Leucine-Enriched Essential Amino Acids Augment Muscle Glycogen Content in Rats Seven Days after Eccentric Contraction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Leucine-Enriched Essential Amino Acids
2.4. Measurement of High Energy Phosphate Compounds and Muscle Glycogen
2.5. Statistical Analysis
3. Results
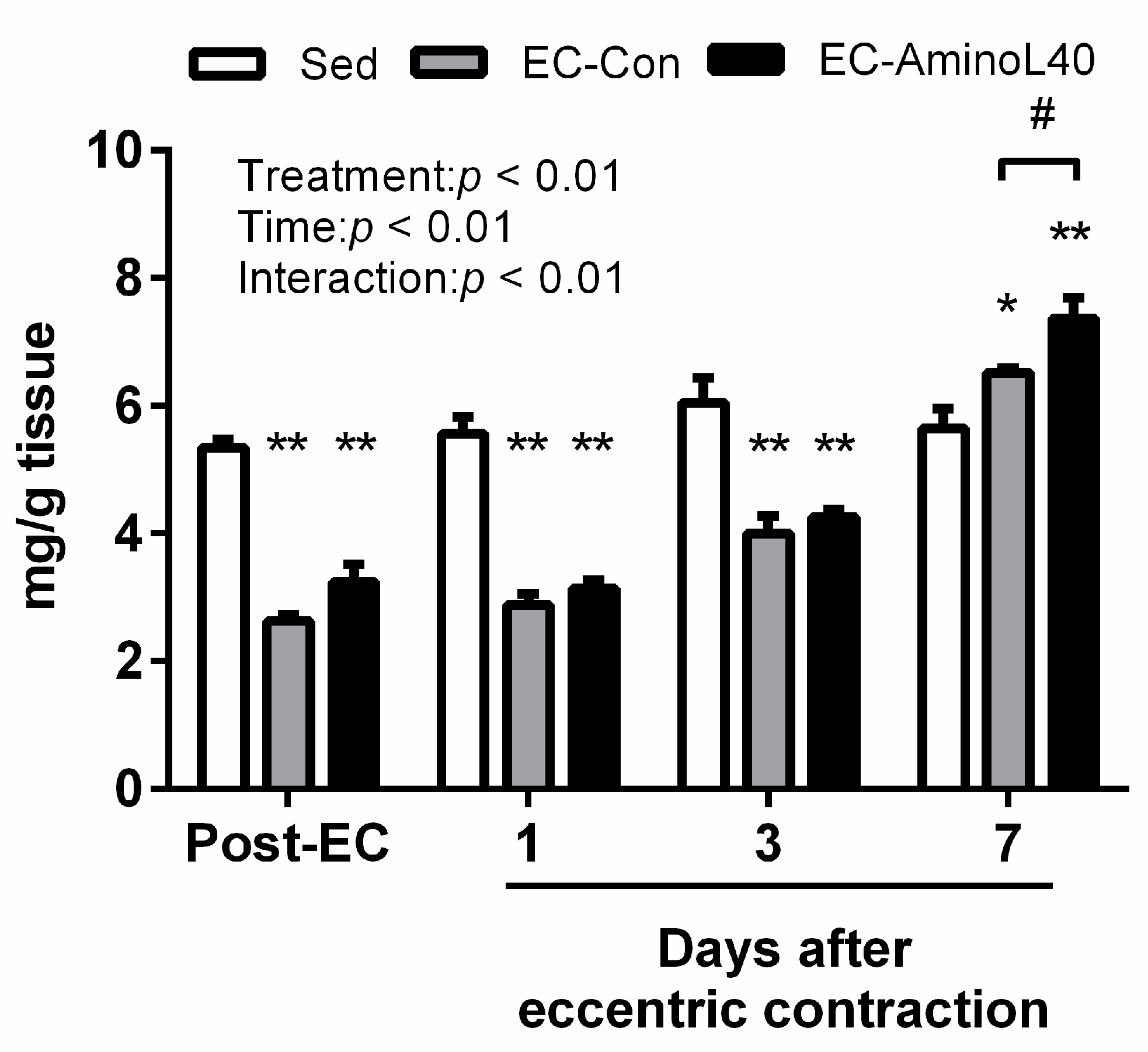
3.1. Muscle Glycogen Content Following Eccentric Contractions
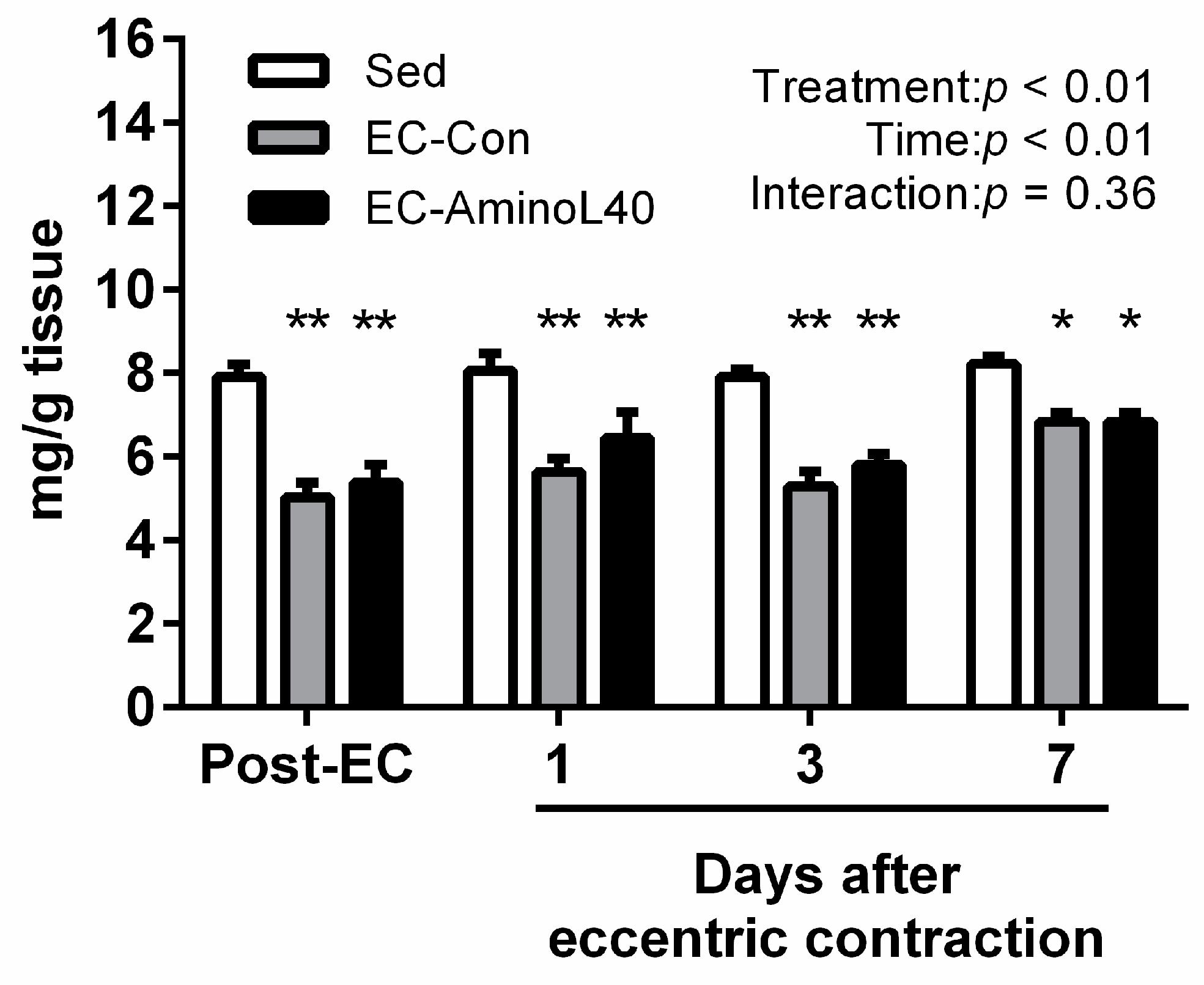
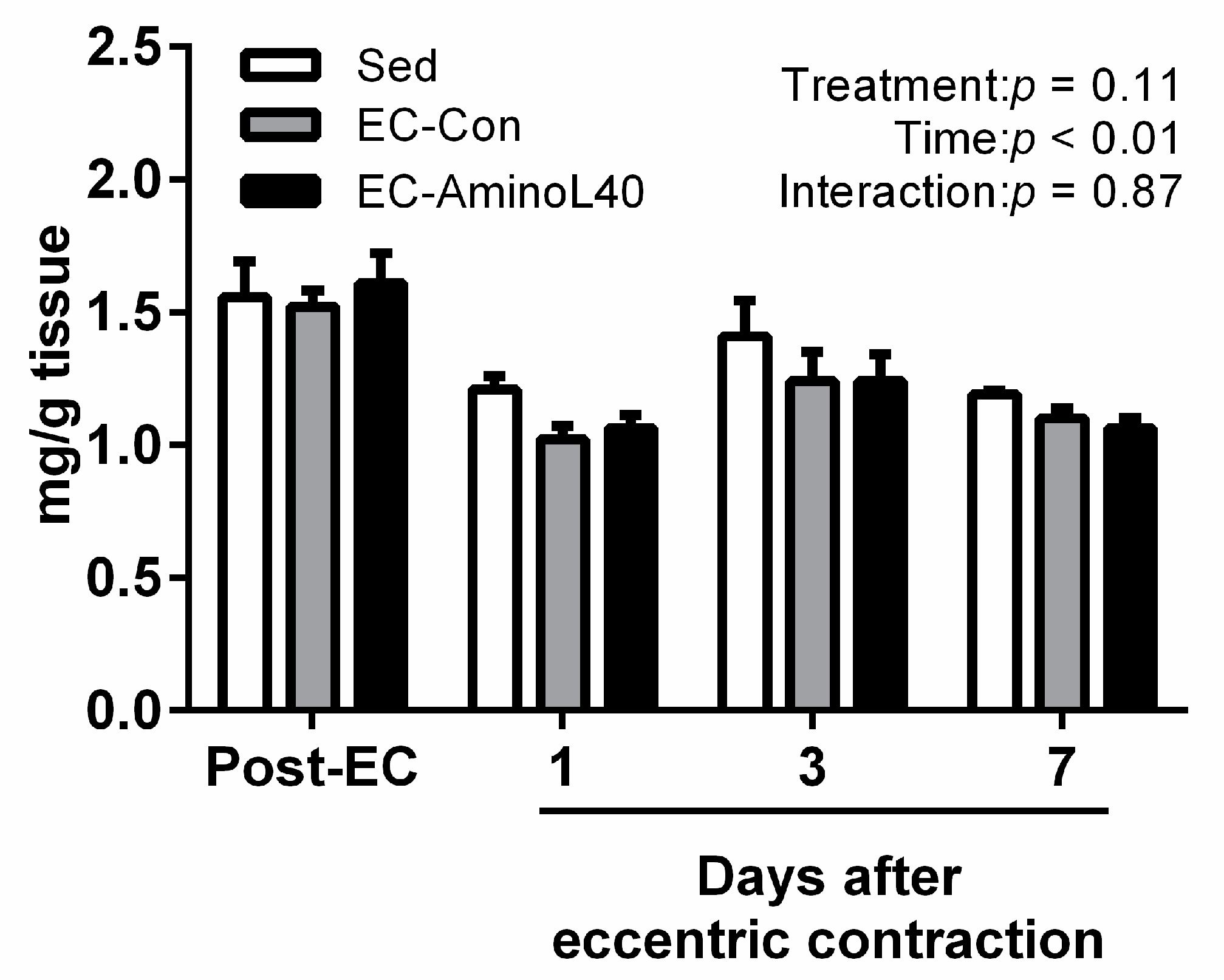
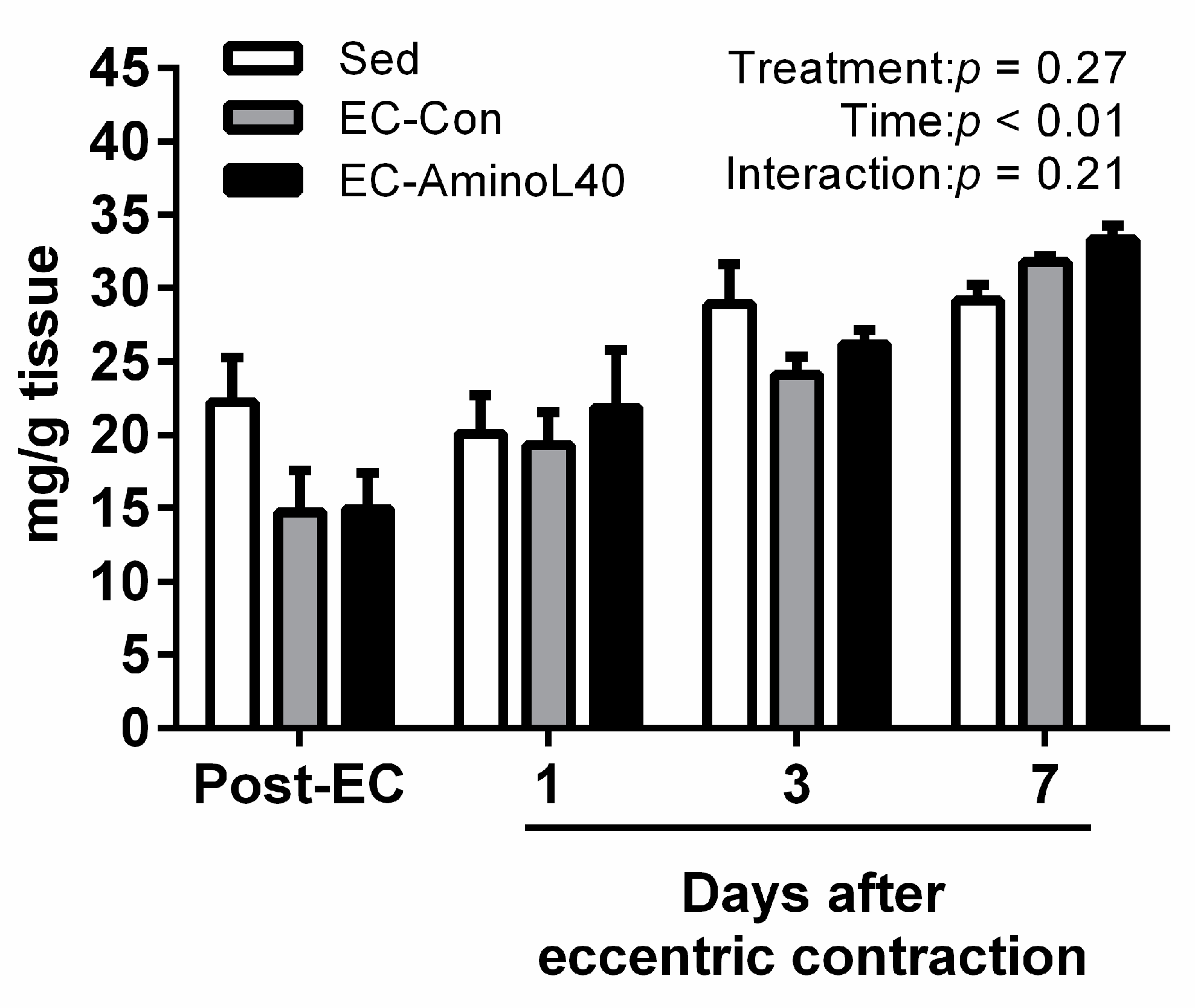
3.2. High Energy Phosphate Compounds Content following Eccentric Contraction
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cleak, M.J.; Eston, R.G. Delayed onset muscle soreness: Mechanisms and management. J. Sports Sci. 1992, 10, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Raastad, T.; Owe, S.G.; Paulsen, G.; Enns, D.; Overgaard, K.; Crameri, R.; Kiil, S.; Belcastro, A.; Bergersen, L.; Hallen, J. Changes in calpain activity, muscle structure, and function after eccentric exercise. Med. Sci. Sports Exerc. 2010, 42, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Costill, D.L.; Pascoe, D.D.; Fink, W.J.; Robergs, R.A.; Barr, S.I.; Pearson, D. Impaired muscle glycogen resynthesis after eccentric exercise. J. Appl. Physiol. 1990, 69, 46–50. [Google Scholar] [PubMed]
- Widrick, J.J.; Costill, D.L.; McConell, G.K.; Anderson, D.E.; Pearson, D.R.; Zachwieja, J.J. Time course of glycogen accumulation after eccentric exercise. J. Appl. Physiol. 1992, 72, 1999–2004. [Google Scholar] [PubMed]
- Ferry, A.; Amiridis, I.; Rieu, M. Glycogen depletion and resynthesis in the rat after downhill running. Eur. J. Appl. Physiol. Occup. Physiol. 1992, 64, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Van der Meulen, J.H.; Kuipers, H.; Stassen, F.R.; Keizer, H.A.; van der Vusse, G.J. High energy phosphates and related compounds, glycogen levels and histology in the rat tibialis anterior muscle after forced lengthening and isometric exercise. Pflugers Arch. 1992, 420, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Hesselink, M.K.; Kuipers, H.; Keizer, H.A.; Drost, M.R.; van der Vusse, G.J. Acute and sustained effects of isometric and lengthening muscle contractions on high-energy phosphates and glycogen metabolism in rat tibialis anterior muscle. J. Muscle Res. Cell Motil. 1998, 19, 373–380. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, K.P.; Warhol, M.J.; Fielding, R.A.; Frontera, W.R.; Meredith, C.N.; Evans, W.J. Eccentric exercise-induced muscle damage impairs muscle glycogen repletion. J. Appl. Physiol. 1987, 63, 252–256. [Google Scholar] [PubMed]
- Hermansen, L.; Hultman, E.; Saltin, B. Muscle glycogen during prolonged severe exercise. Acta Physiol. Scand. 1967, 71, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Maehlum, S.; Hermansen, L. Muscle glycogen concentration during recovery after prolonged severe exercise in fasting subjects. Scand. J. Clin. Lab. Investig. 1978, 38, 557–560. [Google Scholar] [CrossRef]
- Coyle, E.F.; Coggan, A.R.; Hemmert, M.K.; Ivy, J.L. Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate. J. Appl. Physiol. 1986, 61, 165–172. [Google Scholar] [PubMed]
- Ivy, J.L.; Goforth, H.W., Jr.; Damon, B.M.; McCauley, T.R.; Parsons, E.C.; Price, T.B. Early postexercise muscle glycogen recovery is enhanced with a carbohydrate-protein supplement. J. Appl. Physiol. 2002, 93, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Sherman, W.M.; Costill, D.L.; Fink, W.J.; Hagerman, F.C.; Armstrong, L.E.; Murray, T.F. Effect of a 42.2-km footrace and subsequent rest or exercise on muscle glycogen and enzymes. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 1219–1224. [Google Scholar] [PubMed]
- Schiff, H.B.; MacSearraigh, E.T.; Kallmeyer, J.C. Myoglobinuria, rhabdomyolysis and marathon running. Q. J. Med. 1978, 47, 463–472. [Google Scholar] [PubMed]
- Kato, H.; Miura, K.; Nakano, S.; Suzuki, K.; Bannai, M.; Inoue, Y. Leucine-enriched essential amino acids attenuate inflammation in rat muscle and enhance muscle repair after eccentric contraction. Amino Acids 2016, 48, 2145–2155. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Suzuki, H.; Mimura, M.; Inoue, Y.; Sugita, M.; Suzuki, K.; Kobayashi, H. Leucine-enriched essential amino acids attenuate muscle soreness and improve muscle protein synthesis after eccentric contractions in rats. Amino Acids 2015, 47, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Agata, N.; Itoh, Y.; Miyazu-Inoue, M.; Sokabe, M.; Taguchi, T.; Kawakami, K. Stretch speed-dependent myofiber damage and functional deficits in rat skeletal muscle induced by lengthening contraction. Physiol. Rep. 2014, 2, e12213. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, S.S.; Phillips, B.E.; Wilkinson, D.J.; Limb, M.C.; Rankin, D.; Mitchell, W.K.; Kobayashi, H.; Greenhaff, P.L.; Smith, K.; Atherton, P.J. Intake of low-dose leucine-rich essential amino acids stimulates muscle anabolism equivalently to bolus whey protein in older women at rest and after exercise. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E1056–E1065. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, H.; Keizer, H.A.; Verstappen, F.T.; Costill, D.L. Influence of a prostaglandin-inhibiting drug on muscle soreness after eccentric work. Int. J. Sports Med. 1985, 6, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.B.; Warren, G.L.; Warren, J.A. Mechanisms of exercise-induced muscle fibre injury. Sports Med. 1991, 12, 184–207. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, G.; Mikkelsen, U.R.; Raastad, T.; Peake, J.M. Leucocytes, cytokines and satellite cells: What role do they play in muscle damage and regeneration following eccentric exercise? Exerc. Immunol. Rev. 2012, 18, 42–97. [Google Scholar] [PubMed]
- Peake, J.; Nosaka, K.; Suzuki, K. Characterization of inflammatory responses to eccentric exercise in humans. Exerc. Immunol. Rev. 2005, 11, 64–85. [Google Scholar] [PubMed]
- Forster, J.; Morris, A.S.; Shearer, J.D.; Mastrofrancesco, B.; Inman, K.C.; Lawler, R.G.; Bowen, W.; Caldwell, M.D. Glucose uptake and flux through phosphofructokinase in wounded rat skeletal muscle. Am. J. Physiol. 1989, 256, E788–E797. [Google Scholar] [PubMed]
- Bergstrom, J.; Hultman, E. Muscle glycogen synthesis after exercise: An enhancing factor localized to the muscle cells in man. Nature 1966, 210, 309–310. [Google Scholar] [CrossRef] [PubMed]
- Mamedova, L.K.; Shneyvays, V.; Katz, A.; Shainberg, A. Mechanism of glycogen supercompensation in rat skeletal muscle cultures. Mol. Cell. Biochem. 2003, 250, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Asp, S.; Daugaard, J.R.; Kristiansen, S.; Kiens, B.; Richter, E.A. Eccentric exercise decreases maximal insulin action in humans: Muscle and systemic effects. J. Physiol. 1996, 494, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Asp, S.; Daugaard, J.R.; Richter, E.A. Eccentric exercise decreases glucose transporter glut4 protein in human skeletal muscle. J. Physiol. 1995, 482, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Vazquez, I.; Fernandez-Veledo, S.; de Alvaro, C.; Lorenzo, M. Dual role of interleukin-6 in regulating insulin sensitivity in murine skeletal muscle. Diabetes 2008, 57, 3211–3221. [Google Scholar] [CrossRef] [PubMed]
- Di Camillo, B.; Eduati, F.; Nair, S.K.; Avogaro, A.; Toffolo, G.M. Leucine modulates dynamic phosphorylation events in insulin signaling pathway and enhances insulin-dependent glycogen synthesis in human skeletal muscle cells. BMC Cell Biol. 2014, 15, 9. [Google Scholar] [CrossRef] [PubMed]
- Peyrollier, K.; Hajduch, E.; Blair, A.S.; Hyde, R.; Hundal, H.S. L-leucine availability regulates phosphatidylinositol 3-kinase, p70 s6 kinase and glycogen synthase kinase-3 activity in l6 muscle cells: Evidence for the involvement of the mammalian target of rapamycin (mTOR) pathway in the l-leucine-induced up-regulation of system a amino acid transport. Biochem. J. 2000, 350, 361–368. [Google Scholar] [PubMed]
- Doi, M.; Yamaoka, I.; Nakayama, M.; Mochizuki, S.; Sugahara, K.; Yoshizawa, F. Isoleucine, a blood glucose-lowering amino acid, increases glucose uptake in rat skeletal muscle in the absence of increases in amp-activated protein kinase activity. J. Nutr. 2005, 135, 2103–2108. [Google Scholar] [PubMed]
- Liu, H.; Liu, R.; Xiong, Y.; Li, X.; Wang, X.; Ma, Y.; Guo, H.; Hao, L.; Yao, P.; Liu, L.; et al. Leucine facilitates the insulin-stimulated glucose uptake and insulin signaling in skeletal muscle cells: Involving mTORC1 and mTORC2. Amino Acids 2014, 46, 1971–1979. [Google Scholar] [CrossRef] [PubMed]
- Nishitani, S.; Matsumura, T.; Fujitani, S.; Sonaka, I.; Miura, Y.; Yagasaki, K. Leucine promotes glucose uptake in skeletal muscles of rats. Biochem. Biophys. Res. Commun. 2002, 299, 693–696. [Google Scholar] [CrossRef]
- Kalogeropoulou, D.; Lafave, L.; Schweim, K.; Gannon, M.C.; Nuttall, F.Q. Leucine, when ingested with glucose, synergistically stimulates insulin secretion and lowers blood glucose. Metabolism 2008, 57, 1747–1752. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Dolinger, M.; Ritaccio, G.; Mazurkiewicz, J.; Conti, D.; Zhu, X.; Huang, Y. Leucine stimulates insulin secretion via down-regulation of surface expression of adrenergic alpha2a receptor through the mtor (mammalian target of rapamycin) pathway: Implication in new-onset diabetes in renal transplantation. J. Biol. Chem. 2012, 287, 24795–24806. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Suzuki, H.; Inoue, Y.; Takimoto, T.; Suzuki, K.; Kobayashi, H. Co-ingestion of carbohydrate with leucine-enriched essential amino acids does not augment acute postexercise muscle protein synthesis in a strenuous exercise-induced hypoinsulinemic state. Springerplus 2016, 5, 1299. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Murakami, T.; Nakai, N.; Nagasaki, M.; Obayashi, M.; Li, Z.; Xu, M.; Sato, Y.; Kato, T.; Shimomura, N.; et al. Suppression of glycogen consumption during acute exercise by dietary branched-chain amino acids in rats. J. Nutr. Sci. Vitaminol. 2000, 46, 71–77. [Google Scholar] [CrossRef] [PubMed]
- De Araujo, J.A., Jr.; Falavigna, G.; Rogero, M.M.; Pires, I.S.; Pedrosa, R.G.; Castro, I.A.; Donato, J., Jr.; Tirapegui, J. Effect of chronic supplementation with branched-chain amino acids on the performance and hepatic and muscle glycogen content in trained rats. Life Sci. 2006, 79, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Murakami, T.; Nakai, N.; Nagasaki, M.; Harris, R.A. Exercise promotes BCAA catabolism: Effects of BCAA supplementation on skeletal muscle during exercise. J. Nutr. 2004, 134, 1583S–1587S. [Google Scholar] [PubMed]
- Goodman, C.A.; Bennie, J.A.; Leikis, M.J.; McKenna, M.J. Unaccustomed eccentric contractions impair plasma k+ regulation in the absence of changes in muscle Na+,k+-atpase content. PLoS ONE 2014, 9, e101039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rattray, B.; Caillaud, C.; Ruell, P.A.; Thompson, M.W. Heat exposure does not alter eccentric exercise-induced increases in mitochondrial calcium and respiratory dysfunction. Eur. J. Appl. Physiol. 2011, 111, 2813–2821. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Delp, M.D.; Hayes, D.A.; Delp, P.D.; Armstrong, R.B. Rat skeletal muscle mitochondrial [Ca2+] and injury from downhill walking. J. Appl. Physiol. 1990, 68, 1241–1251. [Google Scholar] [PubMed]
- Wojcik, J.R.; Walber-Rankin, J.; Smith, L.L.; Gwazdauskas, F.C. Comparison of carbohydrate and milk-based beverages on muscle damage and glycogen following exercise. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 406–419. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.R.; Ratamess, N.A.; Tranchina, C.P.; Rashti, S.L.; Kang, J.; Faigenbaum, A.D. Effect of a proprietary protein supplement on recovery indices following resistance exercise in strength/power athletes. Amino Acids 2010, 38, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Flakoll, P.J.; Judy, T.; Flinn, K.; Carr, C.; Flinn, S. Postexercise protein supplementation improves health and muscle soreness during basic military training in marine recruits. J. Appl. Physiol. 2004, 96, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Inaguma, A.; Watanabe, S.; Yamamoto, Y.; Muramatsu, Y.; Bajotto, G.; Sato, J.; Shimomura, N.; Kobayashi, H.; Mawatari, K. Branched-chain amino acid supplementation before squat exercise and delayed-onset muscle soreness. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Greer, B.K.; Woodard, J.L.; White, J.P.; Arguello, E.M.; Haymes, E.M. Branched-chain amino acid supplementation and indicators of muscle damage after endurance exercise. Int. J. Sport Nutr. Exerc. Metab. 2007, 17, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Jackman, S.R.; Witard, O.C.; Jeukendrup, A.E.; Tipton, K.D. Branched-chain amino acid ingestion can ameliorate soreness from eccentric exercise. Med. Sci. Sports Exerc. 2010, 42, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Jackman, S.R.; Witard, O.C.; Philp, A.; Wallis, G.A.; Baar, K.; Tipton, K.D. Branched-chain amino acid ingestion stimulates muscle myofibrillar protein synthesis following resistance exercise in humans. Front. Physiol. 2017, 8, 390. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato, H.; Miura, K.; Suzuki, K.; Bannai, M. Leucine-Enriched Essential Amino Acids Augment Muscle Glycogen Content in Rats Seven Days after Eccentric Contraction. Nutrients 2017, 9, 1159. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9101159
Kato H, Miura K, Suzuki K, Bannai M. Leucine-Enriched Essential Amino Acids Augment Muscle Glycogen Content in Rats Seven Days after Eccentric Contraction. Nutrients. 2017; 9(10):1159. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9101159
Chicago/Turabian StyleKato, Hiroyuki, Kyoko Miura, Katsuya Suzuki, and Makoto Bannai. 2017. "Leucine-Enriched Essential Amino Acids Augment Muscle Glycogen Content in Rats Seven Days after Eccentric Contraction" Nutrients 9, no. 10: 1159. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9101159