Mass Spectrometry-Based Metabolomic and Lipidomic Analyses of the Effects of Dietary Platycodon grandiflorum on Liver and Serum of Obese Mice under a High-Fat Diet



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of PG Extract
2.3. Experimental Design
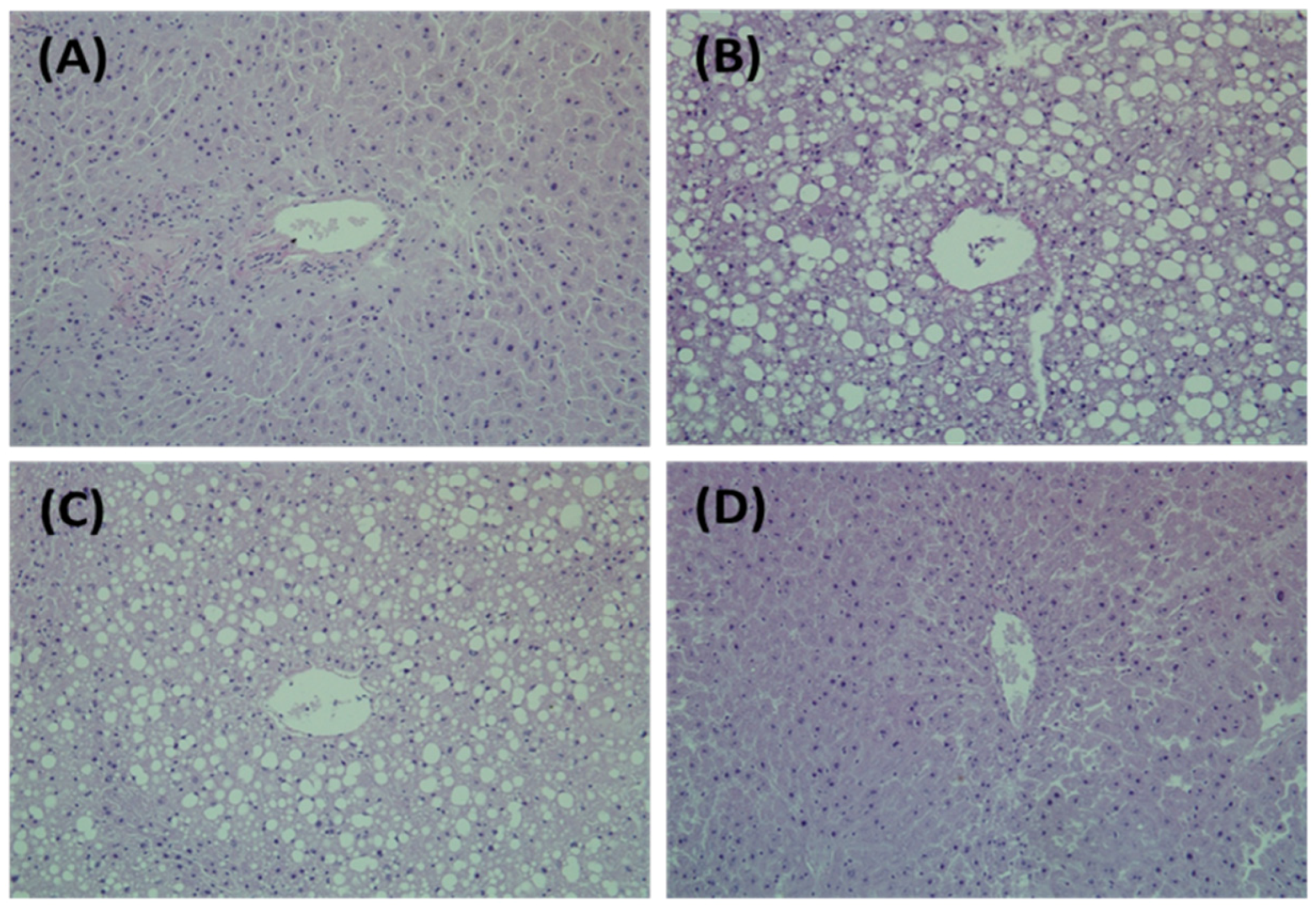
2.4. Hepatic Morphology
2.5. Hepatic Biochemical Parameters in Serum
2.6. Serum Extraction
2.7. Liver Tissue Extraction
2.8. GC-TOF-MS Analysis
2.9. UPLC-Q-TOF-MS Analysis
2.10. Lipid Profiling by Direct Infusion MS
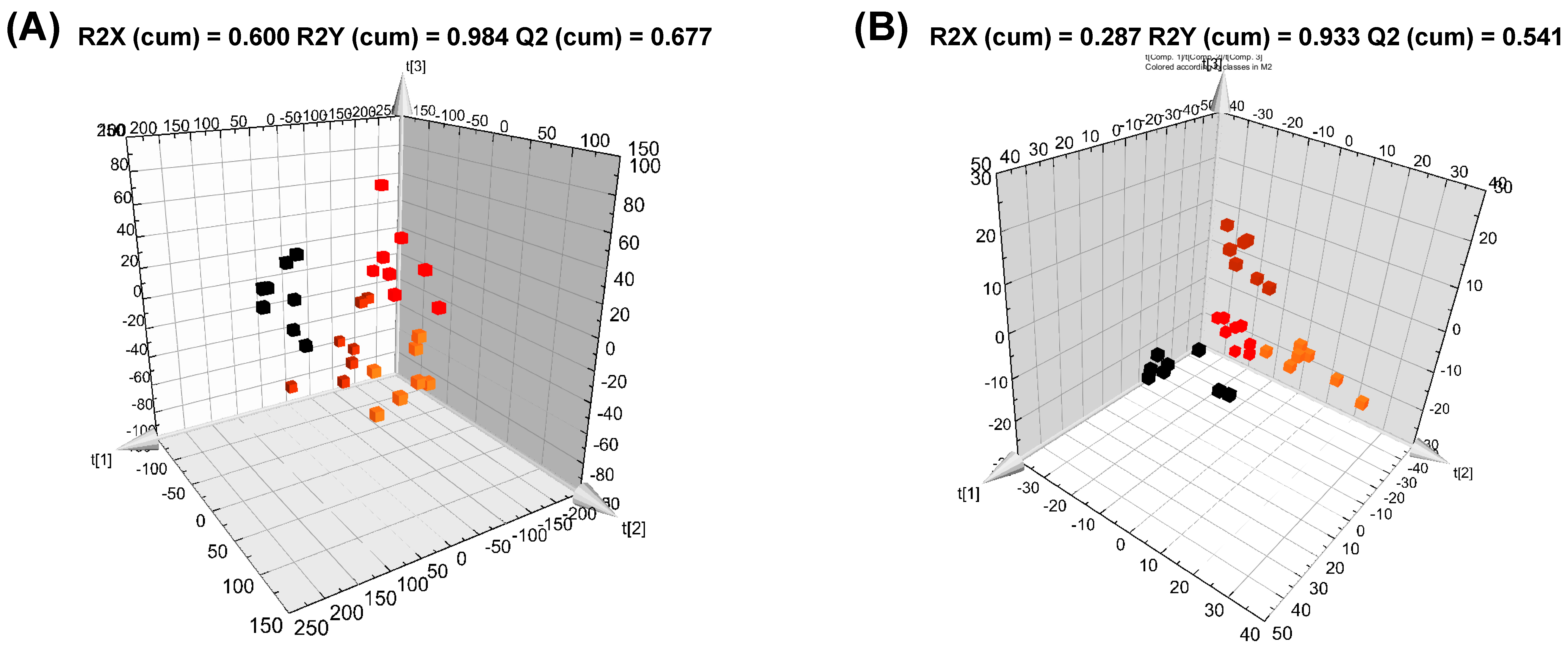
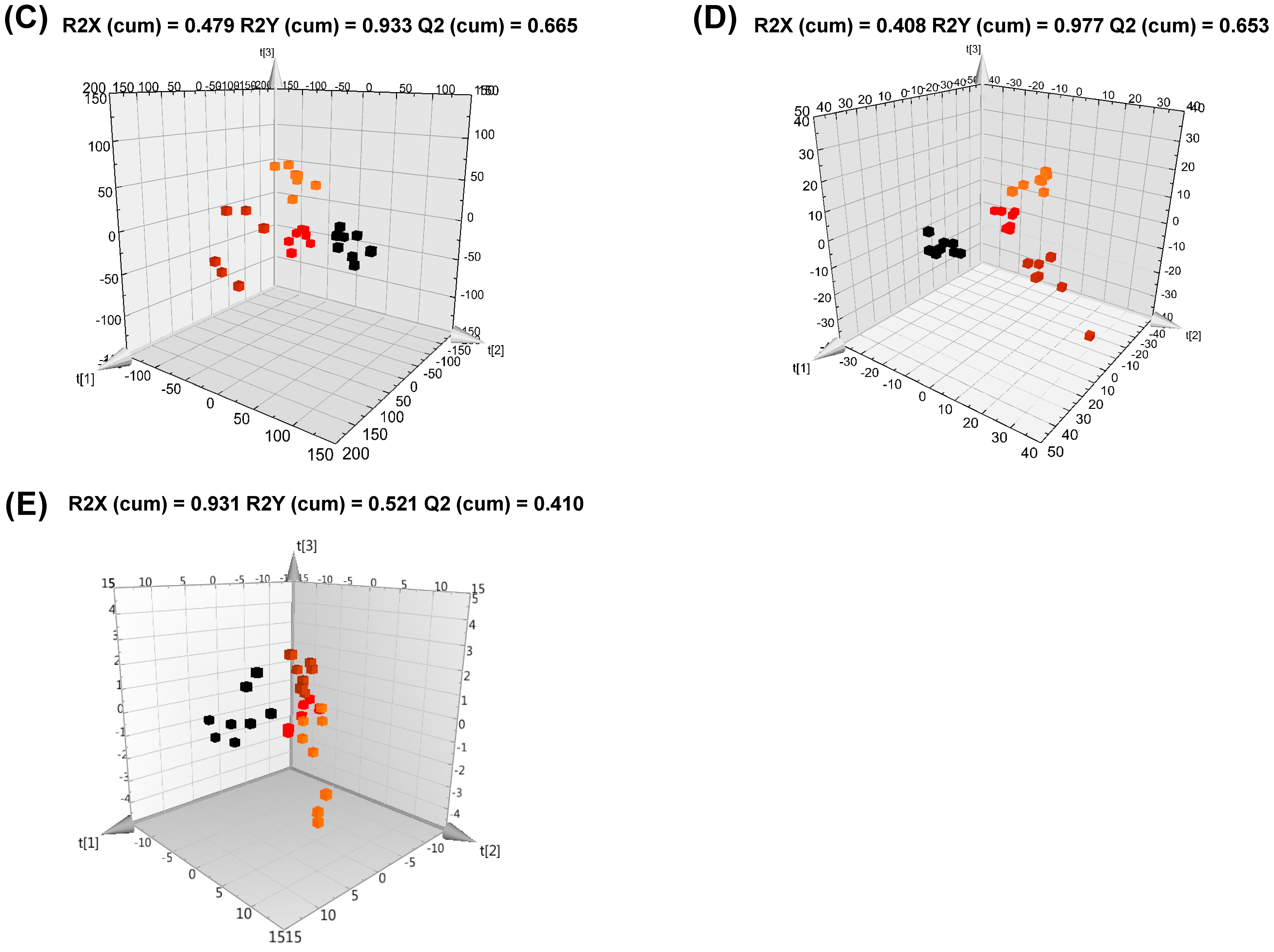
2.11. Data Processing and Multivariate Analysis
3. Results
3.1. Effects of PG on HFD-Induced Obesity in C57BL/6J Mice
3.2. Serum Metabolite Changes in HFD and PG Extract-Fed Mice
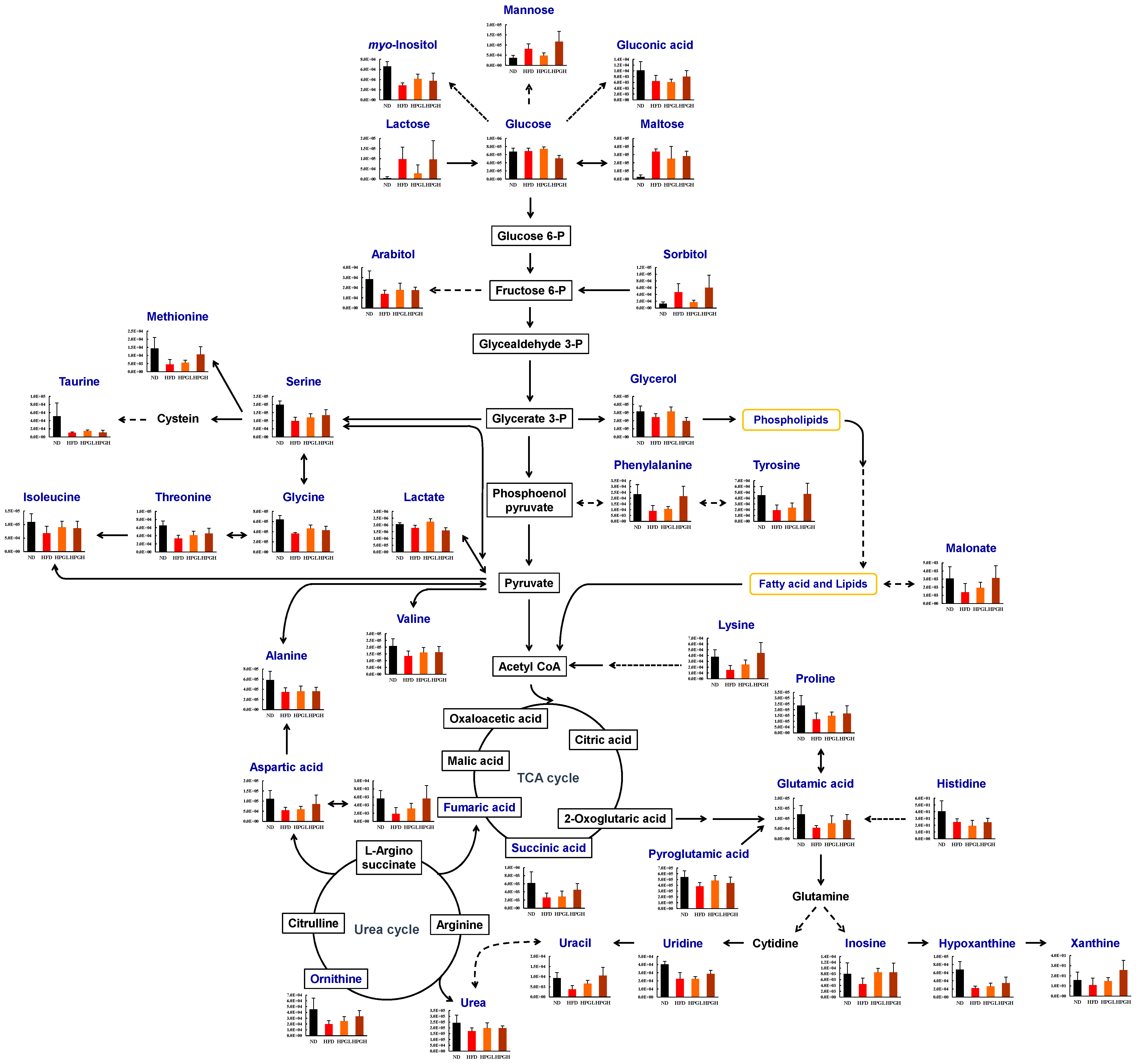
3.3. Hepatic Metabolite Changes in HFD and PG Extract-Fed Mice
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nikaido, T.; Koike, K.; Mitsunaga, K.; Saeki, T. Two new triterpenoid saponins from Platycodon grandiflorum. Chem. Pharm. Bull. 1999, 47, 903–904. [Google Scholar] [CrossRef] [PubMed]
- Jeonga, C.-H.; Choia, G.N.; Kima, J.H.; Kwaka, J.H.; Kimb, D.O.; Kimc, Y.J.; Heoa, H.J. Antioxidant activities from the aerial parts of Platycodon grandiflorum. Food Chem. 2010, 118, 278–282. [Google Scholar] [CrossRef]
- Ma, G.; Guo, W.; Zhao, L.; Zheng, Q.; Sun, Z.; Wei, J.; Yang, J.; Xu, X. Two new triterpenoid saponins from the root of Platycodon grandiflorum. Chem. Pharm. Bull. 2013, 61, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; You, H.J.; Park, S.J.; Kim, Y.S.; Chung, Y.C.; Jeong, T.C.; Jeong, H.G. Hepatoprotective effects of Platycodon grandiflorum on acetaminophen-induced liver damage in mice. Cancer Lett. 2001, 174, 73–81. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hwang, W.I.; Lim, S.T. Antioxidant and anticancer activities of organic extracts from Platycodon grandiflorum A. De candolle roots. J. Ethnopharmacol. 2004, 93, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.S.; Noh, E.J.; Zhao, H.L.; Jung, S.H.; Kang, S.S.; Kim, Y.S. Inhibition of inducible nitric oxide synthase and cyclooxygenase II by Platycodon grandiflorum saponins via suppression of nuclear factor-kappab activation in raw 264.7 cells. Life Sci. 2005, 76, 2315–2328. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Seo, E.K.; Lee, Y.C.; Lee, T.K.; Cho, Y.W.; Ezaki, O.; Kim, C.H. Effect of dietary Platycodon grandiflorum on the improvement of insulin resistance in obese zucker rats. J. Nutr. Biochem. 2000, 11, 420–424. [Google Scholar] [CrossRef]
- Han, L.K.; Xu, B.J.; Kimura, Y.; Zheng, Y.; Okuda, H. Platycodi radix affects lipid metabolism in mice with high fat diet-induced obesity. J. Nutr. 2000, 130, 2760–2764. [Google Scholar] [PubMed]
- Zhao, H.L.; Harding, S.V.; Marinangeli, C.P.; Kim, Y.S.; Jones, P.J. Hypocholesterolemic and anti-obesity effects of saponins from Platycodon grandiflorum in hamsters fed atherogenic diets. J. Food Sci. 2008, 73, H195–H200. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Obesity and Overweight Fact Sheet N°311; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Akiyama, T.; Tachibana, I.; Shirohara, H.; Watanabe, N.; Otsuki, M. High-fat hypercaloric diet induces obesity, glucose intolerance and hyperlipidemia in normal adult male wistar rat. Diabetes Res. Clin. Pract. 1996, 31, 27–35. [Google Scholar] [CrossRef]
- Lin, S.; Thomas, T.C.; Storlien, L.H.; Huang, X.F. Development of high fat diet-induced obesity and leptin resistance in C57BL/6J mice. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Ellis, D.I. Metabolomics: Current analytical platforms and methodologies. TrAC Trends Anal. Chem. 2005, 24, 285–294. [Google Scholar]
- Zhang, A.; Sun, H.; Wang, P.; Han, Y.; Wang, X. Recent and potential developments of biofluid analyses in metabolomics. J. Proteom. 2012, 75, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Abu Bakar, M.H.; Sarmidi, M.R.; Cheng, K.K.; Ali Khan, A.; Suan, C.L.; Zaman Huri, H.; Yaakob, H. Metabolomics—The complementary field in systems biology: A review on obesity and type 2 diabetes. Mol. BioSyst. 2015, 11, 1742–1774. [Google Scholar] [CrossRef] [PubMed]
- Shearer, J.; Duggan, G.; Weljie, A.; Hittel, D.S.; Wasserman, D.H.; Vogel, H.J. Metabolomic profiling of dietary-induced insulin resistance in the high fat-fed C57BL/6J mouse. Diabetes Obes. Metab. 2008, 10, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Serkova, N.J.; Jackman, M.; Brown, J.L.; Liu, T.; Hirose, R.; Roberts, J.P.; Maher, J.J.; Niemann, C.U. Metabolic profiling of livers and blood from obese zucker rats. J. Hepatol. 2006, 44, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, J.H.; Noh, S.; Hur, H.J.; Sung, M.J.; Hwang, J.T.; Park, J.H.; Yang, H.J.; Kim, M.S.; Kwon, D.Y.; et al. Metabolomic analysis of livers and serum from high-fat diet induced obese mice. J. Proteome Res. 2011, 10, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Lee, M.Y.; Park, H.M.; Park, Y.K.; Shon, J.C.; Liu, K.H.; Lee, C.H. Urine and serum metabolite profiling of rats fed a high-fat diet and the anti-obesity effects of caffeine consumption. Molecules 2015, 20, 3107–3128. [Google Scholar] [CrossRef] [PubMed]
- Gooda Sahib Jambocus, N.; Saari, N.; Ismail, A.; Khatib, A.; Mahomoodally, M.F.; Abdul Hamid, A. An investigation into the antiobesity effects of Morinda citrifolia L. Leaf extract in high fat diet induced obese rats using a 1H NMR metabolomics approach. J. Diabetes Res. 2016, 2016, 2391592. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Choi, J.Y.; Ryu, R.; Lee, J.; Cho, S.J.; Kwon, E.Y.; Lee, M.K.; Liu, K.H.; Rina, Y.; Sung, M.K.; et al. Platycodon grandiflorus root extract attenuates body fat mass, hepatic steatosis and insulin resistance through the interplay between the liver and adipose tissue. Nutrients 2016, 8, 532. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Yuan, H.D.; Chung, I.K.; Chung, S.H. Compound K, intestinal metabolite of ginsenoside, attenuates hepatic lipid accumulation via AMPK activation in human hepatoma cells. J. Agric. Food Chem. 2009, 57, 1532–1537. [Google Scholar] [CrossRef] [PubMed]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Masson, P.; Alves, A.C.; Ebbels, T.M.; Nicholson, J.K.; Want, E.J. Optimization and evaluation of metabolite extraction protocols for untargeted metabolic profiling of liver samples by UPLC-MS. Anal. Chem. 2010, 82, 7779–7786. [Google Scholar] [CrossRef] [PubMed]
- Shon, J.C.; Shin, H.S.; Seo, Y.K.; Yoon, Y.R.; Shin, H.; Liu, K.H. Direct infusion MS-based lipid profiling reveals the pharmacological effects of compound K-reinforced ginsenosides in high-fat diet induced obese mice. J. Agric. Food Chem. 2015, 63, 2919–2929. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Liu, K.H.; Lee, D.Y.; DeFelice, B.; Meissen, J.K.; Fiehn, O. Lipidblast in silico tandem mass spectrometry database for lipid identification. Nat. Methods 2013, 10, 755–758. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.S.; Chen, W.C.; Kuo, T.C.; Ho, C.T.; Kuo, C.H.; Tseng, Y.J.; Lu, K.H.; Lin, S.H.; Panyod, S.; Sheen, L.Y. Mass-spectrometry-based serum metabolomics of a C57BL/6J mouse model of high-fat-diet-induced non-alcoholic fatty liver disease development. J. Agric. Food Chem. 2015, 63, 7873–7884. [Google Scholar] [CrossRef] [PubMed]
- Me, B.; Jt, B. The Textbook of Hepatology: From Basic Science to Clinical Practice, 3rd ed.; Wiley-Blackwell: Oxford, UK, 2007; pp. 142–149. [Google Scholar]
- Newsholme, P.; Procopio, J.; Lima, M.M.; Pithon-Curi, T.C.; Curi, R. Glutamine and glutamate—Their central role in cell metabolism and function. Cell Biochem. Funct. 2003, 21, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Satapati, S.; Sunny, N.E.; Kucejova, B.; Fu, X.; He, T.T.; Mendez-Lucas, A.; Shelton, J.M.; Perales, J.C.; Browning, J.D.; Burgess, S.C. Elevated tca cycle function in the pathology of diet-induced hepatic insulin resistance and fatty liver. J. Lipid Res. 2012, 53, 1080–1092. [Google Scholar] [CrossRef] [PubMed]
- Shambaugh, G.E., III. Urea biosynthesis I. The urea cycle and relationships to the citric acid cycle. Am. J. Clin. Nutr. 1977, 30, 2083–2087. [Google Scholar] [PubMed]
- Liao, C.C.; Lin, Y.L.; Kuo, C.F. Effect of high-fat diet on hepatic proteomics of hamsters. J. Agric. Food Chem. 2015, 63, 1869–1881. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Brunengraber, H. Glutamate, a window on liver intermediary metabolism. J. Nutr. 2000, 130, 991S–994S. [Google Scholar] [PubMed]
- Brosnan, M.E.; Brosnan, J.T. Hepatic glutamate metabolism: A tale of 2 hepatocytes. Am. J. Clin. Nutr. 2009, 90, 857S–861S. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T. Glutamate, at the interface between amino acid and carbohydrate metabolism. J. Nutr. 2000, 130, 988S–990S. [Google Scholar] [PubMed]
- Setzer, B.; Lebrecht, D.; Walker, U.A. Pyrimidine nucleoside depletion sensitizes to the mitochondrial hepatotoxicity of the reverse transcriptase inhibitor stavudine. Am. J. Pathol. 2008, 172, 681–690. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Xu, W.; Li, H.; Lei, H.; Zhang, L.; Hao, F.; Duan, Y.; Yan, X.; Zhao, Y.; Wu, J.; et al. High-fat diet induces dynamic metabolic alterations in multiple biological matrices of rats. J. Proteome Res. 2013, 12, 3755–3768. [Google Scholar] [CrossRef] [PubMed]
- Oosterveer, M.H.; van Dijk, T.H.; Tietge, U.J.; Boer, T.; Havinga, R.; Stellaard, F.; Groen, A.K.; Kuipers, F.; Reijngoud, D.J. High fat feeding induces hepatic fatty acid elongation in mice. PLoS ONE 2009, 4, e6066. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.R.; Kim, Y.H.; Gang, G.T.; Yang, K.J.; Kim, S.K.; Ryu, S.Y.; Kim, Y.S.; Lee, C.H.; Lee, H.S. Preventative effects of Platycodon grandiflorum treatment on hepatic steatosis in high fat diet-fed C57BL/6 mice. Biol. Pharm. Bull. 2010, 33, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.; Choi, M.S.; Jung, S.; Jung, Y.; Choi, J.Y.; Ryu do, H.; Hwang, G.S. Lipidomic profiling of liver tissue from obesity-prone and obesity-resistant mice fed a high fat diet. Sci. Rep. 2015, 5, 16984. [Google Scholar] [CrossRef] [PubMed]
- Gorden, D.L.; Ivanova, P.T.; Myers, D.S.; McIntyre, J.O.; VanSaun, M.N.; Wright, J.K.; Matrisian, L.M.; Brown, H.A. Increased diacylglycerols characterize hepatic lipid changes in progression of human nonalcoholic fatty liver disease; comparison to a murine model. PLoS ONE 2011, 6, e22775. [Google Scholar] [CrossRef] [PubMed]
- Eisinger, K.; Krautbauer, S.; Hebel, T.; Schmitz, G.; Aslanidis, C.; Liebisch, G.; Buechler, C. Lipidomic analysis of the liver from high-fat diet induced obese mice identifies changes in multiple lipid classes. Exp. Mol. Pathol. 2014, 97, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyn, A.; Dobrzyn, P.; Miyazaki, M.; Sampath, H.; Chu, K.; Ntambi, J.M. Stearoyl-coa desaturase 1 deficiency increases ctp: Choline cytidylyltransferase translocation into the membrane and enhances phosphatidylcholine synthesis in liver. J. Biol. Chem. 2005, 280, 23356–23362. [Google Scholar] [CrossRef] [PubMed]
- Van der Veen, J.N.; Lingrell, S.; da Silva, R.P.; Jacobs, R.L.; Vance, D.E. The concentration of phosphatidylethanolamine in mitochondria can modulate atp production and glucose metabolism in mice. Diabetes 2014, 63, 2620–2630. [Google Scholar] [CrossRef] [PubMed]
- DeLong, C.J.; Shen, Y.J.; Thomas, M.J.; Cui, Z. Molecular distinction of phosphatidylcholine synthesis between the cdp-choline pathway and phosphatidylethanolamine methylation pathway. J. Biol. Chem. 1999, 274, 29683–29688. [Google Scholar] [CrossRef] [PubMed]
- Khanal, T.; Choi, J.H.; Hwang, Y.P.; Chung, Y.C.; Jeong, H.G. Protective effects of saponins from the root of Platycodon grandiflorum against fatty liver in chronic ethanol feeding via the activation of AMP-dependent protein kinase. Food Chem. Toxicol. 2009, 47, 2749–2754. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.E.; Hur, H.J.; Hwang, J.T.; Sung, M.J.; Yang, H.J.; Kim, H.J.; Park, J.H.; Kwon, D.Y.; Kim, M.S. Long-term consumption of platycodi radix ameliorates obesity and insulin resistance via the activation of AMPK pathways. Evid.-Based Complement. Altern. Med. 2012, 2012, 759143. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C.; Mato, J.M. S-adenosylmethionine in liver health, injury, and cancer. Physiol. Rev. 2012, 92, 1515–1542. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, R.L.; van der Veen, J.N.; Vance, D.E. Finding the balance: The role of S-adenosylmethionine and phosphatidylcholine metabolism in development of nonalcoholic fatty liver disease. Hepatology 2013, 58, 1207–1209. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.N.; Risis, S.; Yang, C.; Meikle, P.J.; Staples, M.; Febbraio, M.A.; Bruce, C.R. Plasma lysophosphatidylcholine levels are reduced in obesity and type 2 diabetes. PLoS ONE 2012, 7, e41456. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, R.; Barrera, C.; Espinosa, A.; Llanos, P.; Orellana, P.; Videla, L.A. Reduction in the desaturation capacity of the liver in mice subjected to high fat diet: Relation to lcpufa depletion in liver and extrahepatic tissues. Prostaglandins Leukot. Essent. Fat. Acids 2015, 98, 7–14. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weight (g) | ND | HFD | HPGL | HPGH |
|---|---|---|---|---|
| Body | 31 ± 4 a | 49 ± 1 b | 43 ± 3 b | 32 ± 3 a |
| Liver | 1 ± 0.2 a | 2 ± 0.9 b | 2 ± 0.1 b | 1 ± 0.1 a |
| Subcutaneous fat | 1 ± 0.4 a | 5 ± 2 b | 5 ± 0.6 b | 2 ± 1 a |
| Visceral fat | 1 ± 0.4 a | 3 ± 0.7 c | 4 ± 0.4 c | 2 ± 1 b |
| Parameters | ND | HFD | HPGL | HPGH |
|---|---|---|---|---|
| AST (U/L) | 159 ± 90 a | 254 ± 96 b | 199 ± 70 a,b | 175 ± 50 a,b |
| ALT (U/L) | 39 ± 19 a | 201 ± 133 b | 187 ± 92 b | 35 ± 20 a |
| Total cholesterol (mg/dL) | 141 ± 18 a | 264 ± 62 b | 254 ± 25 b | 195 ± 51 c |
| No. | Metabolite | Fold Change 1 | ||
|---|---|---|---|---|
| HFD/ND | HPGL/HFD | HPGH/HFD | ||
| 1 | Ornithine | 1.38 # | 0.76 # | 0.72 # |
| 2 | Tryptophan | 1.26 # | 0.85 # | 0.78 # |
| 3 | Saccharide * | 1.67 # | 0.92 | 0.79 # |
| 4 | LysoPC 20:4 | 1.65 # | 0.98 | 0.66 # |
| 5 | PC 34:2 (16:0/18:2) | 0.55 # | 0.88 | 1.39 # |
| 6 | PC 36:4 (16:0/20:4) | 1.50 # | 1.11 | 0.71 # |
| 7 | PC 38:4 (18:0/20:4) | 2.28 # | 1.01 | 0.60 # |
| 8 | PC 38:3 (18:0/20:3) | 2.07 # | 1.00 | 0.65 # |
| 9 | PC 38:3 | 1.29 # | 1.07 | 0.77 # |
| 10 | TG 52:3 | 0.53 # | 0.66 | 2.06 # |
| No. | Metabolite | Fold Change 1 | ||
|---|---|---|---|---|
| HFD/ND | HPGL/HFD | HPGH/HFD | ||
| 1 | Glycine | 0.57 # | 1.27 # | 1.18 # |
| 2 | Serine | 0.50 # | 1.21 | 1.34 # |
| 3 | Threonine | 0.51 # | 1.25 | 1.37 # |
| 4 | Methionine a | 0.31 # | 1.25 | 2.38 # |
| 5 | Glutamic acid | 0.45 # | 1.42 | 1.71 # |
| 6 | Phenylalanine a | 0.39 # | 1.19 | 2.40 # |
| 7 | Ornithine | 0.44 # | 1.26 | 1.67 # |
| 8 | Lysine a | 0.39 # | 1.63 # | 2.98 # |
| 9 | Tyrosine a | 0.42 # | 1.24 | 2.47 # |
| 10 | Urea | 0.71 # | 1.15 | 1.15 # |
| 11 | Succinic acid | 0.42 # | 1.11 | 1.75 # |
| 12 | Fumaric acid a | 0.34 # | 1.67 | 2.96 # |
| 13 | Linolelidic acid methyl ester a | 1.53 # | 0.85 # | 0.54 # |
| 14 | Oleic acid methyl ester a | 2.45 # | 1.56 # | 0.54 # |
| 15 | Oleamide a | 1.21 # | 0.90 | 0.72 # |
| 16 | Cholesterol | 0.24 # | 1.97 # | 2.82 # |
| 17 | Maltose | 12.33 # | 0.75 | 0.83 # |
| 18 | Uracil a | 0.40 # | 1.75 # | 2.81 # |
| 19 | Hypoxanthine | 0.33 # | 1.18 | 1.53 # |
| 20 | LysoPC 22:6 | 4.12 # | 0.51 # | 0.58 # |
| 21 | LysoPC 20:4 | 5.88 # | 0.50 # | 0.48 # |
| 22 | LysoPC 22:6 | 1.72 # | 0.86 # | 0.78 # |
| 23 | LysoPE 18:2 a | 0.21 # | 1.11 | 2.25 # |
| 24 | LysoPC 20:4 | 2.50 # | 0.79 # | 0.67 # |
| 25 | LysoPC 18:2 a | 0.50 # | 1.01 | 1.53 # |
| 26 | LysoPC 18:2 | 0.45 # | 1.25 | 2.32 # |
| 27a | LysoPC 16:0 a | 0.64 # | 1.70 # | 1.39 # |
| 27b | LysoPC 20:3 | 3.44 # | 0.79 | 0.64 # |
| 28 | LysoPE 18:0 | 0.23 # | 5.76 # | 3.06 # |
| 29 | PE 36:3 | 0.51 # | 0.89 | 1.78 # |
| 30 | PE 36:2 | 0.42 # | 0.90 | 2.22 # |
| 31 | PC 38:9 | 1.79 # | 0.92 | 0.60 # |
| 32 | TG 50:2 | 2.52 # | 0.78 | 0.45 # |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.M.; Park, K.-T.; Park, E.C.; Kim, S.I.; Choi, M.S.; Liu, K.-H.; Lee, C.H. Mass Spectrometry-Based Metabolomic and Lipidomic Analyses of the Effects of Dietary Platycodon grandiflorum on Liver and Serum of Obese Mice under a High-Fat Diet. Nutrients 2017, 9, 71. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9010071
Park HM, Park K-T, Park EC, Kim SI, Choi MS, Liu K-H, Lee CH. Mass Spectrometry-Based Metabolomic and Lipidomic Analyses of the Effects of Dietary Platycodon grandiflorum on Liver and Serum of Obese Mice under a High-Fat Diet. Nutrients. 2017; 9(1):71. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9010071
Chicago/Turabian StylePark, Hye Min, Kab-Tae Park, Edmond Changkyun Park, Seung II Kim, Myung Sook Choi, Kwang-Hyeon Liu, and Choong Hwan Lee. 2017. "Mass Spectrometry-Based Metabolomic and Lipidomic Analyses of the Effects of Dietary Platycodon grandiflorum on Liver and Serum of Obese Mice under a High-Fat Diet" Nutrients 9, no. 1: 71. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9010071