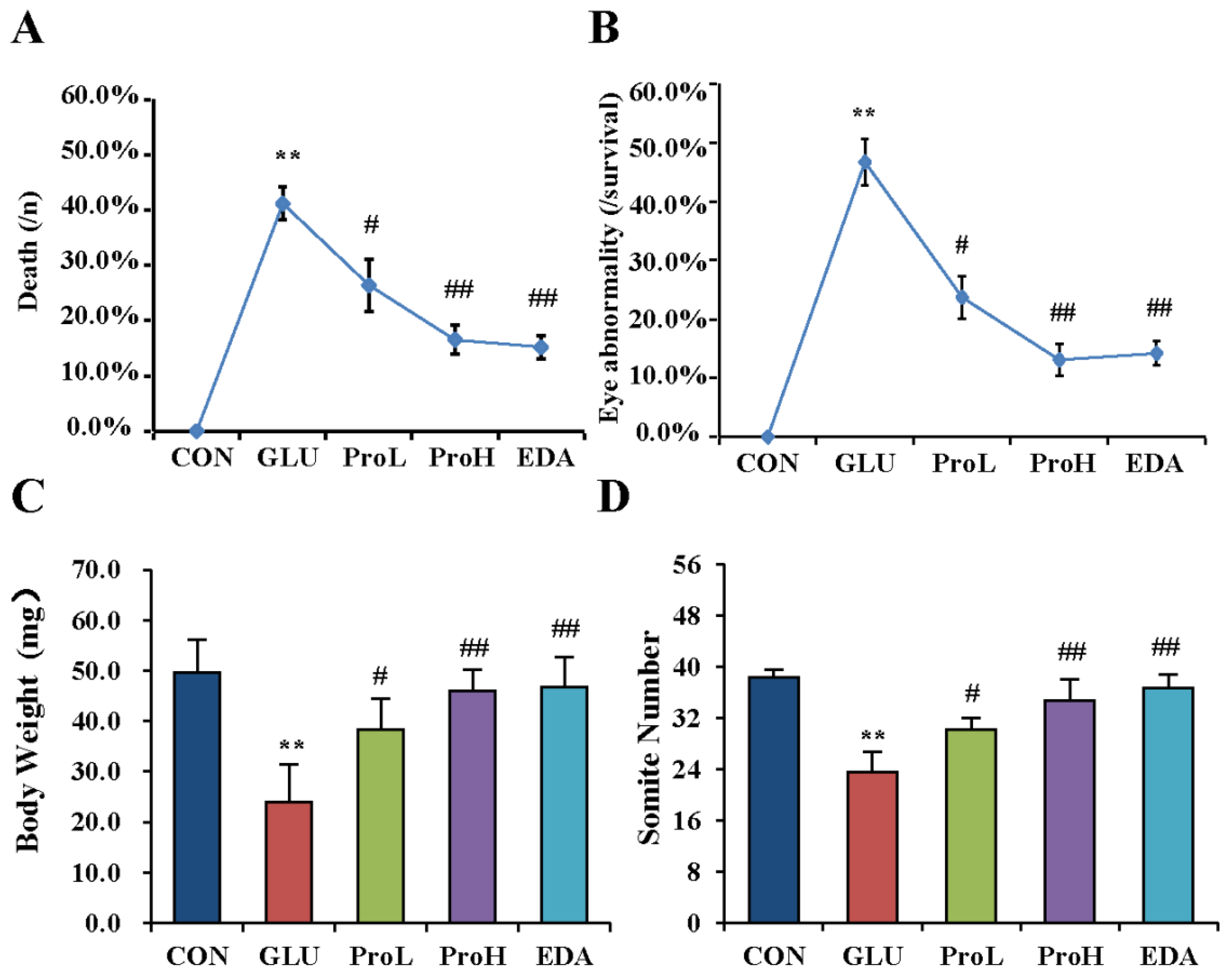
2.2. Mortality, Abnormality, Body Weight and Somite Number Measurements
2.3. Eye Glucose Measurements
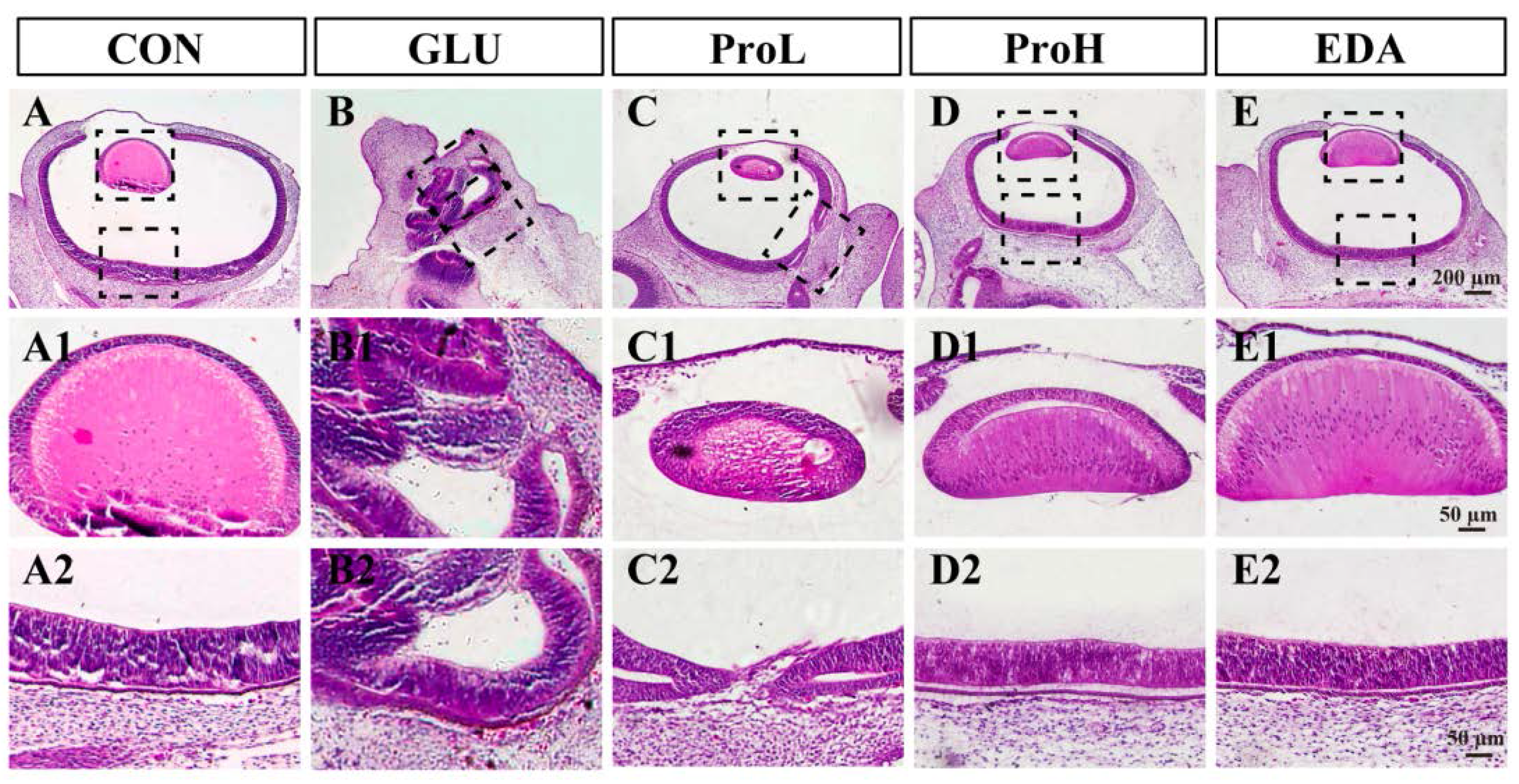
2.4. Histological Analysis
2.5. Measurements of MDA Contents, SOD and GSH-PX Activities and ROS Generation Ratio
2.6. Quantitative PCR
2.7. Western Blotting Analysis
2.8. Statistical Analysis
3. Results
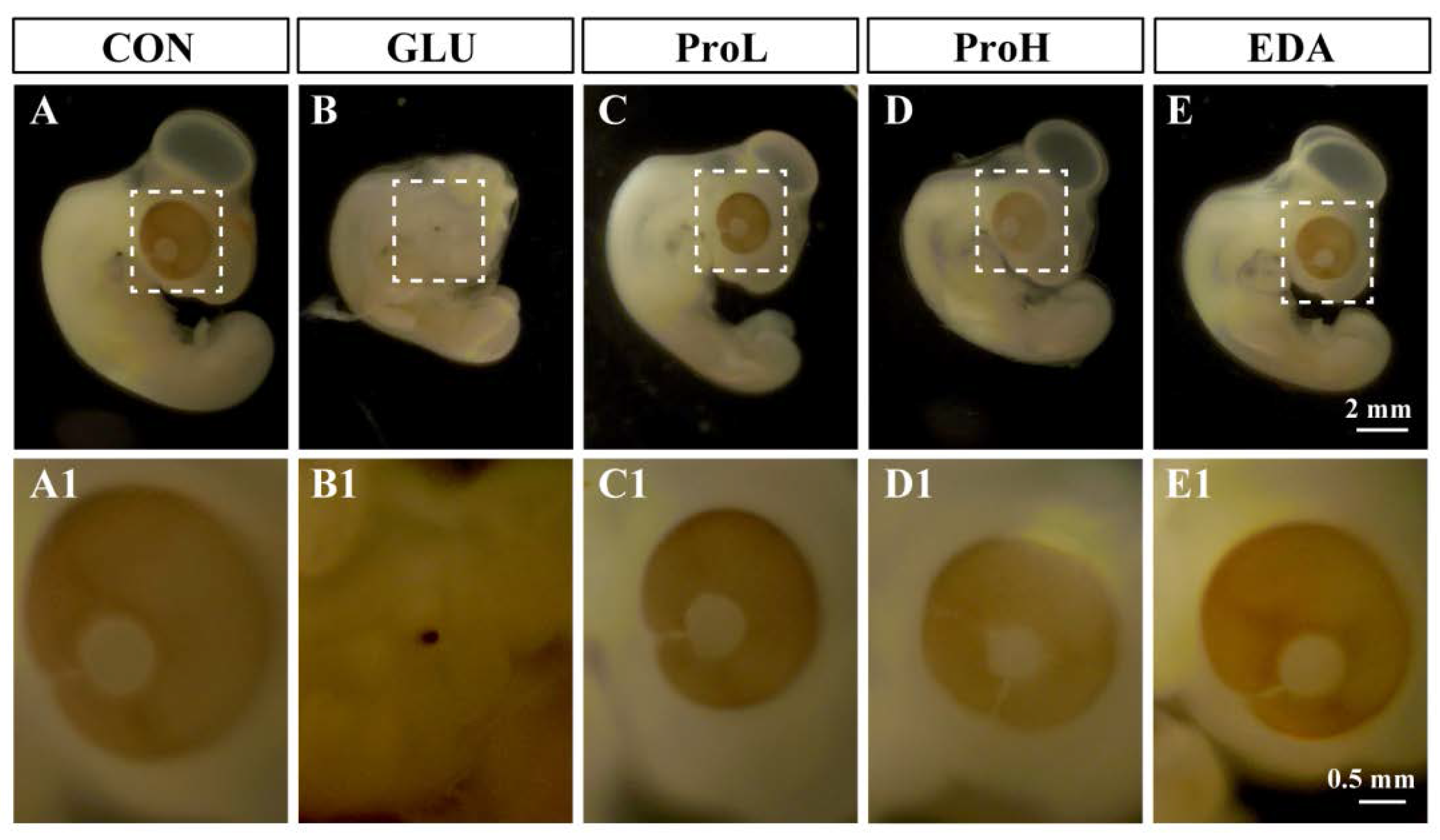
3.1. Proanthocyanidins Ameliorate High Glucose-Induced Eye Malformation in Chick Embryos



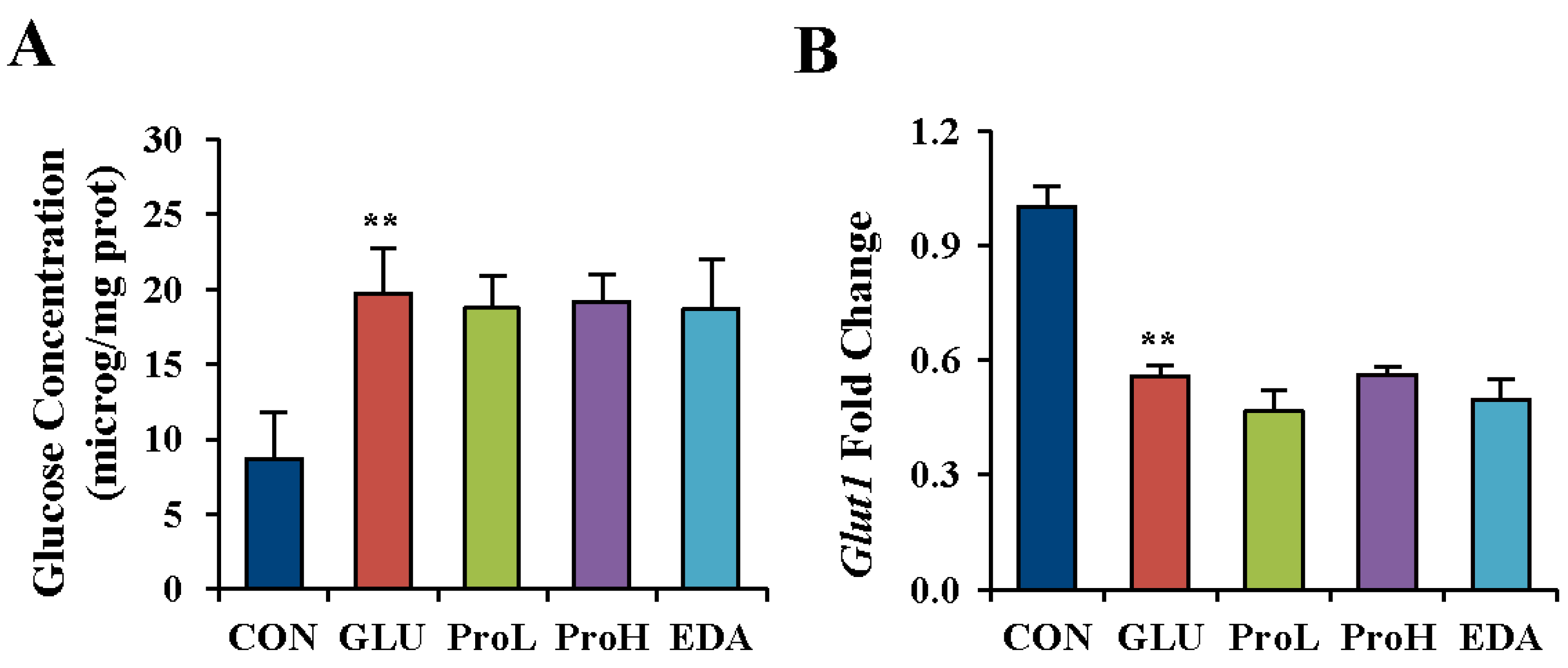
3.2. Proanthocyanidins have No Effects on Glucose Concentration in High Glucose-Treated Eye of Chick Embryos

3.3. Proanthocyanidins Restore the Expression of Eye Development Marker in the Eye of Chick Embryos

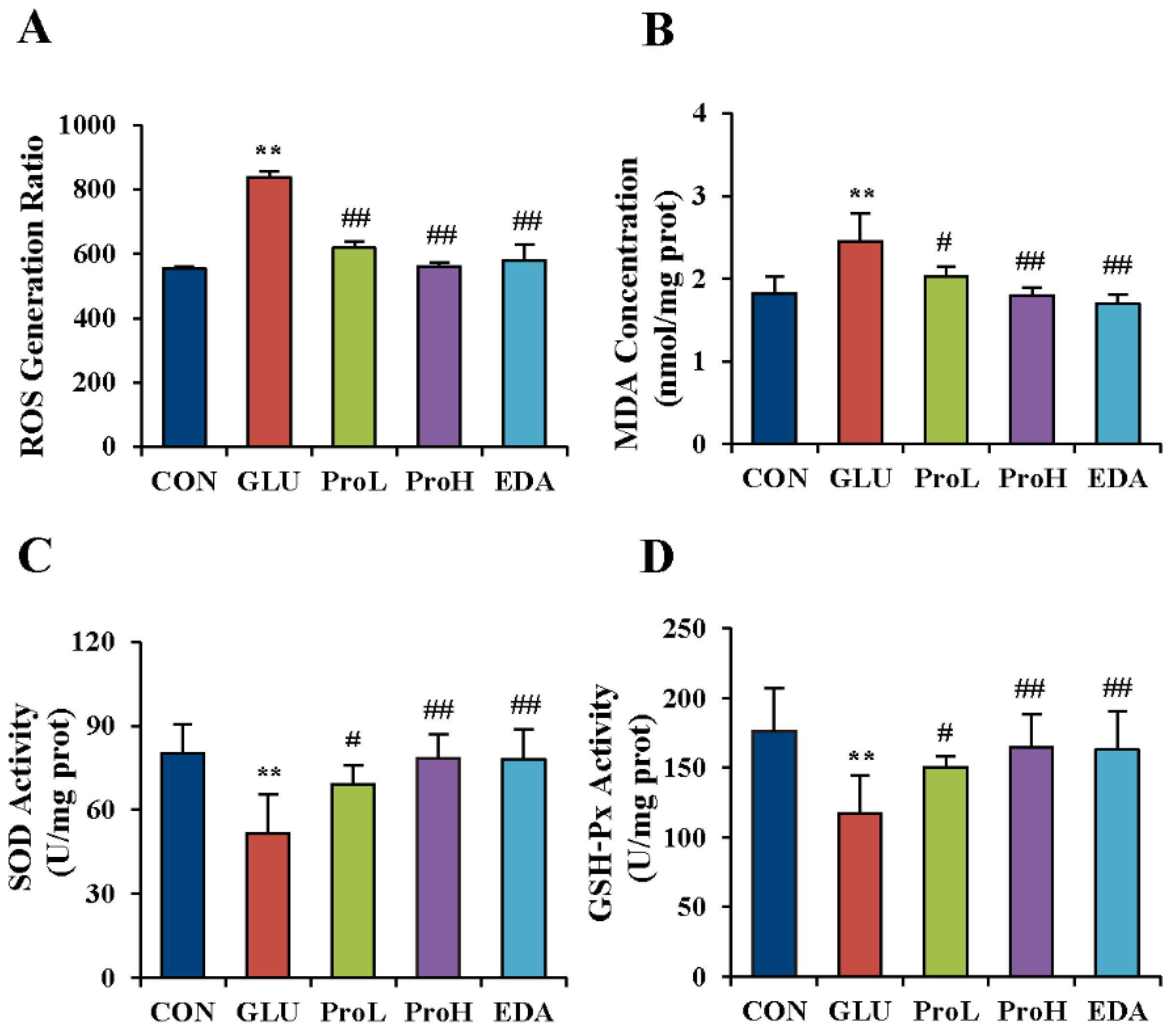
3.4. Proanthocyanidins Ameliorated the Oxidative Stress State Induced by High Glucose in the Eye of Chick Embryos

4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Crowther, C.A.; Hiller, J.E.; Moss, J.R.; McPhee, A.J.; Jeffries, W.S.; Robinson, J.S.; Grp, A.T. Effect of treatment of gestational diabetes mellitus on pregnancy outcomes. N. Engl. J. Med. 2005, 352, 2477–2486. [Google Scholar] [CrossRef] [PubMed]
- Suhonen, L.; Hiilesmaa, V.; Kaaja, R.; Teramo, K. Detection of pregnancies with high risk of fetal macrosomia among women with gestational diabetes mellitus. Acta Obstet. Gynecol. Scand. 2008, 87, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Flores-le Roux, J.A.; Sagarra, E.; Benaiges, D.; Hernandez-Rivas, E.; Chillaron, J.J.; de Dou, J.P.; Mur, A.; Lopez-Vilchez, M.A.; Pedro-Botet, J. A prospective evaluation of neonatal hypoglycaemia in infants of women with gestational diabetes mellitus. Diabetes Res. Clin. Pract. 2012, 97, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Mitanchez, D. Foetal and neonatal complications in gestational diabetes: Perinatal mortality, congenital malformations, macrosomia, shoulder dystocia, birth injuries, neonatal complications. Diabetes Metab. 2010, 36, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Tundidor, D.; Garcia-Patterson, A.; Maria, M.A.; Ubeda, J.; Ginovart, G.; Adelantado, J.M.; de Leiva, A.; Corcoy, R. Perinatal maternal and neonatal outcomes in women with gestational diabetes mellitus according to fetal sex. Gend. Med. 2012, 9, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Landau, K.; Bajka, J.D.; Kirchschlager, B.M. Topless optic disks in children of mothers with type i diabetes mellitus. Am. J. Ophthalmol. 1998, 125, 605–611. [Google Scholar] [CrossRef]
- Tariq, Y.M.; Samarawickrama, C.; Li, H.; Huynh, S.C.; Burlutsky, G.; Mitchell, P. Retinal thickness in the offspring of diabetic pregnancies. Am. J. Ophthalmol. 2010, 150, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Ashery-Padan, R.; Gruss, P. Pax6 lights-up the way for eye development. Curr. Opin. Cell Biol. 2001, 13, 706–714. [Google Scholar] [CrossRef]
- Grindley, J.C.; Davidson, D.R.; Hill, R.E. The role of pax-6 in eye and nasal development. Development 1995, 121, 1433–1442. [Google Scholar] [PubMed]
- Koroma, B.M.; Yang, J.M.; Sundin, O.H. The pax-6 homeobox gene is expressed throughout the corneal and conjunctival epithelia. Investig. Ophthalmol. Vis. Sci. 1997, 38, 108–120. [Google Scholar]
- Hill, R.E.; Favor, J.; Hogan, B.L.; Ton, C.C.; Saunders, G.F.; Hanson, I.M.; Prosser, J.; Jordan, T.; Hastie, N.D.; van Heyningen, V. Mouse small eye results from mutations in a paired-like homeobox-containing gene. Nature 1991, 354, 522–525. [Google Scholar] [CrossRef] [PubMed]
- Glaser, T.; Walton, D.S.; Maas, R.L. Genomic structure, evolutionary conservation and aniridia mutations in the human pax6 gene. Nat. Genet. 1992, 2, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Prosser, J.; van Heyningen, V. Pax6 mutations reviewed. Hum. Mutat. 1998, 11, 93–108. [Google Scholar] [CrossRef]
- Xiao, X.; Li, S.; Zhang, Q. Microphthalmia, late onset keratitis, and iris coloboma/aniridia in a family with a novel pax6 mutation. Ophthalmic Genet. 2012, 33, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Natella, F.; Belelli, F.; Gentili, V.; Ursini, F.; Scaccini, C. Grape seed proanthocyanidins prevent plasma postprandial oxidative stress in humans. J. Agric. Food Chem. 2002, 50, 7720–7725. [Google Scholar] [CrossRef] [PubMed]
- Blade, C.; Arola, L.; Salvado, M.-J. Hypolipidemic effects of proanthocyanidins and their underlying biochemical and molecular mechanisms. Mol. Nutr. Food Res. 2010, 54, 37–59. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Zoheir, K.M.A.; Abdel-Hamied, H.E.; Ashour, A.E.; Bakheet, S.A.; Attia, S.M.; Abd-Allah, A.R.A. Grape seed proanthocyanidin extract has potent anti-arthritic effects on collagen-induced arthritis by modifying the t cell balance. Int. Immunopharmacol. 2013, 17, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, V.; Singh, T.; Katiyar, S.K. Multi-targeted prevention and therapy of cancer by proanthocyanidins. Cancer Lett. 2008, 269, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Roca-Rodriguez, M.M.; Lopez-Tinoco, C.; Murri, M.; Fernandez-Deudero, A.; Garcia-Palacios, M.V.; Garcia-Valero, M.A.; Tinahones-Madueno, F.J.; Aguilar-Diosdado, M. Postpartum development of endothelial dysfunction and oxidative stress markers in women with previous gestational diabetes mellitus. J. Endocrinol. Investig. 2014, 37, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, P.C.; Anderson, D.; Gangolli, S.D. Malformations induced in cultured rat embryos by enzymically generated active oxygen species. Teratog. Carcinog. Mutagen. 1986, 6, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.D.; Meeran, S.M.; Katiyar, S.K. Dietary grape seed proanthocyanidins inhibit uvb-induced oxidative stress and activation of mitogen-activated protein kinases and nuclear factor-kappa b signaling in in vivo skh-1 hairless mice. Mol. Cancer Ther. 2007, 6, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Bagchi, M.; Stohs, S.J.; Ray, S.D.; Sen, C.K.; Preuss, H.G. Cellular protection with proanthocyanidins derived from grape seeds. In Alcohol and Wine in Health and Disease; Das, D.K., Ursini, F., Eds.; New York Academy of Sciences: New York, NY, USA, 2002; pp. 260–270. [Google Scholar]
- Lee, Y.A.; Cho, E.J.; Tanaka, T.; Yokozawa, T. Inhibitory activities of proanthocyanidins from persimmon against oxidative stress and digestive enzymes related to diabetes. J. Nutr. Sci. Vitaminol. (Tokyo) 2007, 53, 287–292. [Google Scholar] [CrossRef] [PubMed]
- El-Alfy, A.T.; Ahmed, A.A.E.; Fatani, A.J. Protective effect of red grape seeds proanthocyanidins against induction of diabetes by alloxan in rats. Pharmacol. Res. 2005, 52, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Fukudome, D.; Matsuda, M.; Kawasaki, T.; Ago, Y.; Matsuda, T. The radical scavenger edaravone counteracts diabetes in multiple low-dose streptozotocin-treated mice. Eur. J. Pharmacol. 2008, 583, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Gehring, W.J. New perspectives on eye development and the evolution of eyes and photoreceptors. J. Hered. 2005, 96, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Shaham, O.; Gueta, K.; Mor, E.; Oren-Giladi, P.; Grinberg, D.; Xie, Q.; Cvekl, A.; Shomron, N.; Davis, N.; Keydar-Prizant, M.; et al. Pax6 regulates gene expression in the vertebrate lens through mir-204. PLoS Genet. 2013, 9, e1003357. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Q.C.; Dyer, M.A.; Uchikawa, M.; Kondoh, H.; Lagutin, O.V.; Oliver, G. Six3-mediated auto repression and eye development requires its interaction with members of the groucho-related family of co-repressors. Development 2002, 129, 2835–2849. [Google Scholar] [PubMed]
- Rath, M.F.; Morin, F.; Shi, Q.; Klein, D.C.; Moller, M. Ontogenetic expression of the otx2 and crx homeobox genes in the retina of the rat. Exp. Eye Res. 2007, 85, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Bharti, K.; Liu, W.; Csermely, T.; Bertuzzi, S.; Arnheiter, H. Alternative promoter use in eye development: The complex role and regulation of the transcription factor mitf. Development 2008, 135, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Giudetti, G.; Giannaccini, M.; Biasci, D.; Mariotti, S.; Degl’innocenti, A.; Perrotta, M.; Barsacchi, G.; Andreazzoli, M. Characterization of the rx1-dependent transcriptome during early retinal development. Dev. Dyn. 2014, 243, 1352–1361. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.M.A.; Cepko, C.L. Expression of chx10 and chx10-1 in the developing chicken retina. Mech. Dev. 2000, 90, 293–297. [Google Scholar] [CrossRef]
- Sethi, A.S.; Lees, D.M.; Douthwaite, J.A.; Dawnay, A.B.; Corder, R. Homocysteine-induced endothelin-1 release is dependent on hyperglycaemia and reactive oxygen species production in bovine aortic endothelial cells. J. Vasc. Res. 2006, 43, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Baack, M.L.; Wang, C.; Hu, S.; Segar, J.L.; Norris, A.W. Hyperglycemia induces embryopathy, even in the absence of systemic maternal diabetes: An in vivo test of the fuel mediated teratogenesis hypothesis. Reprod. Toxicol. 2014, 46, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, A.K.; Glasgow, B.J.; Pardridge, W.M. Glut1 glucose transporter expression in the diabetic and nondiabetic human eye. Investig. Ophthalmol. Vis. Sci. 1994, 35, 2887–2894. [Google Scholar]
- Moley, K.H.; Chi, M.M.; Mueckler, M.M. Maternal hyperglycemia alters glucose transport and utilization in mouse preimplantation embryos. Am. J. Physiol. 1998, 275, E38–E47. [Google Scholar] [PubMed]
- Park, M.-S. Molecular dynamics simulations of the human glucose transporter glut1. PLoS ONE 2015, 10, e0125361. [Google Scholar] [CrossRef] [PubMed]
- Marques, T.; Patente, T.A.; Monteiro, M.B.; Cavaleiro, A.M.; Queiroz, M.S.; Nery, M.; de Azevedo, M.J.; Canani, L.H.; Parisi, M.C.; Moura-Neto, A.; et al. Association of single nucleotide polymorphisms in the gene encoding glut1 and diabetic nephropathy in brazilian patients with type 1 diabetes mellitus. Clin. Chim. Acta 2015, 444, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, D.; Lee, E.K.; Kim, S.; Choi, C.S.; Jun, H.S. Sodium meta-arsenite prevents the development of autoimmune diabetes in nod mice. Toxicol. Appl. Pharmacol. 2015, 284, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, J.; Zhang, X.-Y. 2-NBDG as a marker for detecting glucose uptake in reactive astrocytes exposed to oxygen-glucose deprivation in vitro. J. Mol. Neurosci. 2015, 55, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Gunnink, S.M.; Kerk, S.A.; Kuiper, B.D.; Alabi, O.D.; Kuipers, D.P.; Praamsma, R.C.; Wrobel, K.E.; Louters, L.L. Alkaline ph activates the transport activity of GLUT1 in L929 fibroblast cells. Biochimie 2014, 99, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Loeken, M.R. Advances in understanding the molecular causes of diabetes-induced birth defects. J. Soc. Gynecol. Investig. 2006, 13, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Dieterlen-Lievre, F.; Beaupain, D. Immunocytological study of endocrine pancreas ontogeny in the chick embryo: Normal development and pancreatic potentialities in the early splanchnopleura. In The Evolution of Pancreatic Islets; Pergamon Press: Oxford, UK, 1976; pp. 37–50. [Google Scholar]
- Naftolin, F.; Diamond, M.P.; Pinter, E.; Reece, E.A.; Sanyal, M.K. A hypothesis concerning the general basis of organogenetic congenital anomalies. Am. J. Obstet. Gynecol. 1987, 157, 1–4. [Google Scholar] [CrossRef]
- Newsholme, P.; de Bittencourt, P.I.H., Jr.; O’Hagan, C.; de Vito, G.; Murphy, C.; Krause, M.S. Exercise and possible molecular mechanisms of protection from vascular disease and diabetes: The central role of ros and nitric oxide. Clin. Sci. 2009, 118, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Araki, E. Impact of mitochondrial ros production in the pathogenesis of diabetes mellitus and its complications. Antioxid. Redox Signal. 2007, 9, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.M.; Zhao, S.Z.; Zhang, Z.L.; Chen, Y.; Cheng, X.; Chuai, M.; Liu, G.S.; Lee, K.K.H.; Yang, X. High glucose level induces cardiovascular dysplasia during early embryo development. Exp. Clin. Endocrinol. Diabetes 2013, 121, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Bitar, M.S.; Al-Mulla, F. Ros constitute a convergence nexus in the development of igf1 resistance and impaired wound healing in a rat model of type 2 diabetes. Dis. Model. Mech. 2012, 5, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Tang, J.; Kern, T.S. Abnormalities of retinal metabolism in diabetes and experimental galactosemia vii. Effect of long-term administration of antioxidants on the development of retinopathy. Diabetes 2001, 50, 1938–1942. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Walczysko, P.; Kucerova, R.; Rajnicek, A.M.; McCaig, C.D.; Zhao, M.; Collinson, J.M. Chronic wound state exacerbated by oxidative stress in pax6(+/−) aniridia-related keratopathy. J. Pathol. 2008, 215, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Pataki, T.; Bak, I.; Kovacs, P.; Bagchi, D.; Das, D.K.; Tosaki, A. Grape seed proanthocyanidins improved cardiac recovery during reperfusion after ischemia in isolated rat hearts. Am. J. Clin. Nutr. 2002, 75, 894–899. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, R.-R.; Zhang, S.-J.; Li, Y.-F.; Tsoi, B.; Huang, W.-S.; Yao, N.; Hong, M.; Zhai, Y.-J.; Mao, Z.-F.; Tang, L.-P.; et al. Proanthocyanidins Prevent High Glucose-Induced Eye Malformation by Restoring Pax6 Expression in Chick Embryo. Nutrients 2015, 7, 6567-6581. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7085299
Tan R-R, Zhang S-J, Li Y-F, Tsoi B, Huang W-S, Yao N, Hong M, Zhai Y-J, Mao Z-F, Tang L-P, et al. Proanthocyanidins Prevent High Glucose-Induced Eye Malformation by Restoring Pax6 Expression in Chick Embryo. Nutrients. 2015; 7(8):6567-6581. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7085299
Chicago/Turabian StyleTan, Rui-Rong, Shi-Jie Zhang, Yi-Fang Li, Bun Tsoi, Wen-Shan Huang, Nan Yao, Mo Hong, Yu-Jia Zhai, Zhong-Fu Mao, Lu-Ping Tang, and et al. 2015. "Proanthocyanidins Prevent High Glucose-Induced Eye Malformation by Restoring Pax6 Expression in Chick Embryo" Nutrients 7, no. 8: 6567-6581. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7085299