Benefits of Nut Consumption on Insulin Resistance and Cardiovascular Risk Factors: Multiple Potential Mechanisms of Actions



School of Pharmacy and Medical Sciences, University of South Australia, General Post Office Box 2471, Adelaide, SA 5001, Australia
*
Author to whom correspondence should be addressed.
Nutrients 2017, 9(11), 1271; https://0-doi-org.brum.beds.ac.uk/10.3390/nu9111271
Submission received: 15 September 2017
/
Revised: 2 November 2017
/
Accepted: 14 November 2017
/
Published: 22 November 2017
(This article belongs to the Special Issue Nut Consumption for Human Health)
Abstract
:Epidemiological and clinical studies have indicated that nut consumption could be a healthy dietary strategy to prevent and treat type 2 diabetes (T2DM) and related cardiovascular disease (CVD). The objective of this review is to examine the potential mechanisms of action of nuts addressing effects on glycemic control, weight management, energy balance, appetite, gut microbiota modification, lipid metabolism, oxidative stress, inflammation, endothelial function and blood pressure with a focus on data from both animal and human studies. The favourable effects of nuts could be explained by the unique nutrient composition and bioactive compounds in nuts. Unsaturated fatty acids (monounsaturated fatty acids and polyunsaturated fatty acids) present in nuts may play a role in glucose control and appetite suppression. Fiber and polyphenols in nuts may also have an anti-diabetic effect by altering gut microbiota. Nuts lower serum cholesterol by reduced cholesterol absorption, inhibition of HMG-CoA reductase and increased bile acid production by stimulation of 7-α hydroxylase. Arginine and magnesium improve inflammation, oxidative stress, endothelial function and blood pressure. In conclusion, nuts contain compounds that favourably influence glucose homeostasis, weight control and vascular health. Further investigations are required to identify the most important mechanisms by which nuts decrease the risk of T2DM and CVD.
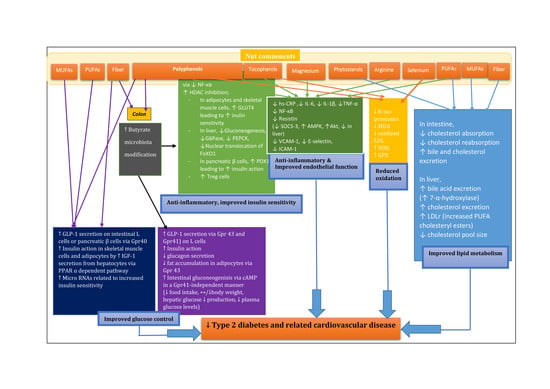
1. Introduction
The prevalence of diabetes mellitus will increase worldwide from 382 million individuals in 2013 to 592 million individuals in 2035. Type 2 diabetes mellitus (T2DM) accounts for 90–95% of individuals with diabetes mellitus [1]. T2DM is attributable to poor diet, obesity, age, genetics, smoking, sedentary lifestyle and hypertension [2,3]. T2DM is associated with an increased risk of adverse cardiovascular events [3]. A 1 mmol/L increase in fasting plasma glucose is related to a 17% increase in the risk of developing of cardiovascular disease (CVD) and death [4]. Individuals with diabetes have a 2- to 4-fold higher risk of CVD than individuals without diabetes [5]. As a consequence, T2DM dramatically increases health care costs and disease burden [6]. Lifestyle modification including diet can decrease the risk of developing T2DM [7]. A large number of epidemiological and clinical studies have shown that consumption of different types of nuts (almonds, walnuts, hazelnuts, pecans, pistachios, macadamia nuts, cashews and Brazil nuts) is associated with a reduced risk of T2DM [8,9,10] and CVD [8,9,10,11,12].
This review aims to address the potential mechanisms of nut consumption on the prevention and treatment of T2DM and CVD, focusing on body weight control, glucose control, gut microbiota composition, inflammation, oxidative stress, lipid profiles, endothelial function and hypertension. All human interventions available on PubMed that covered these areas were included. This is the only systematic review that covers all these areas.
2. Nut Composition
Nuts are an energy dense food (Table 1). Nuts consist of fat (43–67% by weight), protein (8–22%), ash (1–3%), total soluble sugars (0.6–4%), polyphenols (0.2–1.6%), phytates (0.2–0.4%) [13]. Nuts also contain bioactive components such as lipids (carotenoids and phytosterols), vitamins and minerals (particularly magnesium, folate and potassium).
Nuts predominantly contain unsaturated fatty acids (monounsaturated fatty acids (MUFAs) and polyunsaturated fatty acids (PUFAs)) with a low amount of saturated fatty acids (SFAs—only 4–5%). Oleic acid (C18:1) is the major MUFA in all nuts [13]. Linoleic acid (C18:2n-6) is the major PUFA in nuts except for macadamias [13]. Walnuts have the highest content of PUFAs at 47%, 38% linoleic acid and 9% linolenic acid (C18:3n-3). Nuts are also a good source of polyphenols and fiber (especially nuts covered with skin). In the US diet in 2008, nuts provided 162 mg/day of polyphenols (19%) compared with 204 mg/day from vegetables (24%), 213 mg/day from grains (25%) and 223 mg/day from fruits (26%), with similar figures in Europe [14]. Pistachios contain the highest quantity of lutein and zeaxanthin which are xanthophyll carotenoids. These carotenoids appear to protect from age-related macular degeneration [15].
3. Glycemic Control
A meta-analysis of 12 randomised controlled trials (RCTs) with a ≥3-week follow-up period in subjects with T2DM comparing a diet supplemented with tree nuts (almonds, Brazil nuts, cashews, hazelnuts, macadamia nuts, pecans, pine nuts, pistachios and walnuts) and an isocaloric diet without tree nuts, showed that consumption of tree nuts at a median dose of 56 g/day improved glycemic control in subjects with T2DM, showing significantly decreased glycosylated haemoglobin levels (HbA1c; mean difference 0.07%, p = 0.0003) and fasting glucose (p = 0.03), with no effect on fasting insulin or homeostasis model assessment of insulin resistance index (HOMA-IR). Most RCTs were of poor quality and had short study periods [20]. Human interventions evaluating the effect of nut consumption on glycemic control are shown in Table 2. In summary, given the results from the meta-analysis of 12 RCTs [20] and chronic studies [21,22,23,24,25,26] that were not included in the meta-analysis [20], nut consumption benefits glycemic control regardless of the type of nut but the effect is small and the amount of nuts required is large. Moreover, nut consumption reduces postprandial glycemia responses as shown in all nine studies [27,28,29,30,31,32,33,34,35].
A randomized control clinical trial in subjects with T2DM showed that consumption of walnut oil (15 g/day for 3 months) significantly reduced glycosylated HbA1c by 8 ± 22% (p = 0.005) and fasting glucose levels by 8 ± 17% (p = 0.001) compared with the baseline while a control group showed no significant differences in HbA1c and fasting glucose levels. [21].
Pistachio consumption improved glycemic status in subjects with T2DM [22,23] and in healthy young men [24]. In the 8-week dietary intervention trial [24], healthy young men aged on average 22 years consumed a Mediterranean diet for 4 weeks and in the next 4 weeks they supplemented the Mediterranean diet with pistachios for 4 weeks replacing 32% of the energy obtained from MUFAs in the control diet (20% of energy). A Mediterranean diet rich in pistachios improved glucose levels (−8.8 ± 8.5% p < 0.001), low-density lipoprotein cholesterol (LDL-C; p < 0.001), total cholesterol (TC; p < 0.001), triacylglycerol (p = 0.008), TC-C/high density lipoprotein cholesterol (HDL-C) ratio (p < 0.001), LDL-C/HDL-C ratio (p < 0.001) and endothelium-dependent vasodilation (p = 0.002; 30% relative increase). In addition, the pistachio diet significantly decreased serum oxidation (lipid hydroperoxide (p < 0.001) and malondialdehyde (MDA; p < 0.001)) and inflammatory markers (interleukin-6 (IL-6; p < 0.001)) and increased superoxide dismutase (SOD—a superoxide anion radical (O2−) scavenger; p < 0.001) in comparison with a Mediterranean diet rich in vegetable and fish and limited in red meat, fat and egg products [24].
Almond consumption improved glucose control in subjects with impaired fasting glucose [25] and prediabetes [36], while one study showed no effect of an almond diet on glucose and lipid profiles in subjects with T2DM [37]. In a randomized parallel trial [36], subjects with prediabetes who followed an ADA (American Diabetes Association) diet containing 20% of energy from almonds (60 g/day of pre-packaged raw or dry roasted almonds) for 16 weeks showed a significant reduction in fasting insulin concentrations (−23% vs. +19%; p = 0.002), HOMA-IR (−25% vs. +0.3%; p = 0.007) and homeostasis model analysis for beta-cell function (HOMA-B; −18% vs. +30.0%; p = 0.001) with no alteration in fasting glucose, compared with an almond-free control group. LDL-C concentrations were significantly lowered after the almond diet compared with the control diet containing 60–70% carbohydrate and MUFA together, 15–20% protein, 10% saturated fat, and 300 mg/day cholesterol. Adjustment for weight did not abolish changes in insulin, HOMA-IR, and HOMA-B. Subjects in the almond group were instructed to use the ADA Food Exchange System for 80% of their energy needs. The average body mass index (BMI) was 30 ± 5 kg/m2 for the almond group (n = 32) and 29 ± 5 kg/m2 for the control group (n = 33). Energy intake was limited for 14 subjects with a BMI > 25 kg/m2 out of 65 subjects following the ADA’s recommendations of modest weight loss in people with prediabetes. No significant differences in weight, BMI and waist circumference were observed between two groups at weeks 0, 4, 8, 12, and 16, even though the reductions in these measurements occurred in both groups over the course of the study (the average weight loss was 1.1 kg for the almond group and 2.0 kg for the control group) [36].
The negative study [37], which was included in the meta-analysis of T2DM subjects [20], consisted of two studies to assess insulin sensitivity. Both studies showed no effect on insulin sensitivity in healthy subjects or on glycemic control in subjects with T2DM, but beneficial effects were seen on lipid profiles. In study 1, 20 healthy adults were instructed to adhere to their habitual diet supplemented with 100 g almonds/day for 4 weeks. No changes in fasting glucose and fasting insulin, and insulin sensitivity index (SI) and glucose effectiveness (SG) as calculated using the Minimal Model method were observed before and after the diet. Body weight significantly increased (p = 0.006) and a significant time-by-sex interaction for SI and SG was observed due to an increase in women with a decrease in men. Changes in body weight had no effect on insulin sensitivity. In study 2 using a randomised, double-blind, crossover design, volunteers were randomly assigned to begin one of four diets for 4 weeks: high-fat, high-almond (37% total fat, 10% from almonds); low-fat, high-almond (25% total fat, 10% from almonds); high-fat control (37% total fat, 10% from olive oil or canola oil); low-fat control (25% total fat, 10% from olive or canola oil). The almond diets contained 57–113 g almonds/day based on the total energy requirements. No changes in fasting glucose, fasting insulin, and 2-h glucose and insulin as assessed by oral glucose tolerance test were observed overall with almonds, while a significant interaction between fat source and fat level for both fasting and 2-h glucose levels was observed due to the lower glucose levels on the low fat, high almond diet than a high fat control diet [37].
3.1. Acute Studies
A favourable effect of pistachios on postprandial glycemia was seen in 10 healthy adults [27] and 20 subjects with metabolic syndrome [28]. Two acute meal studies [29,30] examined the glucose-lowering effect of peanuts. Peanut consumption significantly reduced the 60-min postprandial glucose response to high-glycemic load meals in healthy subjects [29,30]. Five acute meal studies with a crossover design examined the effect of almond intake on postprandial glycemia and insulinemia [31,32,33,34,38]. Almonds (28 g) with a test meal (bagel, juice, and butter) significantly lowered postprandial glycemia in seven subjects with T2DM by 30% (p = 0.04), but not in 13 healthy subjects without T2DM, with no differences in insulinemia and the incretin hormone, glucagon-like peptide-1 (GLP-1) [38]. On the other hand, four acute meal studies showed attenuated postprandial glycemia in impaired glucose tolerant [31], healthy [32,33,35] and diabetic [35] subjects.
3.2. Possible Nutrients Involved in Glucose-Lowering Effects in Nuts
A recent meta-analysis of 102 RCTs showed the substitution of carbohydrates and SFAs with a diet rich in unsaturated fat, particularly PUFAs improved glucose control. The substitution of carbohydrates with PUFAs significantly decreased HbA1c (−0.11%; 95% CI: −0.17, −0.05) and fasting insulin (−1.6 pmol/L; 95% CI: −2.8, −0.4). Substitution of carbohydrates with MUFAs decreased HbA1c (−0.09%; 95% CI: −0.12, −0.05), 2-h postprandial insulin (−20 pmol/L/min; 95% CI: −32, −8), and HOMA-IR (−2%; 95% CI: −5, −0.3). The substitution of SFAs with MUFAs significantly lowered HbA1c (−0.12%; 95% CI: −0.19, −0.05) and HOMA-IR (−3%; 95% CI: −6, −0.4), while the substitution of SFAs with PUFAs significantly lowered HbA1c (−0.15%; 95% CI: −0.2, −0.06), C-peptide (−0.07%; 95% CI: −0.14, −0.01), HOMA-IR (−4%; 95% CI: −6, −2) and 2-h postprandial insulin responses (0.5 pmol/L/min; 95% CI: 0.2, 0.8) [39]. Alpha-linolenic acid (ALA an-3 PUFA; 18:3n-3; a precursor of eicosapentaenoic acid (EPA; 20:5n-3) and docosahexaenoic acid (DHA; 22:6n-3) has been shown to reduce fasting glucose levels in a meta-analysis of 12 RCTs with a ≥4-week intervention and a ≥4-week washout period [40] and to decrease the prevalence of insulin resistance in middle-aged Japanese but only in those with a normal weight [41]. ALA stimulates GLP-1 release from pancreatic β-cells [42,43], from intestinal l-cells [42] through activation of GPR40, one of the G-protein-coupled free fatty acid receptors also called free fatty acid receptor 1 (FFAR1). In addition, ALA induces insulin-like growth factor 1 (IGF-1) gene expression and release from hepatocytes via a peroxisome proliferator-activated receptor α (PPAR α) dependent pathway [44,45] resulting in improved insulin action primarily in skeletal muscle, and in adipose tissue [45].
In summary, unsaturated fatty acids in nuts in place of both carbohydrate and saturated fat appear to improve glucose levels by a small amount but more studies at the molecular level should be undertaken along with a detailed investigation of different nut types and amounts, study duration, background diet, subject ethnicity in subjects with T2DM and at risk of T2DM.
3.3. Modification in Micro RNAs Related to Insulin Sensitivity
Only two nut intervention studies have explored microRNA modification in relation to T2DM risk [46,47]. Ortegaa et al. [46] found significant modification of several common miRNAs in plasma from 10 healthy women who consumed a PUFA-enriched normocaloric diet containing 15 g/day of almonds and 15 g/day of walnuts (55–60% carbohydrates 15% proteins, 30% fat, 10% SFA, 10–15% MUFAs and 10% PUFAs) for 8 weeks. The changes of plasma miR-106a were correlated with changes in circulating PUFAs. The changes in plasma miR-130b and miR-221 (r = 0.46, p = 0.03) were correlated with changes in plasma C-reactive protein. The changes in plasma miR-125a-5p were correlated with changes in plasma fasting triglycerides and adiponectin [46]. Very recently, a 4-month diet supplemented with 57 g/day of pistachio (50% carbohydrates, 33% fat) modified circulating microRNAs (miR-192 and miR-375) and these changes were positively correlated with plasma glucose, insulin and HOMA-IR in 49 subjects with prediabetes [47]. Thus, microRNAs can be modulated by nut consumption and may be involved in glucose metabolism, lipid metabolism and inflammation. Circulating miRNAs have stable and reproducible levels in serum and may be useful biomarkers examining molecular mechanisms in T2DM and CVD [48].
4. Body Weight Control, Appetite, Energy Balance and Lipid Bioaccessibility
Nuts may be an effective appetite suppressant to prevent body weight gain. In addition, nuts may enhance energy expenditure and reduce energy absorption [37,49,50,51,52,53,54].
4.1. Body Weight Control
The high energy density and fat content of nuts has raised concerns that regular nut consumption will cause body weight gain. However, epidemiological studies indicated either an inverse [55,56,57,58] or no association [59,60,61] between nut consumption and BMI or body fat level. Most randomised crossover interventions investigating the effect of adding nuts to a habitual diet on body weight as a primary endpoint reported either no weight gain or lower weight gain than expected from the additional energy intake from the nut consumption (peanuts [49], almonds [37,50,51,53,62,63,64,65], walnuts [52,53,66,67,68,69], pecans [70], pistachios [71,72,73,74], macadamia nuts [75,76] and hazelnuts [77]).
Human interventions evaluating the effect of nut consumption on body weight control are shown in Table 3. Seventeen of 21 clinical studies in Table 3 showed no change in body weight but five [37,49,50,66,77] of 21 studies showed body weight gain ranging from 0.4 to 1 kg regardless of the types of nuts. Four studies [26,78,79,80] showed body weight reduction.
The PREDIMED study showed that adherence to the Mediterranean diets enriched with either 50 mL/day (350 mL/week) of extra virgin olive oil or 30 g/day of nuts for 3 years resulted in a reduction in body weight. The increased energy density was not associated with body weight gain [78]. Other short-term interventions also showed a greater weight loss effect of a diet enriched in nuts in comparison with a low-fat diet [79,80], or a calorie-matched complex carbohydrate diet [26]. In a randomized controlled trial comparing the effect of a moderate fat low energy diet (fat: 35% of energy; predominantly PUFAs), with a low-fat, low-energy diet (fat: 20% of energy) on weight loss in 101 overweight subjects for 18 months, the nut-enriched diet resulted in greater sustained weight reduction at 18 months compared with a low-fat diet [79]. Supplementation with either a high dose (70 g/day) or a recommended dose of pistachio nuts (42 g/day) for 12 weeks in a parallel design did not alter BMI or waist-to-hip ratio compared with a control diet in Chinese subjects with metabolic syndrome [73].
In summary, it would appear that nuts can be added to a diet without significant body weight gain under most circumstances.
4.2. Appetite
Two studies showed a favourable effect on appetite of almond snacks in subjects at increased risk of T2DM [81] and in healthy women [82]. In a 4-week randomized, parallel-arm study, 43 g almonds/day with breakfast or lunch, or alone as a morning or afternoon snack for 4 weeks, attenuated postprandial glucose responses and decreased hunger and desire to eat in subjects with increased risk for T2DM [81].
Cholecystokinin (CCK), GLP-1 and peptide YY (PYY) are gut satiety hormones. Cholecystokinin (CCK-8) is released from duodenal enteroendocrine cells in response to fatty acid and protein. GLP-1 and PYY are secreted from the ileum in response to carbohydrates, proteins and fatty acids. Ghrelin is also a gut hunger hormone released from the stomach which is suppressed by food intake. It increases appetite [83,84].
A study consisting of an in vitro (STC-1 cell–intestinal secretin tumor cell line) study and a human intervention suggested Korean pine nuts containing ≥92% PUFA (15% pinolenic acid (C18:3), linoleic acid (C18:2) and MUFAs (oleic acid (C18:1)) might be a good appetite suppressant (assessed by decreased prospective food intake) by enhancement of CCK-8 and GLP-1 secretion [85].
A favourable acute and second meal effect of added nuts was seen with almonds ingested by subjects with impaired glucose tolerance [31] and peanuts consumed by obese women with a high T2DM risk [29] with breakfast on postprandial glucose responses and appetite.
Leptin is a hormone secreted from adipocytes which regulates energy balance by suppressing appetite along with other metabolic effects. Leptin plays an opposite role to that of ghrelin [86]. A recent meta-analysis of 20 RCTs showed that nut consumption (tree nut, peanut, and soy nut) significantly reduced leptin but did not significantly alter high sensitivity C-reactive protein (hs-CRP), IL-6, adiponectin, IL-10, and tumor necrosis factor-α (TNF-α) [87].
4.3. Energy Balance
Minimal [37,49,50,66,77] or no body weight gain [51,52,53,62,63,64,65,67,68,69,70,71,72,73,75,76] following nut consumption may also be due to increased energy expenditure. Several studies [49,54,90] have reported an increased resting metabolic rate which could be attributed to both the high unsaturated fat in nuts as well as the protein. A Mediterranean diet supplemented with nuts decreased waist circumference after 4.8 years of follow-up compared with a low-fat diet [91].
4.4. Bioaccessibility of Nutrients
Bioaccessibility refers to the proportion of nutrients released from a complex food matrix, which become potentially available for absorption in the gastrointestinal tract [92]. One proposed mechanism of low or no weight gain with nuts includes a reduced level of lipid bioaccessibility from nuts. The properties of cell walls and fibre-rich skins in nuts may play a role in the rate and extent of lipid release [93].
Several studies [93,94] have investigated this question. In the first study [93], only the first layer of cells at the fracture surface was ruptured by chewing and released lipids. In faecal samples from healthy subjects who consumed the almond rich diet, intact almond tissues, intact cotyledon cells containing intracellular lipid, increased lipid concentrations, and large numbers of bacteria adherent to the cotyledon cells were observed [93]. Cell walls of almonds consist of non-starch polysaccharides (especially, arabinose-rich polysaccharides) and phenolic compounds (predominantly, protocatechuic acid, p-hydroxybenzoic acid, and vanillic acid).
Other intervention studies also showed an increased excretion of stool fat after consumption of almonds [51,88,95], pecans [96] and peanuts [97,98].
The poor lipid bioaccessibility of almonds reduces postprandial lipidemia which may be associated with a lower risk of coronary heart disease [99,100,101]. A randomised crossover study [102] in healthy human subjects compared 54 g fat in four different meals (whole almond seed, almond oil and defatted almond flour, or a sunflower oil blend), and showed a favourable effect of whole almond seed on postprandial lipidemia compared with other forms of lipid. However, no significant differences in insulin, 8-isoprostane F2 α (a marker of oxidative stress) and peripheral augmentation index (a measure of vascular tone) were seen between meals. The iAUC for plasma glucose, however, was higher on the whole almond seed meal than on the almond oil and defatted almond flour meal [102].
5. Gut Microbiota Modification
Fiber and polyphenols (flavonoids and nonflavonoids) abundant in nuts may exert a prebiotic effect and influence glucose metabolism [103]. Nuts with the highest contents of polymerized polyphenols are hazelnuts and pecans followed by pistachios, almonds and walnuts [104]. Walnuts, pecans and chestnuts are high in ellagitannins (hydrolysable tannins), whereas proanthocyanidins (condensed tannins) are abundant in the kernel of nuts such as hazelnuts, pecans, pistachios, almonds and cashew [19].
Proanthocyanidins are oligomers or polymers of polyhydroxy flavan-3-ol units, such as (−)-epicatechin, and (+)-catechin [105]. Epidemiological studies have shown an inverse association between proanthocyanidin intake and incident T2DM [106,107]. No dietary intervention examining the prebiotic effects of nut-derived proanthocyanidin has been performed while two dietary interventions have shown a favourable prebiotic effect of proanthocyanidin derived from grape seeds [108] and cocoa [109] modifying the proportion of Bifidobacterium and Enterobacteriaceae [108,109] which was not dose related. The doses of proanthocyanidins administered were 190 mg/day for healthy adults aged 40–59 years for 2 weeks [108], 380 mg/day for elderly adults aged 67–98 years for 2 weeks [108], and 494 mg/day for young healthy adults with a mean age of 30 years for 4 weeks [109].
5.1. Urolithin (Microflora Metabolite)
Walnuts and pecans which have the highest total phenol contents are the richest sources of ellagic acid that possesses 2,2-diphenyl-1-picrylhydrazyl (DPPH•) scavenging capacities [19]. Brazil nuts contain no polyphenols nor have an antioxidant capacity [19]. Ellagitannins and ellagic acid are not absorbed in the gastrointestinal tract but are metabolized by the human gut microbiota to urolithins (dibenzo[b,d]pyran-6-one derivatives with different hydroxyl substitutions) which are much better absorbed than the parent ellagitannins and ellagic acid [110] while the type of urolithin formed depends on particular species in the gut microflora [111]. There is a wide range of urolithins based on a decreasing number of phenolic hydroxyl groups; urolithin D→C→A→B [112]. Aromatic and phenolic structures including phenylvalerolactones and phenylvaleric, phenylpropionic, phenylacetic, hippuric, and benzoic acids, with different hydroxylation patterns which are microbial metabolites of flavan-3-ol dimers, oligomers, flavan-3-ol polymers [113,114,115,116]. Hydroxyphenylvalerolactones in plasma or urine are proposed to be the main metabolites of flavan-3-ol compounds as they come from reactions of the flavonoid C-ring-opening followed by lactonization [113,114]. In vitro and in vivo studies have shown beneficial effects of urolithins on oxidation, inflammation and glycation [110,117].
In a randomized parallel intervention in subjects with the metabolic syndrome comparing a 12-week healthy diet supplemented with 30 g/day mixed nuts (15 g walnuts, 7.5 g almonds and 7.5 g hazelnuts) with a healthy diet without nuts [118,119,120,121], a healthy diet with added nuts significantly increased walnut ellagitannin-derived urolithins A and B in urine [118,119], significantly improved fasting insulin, insulin sensitivity assessed by HOMA-IR, and IL-6 and significantly decreased DNA damage assessed by urinary 8-oxo-7,8-dihydro-2′-deoxyguanosine (a measure of oxidative stress) [121]. Adjustment for weight loss attenuated the statistical significance of IL-6 (p = 0.08) [120]. No effects were seen on cholesterol levels [120], biomarkers of oxidative stress (plasma oxidized LDL, plasma conjugated diene and urinary 8-isoprostanes), antioxidant capacity or endothelial function determined by non-invasive peripheral artery tonometry [121].
In a study with a metabolomic approach, nut consumption resulted in increased metabolism and excretion of unsaturated fatty acids with increased detection of fatty acid conjugated metabolites and increased excretion of serotonin metabolites in 24-h urine samples [118].
In summary, urolithins are phenolic metabolites which are increased following nut consumption, but to our knowledge, no human intervention study has investigated the prebiotic effect of nut derived urolithins. Therefore, it is unclear what role urolithins play in decreasing the risk of T2DM.
5.2. Butyrate (Microflora Metabolite)
A few studies have examined the effect of nut consumption on gut microbiota and the gut metabolome [122,123,124,125]. In vitro, chestnut extract enhanced the viability of Lactobacillus strains in a simulated GI tract [122]. In vitro using a model stomach, small intestine and colon finely ground almond seeds significantly increased the growth of bifidobacteria and Eubacterium rectale with a butyrogenic prebiotic effect [123].
Butyrate is a short chain fatty acid (SCFAs). SCFAs stimulate the expression of peptide YY via the G-protein-coupled receptors (Gpr41 and Gpr43) leading to the inhibition of gut motility and suppression of appetite [126,127]. Activation of G-protein-coupled receptors (Gpr43; and Gpr41) on l-cells by SCFAs triggers the secretion of GLP-1 which improve glucose homeostasis by increasing insulin and decreasing glucagon secretion [128,129]. SCFAs play a role in controlling body energy utilization by suppressing insulin signalling in adipocytes and fat accumulation in adipose tissues through Gpr43 activation [130]. Intestinal gluconeogenesis (IGN) may have a favourable impact on the risk of T2DM by decreasing food intake, body weight, hepatic glucose production and plasma glucose [131,132,133]. Butyrate promotes IGN via cyclic adenosine monophosphate (cAMP) in a Gpr41-independent manner, while propionate (a longer SCFA) promotes IGN via a gut-brain neuronal circuit by binding to Gpr41 [134].
Anti-inflammatory effects of butyrate have been demonstrated through two pathways: nuclear factor-κB (NF-κB) and histone deacetylase (HDAC) inhibition [135,136]. Butyrate suppresses the expression of inducible nitric oxide synthase (iNOS), IL-6 and TNF-α by downregulating NF-κB and extracellular signal-regulated kinase (ERK) signalling pathways in part with no involvement of the Jak/STAT pathway in IFN-γ induced-RAW 264.7 murine macrophage cells [137]. Butyrate also acts as a histone deacetylase (HDAC) inhibitor [135,136,138,139]. HDAC inhibitors could be a potential T2DM therapy [140,141]. HDACs inhibit the expression of glucose transporter type 4 (GLUT4) leading to decreased insulin sensitivity in adipocytes and muscle cells [140,141]. In the liver, HDACs stimulate forkhead box O (FoxO) DNA-binding leading to facilitated gluconeogenetic gene expression (e.g., glucose-6-phosphatase (G6Pase) and phosphoenolpyruvate carboxykinase (PEPCK)) [142], and HDACs act on a signal transducer and activator of transcription 3 (STAT3)-mediated gluconeogenesis [143]. In pancreatic β-cells, HDAC1 reduced the expression of duodenal homeobox 1 (PDX1) leading to a decreased expression of insulin [144,145]. Butyrate also induced regulatory T cells (Treg) that contribute to shutting down inflammatory responses, possibly by enhancing histone H3 acetylation and stimulating the expression of the transcription factor forkhead box P3 (FOXP3) in TReg cells [146,147,148,149]. In addition, butyrate attenuates MCP-1 levels in human PBMC [150] and represses the expression of vascular cell adhesion molecule-1 (VCAM-1) in human umbilical vein endothelial cells (HUVEC) [151,152].
Ukhanova et al. [124] conducted two separate randomised, controlled, crossover nut feeding studies in healthy subjects (n = 18 for an almond group; n = 16 for a pistachio group) who consumed 0, 43 g and 86 g per day for 18 days. Analyses of faecal samples using a 16S rRNA-based approach showed that nuts altered the composition of the faecal bacterial and fungal microbiota with an increase in butyrate producing bacteria. This effect was much stronger in the pistachio group. Neither nut altered Lactobacillus or Bifidobacteria levels [124]. An intervention with a much longer period (6 weeks) in healthy subjects indicated that whole almond or almond skin could beneficially modify gut microbiota and bacterial activities [125]. In this study [125], daily consumption of 56 g roasted, unsaltednwhole almonds or 10 g almond skin significantly increased populations of Bifidobacterium spp. and Lactobacillus spp., and suppressed the growth of Clostridium perfringens, leading to changes in bacterial enzyme activities (increased faecal β-galactosidase and decreased faecal β-glucuronidase, nitroreductase and azoreductase) [125].
In summary, butyrate is a major product of a nut-related change in the microbiota. Butyrate appears to be a strong candidate molecule which can be mechanistically linked to a reduced T2DM risk.
6. Changes in Blood Lipids and Lipoproteins
Nut consumption can favourably alter blood lipids and lipoproteins. Mediterranean diets enriched with 30 g/day of nuts after 1 year of follow up in subjects with a high CVD risk showed an increase in cholesterol efflux capacity and an increase in the percentage of large HDL-C particles [153].
Meta-analyses of nut intervention studies showed a favourable alteration in lipid profiles and apolipoproteins [9,154,155,156]. A meta-analysis of 61 interventions with the study period ranging from 3 to 26 weeks [9] showed that nut consumption reduced TC (−4.7 mg/dL), LDL-C (−4.8 mg/dL), apolipoprotein B (apo B) (−3.7 mg/dL), and triglyceride (TG; −2.2 mg/dL) regardless of type of nut or background diets. A favourable effect on Apo B was stronger in individuals with T2DM. An association between tree nut consumption and a reduction in TC and LDL-C was seen at ≥60 g/day of nuts. The LDL-C lowering effect of 100 g/day of nuts was up to 35 mg/dL (0.9 mmol/L) [9]. Coronary deaths may be reduced by 20% (95% CI: 0.76, 0.85) per 1.0 mmol/L reduction in LDL-C [157]. Moreover, a separate meta-analysis of only almonds [154] or only walnut [156] intervention studies showed a significant reduction in TC, LDL-C and TG and no alteration in HDL-C [154] and reduction in TC, LDL-C and no alteration in HDL-C and TG compared with the diet [156]. Incorporation of 50–100 g/day nuts providing 35% of energy to a low-fat diet five or more times per week can lower TC and LDL-C for the prevention of CVD [158,159,160].
In a randomised, cross-over, controlled study, which was not included in the meta-analyses [9,154,155,156], comparing three 4-week iso-energetic dietary regimes in subjects with an increased risk of CVD, a diet providing 20% of energy from pistachios (63–126 g/day of pistachios; 34% total fat and 8% SFA) significantly reduced small, dense LDL-C levels (sdLDL, associated with more CVD events than total LDL-C) and the triacylglycerol:HDL-C ratio compared with the low-fat control diet (control; 25% total fat and 8% SFA), as well as improved ATP-binding cassette transporter A1 (ABCA1)-mediated serum cholesterol efflux capacity but only in participants with low hs-CRP status at baseline (<10 mg/L), compared with a diet providing 10% of energy from pistachios (32–63 g/day of pistachio; 30% total fat and 8% SFA) [74].
Favourable effects of nut consumption on lipid profiles could in part result from unsaturated fat or bioactive compounds including plant sterols and dietary fibre [158,161]. Meta-analyses of prospective cohort studies showed that PUFAs in nuts (especially in walnuts) and ALA (n-3 PUFA; 18:3n-3) were inversely associated with the risk of CVD [162,163].
6.1. Unsaturated Fat
Unsaturated fat in nuts may alter the composition of very low density lipoprotein (VLDL) [164] and the activities of lipoprotein lipase and hepatic lipase [165,166] and decrease apo C-III in HDL.
Park et al. demonstrated that PUFAs decreased the expression of cholesterol transporter NPC1Ll in Caco-2 cells (human colon adenocarcinoma cell line) and HepG2 cells (human liver carcinoma cell line) compared with SFAs and phytosterols [167]. Harini et al. [168] proposed several potential mechanisms for PUFA to lower lipids. The expression of several genes involved in lipid metabolism can be mediated by PUFAs via nuclear receptors including the nuclear receptors (PPAR), liver X receptor (LXR), and hepatocyte nuclear factor-(HNF)-4α], via NFκB and the transcription factors sterol-regulatory element binding protein (SREBP) [168].
6.2. The Lysine to Arginine Ratio
One potential mechanism of the hypolipidemic effects of nuts may be the low content of lysine and the relatively high content of arginine. In animals, a diet with a low Lys: Arg ratio promoted 7-α hydroxylase activity resulting in elevated bile acid production and excretion of neutral and acidic steroids decreasing the hepatic cholesterol pool size. This can result in a decrease in LDL-C [169,170].
6.3. Phytosterols
A meta-analysis of RCTs showed that a daily intake of approximately 3 g phytosterols lowered LDL-C concentrations by 12% [171]. Possible molecular mechanisms have been proposed but they are still controversial [172]. Phytosterols are plant-specific phytochemicals and their chemical structure is similar to that of cholesterol. Phytosterols have been known to compete against dietary cholesterol and biliary cholesterol incorporation into mixed micelles in the intestinal lumen [172,173,174].
Intestinal transport proteins and receptors have also been proposed as a mechanism underlying the cholesterol-lowering effect of phytosterols [172]. The NPC1L1 (a target of ezetimibe which lowers cholesterol uptake) is a protein expressed in jejunal enterocytes and localized to the brush border membrane and which is required for intestinal cholesterol absorption and phytosterol absorption [175,176].
On the other hand, the heterodimer of ATP-binding cassette (ABC) transporters, G5 (ABCG5) and G8 (ABCG8) inhibit the absorption of cholesterol and phytosterols from the diet by driving the efflux of cholesterol and phytosterols from enterocytes back into the intestinal lumen, and by facilitating the release of cholesterol and phytosterols from hepatocytes into bile [177].
Intestinal acyl CoA: cholesterol acyltransferase (ACAT2) converts cholesterol to cholesteryl ester which is packed into chylomicrons by microsomal triglyceride protein (MTP—a lipid transfer protein) for secretion into chyle [178]. In hamsters fed a high cholesterol diet, β-sitosterol and stigmasterol down-regulated mRNA levels of intestinal ACAT2 and MTP [179]. Brauner et al. [180] demonstrated that phytosterols have lower uptake and lower esterification by ACAT2 in the intestine than cholesterol. In Caco-2 enterocytes, phytosterols decreased cholesterol absorption by suppressing 27-hydroxycholesterol generation by CYP27, liver X receptor α (LXRα) and the basolateral sterol exporter ABCA1 expression, and delivering sterol back into the gut lumen by ABCG5/G8 [180].
3-hydroxy-3-methylglutary-CoA (HMG-CoA) reductase is the rate-limiting enzyme in cholesterol synthesis. HMG-CoA reductase gene expression was reduced in CaCo-2 cells incubated with beta-sitosterol [181]. An in vitro study showed that stigmasterol, campesterol and β-sitosterol decreased secretion of apo B48 in Caco2 human intestinal cells by 15%, 16% and 19% respectively. Moreover, those three phytosterols led to a 30% reduction in VLDL levels as measured by secretion of apo B100 in HepG2 human liver cells [182].
In a placebo-controlled, crossover feeding trial, phytosterol intakes at moderate (458 mg/day) and high (2059 mg/day) doses significantly increased total faecal cholesterol excretion and biliary cholesterol excretion and decreased intestinal cholesterol absorption [183].
6.4. Fiber
Meta-analyses of prospective cohort studies showed that total dietary fibre consumption (average of 7 g/day) was inversely associated with the risk of CVD [184,185]. Insoluble fiber in nuts increases satiety and faecal bulk. Soluble fiber (viscous fiber) decreases gastric emptying and impairs diffusion across the unstirred water layer in the small intestine and increases bile acid excretion [186]. These favourable effects of fiber may partly contribute to the cholesterol-lowering effects of nuts.
7. Antioxidant Activity and Decreased Oxidative Stress
Nuts have very different antioxidant capacities varying from 1.2 to 120 mg of Trolox equivalents per 100 g fresh weight [19]. In a systematic review of in vitro, in vivo and human intervention studies, most nuts showed favourable effects on oxidation while walnuts showed inconsistent antioxidant effects [187]. The PREDIMED study showed that adherence to the Mediterranean diets enriched with either an average of 50 mL/day of virgin olive oil or 30 g/day of nuts for 5 years in subjects with metabolic syndrome decreased xanthine oxidase (an enzymatic source of reactive oxygen species (ROS)) activity and increased plasma SOD and catalase activities compared with a low-fat diet [188].
MUFAs which are abundant in nuts resist oxidation [189,190] while PUFAs from walnuts which are potentially very oxidisable due to their double bonds [191] appear not to influence LDL oxidation compared with a walnut-free, lower-PUFA diet [68,192,193,194], indicating that other bioactive compounds in nuts such as polyphenols [195,196,197,198], tocopherols [199], phytosterols (β-sitosterol) [200,201] and selenium [202,203] could be possible candidates for the antioxidative effects, possibly in a synergistic manner [204]. Human interventions evaluating the effect of nut consumption on oxidative stress are shown in Table 4. Thirteen [24,71,77,121,203,205,206,207,208,209,210,211,212] of 14 clinical studies in Table 4 showed a protective effect on oxidative stress. Only one study [213] showed no effect on oxidative stress.
Walnuts have the highest content of polyphenols in both raw and roasted nuts. Walnut polyphenols have the highest antioxidant efficacy compared with other nuts. Walnuts, almonds, Brazil, cashews, hazelnuts, macadamias, peanuts, pecans, pistachios and peanut butter have greater antioxidant power compared with α-tocopherol ranging from 15 times for raw walnuts to 1.9 times for roasted cashews [14].
In in vitro studies, 14 phenolic compounds obtained from walnuts showed SOD-like activity and 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging activity [195].
Walnut extracts consisting of ellagic acid monomers, polymeric ellagitannins and other phenolics, principally nonflavonoid compounds inhibited Cu2+-induced LDL oxidation by 84% while ellagic acid alone at the same level inhibited Cu2+-induced LDL oxidation by 14% [196]. Approximately 120 phenolic compounds have been identified in walnuts [214]. Almond polyphenols and α-tocopherol synergistically inhibited Cu2+-induced LDL oxidation [198,204]. A decrease in Cu2+-induced LDL oxidation was also observed in clinical studies following 4 weeks of almond consumption (60 g/day) in men with mild hyperlipidemia [215] and in Chinese patients with T2DM [216] but not in normocholesterolemic healthy subjects who consumed 66 g/day of almonds for 6 weeks [65].
Walnuts and cashews (63–108 g/day for each nut) significantly improved antioxidant status with a reduction in oxidized glutathione (GSSG) and an increase in oxygen radical absorbance capacity (ORAC) levels but only in comparison with baseline and not the control diet [206]. An acute study examined postprandial oxidative stress compared a walnut meal containing 90 g walnuts and a isocaloric control meal (50% carbohydrate; 20% protein; 30% fat) in healthy young subjects. The walnut meal reduced oxidative stress by significantly increasing iAUC for hydrophilic and lipophilic ORAC and decreasing iAUC for MDA over the course of a 5-h test. There was decreased oxidized LDL at 2 h after a walnut meat. The walnut meal also significantly elevated plasma levels of epicatechin gallate (ECG), epigallocatechin gallate (EGCG) and gallocatechin gallate (GCG) 1 h after a walnut meal. A significant amount of urolithin-A in urine samples was detected after the walnut meal. A diet effect for plasma γ-tocopherol but not for α-tocopherol was observed in the mixed linear models [211].
An acute study showed that the glucose and insulin-lowering effects of almonds were also related to a lower risk of oxidative damage to proteins [32]. An almond meal improved postprandial protein damage by increasing serum protein thiol (-SH) levels compared with control meals (bread, potato and rice meals containing similar amounts of carbohydrate, protein and fat) when data from three control meals were combined. There were negative correlations between postprandial changes in protein thiols and 2-h glucose iAUC or 2-h insulin iAUC or glucose peak height or insulin peak height, indicating that blood glucose levels increase as damage to protein thiols increases [32].
Four-week pecan [212] and 3-week pistachios [71] consumption in healthy individuals also showed decreased MDA concentrations. Pistachio diets (32–63 or 63–126 g/day) consumed by hypercholesterolemic subjects for 4 weeks significantly reduced serum oxidized-LDL levels compared with a low-fat control diet and elevated plasma lutein, α-carotene, and β-carotene levels compared with baseline [210]. Daily supplementation of two Brazil nuts added to a normal diet for 12 weeks increased plasma selenium concentrations and GPx activities compared with the normal diet [203]. In addition, the consumption of pistachios (65–75 g/day) for 3 weeks and hazelnuts (1 g/kg body weight/day) for 30 days in healthy subjects [71,77] and macadamias (40–90 g/day) for 4 weeks in hypercholesterolemic subjects [207] enhanced oxidative status but these studies had the limitation that the amount of SFA in the nut diet was lower than that in a control diet. Nuts also reduce oxidative DNA damage [121,205,208].
However, whether the antioxidant effects of nuts play a role in their apparent effects in CVD prevention is not clear and more clinical trials of nuts with hard endpoints are required.
8. Anti-Inflammatory Actions
Elevated levels of pro-inflammatory markers (e.g., hs-CRP, IL-6 and TNF-α) are strong predictors of the development of T2DM [217,218] and CVD [219]. In the Nurses’ Health Study (NHS) and the Health Professionals Follow-Up Study (HPFS) of US men and women with no diabetes, nut consumption (≥5 times per week, 28 g per serving) was associated with lower inflammatory markers of hs-CRP (relative concentrations 0.80 (95% CI: 0.69, 0.90); p = 0.0003) and IL-6 (0.86 (95% CI: 0.77, 0.97); p = 0.006) after adjusting for demographic, medical, dietary, and lifestyle variables, compared with no or almost no nut consumption [220]. A sub-study of the PREDIMED study (two Mediterranean diets enriched with either 50 mL/day of virgin olive oil or 30 g/day of nuts and a control lower fat Mediterranean diet) investigated methylation changes in inflammation-related genes in peripheral blood cells between baseline and 5 years in 36 subjects at high cardiovascular risks. This study showed a correlation between methylation changes of eight genes (EEF2, COL18A1, IL4I1, LEPR, PLAGL1, IFRD1, MAPKAPK2, and PPARGC1B) related to inflammation and adherence to a Mediterranean diet. A positive correlation between levels of EEF2 methylation and TNF-α and hs-CRP was observed, suggesting that Mediterranean diets might have anti-inflammatory actions possibly via epigenetic mechanisms [221]. Human interventions evaluating the effect of nut consumption on inflammation are shown in Table 5. Most clinical studies in Table 5 showed beneficial effects on inflammatory markers. Only one pistachio study showed no change in inflammatory markers [22].
In a pre–post intervention study [226], a diet supplemented with raw almond (20% of total energy substituted for fat and carbohydrate) and physical activity (45 min of walking at least 5 days a week) for 24 weeks in subjects with T2DM showed a significant decrease in hs-CRP (p < 0.01), along with a reduction in waist circumference, waist-to-height ratio, serum TC, TG, LDL-C, and HbA1c compared with a baseline 3-week almond-free diet and same level of physical activity [226].
Zhao et al. [222,223] conducted a randomized, controlled cross-over study in 23 hypercholesterolemic subjects who were randomly assigned to one of three diets for 6 weeks each: an average American diet ((AAD)—13% energy from SFA, 13% energy from MUFAs and 8.7% energy from PUFAs (7.7% LA; 0.8% ALA)); two diets high in PUFAs—a LA diet and ALA diet. The LA diet consisted of 16.4% energy from PUFAs (12.6% LA; 3.6% ALA). The ALA diet consisted of 17% energy from PUFAs (10.5% LA; 6.5% ALA). Walnuts and walnut oil were used for half of total fat in the 2 high-PUFA diets. The ALA diet reduced hs-CRP (p < 0.01) while the LA diet had a trend to decrease hs-CRP (p = 0.08) compared with the AAD [222]. The levels of IL-6, IL-1β and TNF-α cultured in peripheral blood mononuclear cells (PBMCs) was significantly lower on the ALA diet than on the LA diet (p < 0.05) [223]. In addition, the ALA diet decreased VCAM-1 by 16% while the LA diet decreased it by only 3% (p < 0.01). This study indicates that ALA and EPA derived from ALA exert a key role in the anti-inflammatory responses after walnut consumption [222] but a large proportion of dietary ALA is oxidized after digestion and only a small proportion is converted to EPA/DHA [227].
Other bioactive compounds in nuts may influence anti-inflammatory activity. Ellagic acid, a major polyphenolic compound in especially walnuts, has shown anti-inflammatory activity [228,229]. Nuts are a good source of magnesium which is inversely associated with the risk of CVD [230,231] and hs-CRP [232,233], and E-selectin levels [233].
Hernandez-Alonso et al. [225] showed an anti-inflammatory effect of pistachio consumption (pistachios are higher in β-carotene, γ-tocopherol and lutein than other nuts) compared with a control diet with olive oil to compensate for the energy from pistachios) [225]. Another pistachio diet (57 g/day) for 4 months significantly decreased IL-6 mRNA (p = 0.004) and resistin gene expression (p = 0.04) in lymphocytes compared with the control diet [225]. Elevated serum resistin levels are associated with insulin resistance, T2DM, and CVD [234,235,236].
Luo et al. [237] demonstrated the mechanism of resistin in increased glucose production and decreased insulin action in human liver HepG2 cells. Resistin led to insulin resistance through both 5′ adenosine monophosphate-activated protein kinase (AMPK)-dependent and AMPK-independent pathways in hepatocytes: resistin upregulated cytokine signalling 3 (SOCS-3) expression, and downregulated insulin receptor substrate 2 (IRS-2) and Akt phosphorylation via an AMPK-independent mechanism. In addition, resistin downregulated AMPK leading to upregulated mRNA expression of gluconeogenic enzymes, G6Pase and PEPCK, and downregulated glucose transporter 2 (GLUT2) mRNA and glycogen synthesis [237].
9. Endothelial Function
Oxidative stress (increased F2-isoprostanes and oxidized LDL), inflammation (hs-CRP) and endothelial dysfunction (e.g., increased E-selectin and intercellular cell-adhesion molecule-1 (ICAM-1)) are positively associated with the risk of T2DM [238,239].
Endothelial dysfunction which is characterized by decreased nitric oxide (NO) bioavailability (either decreased NO synthesis or destruction of NO by reactive oxygen species such as superoxide) [240], is associated with obesity and insulin resistance and is involved in initiation and promotion of atherosclerosis [241] and is related to the risk of CVD [242,243].
Insulin can induce both vasodilation and vasoconstriction in the endothelium. It has been postulated that insulin binding to the insulin receptor (IR) activates IR substrate (IRS) proteins leading to activation of phosphoinositide 3-kinase (PI3K)/protein kinase B (PKB)/endothelial NO synthase (eNOS) at the Ser1177-P pathway [244,245,246,247,248]. Activated eNOS synthesizes NO and l-citrulline from l-arginine and molecular oxygen. NO then stimulates cyclic GMP to induce smooth muscle relaxation.
The vasoconstrictive actions of insulin are mediated by endothelin-1 (ET1). ET-1 is a potent endothelial activator which participates in plaque development and smooth muscle proliferation via the mitogen-activated protein kinase (MAPK)/ERK activation and cyclin D1 pathways [249]) through MAPK/ERK, which antagonizes insulin-induced, PI3K-dependent vasodilatation in skeletal muscle arterioles [250]. NO donors can inhibit cytokine-induced endothelial cell activation and monocyte adhesion partly by inhibiting NF-κB [251]. Ellagic acid (50 μmol/L) significantly blocked the expression of VCAM-1 and E-selectin by inhibiting IL-1 β-induced activation of NF-kB p65 and p50, leading to reduced monocyte adhesion in HUVEC [228].
Elevated inflammatory cytokines promote endothelial dysfunction [241]. The inflammatory cytokine TNF-α has been shown to decrease NO bioavailability in the vascular endothelium [252]. TNF-α is also known to increase the expression of cell adhesion molecules such as VCAM-1, ICAM-1 and E-selectin [253]. TNF-α-induced expressions of VCAM-1 and ICAM-1 were significantly inhibited in human aorta endothelial cells (HAEC) by walnut extract (10–200 μg/mL) or ellagic acid (0.1 and 1 μM) [229].
Unsaturated fatty acids, especially oleic acid in nuts, appear to prevent atherosclerosis by modulating gene expression of endothelial leukocyte adhesion molecules. In HUVECs, oleic acid suppressed the expression of VCAM-1 stimulated with cytokines (e.g., TNF-α, IL-1α, IL-1β and IL-4) or lipopolysaccharide (LPS) or protein-kinase C activator (PMA) and oleic acid also inhibited LPS-stimulated NF-kB activation. Moreover, oleic acid decreased SFAs (palmitic and stearic acids) and increased oleic acid with no change PUFAs in the cell membrane phospholipids which influences prostaglandin production [254].
Adiponectin activates AMPK and PKB signalling resulting in NO generation in endothelial cells [255,256], and suppresses the expression of VCAM-1, ICAM-1 and E-selectin [257]. Walnut consumption (48 g/day) for 4 days increased circulating apo AI and total adiponectin levels in obese subjects with the metabolic syndrome, indicating that even short-term walnut consumption benefits the lipid profile and adiponectin levels which may insulin resistance and CVD [258].
Nut consumption may favourably affect endothelial function. Improved endothelial function was observed after consumption of walnuts in overweight subjects with visceral obesity [259], patients with hypercholesterolemia [192,260], and patients with T2DM [261], as well as after consumption of pistachios in subjects with mild dyslipidemia [262].
In a randomized crossover study of hypercholesterolemic subjects comparing a cholesterol-lowering Mediterranean diet and a walnut diet containing similar energy and fat substituting walnuts for 32% of the energy from MUFA in the Mediterranean diet, walnuts significantly improved endothelium-dependent vasodilation and decreased VCAM-1, TC and LDL-C. Decreased cholesterol correlated with increases in dietary α-linolenic acid and LDL γ-tocopherol levels. Changes in endothelium-dependent vasodilation was inversely correlated with those in the TC/HDL-C ratios suggesting the favourable effect of walnut on endothelial function could be explained in part by improved lipid profiles [192].
A whole-walnut diet (whole walnuts: 61–150 g/kg diet) significantly decreased aortic ET-1 mRNA levels (by up to 75%) and the aortic cholesterol ester compared with the lowest α-tocopherol diet groups (α-tocopherol: 8.1–81 mg/kg diet) in hamsters fed a high-fat atherogenic diet [263].
Other nut bioactive such as l-arginine [264] and magnesium [265,266] are possible candidates to account for improved endothelial function [254].
In a cross-sectional study, subjects who consumed more than 7.5 g/day of arginine had a 30% lower likelihood of having an hs-CRP above 3.0 mg/L compared with those who consumed 2.5–5.0 g/day of arginine [267]. Intervention studies [268,269] also showed that dietary l-arginine improved endothelial function and decreased monocyte adhesion to endothelial cells in young men with coronary artery disease [269] and decreased P-selectin, ICAM-1, E-selectin, IL-1 and IL-6 levels in subjects with intractable angina pectoris [268]. Similarly, daily supplementation of 10 g of arginine for 4 weeks improved endothelial function and decreased LDL-C oxidation in subjects with stable coronary artery disease without significant changes in inflammatory markers [270].
10. Blood Pressure
A meta-analysis of RCTs [271] showed that nut consumption reduced systolic blood pressure (SBP) by 1.3 mm Hg (p = 0.02) and diastolic blood pressure (DBP) in subjects without T2DM. Only pistachios and mixed nuts showed a significant effect on BP [271].
Mediterranean diets enriched with either 350 mL/week of extra virgin olive oil or 30 g/day of nuts lowered BP after a 4-year follow-up period [272] and 24-h ambulatory BP after a 1-year follow-up period [273]. Polyphenol consumption may have been responsible for the hypotensive effects [274]. The reduction in BP in 200 participants was positively associated with an increase in total polyphenol excretion and plasma NO markers and a positive correlation between urinary total polyphenol excretion and plasma NO was observed. These findings suggest that polyphenols could also exert a protective effect on endothelial function [274].
11. Conclusions
In conclusion, frequent nut consumption could play a role in reducing the risk of T2DM and CVD through improvement in glucose and lipid metabolism, weight maintenance and improved endothelial function. The protective effect of nuts could be explained by their distinctive nutrient profile and non-nutrient bioactive compounds. However, the specific mechanisms underlying these effects are not fully understood. Further targeted research should be undertaken to clarify the biological mechanisms.
Acknowledgments
Peter Clifton is supported by a NHMRC Principal Research Fellowship.
Author Contributions
All authors conceived of the manuscript structure and contributed to the writing and editing.
Conflicts of Interest
The authors have no conflicts of interest related to this manuscript.
References
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Sherwin, R.; Anderson, R.; Buse, J.; Chin, M.; Eddy, D.; Fradkin, J.; Ganiats, T.; Ginsberg, H.; Kahn, R.; Nwankwo, R. The prevention or delay of type 2 diabetes. Diabetes Care 2003, 26, S62. [Google Scholar] [PubMed]
- Nanditha, A.; Ma, R.C.; Ramachandran, A.; Snehalatha, C.; Chan, J.C.; Chia, K.S.; Shaw, J.E.; Zimmet, P.Z. Diabetes in asia and the pacific: Implications for the global epidemic. Diabetes Care 2016, 39, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.S.; Dagenais, G.R.; Mohan, V.; Diaz, R.; Probstfield, J.; Freeman, R.; Shaw, J.; Lanas, F.; Avezum, A.; Budaj, A.; et al. Glucose levels are associated with cardiovascular disease and death in an international cohort of normal glycaemic and dysglycaemic men and women: The epidream cohort study. Eur. J. Prev. Cardiol. 2012, 19, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Kirpichnikov, D.; Sowers, J.R. Diabetes mellitus and diabetes-associated vascular disease. Trends Endocrinol. Metab. 2001, 12, 225–230. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, X.; Brown, J.; Vistisen, D.; Sicree, R.; Shaw, J.; Nichols, G. Global healthcare expenditure on diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Pfeiffer, A.F. Foods for the prevention of diabetes: How do they work? Diabetes Metab. Res. Rev. 2012, 28, 25–49. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Zhang, Y.; Ding, Y.; Shan, Z.; Chen, S.; Yu, M.; Hu, F.B.; Liu, L. Nut consumption and risk of type 2 diabetes, cardiovascular disease, and all-cause mortality: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2014, 100, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Del Gobbo, L.C.; Falk, M.C.; Feldman, R.; Lewis, K.; Mozaffarian, D. Effects of tree nuts on blood lipids, apolipoproteins, and blood pressure: Systematic review, meta-analysis, and dose-response of 61 controlled intervention trials. Am. J. Clin. Nutr. 2015, 102, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Jaceldo-Siegl, K.; Haddad, E.; Oda, K.; Fraser, G.E.; Sabate, J. Tree nuts are inversely associated with metabolic syndrome and obesity: The adventist health study-2. PLoS ONE 2014, 9, e85133. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, A.J.; de Souza, R.J.; Meyre, D.; Anand, S.S.; Mente, A. A systematic review and meta-analysis of nut consumption and incident risk of cvd and all-cause mortality. Br. J. Nutr. 2016, 115, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Yang, J.; Marventano, S.; Micek, A.; Galvano, F.; Kales, S.N. Nut consumption on all-cause, cardiovascular, and cancer mortality risk: A systematic review and meta-analysis of epidemiologic studies. Am. J. Clin. Nutr. 2015, 101, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, M.; Sathe, S.K. Chemical composition of selected edible nut seeds. J. Agric. Food Chem. 2006, 54, 4705–4714. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.A.; Cai, Y. Nuts, especially walnuts, have both antioxidant quantity and efficacy and exhibit significant potential health benefits. Food Funct. 2012, 3, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Koo, E.; Neuringer, M.; SanGiovanni, J.P. Macular xanthophylls, lipoprotein-related genes, and age-related macular degeneration. Am. J. Clin. Nutr. 2014, 100 (Suppl. 1), 336S–346S. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture Agricultural Research Service USDA Food Composition Databases. Available online: https://ndb.nal.usda.gov/ndb/search/list (accessed on 10 March 2017).
- Phenol Explorer Database on Polyphenol Content in Foods. Available online: http://www.phenol-explorer.eu (accessed on 24 May 2017).
- Yang, J. Brazil nuts and associated health benefits: A review. Food Sci. Technol. 2009, 42, 1573–1580. [Google Scholar] [CrossRef]
- Abe, L.T.; Lajolo, F.M.; Genovese, M.I. Comparison of phenol content and antioxidant capacity of nuts. Food Sci. Technol. 2010, 30, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Viguiliouk, E.; Kendall, C.W.; Blanco Mejia, S.; Cozma, A.I.; Ha, V.; Mirrahimi, A.; Jayalath, V.H.; Augustin, L.S.; Chiavaroli, L.; Leiter, L.A.; et al. Effect of tree nuts on glycemic control in diabetes: A systematic review and meta-analysis of randomized controlled dietary trials. PLoS ONE 2014, 9, e103376. [Google Scholar] [CrossRef] [PubMed]
- Zibaeenezhad, M.; Aghasadeghi, K.; Hakimi, H.; Yarmohammadi, H.; Nikaein, F. The effect of walnut oil consumption on blood sugar in patients with diabetes mellitus type 2. Int. J. Endocrinol. Metab. 2016, 14, e34889. [Google Scholar] [PubMed]
- Sauder, K.A.; McCrea, C.E.; Ulbrecht, J.S.; Kris-Etherton, P.M.; West, S.G. Effects of pistachios on the lipid/lipoprotein profile, glycemic control, inflammation, and endothelial function in type 2 diabetes: A randomized trial. Metabolism 2015, 64, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Parham, M.; Heidari, S.; Khorramirad, A.; Hozoori, M.; Hosseinzadeh, F.; Bakhtyari, L.; Vafaeimanesh, J. Effects of pistachio nut supplementation on blood glucose in patients with type 2 diabetes: A randomized crossover trial. Rev. Diabet. Stud. 2014, 11, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Sari, I.; Baltaci, Y.; Bagci, C.; Davutoglu, V.; Erel, O.; Celik, H.; Ozer, O.; Aksoy, N.; Aksoy, M. Effect of pistachio diet on lipid parameters, endothelial function, inflammation, and oxidative status: A prospective study. Nutrition 2010, 26, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.W.; Balasubramanyam, A.; Kimball, K.T.; Aherns, A.K.; Fordis, C.M., Jr.; Ballantyne, C.M. Long-term, randomized clinical trial of two diets in the metabolic syndrome and type 2 diabetes. Diabetes Care 2003, 26, 2481–2482. [Google Scholar] [CrossRef] [PubMed]
- Wien, M.A.; Sabate, J.M.; Ikle, D.N.; Cole, S.E.; Kandeel, F.R. Almonds vs. complex carbohydrates in a weight reduction program. Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study Obes. 2003, 27, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Kendall, C.W.; Josse, A.R.; Esfahani, A.; Jenkins, D.J. The impact of pistachio intake alone or in combination with high-carbohydrate foods on post-prandial glycemia. Eur. J. Clin. Nutr. 2011, 65, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Kendall, C.W.; West, S.G.; Augustin, L.S.; Esfahani, A.; Vidgen, E.; Bashyam, B.; Sauder, K.A.; Campbell, J.; Chiavaroli, L.; Jenkins, A.L.; et al. Acute effects of pistachio consumption on glucose and insulin, satiety hormones and endothelial function in the metabolic syndrome. Eur. J. Clin. Nutr. 2014, 68, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Reis, C.E.; Ribeiro, D.N.; Costa, N.M.; Bressan, J.; Alfenas, R.C.; Mattes, R.D. Acute and second-meal effects of peanuts on glycaemic response and appetite in obese women with high type 2 diabetes risk: A randomised cross-over clinical trial. Br. J. Nutr. 2013, 109, 2015–2023. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.S.; Buller, A.J. Vinegar and peanut products as complementary foods to reduce postprandial glycemia. J. Am. Diet. Assoc. 2005, 105, 1939–1942. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.M.; Considine, R.V.; Mattes, R.D. Acute and second-meal effects of almond form in impaired glucose tolerant adults: A randomized crossover trial. Nutr. Metab. 2011, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Kendall, C.W.; Josse, A.R.; Salvatore, S.; Brighenti, F.; Augustin, L.S.; Ellis, P.R.; Vidgen, E.; Rao, A.V. Almonds decrease postprandial glycemia, insulinemia, and oxidative damage in healthy individuals. J. Nutr. 2006, 136, 2987–2992. [Google Scholar] [PubMed]
- Josse, A.R.; Kendall, C.W.; Augustin, L.S.; Ellis, P.R.; Jenkins, D.J. Almonds and postprandial glycemia—A dose-response study. Metabolism 2007, 56, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Crouch, M.A.; Slater, R.T., III. Almond “appetizer” effect on glucose tolerance test (GTT) results. J. Am. Board Fam. Med. 2016, 29, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Kendall, C.W.; Esfahani, A.; Josse, A.R.; Augustin, L.S.; Vidgen, E.; Jenkins, D.J. The glycemic effect of nut-enriched meals in healthy and diabetic subjects. Nutr. Metab. Cardiovasc. Dis. 2011, 21 (Suppl. 1), S34–S39. [Google Scholar] [CrossRef] [PubMed]
- Wien, M.; Bleich, D.; Raghuwanshi, M.; Gould-Forgerite, S.; Gomes, J.; Monahan-Couch, L.; Oda, K. Almond consumption and cardiovascular risk factors in adults with prediabetes. J. Am. Coll. Nutr. 2010, 29, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, J.C.; Most, M.M.; Lefevre, M.; Greenway, F.L.; Rood, J.C. Effect of diets enriched in almonds on insulin action and serum lipids in adults with normal glucose tolerance or type 2 diabetes. Am. J. Clin. Nutr. 2002, 76, 1000–1006. [Google Scholar] [PubMed]
- Cohen, A.E.; Johnston, C.S. Almond ingestion at mealtime reduces postprandial glycemia and chronic ingestion reduces hemoglobin A(1c) in individuals with well-controlled type 2 diabetes mellitus. Metabolism 2011, 60, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Imamura, F.; Micha, R.; Wu, J.H.; de Oliveira Otto, M.C.; Otite, F.O.; Abioye, A.I.; Mozaffarian, D. Effects of saturated fat, polyunsaturated fat, monounsaturated fat, and carbohydrate on glucose-insulin homeostasis: A systematic review and meta-analysis of randomised controlled feeding trials. PLoS Med. 2016, 13, e1002087. [Google Scholar] [CrossRef] [PubMed]
- Wendland, E.; Farmer, A.; Glasziou, P.; Neil, A. Effect of α linolenic acid on cardiovascular risk markers: A systematic review. Heart 2006, 92, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, T.; Yatsuya, H.; Toyoshima, H.; Sasaki, S.; Li, Y.; Otsuka, R.; Wada, K.; Hotta, Y.; Mitsuhashi, H.; Matsushita, K.; et al. Higher dietary intake of alpha-linolenic acid is associated with lower insulin resistance in middle-aged japanese. Prev. Med. 2010, 50, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Swaminath, G.; Cao, Q.; Yang, L.; Guo, Q.; Salomonis, H.; Lu, J.; Houze, J.B.; Dransfield, P.J.; Wang, Y.; et al. Activation of FFA1 mediates GLP-1 secretion in mice. Evidence for allosterism at FFA1. Mol. Cell. Endocrinol. 2013, 369, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Tuo, Y.; Feng, D.D.; Wang, D.F.; Sun, J.; Li, S.B.; Chen, C. Long-term in vitro treatment of INS-1 rat pancreatic beta-cells by unsaturated free fatty acids protects cells against gluco- and lipotoxicities via activation of GPR40 receptors. Clin. Exp. Pharmacol. Physiol. 2012, 39, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.L.; Shu, G.; Zhang, Z.Q.; Wang, S.B.; Zhu, X.T.; Gao, P.; Xi, Q.Y.; Zhang, Y.L.; Jiang, Q.Y. Roles of alpha-linolenic acid on IGF-I secretion and GH/IGF system gene expression in porcine primary hepatocytes. Mo. Biol. Rep. 2012, 39, 10987–10996. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, D.R. The relative roles of growth hormone and IGF-1 in controlling insulin sensitivity. J. Clin. Investig. 2004, 113, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Ortega, F.J.; Cardona-Alvarado, M.I.; Mercader, J.M.; Moreno-Navarrete, J.M.; Moreno, M.; Sabater, M.; Fuentes-Batllevell, N.; Ramirez-Chavez, E.; Ricart, W.; Molina-Torres, J.; et al. Circulating profiling reveals the effect of a polyunsaturated fatty acid-enriched diet on common micrornas. J. Nutr. Biochem. 2015, 26, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Alonso, P.; Giardina, S.; Salas-Salvado, J.; Arcelin, P.; Bullo, M. Chronic pistachio intake modulates circulating micrornas related to glucose metabolism and insulin resistance in prediabetic subjects. Eur. J. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of micrornas in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Alper, C.M.; Mattes, R.D. Effects of chronic peanut consumption on energy balance and hedonics. Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study Obes. 2002, 26, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Fraser, G.E.; Bennett, H.W.; Jaceldo, K.B.; Sabate, J. Effect on body weight of a free 76 kilojoule (320 calorie) daily supplement of almonds for six months. J. Am. Coll. Nutr. 2002, 21, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Hollis, J.; Mattes, R. Effect of chronic consumption of almonds on body weight in healthy humans. Br. J. Nutr. 2007, 98, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Almario, R.U.; Vonghavaravat, V.; Wong, R.; Kasim-Karakas, S.E. Effects of walnut consumption on plasma fatty acids and lipoproteins in combined hyperlipidemia. Am. J. Clin. Nutr. 2001, 74, 72–79. [Google Scholar] [PubMed]
- Abbey, M.; Noakes, M.; Belling, G.B.; Nestel, P.J. Partial replacement of saturated fatty acids with almonds or walnuts lowers total plasma cholesterol and low-density-lipoprotein cholesterol. Am. J. Clin. Nutr. 1994, 59, 995–999. [Google Scholar] [PubMed]
- Coelho, S.B.; de Sales, R.L.; Iyer, S.S.; Bressan, J.; Costa, N.M.; Lokko, P.; Mattes, R. Effects of peanut oil load on energy expenditure, body composition, lipid profile, and appetite in lean and overweight adults. Nutrition 2006, 22, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B.; Stampfer, M.J.; Manson, J.E.; Rimm, E.B.; Colditz, G.A.; Rosner, B.A.; Speizer, F.E.; Hennekens, C.H.; Willett, W.C. Frequent nut consumption and risk of coronary heart disease in women: Prospective cohort study. BMJ 1998, 317, 1341–1345. [Google Scholar] [CrossRef] [PubMed]
- Albert, C.M.; Gaziano, J.M.; Willett, W.C.; Manson, J.E. Nut consumption and decreased risk of sudden cardiac death in the physicians’ health study. Arch. Intern. Med. 2002, 162, 1382–1387. [Google Scholar] [CrossRef] [PubMed]
- Fraser, G.E.; Sabate, J.; Beeson, W.L.; Strahan, T.M. A possible protective effect of nut consumption on risk of coronary heart disease. The adventist health study. Arch. Intern. Med. 1992, 152, 1416–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellsworth, J.L.; Kushi, L.H.; Folsom, A.R. Frequent nut intake and risk of death from coronary heart disease and all causes in postmenopausal women: The iowa women’s health study. Nutr. Metab. Cardiovasc. Dis. 2001, 11, 372–377. [Google Scholar] [PubMed]
- Bes-Rastrollo, M.; Wedick, N.M.; Martinez-Gonzalez, M.A.; Li, T.Y.; Sampson, L.; Hu, F.B. Prospective study of nut consumption, long-term weight change, and obesity risk in women. Am. J. Clin. Nutr. 2009, 89, 1913–1919. [Google Scholar] [CrossRef] [PubMed]
- Bes-Rastrollo, M.; Sabate, J.; Gomez-Gracia, E.; Alonso, A.; Martinez, J.A.; Martinez-Gonzalez, M.A. Nut consumption and weight gain in a Mediterranean cohort: The sun study. Obesity 2007, 15, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Casas-Agustench, P.; Bullo, M.; Ros, E.; Basora, J.; Salas-Salvado, J. Cross-sectional association of nut intake with adiposity in a Mediterranean population. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Kendall, C.W.; Marchie, A.; Parker, T.L.; Connelly, P.W.; Qian, W.; Haight, J.S.; Faulkner, D.; Vidgen, E.; Lapsley, K.G.; et al. Dose response of almonds on coronary heart disease risk factors: Blood lipids, oxidized low-density lipoproteins, lipoprotein(a), homocysteine, and pulmonary nitric oxide: A randomized, controlled, crossover trial. Circulation 2002, 106, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Spiller, G.A.; Jenkins, D.A.; Bosello, O.; Gates, J.E.; Cragen, L.N.; Bruce, B. Nuts and plasma lipids: An almond-based diet lowers LDL-C while preserving HDL-C. J. Am. Coll. Nutr. 1998, 17, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Spiller, G.A.; Jenkins, D.J.; Cragen, L.N.; Gates, J.E.; Bosello, O.; Berra, K.; Rudd, C.; Stevenson, M.; Superko, R. Effect of a diet high in monounsaturated fat from almonds on plasma cholesterol and lipoproteins. J. Am. Coll. Nutr. 1992, 11, 126–130. [Google Scholar] [PubMed]
- Hyson, D.A.; Schneeman, B.O.; Davis, P.A. Almonds and almond oil have similar effects on plasma lipids and ldl oxidation in healthy men and women. J. Nutr. 2002, 132, 703–707. [Google Scholar] [PubMed]
- Sabate, J.; Cordero-Macintyre, Z.; Siapco, G.; Torabian, S.; Haddad, E. Does regular walnut consumption lead to weight gain? Br. J. Nutr. 2005, 94, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, A.; Mann, J.; Skeaff, M.; Frampton, C.; Sutherland, W.; Duncan, A.; Tiszavari, S. A diet rich in walnuts favourably influences plasma fatty acid profile in moderately hyperlipidaemic subjects. Eur. J. Clin. Nutr. 1998, 52, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Zambon, D.; Sabate, J.; Munoz, S.; Campero, B.; Casals, E.; Merlos, M.; Laguna, J.C.; Ros, E. Substituting walnuts for monounsaturated fat improves the serum lipid profile of hypercholesterolemic men and women. A randomized crossover trial. Ann. Intern. Med. 2000, 132, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.M.; Horton, K.; Reese, D.; Carey, C.; Walker, K.; Capuzzi, D.M. Effects of walnut consumption as part of a low-fat, low-cholesterol diet on serum cardiovascular risk factors. Int. J. Vitam. Nutr. Res. 2002, 72, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.A.; Clayshulte, B.J. Pecans lower low-density lipoprotein cholesterol in people with normal lipid levels. J. Am. Diet. Assoc. 2000, 100, 312–318. [Google Scholar] [CrossRef]
- Kocyigit, A.; Koylu, A.A.; Keles, H. Effects of pistachio nuts consumption on plasma lipid profile and oxidative status in healthy volunteers. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.; Kwaw, I.; Matud, J.; Kurtz, I. Effect of pistachio nuts on serum lipid levels in patients with moderate hypercholesterolemia. J. Am. Coll. Nutr. 1999, 18, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Z.; Liu, Y.; Lv, X.; Yang, W. Effects of pistachios on body weight in chinese subjects with metabolic syndrome. Nutr. J. 2012, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Holligan, S.D.; West, S.G.; Gebauer, S.K.; Kay, C.D.; Kris-Etherton, P.M. A moderate-fat diet containing pistachios improves emerging markers of cardiometabolic syndrome in healthy adults with elevated ldl levels. Br. J. Nutr. 2014, 112, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Curb, J.D.; Wergowske, G.; Dobbs, J.C.; Abbott, R.D.; Huang, B. Serum lipid effects of a high-monounsaturated fat diet based on macadamia nuts. Arch. Intern. Med. 2000, 160, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, D.; Humphries, J.; Moores, D.; Somerset, S. Effects of a macadamia nut enriched diet on serum lipids and lipoproteins compared to a low fat diet. Food Aust. 1996, 48, 216–222. [Google Scholar]
- Durak, I.; Koksal, I.; Kacmaz, M.; Buyukkocak, S.; Cimen, B.M.; Ozturk, H.S. Hazelnut supplementation enhances plasma antioxidant potential and lowers plasma cholesterol levels. Clin. Chim. Acta 1999, 284, 113–115. [Google Scholar] [CrossRef]
- Razquin, C.; Sanchez-Tainta, A.; Salas-Salvado, J.; Buil-Cosiales, P.; Corella, D.; Fito, M.; Ros, E.; Estruch, R.; Aros, F.; Gomez-Gracia, E.; et al. Dietary energy density and body weight changes after 3 years in the predimed study. Int. J. Food Sci. Nutr. 2017, 68, 865–872. [Google Scholar] [CrossRef] [PubMed]
- McManus, K.; Antinoro, L.; Sacks, F. A randomized controlled trial of a moderate-fat, low-energy diet compared with a low fat, low-energy diet for weight loss in overweight adults. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Pelkman, C.L.; Fishell, V.K.; Maddox, D.H.; Pearson, T.A.; Mauger, D.T.; Kris-Etherton, P.M. Effects of moderate-fat (from monounsaturated fat) and low-fat weight-loss diets on the serum lipid profile in overweight and obese men and women. Am. J. Clin. Nutr. 2004, 79, 204–212. [Google Scholar] [PubMed]
- Tan, S.Y.; Mattes, R.D. Appetitive, dietary and health effects of almonds consumed with meals or as snacks: A randomized, controlled trial. Eur. J. Clin. Nutr. 2013, 67, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Hull, S.; Re, R.; Chambers, L.; Echaniz, A.; Wickham, M.S. A mid-morning snack of almonds generates satiety and appropriate adjustment of subsequent food intake in healthy women. Eur. J. Nutr. 2015, 54, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.E.J.; Bloom, S.R.; Tan, T. Gut Hormones. In Encyclopedia of Food and Health; Benjamin, C., Paul, M.F., Fidel, T., Eds.; Elsevier Ltd.: Oxford, UK, 2016. [Google Scholar]
- Frost, G.; Brynes, A.E.; Ellis, S.; Milton, J.E.; Nematy, M.; Philippou, E. Nutritional influences on gut hormone release. Curr. Opin. Endocrinol. Diabetes 2006, 13, 42–48. [Google Scholar] [CrossRef]
- Pasman, W.J.; Heimerikx, J.; Rubingh, C.M.; van den Berg, R.; O’Shea, M.; Gambelli, L.; Hendriks, H.F.; Einerhand, A.W.; Scott, C.; Keizer, H.G.; et al. The effect of korean pine nut oil on in vitro CCK release, on appetite sensations and on gut hormones in post-menopausal overweight women. Lipids Health Dis. 2008, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Brennan, A.M.; Mantzoros, C.S. Drug insight: The role of leptin in human physiology and pathophysiology—emerging clinical applications. Nat. Clin. Pract. Endocrinol. Metab. 2006, 2, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Mazidi, M.; Rezaie, P.; Ferns, G.A.; Gao, H.K. Impact of different types of tree nut, peanut, and soy nut consumption on serum C-reactive protein (CRP): A systematic review and meta-analysis of randomized controlled clinical trials. Medicine 2016, 95, e5165. [Google Scholar] [CrossRef] [PubMed]
- Cassady, B.A.; Hollis, J.H.; Fulford, A.D.; Considine, R.V.; Mattes, R.D. Mastication of almonds: Effects of lipid bioaccessibility, appetite, and hormone response. Am. J. Clin. Nutr. 2009, 89, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Frecka, J.M.; Hollis, J.H.; Mattes, R.D. Effects of appetite, bmi, food form and flavor on mastication: Almonds as a test food. Eur. J. Clin. Nutr. 2008, 62, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.J.; Ridgen, J.E.; Phang, P.T.; Birmingham, C.L. Influence of dietary fat polyunsaturated to saturated ratio on energy substrate utilization in obesity. Metabolism 1992, 41, 396–401. [Google Scholar] [CrossRef]
- Babio, N.; Toledo, E.; Estruch, R.; Ros, E.; Martinez-Gonzalez, M.A.; Castaner, O.; Bullo, M.; Corella, D.; Aros, F.; Gomez-Gracia, E.; et al. Mediterranean diets and metabolic syndrome status in the predimed randomized trial. Can. Med. Assoc. J. 2014, 186, E649–E657. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garcia, E.; Carvajal-Lerida, I.; Perez-Galvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Ellis, P.R.; Kendall, C.W.; Ren, Y.; Parker, C.; Pacy, J.F.; Waldron, K.W.; Jenkins, D.J. Role of cell walls in the bioaccessibility of lipids in almond seeds. Am. J. Clin. Nutr. 2004, 80, 604–613. [Google Scholar] [PubMed]
- Mandalari, G.; Grundy, M.M.-L.; Grassby, T.; Parker, M.L.; Cross, K.L.; Chessa, S.; Bisignano, C.; Barreca, D.; Bellocco, E.; Laganà, G.; et al. The effects of processing and mastication on almond lipid bioaccessibility using novel methods of in vitro digestion modelling and micro-structural analysis. Br. J. Nutr. 2014, 112, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Kendall, C.; Jenkins, D.; Marchie, A.; Ren, Y.; Ellis, P.; Lapsley, K. Energy Availability from Almonds: Implications for Weight Loss and Cardiovascular Health. A Randomized Controlled Dose-Response Trial. FASEB J. 2003, 17, A339. [Google Scholar]
- Haddad, E.; Sabate, J. Effect of Pecan Consumption on Stool Fat. FASEB J. 2000, 14, A294. [Google Scholar]
- Levine, A.S.; Silvis, S.E. Absorption of whole peanuts, peanut oil, and peanut butter. N. Engl. J. Med. 1980, 303, 917–918. [Google Scholar] [CrossRef] [PubMed]
- Traoret, C.J.; Lokko, P.; Cruz, A.C.; Oliveira, C.G.; Costa, N.M.; Bressan, J.; Alfenas, R.C.; Mattes, R.D. Peanut digestion and energy balance. Int. J. Obes. 2008, 32, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Sethi, S.; Gibney, M.; Williams, C.M. Postprandial lipoprotein metabolism. Nutr. Res. Rev. 1993, 6, 161–183. [Google Scholar] [CrossRef] [PubMed]
- Karpe, F. Postprandial lipid metabolism in relation to coronary heart disease. Proc. Nutr. Soc. 1997, 56, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.-H.; Bassenge, E.; Kim, K.-B.; Kim, Y.-N.; Kim, K.-S.; Lee, H.-J.; Moon, K.-C.; Lee, M.-S.; Park, K.-Y.; Schwemmer, M. Postprandial hypertriglyceridemia impairs endothelial function by enhanced oxidant stress. Atherosclerosis 2001, 155, 517–523. [Google Scholar] [CrossRef]
- Berry, S.E.; Tydeman, E.A.; Lewis, H.B.; Phalora, R.; Rosborough, J.; Picout, D.R.; Ellis, P.R. Manipulation of lipid bioaccessibility of almond seeds influences postprandial lipemia in healthy human subjects. Am. J. Clin. Nutr. 2008, 88, 922–929. [Google Scholar] [PubMed]
- Kim, Y.; Keogh, J.B.; Clifton, P.M. Polyphenols and glycemic control. Nutrients 2016, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Perez-Jimenez, J.; Neveu, V.; Vos, F.; Scalbert, A. Identification of the 100 richest dietary sources of polyphenols: An application of the phenol-explorer database. Eur. J. Clin. Nutr. 2010, 64 (Suppl. 3), S112–S120. [Google Scholar] [CrossRef] [PubMed]
- Bittner, K.; Rzeppa, S.; Humpf, H.U. Distribution and quantification of flavan-3-ols and procyanidins with low degree of polymerization in nuts, cereals, and legumes. J. Agric. Food Chem. 2013, 61, 9148–9154. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Forouhi, N.G.; Sharp, S.J.; Gonzalez, C.A.; Buijsse, B.; Guevara, M.; van der Schouw, Y.T.; Amiano, P.; Boeing, H.; Bredsdorff, L.; et al. Dietary intakes of individual flavanols and flavonols are inversely associated with incident type 2 diabetes in european populations. J. Nutr. 2014, 144, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Forouhi, N.G.; Sharp, S.J.; Gonzalez, C.A.; Buijsse, B.; Guevara, M.; van der Schouw, Y.T.; Amiano, P.; Boeing, H.; Bredsdorff, L.; et al. The association between dietary flavonoid and lignan intakes and incident type 2 diabetes in european populations: The epic-interact study. Diabetes Care 2013, 36, 3961–3970. [Google Scholar] [CrossRef] [PubMed]
- Yamakoshi, J.; Tokutake, S.; Kikuchi, M.; Kubota, Y.; Konishi, H.; Mitsuoka, T. Effect of proanthocyanidin-rich extract from grape seeds on human fecal flora and fecal odor. Microb. Ecol. Health Dis. 2001, 13, 25–31. [Google Scholar] [CrossRef]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P.E. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2011, 93, 62. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Barberan, F.A.; Gonzalez-Sarrias, A.; Garcia-Villalba, R.; Nunez-Sanchez, M.A.; Selma, M.V.; Garcia-Conesa, M.T.; Espin, J.C. Urolithins, the rescue of “old” metabolites to understand a “new” concept: Metabotypes as a nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Barberan, F.A.; Garcia-Villalba, R.; Gonzalez-Sarrias, A.; Selma, M.V.; Espin, J.C. Ellagic acid metabolism by human gut microbiota: Consistent observation of three urolithin phenotypes in intervention trials, independent of food source, age, and health status. J. Agric. Food Chem. 2014, 62, 6535–6538. [Google Scholar] [CrossRef] [PubMed]
- Espin, J.C.; Gonzalez-Barrio, R.; Cerda, B.; Lopez-Bote, C.; Rey, A.I.; Tomas-Barberan, F.A. Iberian pig as a model to clarify obscure points in the bioavailability and metabolism of ellagitannins in humans. J. Agric. Food Chem. 2007, 55, 10476–10485. [Google Scholar] [CrossRef] [PubMed]
- Appeldoorn, M.M.; Vincken, J.P.; Aura, A.M.; Hollman, P.C.; Gruppen, H. Procyanidin dimers are metabolized by human microbiota with 2-(3,4-dihydroxyphenyl)acetic acid and 5-(3,4-dihydroxyphenyl)-gamma-valerolactone as the major metabolites. J. Agric. Food Chem. 2009, 57, 1084–1092. [Google Scholar] [CrossRef] [PubMed]
- Urpi-Sarda, M.; Garrido, I.; Monagas, M.; Gomez-Cordoves, C.; Medina-Remon, A.; Andres-Lacueva, C.; Bartolome, B. Profile of plasma and urine metabolites after the intake of almond [prunus dulcis (Mill.) D.A. Webb] polyphenols in humans. J. Agric. Food Chem. 2009, 57, 10134–10142. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Urpi-Sarda, M.; Sanchez-Patan, F.; Llorach, R.; Garrido, I.; Gomez-Cordoves, C.; Andres-Lacueva, C.; Bartolome, B. Insights into the metabolism and microbial biotransformation of dietary flavan-3-ols and the bioactivity of their metabolites. Food Funct. 2010, 1, 233–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoupi, S.; Williamson, G.; Drynan, J.W.; Barron, D.; Clifford, M.N. A comparison of the in vitro biotransformation of (-)-epicatechin and procyanidin b2 by human faecal microbiota. Mol. Nutr. Food Res. 2010, 54, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Espin, J.C.; Larrosa, M.; Garcia-Conesa, M.T.; Tomas-Barberan, F. Biological significance of urolithins, the gut microbial ellagic acid-derived metabolites: The evidence so far. Evid.-Based Complement. Altern. Med. 2013, 2013, 270418. [Google Scholar] [CrossRef] [PubMed]
- Tulipani, S.; Llorach, R.; Jauregui, O.; Lopez-Uriarte, P.; Garcia-Aloy, M.; Bullo, M.; Salas-Salvado, J.; Andres-Lacueva, C. Metabolomics unveils urinary changes in subjects with metabolic syndrome following 12-week nut consumption. J. Proteome Res. 2011, 10, 5047–5058. [Google Scholar] [CrossRef] [PubMed]
- Tulipani, S.; Urpi-Sarda, M.; Garcia-Villalba, R.; Rabassa, M.; Lopez-Uriarte, P.; Bullo, M.; Jauregui, O.; Tomas-Barberan, F.; Salas-Salvado, J.; Espin, J.C.; et al. Urolithins are the main urinary microbial-derived phenolic metabolites discriminating a moderate consumption of nuts in free-living subjects with diagnosed metabolic syndrome. J. Agric. Food Chem. 2012, 60, 8930–8940. [Google Scholar] [CrossRef] [PubMed]
- Casas-Agustench, P.; López-Uriarte, P.; Bulló, M.; Ros, E.; Cabré-Vila, J.; Salas-Salvadó, J. Effects of one serving of mixed nuts on serum lipids, insulin resistance and inflammatory markers in patients with the metabolic syndrome. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Uriarte, P.; Nogues, R.; Saez, G.; Bullo, M.; Romeu, M.; Masana, L.; Tormos, C.; Casas-Agustench, P.; Salas-Salvado, J. Effect of nut consumption on oxidative stress and the endothelial function in metabolic syndrome. Clin. Nutr. 2010, 29, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Blaiotta, G.; La Gatta, B.; Di Capua, M.; Di Luccia, A.; Coppola, R.; Aponte, M. Effect of chestnut extract and chestnut fiber on viability of potential probiotic lactobacillus strains under gastrointestinal tract conditions. Food Microbiol. 2013, 36, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Nueno-Palop, C.; Bisignano, G.; Wickham, M.S.; Narbad, A. Potential prebiotic properties of almond (Amygdalus communis L.) seeds. Appl. Environ. Microbiol. 2008, 74, 4264–4270. [Google Scholar] [CrossRef] [PubMed]
- Ukhanova, M.; Wang, X.; Baer, D.J.; Novotny, J.A.; Fredborg, M.; Mai, V. Effects of almond and pistachio consumption on gut microbiota composition in a randomised cross-over human feeding study. Br. J. Nutr. 2014, 111, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Lin, X.; Huang, G.; Zhang, W.; Rao, P.; Ni, L. Prebiotic effects of almonds and almond skins on intestinal microbiota in healthy adult humans. Anaerobe 2014, 26, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Samuel, B.S.; Shaito, A.; Motoike, T.; Rey, F.E.; Backhed, F.; Manchester, J.K.; Hammer, R.E.; Williams, S.C.; Crowley, J.; Yanagisawa, M.; et al. Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding g protein-coupled receptor, GPR41. Proc. Natl. Acad. Sci. USA 2008, 105, 16767–16772. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.; Stafford, S.; Coope, G.; Heffron, H.; Real, K.; Newman, R.; Davenport, R.; Barnes, M.; Grosse, J.; Cox, H. Selective FFA2 agonism appears to act via intestinal PYY to reduce transit and food intake but does not improve glucose tolerance in mouse models. Diabetes 2015, 64, 3763–3771. [Google Scholar] [CrossRef] [PubMed]
- Madsbad, S. Exenatide and liraglutide: Different approaches to develop GLP-1 receptor agonists (incretin mimetics)—Preclinical and clinical results. Best Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troy, S.; Soty, M.; Ribeiro, L.; Laval, L.; Migrenne, S.; Fioramonti, X.; Pillot, B.; Fauveau, V.; Aubert, R.; Viollet, B.; et al. Intestinal gluconeogenesis is a key factor for early metabolic changes after gastric bypass but not after gastric lap-band in mice. Cell Metab. 2008, 8, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Mithieux, G.; Misery, P.; Magnan, C.; Pillot, B.; Gautier-Stein, A.; Bernard, C.; Rajas, F.; Zitoun, C. Portal sensing of intestinal gluconeogenesis is a mechanistic link in the diminution of food intake induced by diet protein. Cell Metab. 2005, 2, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Gautier-Stein, A.; Zitoun, C.; Lalli, E.; Mithieux, G.; Rajas, F. Transcriptional regulation of the glucose-6-phosphatase gene by cAMP/vasoactive intestinal peptide in the intestine. Role of HNF4alpha, CREM, HNF1alpha, and C/EbPalpha. J. Biol. Chem. 2006, 281, 31268–31278. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Backhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Usami, M.; Kishimoto, K.; Ohata, A.; Miyoshi, M.; Aoyama, M.; Fueda, Y.; Kotani, J. Butyrate and trichostatin a attenuate nuclear factor kappab activation and tumor necrosis factor alpha secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells. Nutr. Res. 2008, 28, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Park, G.Y.; Joo, M.; Pedchenko, T.; Blackwell, T.S.; Christman, J.W. Regulation of macrophage cyclooxygenase-2 gene expression by modifications of histone H3. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 286, L956–L962. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Lee, E.J.; Lee, J.C.; Kim, W.K.; Kim, H.S. Anti-inflammatory effects of short chain fatty acids in ifn-gamma-stimulated raw 264.7 murine macrophage cells: Involvement of NF-kappab and ERK signaling pathways. Int. Immunopharmacol. 2007, 7, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Boffa, L.C.; Vidali, G.; Mann, R.S.; Allfrey, V.G. Suppression of histone deacetylation in vivo and in vitro by sodium butyrate. J. Biol. Chem. 1978, 253, 3364–3366. [Google Scholar] [PubMed]
- Davie, J.R. Inhibition of histone deacetylase activity by butyrate. J. Nutr. 2003, 133, 2485s–2493s. [Google Scholar] [PubMed]
- Wang, X.; Wei, X.; Pang, Q.; Yi, F. Histone deacetylases and their inhibitors: Molecular mechanisms and therapeutic implications in diabetes mellitus. Acta Pharm. Sin. B 2012, 2, 387–395. [Google Scholar] [CrossRef]
- Christensen, D.P.; Dahllof, M.; Lundh, M.; Rasmussen, D.N.; Nielsen, M.D.; Billestrup, N.; Grunnet, L.G.; Mandrup-Poulsen, T. Histone deacetylase (HDAC) inhibition as a novel treatment for diabetes mellitus. Mol. Med. 2011, 17, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, M.M.; Vasquez, D.S.; Ravnskjaer, K.; Denechaud, P.D.; Yu, R.T.; Alvarez, J.G.; Downes, M.; Evans, R.M.; Montminy, M.; Shaw, R.J. Class IIA histone deacetylases are hormone-activated regulators of foxo and mammalian glucose homeostasis. Cell 2011, 145, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Yamada, T.; Matsumoto, M.; Kido, Y.; Hosooka, T.; Asahara, S.; Matsuda, T.; Ota, T.; Watanabe, H.; Sai, Y.; et al. Endoplasmic reticulum stress inhibits stat3-dependent suppression of hepatic gluconeogenesis via dephosphorylation and deacetylation. Diabetes 2012, 61, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Mosley, A.L.; Ozcan, S. The pancreatic duodenal homeobox-1 protein (PDX-1) interacts with histone deacetylases HDAC-1 and HDAC-2 on low levels of glucose. J. Biol. Chem. 2004, 279, 54241–54247. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Stoffers, D.A.; Nicholls, R.D.; Simmons, R.A. Development of type 2 diabetes following intrauterine growth retardation in rats is associated with progressive epigenetic silencing of PDX1. J. Clin. Investig. 2008, 118, 2316–2324. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory t-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Miyara, M.; Costantino, C.M.; Hafler, D.A. Foxp3+ regulatory t cells in the human immune system. Nat. Rev. Immunol. 2010, 10, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.A.; Jackson, J.; Stanton, M.; Rojas-Triana, A.; Bober, L.; Laverty, M.; Yang, X.; Zhu, F.; Liu, J.; Wang, S. Short-chain fatty acids act as antiinflammatory mediators by regulating prostaglandin E(2) and cytokines. World J. Gastroenterol. 2009, 15, 5549–5557. [Google Scholar] [CrossRef] [PubMed]
- Zapolska-Downar, D.; Siennicka, A.; Kaczmarczyk, M.; Kolodziej, B.; Naruszewicz, M. Butyrate inhibits cytokine-induced VCAM-1 and ICAM-1 expression in cultured endothelial cells: The role of NF-kappab and pparalpha. J. Nutr. Biochem. 2004, 15, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Menzel, T.; Luhrs, H.; Zirlik, S.; Schauber, J.; Kudlich, T.; Gerke, T.; Gostner, A.; Neumann, M.; Melcher, R.; Scheppach, W. Butyrate inhibits leukocyte adhesion to endothelial cells via modulation of VCAM-1. Inflamm. Bowel Dis. 2004, 10, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Hernaez, A.; Castaner, O.; Elosua, R.; Pinto, X.; Estruch, R.; Salas-Salvado, J.; Corella, D.; Aros, F.; Serra-Majem, L.; Fiol, M.; et al. Mediterranean diet improves high-density lipoprotein function in high-cardiovascular-risk individuals: A randomized controlled trial. Circulation 2017, 135, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Musa-Veloso, K.; Paulionis, L.; Poon, T.; Lee, H.Y. The effects of almond consumption on fasting blood lipid levels: A systematic review and meta-analysis of randomised controlled trials. J. Nutr. Sci. 2016, 5, e34. [Google Scholar] [CrossRef] [PubMed]
- Sabate, J.; Oda, K.; Ros, E. Nut consumption and blood lipid levels: A pooled analysis of 25 intervention trials. Arch. Intern. Med. 2010, 170, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Banel, D.K.; Hu, F.B. Effects of walnut consumption on blood lipids and other cardiovascular risk factors: A meta-analysis and systematic review. Am. J. Clin. Nutr. 2009, 90, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, B.; Emberson, J.; Blackwell, L.; Keech, A.; Simes, J.; Barnes, E.H.; Voysey, M.; Gray, A.; Collins, R.; Baigent, C. The effects of lowering LDLcholesterol with statin therapy in people at low risk of vascular disease: Meta-analysis of individual data from 27 randomised trials. Lancet 2012, 380, 581–590. [Google Scholar] [PubMed]
- Griel, A.E.; Kris-Etherton, P.M. Tree nuts and the lipid profile: A review of clinical studies. Br. J. Nutr. 2006, 96 (Suppl. 2), S68–S78. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, S.K.; West, S.G.; Kay, C.D.; Alaupovic, P.; Bagshaw, D.; Kris-Etherton, P.M. Effects of pistachios on cardiovascular disease risk factors and potential mechanisms of action: A dose-response study. Am. J. Clin. Nutr. 2008, 88, 651–659. [Google Scholar] [PubMed]
- Mukuddem-Petersen, J.; Oosthuizen, W.; Jerling, J.C. A systematic review of the effects of nuts on blood lipid profiles in humans. J. Nutr. 2005, 135, 2082–2089. [Google Scholar] [PubMed]
- Ostlund Jr, R.E. Phytosterols and cholesterol metabolism. Curr. Opin. Lipidol. 2004, 15, 37–41. [Google Scholar] [CrossRef]
- Pan, A.; Chen, M.; Chowdhury, R.; Wu, J.H.; Sun, Q.; Campos, H.; Mozaffarian, D.; Hu, F.B. Alpha-linolenic acid and risk of cardiovascular disease: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2012, 96, 1262–1273. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Micha, R.; Wallace, S. Effects on coronary heart disease of increasing polyunsaturated fat in place of saturated fat: A systematic review and meta-analysis of randomized controlled trials. PLoS Med. 2010, 7, e1000252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, D.J. Dietary fatty acids, lipoproteins, and cardiovascular disease. Adv. Food Nutr. Res. 1992, 36, 253–351. [Google Scholar] [PubMed]
- Campos, H.; Dreon, D.M.; Krauss, R.M. Associations of hepatic and lipoprotein lipase activities with changes in dietary composition and low density lipoprotein subclasses. J. Lipid Res. 1995, 36, 462–472. [Google Scholar] [PubMed]
- Brousseau, M.E.; Ordovas, J.M.; Osada, J.; Fasulo, J.; Robins, S.J.; Nicolosi, R.J.; Schaefer, E.J. Dietary monounsaturated and polyunsaturated fatty acids are comparable in their effects on hepatic apolipoprotein mrna abundance and liver lipid concentrations when substituted for saturated fatty acids in cynomolgus monkeys. J. Nutr. 1995, 125, 425–436. [Google Scholar] [PubMed]
- Park, Y.; Carr, T.P. Unsaturated fatty acids and phytosterols regulate cholesterol transporter genes in Caco-2 and HepG 2 cell lines. Nutr. Res. 2013, 33, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Sampath, H.; Ntambi, J.M. Polyunsaturated fatty acid regulation of genes of lipid metabolism. Annu. Rev. Nutr. 2005, 25, 317–340. [Google Scholar] [CrossRef] [PubMed]
- Kritchevsky, D. Protein and atherosclerosis. J. Nutr. Sci. Vitaminol. 1990, 36, S35–S45. [Google Scholar] [CrossRef]
- Huff, M.W.; Carroll, K.K. Effects of dietary protein on turnover, oxidation, and absorption of cholesterol, and on steroid excretion in rabbits. J. Lipid Res. 1980, 21, 546–548. [Google Scholar] [PubMed]
- Ras, R.T.; Geleijnse, J.M.; Trautwein, E.A. LDL-cholesterol-lowering effect of plant sterols and stanols across different dose ranges: A meta-analysis of randomised controlled studies. Br. J. Nutr. 2014, 112, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Calpe-Berdiel, L.; Escola-Gil, J.C.; Blanco-Vaca, F. New insights into the molecular actions of plant sterols and stanols in cholesterol metabolism. Atherosclerosis 2009, 203, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T. The cholesterol-lowering action of plant stanol esters. J. Nutr. 1999, 129, 2109–2112. [Google Scholar] [PubMed]
- Trautwein, E.A.; Duchateau, G.S.; Lin, Y.; Mel’nikov, S.M.; Molhuizen, H.O.; Ntanios, F.Y. Proposed mechanisms of cholesterol-lowering action of plant sterols. Eur. J. Lipid Sci. Technol. 2003, 105, 171–185. [Google Scholar] [CrossRef]
- Yamanashi, Y.; Takada, T.; Suzuki, H. Niemann-pick C1-like 1 overexpression facilitates ezetimibe-sensitive cholesterol and beta-sitosterol uptake in Caco-2 cells. J. Pharmacol. Exp. Ther. 2007, 320, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.R., Jr.; Zhu, L.J.; Hoos, L.M.; Tetzloff, G.; Maguire, M.; Liu, J.; Yao, X.; Iyer, S.P.; Lam, M.H.; Lund, E.G.; et al. Niemann-pick C1 like 1 (NPC1L1) is the intestinal phytosterol and cholesterol transporter and a key modulator of whole-body cholesterol homeostasis. J. Biol. Chem. 2004, 279, 33586–33592. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Yu, L. Opposing gatekeepers of apical sterol transport: Niemann-pick C1-like 1 (NPC1L1) and atp-binding cassette transporters G5 and G8 (ABCG5/ABCG8). Immunol. Endocr. Metab. Agents Med. Chem. 2009, 9, 18–29. [Google Scholar] [CrossRef] [PubMed]
- De Smet, E.; Mensink, R.P.; Plat, J. Effects of plant sterols and stanols on intestinal cholesterol metabolism: Suggested mechanisms from past to present. Mol. Nutr. Food Res. 2012, 56, 1058–1072. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.T.; Wong, W.T.; Guan, L.; Tian, X.Y.; Ma, K.Y.; Huang, Y.; Chen, Z.Y. Effect of phytosterols and their oxidation products on lipoprotein profiles and vascular function in hamster fed a high cholesterol diet. Atherosclerosis 2011, 219, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Brauner, R.; Johannes, C.; Ploessl, F.; Bracher, F.; Lorenz, R.L. Phytosterols reduce cholesterol absorption by inhibition of 27-hydroxycholesterol generation, liver X receptor alpha activation, and expression of the basolateral sterol exporter ATP-binding cassette A1 in Caco-2 enterocytes. J. Nutr. 2012, 142, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Field, F.J.; Born, E.; Mathur, S.N. Effect of micellar beta-sitosterol on cholesterol metabolism in Caco-2 cells. J. Lipid Res. 1997, 38, 348–360. [Google Scholar] [PubMed]
- Ho, S.S.; Pal, S. Margarine phytosterols decrease the secretion of atherogenic lipoproteins from HEPG2 liver and Caco-2 intestinal cells. Atherosclerosis 2005, 182, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Racette, S.B.; Lin, X.; Lefevre, M.; Spearie, C.A.; Most, M.M.; Ma, L.; Ostlund, R.E. Dose effects of dietary phytosterols on cholesterol metabolism: A controlled feeding study. Am. J. Clin. Nutr. 2010, 91, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Threapleton, D.E.; Greenwood, D.C.; Evans, C.E.; Cleghorn, C.L.; Nykjaer, C.; Woodhead, C.; Cade, J.E.; Gale, C.P.; Burley, V.J. Dietary fibre intake and risk of cardiovascular disease: Systematic review and meta-analysis. BMJ 2013, 347, f6879. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Je, Y. Dietary fibre intake and mortality from cardiovascular disease and all cancers: A meta-analysis of prospective cohort studies. Arch. Cardiovasc. Dis. 2016, 109, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Salas-Salvadó, J.; Bulló, M.; Pérez-Heras, A.; Ros, E. Dietary fibre, nuts and cardiovascular diseases. Br. J. Nutr. 2006, 96, S45–S51. [Google Scholar] [CrossRef]
- Lopez-Uriarte, P.; Bullo, M.; Casas-Agustench, P.; Babio, N.; Salas-Salvado, J. Nuts and oxidation: A systematic review. Nutr. Rev. 2009, 67, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Bibiloni, M.D.M.; Martorell, M.; Buil-Cosiales, P.; Marti, A.; Pons, A.; Tur, J.A.; Martinez-Gonzalez, M.Á. Mediterranean diets supplemented with virgin olive oil and nuts enhance plasmatic antioxidant capabilities and decrease xanthine oxidase activity in people with metabolic syndrome: The predimed study. Mol. Nutr. Food Res. 2016, 60, 2654–2664. [Google Scholar] [CrossRef] [PubMed]
- Hargrove, R.L.; Etherton, T.D.; Pearson, T.A.; Harrison, E.H.; Kris-Etherton, P.M. Low fat and high monounsaturated fat diets decrease human low density lipoprotein oxidative susceptibility in vitro. J. Nutr. 2001, 131, 1758–1763. [Google Scholar] [PubMed]
- Berry, E.M.; Eisenberg, S.; Friedlander, Y.; Harats, D.; Kaufmann, N.A.; Norman, Y.; Stein, Y. Effects of diets rich in monounsaturated fatty acids on plasma lipoproteins—The jerusalem nutrition study. II. Monounsaturated fatty acids vs carbohydrates. Am. J. Clin. Nutr. 1992, 56, 394–403. [Google Scholar] [PubMed]
- Bonanome, A.; Pagnan, A.; Biffanti, S.; Opportuno, A.; Sorgato, F.; Dorella, M.; Maiorino, M.; Ursini, F. Effect of dietary monounsaturated and polyunsaturated fatty acids on the susceptibility of plasma low density lipoproteins to oxidative modification. Arterioscler. Thromb. J. Vasc. Biol. 1992, 12, 529–533. [Google Scholar] [CrossRef]
- Ros, E.; Núñez, I.; Pérez-Heras, A.; Serra, M.; Gilabert, R.; Casals, E.; Deulofeu, R. A walnut diet improves endothelial function in hypercholesterolemic subjects: A randomized crossover trial. Circulation 2004, 109, 1609. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Imaizumi, K.; Sato, M.; Hirooka, Y.; Sakai, K.; Takeshita, A.; Kono, M. Serum lipid profiles in japanese women and men during consumption of walnuts. Eur. J. Clin. Nutr. 2002, 56, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Munoz, S.; Merlos, M.; Zambon, D.; Rodriguez, C.; Sabate, J.; Ros, E.; Laguna, J.C. Walnut-enriched diet increases the association of LDL from hypercholesterolemic men with human HEPG2 cells. J. Lipid Res. 2001, 42, 2069–2076. [Google Scholar] [PubMed]
- Fukuda, T.; Ito, H.; Yoshida, T. Antioxidative polyphenols from walnuts (Juglans regia L.). Phytochemistry 2003, 63, 795–801. [Google Scholar] [CrossRef]
- Anderson, K.J.; Teuber, S.S.; Gobeille, A.; Cremin, P.; Waterhouse, A.L.; Steinberg, F.M. Walnut polyphenolics inhibit in vitro human plasma and LDL oxidation. J. Nutr. 2001, 131, 2837–2842. [Google Scholar] [PubMed]
- Chen, C.Y.; Blumberg, J.B. In vitro activity of almond skin polyphenols for scavenging free radicals and inducing quinone reductase. J. Agric. Food Chem. 2008, 56, 4427–4434. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Milbury, P.E.; Chung, S.K.; Blumberg, J. Effect of almond skin polyphenolics and quercetin on human LDL and apolipoprotein B-100 oxidation and conformation. J. Nutr. Biochem. 2007, 18, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Jambazian, P.R.; Haddad, E.; Rajaram, S.; Tanzman, J.; Sabate, J. Almonds in the diet simultaneously improve plasma alpha-tocopherol concentrations and reduce plasma lipids. J. Am. Diet. Assoc. 2005, 105, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Vivancos, M.; Moreno, J.J. Beta-sitosterol modulates antioxidant enzyme response in raw 264.7 macrophages. Free Radic. Biol. Med. 2005, 39, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Segura, R.; Javierre, C.; Lizarraga, M.A.; Ros, E. Other relevant components of nuts: Phytosterols, folate and minerals. Br. J. Nutr. 2006, 96 (Suppl. 2), S36–S44. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Thomson, C.D.; Chisholm, A.; McLachlan, S.K.; Campbell, J.M. Brazil nuts: An effective way to improve selenium status. Am. J. Clin. Nutr. 2008, 87, 379–384. [Google Scholar] [PubMed]
- Chen, C.Y.; Milbury, P.E.; Lapsley, K.; Blumberg, J.B. Flavonoids from almond skins are bioavailable and act synergistically with vitamins C and E to enhance hamster and human ldl resistance to oxidation. J. Nutr. 2005, 135, 1366–1373. [Google Scholar] [PubMed]
- Jia, X.; Li, N.; Zhang, W.; Zhang, X.; Lapsley, K.; Huang, G.; Blumberg, J.; Ma, G.; Chen, J. A pilot study on the effects of almond consumption on DNA damage and oxidative stress in smokers. Nutr. Cancer 2006, 54, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.; Stonehouse, W.; Mukuddem-Petersen, J.; van der Westhuizen, F.H.; Hanekom, S.M.; Jerling, J.C. The effects of high walnut and cashew nut diets on the antioxidant status of subjects with metabolic syndrome. Eur. J. Nutr. 2007, 46, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.L.; Blake, R.J.; Wills, R.B.; Clayton, E.H. Macadamia nut consumption modulates favourably risk factors for coronary artery disease in hypercholesterolemic subjects. Lipids 2007, 42, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jia, X.; Chen, C.Y.; Blumberg, J.B.; Song, Y.; Zhang, W.; Zhang, X.; Ma, G.; Chen, J. Almond consumption reduces oxidative DNA damage and lipid peroxidation in male smokers. J. Nutr. 2007, 137, 2717–2722. [Google Scholar] [PubMed]
- Jenkins, D.J.; Kendall, C.W.; Marchie, A.; Josse, A.R.; Nguyen, T.H.; Faulkner, D.A.; Lapsley, K.G.; Blumberg, J. Almonds reduce biomarkers of lipid peroxidation in older hyperlipidemic subjects. J. Nutr. 2008, 138, 908–913. [Google Scholar] [PubMed]
- Kay, C.D.; Gebauer, S.K.; West, S.G.; Kris-Etherton, P.M. Pistachios increase serum antioxidants and lower serum oxidized-LDL in hypercholesterolemic adults. J. Nutr. 2010, 140, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Haddad, E.H.; Gaban-Chong, N.; Oda, K.; Sabate, J. Effect of a walnut meal on postprandial oxidative stress and antioxidants in healthy individuals. Nutr. J. 2014, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Haddad, E.; Jambazian, P.; Karunia, M.; Tanzman, J.; Sabaté, J. A pecan-enriched diet increases γ-tocopherol/cholesterol and decreases thiobarbituric acid reactive substances in plasma of adults. Nutr. Res. 2006, 26, 397–402. [Google Scholar] [CrossRef]
- McKay, D.L.; Chen, C.Y.; Yeum, K.J.; Matthan, N.R.; Lichtenstein, A.H.; Blumberg, J.B. Chronic and acute effects of walnuts on antioxidant capacity and nutritional status in humans: A randomized, cross-over pilot study. Nutr. J. 2010, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Regueiro, J.; Sanchez-Gonzalez, C.; Vallverdu-Queralt, A.; Simal-Gandara, J.; Lamuela-Raventos, R.; Izquierdo-Pulido, M. Comprehensive identification of walnut polyphenols by liquid chromatography coupled to linear ion trap-orbitrap mass spectrometry. Food Chem. 2014, 152, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Jalali-Khanabadi, B.A.; Mozaffari-Khosravi, H.; Parsaeyan, N. Effects of almond dietary supplementation on coronary heart disease lipid risk factors and serum lipid oxidation parameters in men with mild hyperlipidemia. J. Altern. Complement. Med. 2010, 16, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Liu, Y.H.; Chen, C.M.; Chang, W.H.; Chen, C.Y. The effect of almonds on inflammation and oxidative stress in chinese patients with type 2 diabetes mellitus: A randomized crossover controlled feeding trial. Eur. J. Nutr. 2013, 52, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, A.D.; Manson, J.E.; Rifai, N.; Buring, J.E.; Ridker, P.M. C-reactive protein, interleukin 6, and risk of developing type 2 diabetes mellitus. JAMA 2001, 286, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Peraldi, P.; Budavari, A.; Ellis, R.; White, M.F.; Spiegelman, B.M. Irs-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF-alpha- and obesity-induced insulin resistance. Science 1996, 271, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Hennekens, C.H.; Buring, J.E.; Rifai, N. C-reactive protein and other markers of inflammation in the prediction of cardiovascular disease in women. N. Engl. J. Med. 2000, 342, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Malik, V.S.; Keum, N.; Hu, F.B.; Giovannucci, E.L.; Stampfer, M.J.; Willett, W.C.; Fuchs, C.S.; Bao, Y. Associations between nut consumption and inflammatory biomarkers. Am. J. Clin. Nutr. 2016, 104, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Arpon, A.; Riezu-Boj, J.I.; Milagro, F.I.; Razquin, C.; Martinez-Gonzalez, M.A.; Corella, D.; Estruch, R.; Casas, R.; Fito, M.; Ros, E.; et al. Adherence to mediterranean diet is associated with methylation changes in inflammation-related genes in peripheral blood cells. J. Physiol. Biochem. 2017, 73, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Etherton, T.D.; Martin, K.R.; West, S.G.; Gillies, P.J.; Kris-Etherton, P.M. Dietary alpha-linolenic acid reduces inflammatory and lipid cardiovascular risk factors in hypercholesterolemic men and women. J. Nutr. 2004, 134, 2991–2997. [Google Scholar] [PubMed]
- Zhao, G.; Etherton, T.D.; Martin, K.R.; Gillies, P.J.; West, S.G.; Kris-Etherton, P.M. Dietary alpha-linolenic acid inhibits proinflammatory cytokine production by peripheral blood mononuclear cells in hypercholesterolemic subjects. Am. J. Clin. Nutr. 2007, 85, 385–391. [Google Scholar] [PubMed]
- Rajaram, S.; Connell, K.M.; Sabate, J. Effect of almond-enriched high-monounsaturated fat diet on selected markers of inflammation: A randomised, controlled, crossover study. Br. J. Nutr. 2010, 103, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Alonso, P.; Salas-Salvado, J.; Baldrich-Mora, M.; Juanola-Falgarona, M.; Bullo, M. Beneficial effect of pistachio consumption on glucose metabolism, insulin resistance, inflammation, and related metabolic risk markers: A randomized clinical trial. Diabetes Care 2014, 37, 3098–3105. [Google Scholar] [CrossRef] [PubMed]
- Gulati, S.; Misra, A.; Pandey, R.M. Effect of almond supplementation on glycemia and cardiovascular risk factors in Asian Indians in North India with type 2 diabetes mellitus: A 24-week study. Metab. Syndr. Relat. Disord. 2017, 15, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of N-3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, 1467S–1476S. [Google Scholar] [PubMed]
- Yu, Y.M.; Wang, Z.H.; Liu, C.H.; Chen, C.S. Ellagic acid inhibits IL-1beta-induced cell adhesion molecule expression in human umbilical vein endothelial cells. Br. J. Nutr. 2007, 97, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Papoutsi, Z.; Kassi, E.; Chinou, I.; Halabalaki, M.; Skaltsounis, L.A.; Moutsatsou, P. Walnut extract (Juglans regia L.) and its component ellagic acid exhibit anti-inflammatory activity in human aorta endothelial cells and osteoblastic activity in the cell line KS483. Br. J. Nutr. 2008, 99, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Del Gobbo, L.C.; Imamura, F.; Wu, J.H.; de Oliveira Otto, M.C.; Chiuve, S.E.; Mozaffarian, D. Circulating and dietary magnesium and risk of cardiovascular disease: A systematic review and meta-analysis of prospective studies. Am. J. Clin. Nutr. 2013, 98, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Jin, F.; Hao, Y.; Li, H.; Tang, T.; Wang, H.; Yan, W.; Dai, K. Magnesium and the risk of cardiovascular events: A meta-analysis of prospective cohort studies. PLoS ONE 2013, 8, e57720. [Google Scholar] [CrossRef] [PubMed]
- Dibaba, D.T.; Xun, P.; He, K. Dietary magnesium intake is inversely associated with serum C-reactive protein levels: Meta-analysis and systematic review. Eur. J. Clin. Nutr. 2014, 68, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, T.Y.; van Dam, R.M.; Manson, J.E.; Hu, F.B. Magnesium intake and plasma concentrations of markers of systemic inflammation and endothelial dysfunction in women. Am. J. Clin. Nutr. 2007, 85, 1068–1074. [Google Scholar] [PubMed]
- Gharibeh, M.Y.; Al Tawallbeh, G.M.; Abboud, M.M.; Radaideh, A.; Alhader, A.A.; Khabour, O.F. Correlation of plasma resistin with obesity and insulin resistance in type 2 diabetic patients. Diabetes Metab. 2010, 36, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Degawa-Yamauchi, M.; Bovenkerk, J.E.; Juliar, B.E.; Watson, W.; Kerr, K.; Jones, R.; Zhu, Q.; Considine, R.V. Serum resistin (FIZZ3) protein is increased in obese humans. J. Clin. Endocrinol. Metab. 2003, 88, 5452–5455. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, D.Y.; Cao, J.; He, Z.Y.; Zhu, B.P.; Long, M. High serum resistin level may be an indicator of the severity of coronary disease in acute coronary syndrome. Chin. Med. Sci. J. 2009, 24, 161–166. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, Y.; Li, F.; He, J.; Ding, H.; Yan, L.; Cheng, H. Resistin induces insulin resistance by both AMPK-dependent and ampk-independent mechanisms in HEPG2 cells. Endocrine 2009, 36, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, A.O.; Jacobs, D.R., Jr.; Sanchez, O.A.; Goff, D.C., Jr.; Reiner, A.P.; Gross, M.D. Oxidative stress, inflammation, endothelial dysfunction and incidence of type 2 diabetes. Cardiovasc. Diabetol. 2016, 15, 51. [Google Scholar] [CrossRef] [PubMed]
- Meigs, J.B.; Hu, F.B.; Rifai, N.; Manson, J.E. Biomarkers of endothelial dysfunction and risk of type 2 diabetes mellitus. JAMA 2004, 291, 1978–1986. [Google Scholar] [CrossRef] [PubMed]
- Laight, D.W.; Carrier, M.J.; Anggard, E.E. Antioxidants, diabetes and endothelial dysfunction. Cardiovasc. Res. 2000, 47, 457–464. [Google Scholar] [CrossRef]
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Versari, D.; Daghini, E.; Virdis, A.; Ghiadoni, L.; Taddei, S. Endothelial dysfunction as a target for prevention of cardiovascular disease. Diabetes Care 2009, 32 (Suppl. 2), S314–S321. [Google Scholar] [CrossRef] [PubMed]
- Hamasaki, S.; Higano, S.; Nishimura, R. Long-term follow-up of patients with mild coronary artery disease and endothelial dysfunction. Circulation 2000, 101, 948–954. [Google Scholar]
- Federici, M.; Pandolfi, A.; De Filippis, E.A.; Pellegrini, G.; Menghini, R.; Lauro, D.; Cardellini, M.; Romano, M.; Sesti, G.; Lauro, R.; et al. G972R IRS-1 variant impairs insulin regulation of endothelial nitric oxide synthase in cultured human endothelial cells. Circulation 2004, 109, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, E.L.; Cho, H.; Birnbaum, M.J. Role of AKT/protein kinase B in metabolism. Trends Endocrinol. Metab. 2002, 13, 444–451. [Google Scholar] [CrossRef]
- Montagnani, M.; Ravichandran, L.V.; Chen, H.; Esposito, D.L.; Quon, M.J. Insulin receptor substrate-1 and phosphoinositide-dependent kinase-1 are required for insulin-stimulated production of nitric oxide in endothelial cells. Mol. Endocrinol. 2002, 16, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Dimmeler, S.; Fleming, I.; Fisslthaler, B.; Hermann, C.; Busse, R.; Zeiher, A.M. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature 1999, 399, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Fulton, D.; Gratton, J.P.; McCabe, T.J.; Fontana, J.; Fujio, Y.; Walsh, K.; Franke, T.F.; Papapetropoulos, A.; Sessa, W.C. Regulation of endothelium-derived nitric oxide production by the protein kinase akt. Nature 1999, 399, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Wang, K.Q.; Zhou, G.M.; Zuo, J.; Ge, J.B. Endothelin-1 promoted proliferation of vascular smooth muscle cell through pathway of extracellular signal-regulated kinase and cyclin D1. Acta Pharmacol. Sin. 2003, 24, 563–568. [Google Scholar] [PubMed]
- Eringa, E.C.; Stehouwer, C.D.; van Nieuw Amerongen, G.P.; Ouwehand, L.; Westerhof, N.; Sipkema, P. Vasoconstrictor effects of insulin in skeletal muscle arterioles are mediated by ERK1/2 activation in endothelium. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, 2043–2048. [Google Scholar] [CrossRef] [PubMed]
- De Caterina, R.; Libby, P.; Peng, H.B.; Thannickal, V.J.; Rajavashisth, T.B.; Gimbrone, M.A., Jr.; Shin, W.S.; Liao, J.K. Nitric oxide decreases cytokine-induced endothelial activation. Nitric oxide selectively reduces endothelial expression of adhesion molecules and proinflammatory cytokines. J. Clin. Investig. 1995, 96, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Paul, R.; Adams, H.R.; Myers, J.L. Effects of cytokines tumor necrosis factor [alpha] and interleukin 1 [beta] on endotoxin-mediated inhibition of endothelium-derived relaxing factor bioactivity and nitric oxide production in vascular endothelium. Shock 1994, 1, 73–78. [Google Scholar] [CrossRef]
- Grau, G.E.; Lou, J. Tnf in vascular pathology: The importance of platelet-endothelium interactions. Res. Immunol. 1993, 144, 355–363. [Google Scholar] [CrossRef]
- Carluccio, M.A.; Massaro, M.; Bonfrate, C.; Siculella, L.; Maffia, M.; Nicolardi, G.; Distante, A.; Storelli, C.; De Caterina, R. Oleic acid inhibits endothelial activation: A direct vascular antiatherogenic mechanism of a nutritional component in the Mediterranean diet. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Montagnani, M.; Funahashi, T.; Shimomura, I.; Quon, M.J. Adiponectin stimulates production of nitric oxide in vascular endothelial cells. J. Biol. Chem. 2003, 278, 45021–45026. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Suzuki, M.; Hattori, S.; Kasai, K. Globular adiponectin upregulates nitric oxide production in vascular endothelial cells. Diabetologia 2003, 46, 1543–1549. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Kihara, S.; Arita, Y.; Maeda, K.; Kuriyama, H.; Okamoto, Y.; Hotta, K.; Nishida, M.; Takahashi, M.; Nakamura, T.; et al. Novel modulator for endothelial adhesion molecules: Adipocyte-derived plasma protein adiponectin. Circulation 1999, 100, 2473–2476. [Google Scholar] [CrossRef] [PubMed]
- Aronis, K.N.; Vamvini, M.T.; Chamberland, J.P.; Sweeney, L.L.; Brennan, A.M.; Magkos, F.; Mantzoros, C.S. Short-term walnut consumption increases circulating total adiponectin and apolipoprotein a concentrations, but does not affect markers of inflammation or vascular injury in obese humans with the metabolic syndrome: Data from a double-blinded, randomized, placebo-controlled study. Metabolism 2012, 61, 577–582. [Google Scholar] [PubMed]
- Katz, D.L.; Davidhi, A.; Ma, Y.; Kavak, Y.; Bifulco, L.; Njike, V.Y. Effects of walnuts on endothelial function in overweight adults with visceral obesity: A randomized, controlled, crossover trial. J. Am. Coll. Nutr. 2012, 31, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Cortés, B.; Núñez, I.; Cofán, M.; Gilabert, R.; Pérez-Heras, A.; Casals, E.; Deulofeu, R.; Ros, E. Acute effects of high-fat meals enriched with walnuts or olive oil on postprandial endothelial function. J. Am. Coll. Cardiol. 2006, 48, 1666–1671. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Njike, V.Y.; Millet, J.; Dutta, S.; Doughty, K.; Treu, J.A.; Katz, D.L. Effects of walnut consumption on endothelial function in type 2 diabetic subjects: A randomized controlled crossover trial. Diabetes Care 2010, 33, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Kasliwal, R.R.; Bansal, M.; Mehrotra, R.; Yeptho, K.P.; Trehan, N. Effect of pistachio nut consumption on endothelial function and arterial stiffness. Nutrition 2015, 31, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.; Valacchi, G.; Pagnin, E.; Shao, Q.; Gross, H.B.; Calo, L.; Yokoyama, W. Walnuts reduce aortic ET-1 mrna levels in hamsters fed a high-fat, atherogenic diet. J. Nutr. 2006, 136, 428–432. [Google Scholar] [PubMed]
- Gornik, H.L.; Creager, M.A. Arginine and endothelial and vascular health. J. Nutr. 2004, 134, 2880S–2887S, discussion 2895S. [Google Scholar] [PubMed]
- Shechter, M.; Sharir, M.; Labrador, M.J.; Forrester, J.; Silver, B.; Bairey Merz, C.N. Oral magnesium therapy improves endothelial function in patients with coronary artery disease. Circulation 2000, 102, 2353–2358. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, M.; Dominguez, L.J.; Galioto, A.; Pineo, A.; Belvedere, M. Oral magnesium supplementation improves vascular function in elderly diabetic patients. Magn. Res. 2010, 23, 131–137. [Google Scholar]
- Wells, B.J.; Mainous, A.G., 3rd; Everett, C.J. Association between dietary arginine and C-reactive protein. Nutrition 2005, 21, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Blum, A.; Porat, R.; Rosenschein, U.; Keren, G.; Roth, A.; Laniado, S.; Miller, H. Clinical and inflammatory effects of dietary l-arginine in patients with intractable angina pectoris. Am. J. Cardiol. 1999, 83, 1488–1490. [Google Scholar] [CrossRef]
- Adams, M.R.; McCredie, R.; Jessup, W.; Robinson, J.; Sullivan, D.; Celermajer, D.S. Oral l-arginine improves endothelium-dependent dilatation and reduces monocyte adhesion to endothelial cells in young men with coronary artery disease. Atherosclerosis 1997, 129, 261–269. [Google Scholar] [CrossRef]
- Yin, W.-H.; Chen, J.-W.; Tsai, C.; Chiang, M.-C.; Young, M.S.; Lin, S.-J. l-arginine improves endothelial function and reduces ldl oxidation in patients with stable coronary artery disease. Clin. Nutr. 2005, 24, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Mohammadifard, N.; Salehi-Abargouei, A.; Salas-Salvado, J.; Guasch-Ferre, M.; Humphries, K.; Sarrafzadegan, N. The effect of tree nut, peanut, and soy nut consumption on blood pressure: A systematic review and meta-analysis of randomized controlled clinical trials. Am. J. Clin. Nutr. 2015, 101, 966–982. [Google Scholar] [CrossRef] [PubMed]
- Toledo, E.; Hu, F.B.; Estruch, R.; Buil-Cosiales, P.; Corella, D.; Salas-Salvado, J.; Covas, M.I.; Aros, F.; Gomez-Gracia, E.; Fiol, M.; et al. Effect of the mediterranean diet on blood pressure in the predimed trial: Results from a randomized controlled trial. BMC Med. 2013, 11, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenech, M.; Roman, P.; Lapetra, J.; Garcia de la Corte, F.J.; Sala-Vila, A.; de la Torre, R.; Corella, D.; Salas-Salvado, J.; Ruiz-Gutierrez, V.; Lamuela-Raventos, R.M.; et al. Mediterranean diet reduces 24-h ambulatory blood pressure, blood glucose, and lipids: One-year randomized, clinical trial. Hypertension 2014, 64, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Medina-Remon, A.; Tresserra-Rimbau, A.; Pons, A.; Tur, J.A.; Martorell, M.; Ros, E.; Buil-Cosiales, P.; Sacanella, E.; Covas, M.I.; Corella, D.; et al. Effects of total dietary polyphenols on plasma nitric oxide and blood pressure in a high cardiovascular risk cohort. The predimed randomized trial. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 60–67. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
Table 1.
Nutrient content of raw nuts per 100 g *.
| Almond | Walnut | Hazelnut | Macadamia | Pecan | Pistachio | Brazil Nut | Cashew Nut | Peanut | Pine Nut | Chest Nut | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Energy (kcal) | 579 | 654 | 628 | 718 | 691 | 560 | 659 | 553 | 567 | 673 | 213 |
| Carbohydrate (g) | 21.55 | 13.71 | 16.7 | 13.82 | 13.86 | 27.17 | 11.74 | 30.19 | 16 | 13.08 | 45.54 |
| Protein (g) | 21.15 | 15.23 | 14.95 | 7.91 | 9.17 | 20.16 | 14.32 | 18.22 | 26 | 13.69 | 2.42 |
| Lysine (g) | 0.568 | 0.424 | 0.42 | 0.018 | 0.287 | 1.138 | 0.49 | 0.928 | 0.926 | 0.54 | 0.143 |
| Arginine (g) | 2.465 | 2.278 | 2.211 | 1.402 | 1.177 | 2.134 | 2.14 | 2.123 | 3.085 | 2.413 | 0.173 |
| Total fat (g) | 49.93 | 65.21 | 60.75 | 75.77 | 71.97 | 45.32 | 67.1 | 43.85 | 49 | 68.37 | 2.26 |
| Saturated fat (g) | 3.8 | 6.126 | 4.464 | 12.061 | 6.18 | 5.907 | 16.134 | 7.783 | 7 | 4.89 | 0.425 |
| MUFA (g) | 31.55 | 8.933 | 45.652 | 58.877 | 40.801 | 23.257 | 23.879 | 23.797 | 24 | 18.76 | 0.78 |
| PUFA (g) | 12.329 | 47.174 | 7.92 | 1.502 | 21.614 | 14.38 | 24.399 | 7.845 | 16 | 34.07 | 0.894 |
| Total Fiber (g) | 12.5 | 6.7 | 9.7 | 8.6 | 9.6 | 10.6 | 7.5 | 3 | 8.5 | 3.7 | 8.1 |
| Folate (μg) | 44 | 98 | 113 | 11 | 22 | 51 | 22 | 25 | 240 | 34 | 62 |
| Calcium (mg) | 269 | 98 | 114 | 85 | 70 | 105 | 160 | 37 | 92 | 16 | 27 |
| Magnesium (mg) | 270 | 158 | 163 | 130 | 121 | 121 | 376 | 292 | 168 | 251 | 32 |
| Sodium (mg) | 1 | 2 | 0 | 5 | 0 | 1 | 3 | 12 | 18 | 2 | 3 |
| Potassium (mg) | 733 | 441 | 680 | 368 | 410 | 1025 | 659 | 660 | 705 | 597 | 518 |
| Copper (mg) | 1.031 | 1586 | 1725 | 0.756 | 1.2 | 1.3 | 1.743 | 2195 | 1.144 | 1.324 | 0.447 |
| Iron (mg) | 3.71 | 2.91 | 4.7 | 3.69 | 2.53 | 3.92 | 2.43 | 6.68 | 4.58 | 5.53 | 1.01 |
| Zinc (mg) | 3.12 | 3.09 | 2.45 | 1.3 | 4.53 | 2.2 | 4.06 | 5.78 | 3.27 | 6.45 | 0.52 |
| Selenium (μg) | 4.1 | 4.9 | 2.4 | 3.6 | 3.8 | 7 | 1917 | 19.9 | 7.2 | 0.7 | NA |
| α-tocopherol (mg) | 25.63 | 0.7 | 15.03 | 0.54 | 1.4 | 2.86 | 5.65 | 0 | 8.33 | 9.33 | NA |
| β-tocopherol (mg) | 0.23 | 0.15 | 0.33 | 0 | 0.39 | 0 | 0.01 | 0.03 | NA | 0 | NA |
| γ-tocopherol (mg) | 0.07 | 20.83 | 0 | 0 | 24.44 | 20.41 | 9.56 | 5.31 | NA | 11.15 | NA |
| δ-tocopherol (mg) | 0.07 | 1.89 | 0 | 0 | 0.47 | 0.8 | 0.63 | 0.36 | NA | 0 | NA |
| Total phytosterol (mg) | ~198 | ~110.2 | ~122 | ~116 | ~158.7 | ~214 | ~123.8 | ~151 | NA | 236.1 | 22 |
| Stigmasterol | 4 | 0 | 1 | 0 | 3 | 5 | 6 | 0 | 0 | - | |
| Campesterol | 5 | 5 | 7 | 8 | 6 | 10 | 2 | 9 | 20 | - | |
| B-sitosterol | 130 | 87 | 102 | 108 | 117 | 198 | 64 | 113 | 132 | - | |
| δ 5-avenasterol | 21 | 7.3 | 2.6 | - | 14.3 | - | 19.7 (δ avenasterol + B sitostanol) | 14 | 40.1 | - | |
| B-sitostanol | 4 | - | 3.9 | - | - | - | - | 5.9 | - | ||
| Campestanol | 2 | 2.3 | 3 | - | 2.8 | - | - | 2 | 3.9 | - | |
| Others | 32 | 8.6 | 2.5 | - | 15.6 | - | 31.8 | 13 | 34.2 | - | |
| Total polyphenol (mg) # | 287 | 1576 | 687 | 126 | 1284 | 867 | 224 | 232.9 | NA | NA | NA |
| Total polyphenol (mg) $ | 212.9 ± 12.3 | 1580.5 ± 58 | 314.8 ± 47.3 | 497.8 ± 52.6 | 1463.9 ± 32.3 | 571.8 ± 12.5 | 169.2 ± 14.6 | 316.4 ± 7.0 | 645.9 ± 47 | 152.9 ± 14.1 | NA |
| Flavonoids (mg) $ | 93.5 ± 10.8 | 744.8 ± 93.3 | 113.7 ± 30.2 | 137.9 ± 9.9 | 704.7 ± 29.5 | 143.3 ± 18.7 | 107.8 ± 6.0 | 63.7 ± 2.1 | 189.8 ± 13.1 | 45.0 ± 5.4 | NA |
| Ellagitannins (mg) ‡ | ND | 823 ± 59 | ND | ND | 301 ± 7 | ND | ND | ND | ND | NA | 149 ± 3 |
| Proanthocyanidins (mg) # | 176 | 60 | 491 | NA | 477 | 226 | NA | 2 | NA | NA | 0 |
| Carotenoids (μg) | 2 | NA | 106 | NA | 55 | 332 | 0 | NA | NA | NA | NA |
| Lutein + zeaxanthin (μg) | 1 | 9 | 92 | 0 | 17 | 2903 | 0 | 22 | 0 | 9 | NA |
* Source: United States Department of Agriculture Nutrient Database for Standard Reference; Nutrient data for 12,061 (nuts, almonds), 12,155 (nuts, walnuts, English), 12,120 (nuts, hazelnuts or filberts), 12,131 (nuts, macadamia nuts, raw), 12,142 (nuts, pecans), 12,151 (nuts, pistachio nuts, raw), 12,078 (nuts, brazilnuts, dried, unblanched), 12,087 (nuts, cashew nuts, raw), 16,087 (peanuts, all types, raw), 12,147 (nuts, pine nuts, dried), 12,097 (nuts, chestnuts, European, raw, unpeeled) [16]. # Data were obtained from [17], $ data were obtained from [18] and data ‡ were obtained from [19]; NA: not available; ND: not detected; MUFA, monounsaturated fatty acids; PUFA, polyunsaturated fatty acids.
Table 2.
Summary of clinical trials examining the effect of nut consumption on glycemic control.
| Description | Reference | Subjects | Study Design/Period | Nut Type | Diet Intervention | Results |
|---|---|---|---|---|---|---|
| Meta-analysis of 12 RCTs for glycemic control | Viguiliouk et al., 2014 [20] | Subjects with T2DM | RCTs/≥3-week follow-up period | Almonds, pistachios, walnuts, pecans, hazelnuts, peanuts, cashews, macadamias | Diet supplemented with tree nuts vs. isocaloric diet without tree nuts | ↓ HbA1c (p = 0.0003), ↓ fasting glucose, |
| ↔ fasting insulin, ↔ HOMA-IR after consumption of tree nuts at a median dose of 56 g/day | ||||||
| Chronic studies that were not included in meta-analysis of Viguiliouk et al., 2014 [20] | Scott et al., 2003 [25] | 17 subjects with IGT (n = 10) or T2DM (n = 7) | Parallel, randomised/42 weeks | Almonds | Almond-enriched diet (High Protein-High MUFA diet; 25% of energy from protein, 40% fat, and 22% MUFAs) vs. AHA diet (15% of energy from protein, 30% fat, and 15% MUFAs) | ↑ glucose control |
| ↔ TG, ↔ LDL-C, ↔ fasting glucose | ||||||
| Wien et al., 2003 [26] | 65 overweight and obese subjects | Randomised prospective/24 weeks | Almonds | Formula-based low-calorie diet enriched with 84 g/day of almonds vs. complex carbohydrates matching calories and protein | ↓ 62% body weight/BMI, | |
| ↓ 50% waist circumference, | ||||||
| ↓ 56% greater reduction in fat mass compared with the complex carbohydrate diet | ||||||
| ↓ glucose, ↓ insulin, ↓ HOMA-IR, ↓ TC, ↓ TG, ↓ LDL-C, ↓ LDL-C/HDL-C in both diets | ||||||
| Sari et al., 2010 [24] | 32 healthy young men | Dietary intervention/8 weeks | Pistachios | Subjects consumed a Mediterranean diet for 4 weeks and in the next 4 weeks they supplemented the Mediterranean diet with pistachios for 4 weeks replacing 32% of the energy obtained from MUFAs in the control diet (20% of energy) | ↓ glucose levels (−8.8 ± 8.5% p < 0.001) | |
| ↓ TC (p < 0.001), ↓ LDL-C (p < 0.001), ↓ triacylglycerol, ↔ HDL-C, | ||||||
| ↓ TC-C/HDL-C, ↓ LDL-C/HDL-C (p < 0.001), | ||||||
| ↑ endothelium-dependent vasodilation (p = 0.002) | ||||||
| ↓ lipid hydroperoxide (p < 0.001), ↓ MDA (p < 0.001), ↓ IL-6 (p < 0.001), | ||||||
| ↑ SOD (p < 0.001), | ||||||
| ↔ hs-CRP, ↔ TNF-α | ||||||
| compared with a Mediterranean diet | ||||||
| Wien et al., 2010 [36] | 65 subjects with prediabetes | Randomized parallel trial/16 weeks | Almonds | ADA diet containing 20% of energy from almonds (60 g/day of pre-packaged raw or dry roasted almonds) vs. almond-free control diet | ↓ fasting insulin (p = 0.002), ↓ HOMA-IR (p = 0.007), ↓ HOMA-B (p = 0.001), ↔ fasting glucose, ↓. LDL-C, ↔ BMI, | |
| ↓ weight, ↔ waist circumference | ||||||
| Parham et al., 2014 [23] | 48 subjects with T2DM | Double-blind, randomized, placebo-controlled, crossover/12 weeks | Pistachios | 25 g pistachio nuts twice a day as a snack vs. control diet without nuts | ↓ HbA1c (−0.4%; p ≤ 0.001), ↓ fasting blood glucose (−16 mg/dL; p ≤ 0.001), | |
| ↔ BMI, ↔ BP, ↔ HOMA-IR, ↔ hs-CRP | ||||||
| Sauder et al., 2015 [22] | 30 subjects with well-controlled T2DM | Randomized, crossover, controlled/4 weeks | Pistachios | Healthy diet with pistachios contributing 20% of total energy. Low-fat or fat-free snacks (e.g., pretzels) in the control diet were substituted with roasted pistachios providing 20% of daily energy (59 to 128 g/day of pistachios). Half of amount of pistachio was ingested unsalted vs. AHA Therapeutic Lifestyle Changes diet (26.9% total fat, 6.7% SFA, 186 mg/day cholesterol). | ↓ fructosamine, ↓ TC, ↓ ratio of TC to HDL-C, ↓ TG (p = 0.003), ↔ HbA1c, | |
| ↔ fasting glucose, ↔ insulin, ↔ hs-CRP, | ||||||
| ↔ ICAM, ↔ VCAM, ↔ endothelial function | ||||||
| Zibaeenezhad et al., 2016 [21] | 100 subjects with T2DM | Randomized control/3 months | Walnut oil | Walnut oil (15 g/day) group (n = 50) vs. a control group without any interventions (n = 50) | ↓ HbA1c by 8 ± 22% (p = 0.005) and | |
| ↓ fasting glucose by 8 ± 17% (p = 0.001) within the walnut group. | ||||||
| But, ↔ HbA1c, ↔ fasting glucose compared with the control. | ||||||
| ↔ body weight, BMI, BP in the two groups | ||||||
| Acute studies that were not included in meta-analysis | Johnston et al., 2005 [30] | 11 healthy subjects | Randomised crossover | Peanuts | 6 meals (bagel and juice meal, bagel and juice meal + vinegar, bagel and juice meal + peanut, chicken and rice meal, chicken and rice meal + vinegar, and chicken and rice meal + peanut) were compared | ↓ glucose response in the 60-min (p < 0.05) following the ingestion of vinegar or peanuts with bagel and juice meal |
| Jenkins et al., 2006 [32] | 15 healthy subjects | Meals consumed in random order on separate days, 4-h postprandial test | Almonds | 3 test meals; almonds (60 g) + bread (97 g), parboiled rice meal (68 g cheese + 14 g butter + 60 g parboiled rice) and mashed potato meal (62 g cheese + 16 g butter + 68 g mashed potatoes) vs. 2 bread control meals | ↓ postprandial glucose and insulin (p < 0.001) responses | |
| ↑ protein thiol | ||||||
| ↔ total antioxidant capacity compared with other test meals | ||||||
| Josse et al., 2007 [33] | 9 healthy subjects | Meals consumed in random order on separate days, 2-h postprandial test | Almonds | White bread + 0 g almond, white bread + 30 g almond, white bread + 60 g almond, white bread + 90 g almond, | ↓ meal glycemic index in a dose-dependent manner (90 g almond + white bread > 60 g almond + white bread > 30 g almond + white bread; p = 0.001) | |
| Each meal contained 50 g CHO from white bread | ||||||
| Kendall et al., 2011 [35] | 14 normoglycemic subjects and 10 subjects with T2DM | Meals consumed in random order on separate days, 2-h postprandial test | Mixed nuts | Mixed nuts (30, 60 and 90 g) were consumed with white bread | ↓ postprandial glycemic responses | |
| Each meal contained 50 g CHO from white bread | ||||||
| Kendall et al., 2011 [27] | 10 healthy adults | Two acute tests; each test consisting of 2-h postprandial test | Pistachios | <Study 1> 8 test meals: white bread (50 g CHO), 28 g pistachio, 56 g pistachio, 84 g pistachio, white bread + 28 g pistachios, white bread + 56 g pistachios and white bread + 84 g pistachios, aiming to assess the dose-response effect of 28, 56 and 84 g pistachios ingested alone or with white bread. <Study 2> 7 test meals: white bread (50 g CHO), parboiled rice, pasta and instant mashed potatoes and parboiled rice + 56 g pistachios, pasta + 56 g pistachios instant mashed potatoes + 56 g pistachios, aiming to assess effective 56 g of pistachio on glycemic response with different types of CHO foods | Study 1: relative glycemic responses; for 28 g, 56 g and 84 g pistachios were 5.7%, 3.8% and 9.3% respectively compared to white bread, ↓ relative glycemic responses; in a dose-dependent manner with 89% (p = 0.1), 67% (p = 0.009) and 52% (p < 0.001) for 28 g, 56 g and 84 g pistachios added to white bread respectively Study 2: ↓ relative glycemic responses; after adding 56 g pistachios to rice, pasta and instant mashed potatoes with 59% (p = 0.02), 56% (p = 0.025) and 87% (p = 0.06) respectively, compared with 73%, 95%, 109% | |
| Mori et al., 2011 [31] | 14 subjects with IGT | Randomized, 5-arm, crossover | Almonds | Whole almonds into breakfast vs. almond butter into breakfast vs. defatted almond flour into breakfast vs. almond oil into breakfast vs. no almonds into breakfast | ↓ postprandial glycemia and ↑ satiety following whole almonds ↓ the second-meal NEFA response following whole almonds or almond oil ↔ GLP-1 between meals | |
| All breakfast meals were matched for 75 g carbohydrate | ||||||
| Reis et al., 2013 [29] | 15 obese women with a high T2DM risk | Randomised cross-over | Peanuts | Consumption of a 75 g CHO-matched breakfast containing 42.5 g of whole peanuts without skins or peanut butter or no peanuts to meal and then consumption of standard lunch, aiming to assess first- and second meal responses | ↓ NEFA iAUC (0–240 min), ↓ glucose iAUC (240–490 min), ↑ insulin (120–370 min) ↑ gut satiety hormones (PYY, GLP-1 and CCK), ↓ desire to eat following peanut butter breakfast compared with no peanut breakfast | |
| Kendall et al., 2014 [28] | 20 subjects with metabolic syndrome | Randomized crossover | Pistachios | 5 test meals: | ↓ glucose response ↑ GLP-1 | |
| - 3 meals containing 50 g available CHO: white bread, white bread, butter and cheese, and white bread and pistachios | ||||||
| - 2 meals containing12 g available CHO: white bread, and pistachios | ||||||
| Crouch et al., 2016 [34] | 20 subjects with prediabetes or isolated 1-h postprandial hyperglycemia | Almonds | one-half ounce (14.2 g) dry-roasted almond preload vs. without the almond preload. | ↓ glucose 1 h after a 75-g glucose challenge (p < 0.001). |
ADA, American Diabetes Association; AHA, American Heart Association; BP, blood pressure; BMI, body mass index; CCK, cholecystokinin; CHO, carbohydrate; GLP-1, glucagon-like-peptide 1; HbA1c, glycosylated haemoglobin; HDL-C, high-density lipoprotein cholesterol; HOMA-IR, homeostasis model assessment of insulin resistance index; HOMA-B, homeostasis model analysis for beta-cell function; hs-CRP, high sensitivity C-reactive protein; iAUC, incremental area under curve; ICAM, intracellular adhesion molecule; LDL-C, low density lipoprotein; IGT, impaired glucose tolerance; IL-6, interleukin-6; MDA, malondialdehyde; MUFA, monounsaturated fatty acids; NEFA, non-esterified fatty acids; PYY, peptide YY; RCT, randomised controlled trial; SFA, saturated fatty acid; SOD, superoxide dismutase; TC, total cholesterol; TG, triglyceride; VCAM, vascular cellular adhesion molecule; ↑, increase; ↓, decrease; ↔, no effect.
Table 3.
Summary of clinical trials examining the effect of nut consumption on body weight control.
| Reference | Subjects | Study Design/Period | Nut Type | Diet Intervention | Results |
|---|---|---|---|---|---|
| Spiller et al., 1992 [64] | 26 subjects with hypercholesterolemia | Pre-post supplemental, dietary advice/9 weeks | Almonds | Raw almonds (100 g/day; 34 g/day MUFA, 12 g/day PUFA 6 g/day SFA) to baseline diet | ↔ body weight, ↑ 81 kcal/day energy intake,↓ LDL-C, ↓ TC (p < 0.001), ↔ HDL-C |
| Abbey et al., 1994 [53] | 16 male health subjects | Pre-post consecutive supplemental, dietary advice/9 weeks | Almonds, Walnuts | In the first 3-week period, addition of raw peanuts (50 g/day), coconut cubes (40 g/day), and a coconut confectionary bar (50 g/day), to Australian diet (reference diet) | ↔ body weight, ↔ energy intake ↓ LDL-C, ↓ TC, ↔ HDL-C compared with reference diet |
| During the following 3 weeks, 84 g/day almonds added to Australian diet | |||||
| During the final 3-week period, 68 g/day walnuts added to Australian diet | |||||
| Colquhoun et al., 1996 [76] | 14 subjects with hypercholesterolemia | Randomised crossover, pre-post, dietary advice/4 weeks | Macadamias | Diet enriched with macadamia (40% energy as fat, 20% energy from macadamia nuts) vs. a high-complex-carbohydrate diet | ↔ body weight, ↔ energy intake |
| ↓ LDL-C (p < 0.01), ↓ TC (p < 0.01) on both diets compared with baseline. | |||||
| ↔ HDL-C on both diets | |||||
| Spiller et al., 1998 [63] | 45 subjects with hypercholesterolemia | Randomised parallel arm, dietary advice/4 weeks | Almonds | Almond-based diet vs. olive oil-based diet vs. dairy-based diet | ↔ body weight, ↔ energy intake |
| ↓ TC (p < 0.001), ↓ LDL-C (p < 0.001) compared with control diets | |||||
| Three diets were matched for total fat | ↓ TC, ↓ LDL-C (p < 0.001), ↓ TC:HDL ratio (p < 0.001), ↔ HDL-C within the almond group | ||||
| Chisholm et al., 1998 [67] | 21 subjects with hypercholesterolemia | Randomised crossover, dietary advice/4 weeks | Walnuts | 20% energy of low-fat diet replaced with 78 g/day walnuts vs. low-fat diet | ↔ body weight, ↔ energy intake, ↓ TC, ↓ LDL-C, ↑ HDL-C on both diets compared with baseline. |
| ↓ apo B on the walnut diet compared with low-fat diet | |||||
| Durak et al., 1999 [77] | 30 healthy subjects | Pre-post supplemental, dietary advice | Hazelnuts | 1 g/kg body weight/day of hazelnuts supplemented with a habitual diet vs. baseline | ↑ 0.5 kg body weight, ↓ TC (p < 0.005), ↓ LDL-C (p < 0.0005), |
| ↑ HDL-C, ↑ TG (p < 0.001), ↑ HDL-C/LDL-C,(p <,0.0005) | |||||
| ↓ MDA(p < 0.0005), ↑AOP (p < 0.0005) after hazelnut supplementation | |||||
| Edwards et al., 1999 [72] | 10 subjects with hypercholesterolemia | Randomised, controlled, crossover, dietary advice/3 weeks | Pistachios | 20% energy of habitual diet replaced with pistachios vs. control diet | ↔Body weight, ↔ BP, ↔ energy intake, ↓ TC, ↓ LDL-C, ↓ TC/HDL (p < 0.01), ↑ HDL-C, ↔ TG |
| Morgan et al., 2000 [70] | 19 subjects with normal lipid levels | Randomised parallel arm, pre-post, supplemental/8 weeks | Pecans | 68 g/day pecans + self-selected diet vs. self-selected diet without nuts | ↔ body weight, ↑ 71 kJ energy intake |
| ↓ TC, ↓ HDL-C | |||||
| Zambone et al., 2000 | 49 subjects with hypercholesterolemia | Randomised crossover, dietary advice/6 weeks | Walnuts | 18% energy of habitual diet replaced with walnuts vs. Mediterranean diet | ↔ body weight, ↔ energy intake |
| ↓ TC (p < 0.001), ↓ LDL-C (p < 0.00), ↓ lipoprotein (a) compared with the Mediterranean diet, | |||||
| Curb et al., 2000 [75] | 30 healthy subjects | Randomized crossover/30 days | Macadamias | 3 dietary periods; American diet high in saturated fat (37% energy from fat) vs. AHA Step 1 diet (30% energy from fat) vs. MUFA diet rich in macadamia (37% energy from fat) | ↔ body weight, ↑ energy intake |
| ↓ TC, ↓ LDL-C after macadamia diet compared with American diet | |||||
| Almario et al., 2001 [52] | 18 subjects (7 men and 16 postmenopausal women) with dyslipidaemia | Pre-post consecutive supplemental, dietary advice/6 weeks | Walnuts | 48 g/day walnuts added to a habitual diet vs. | ↔ body weight, ↑ energy intake |
| 48 g/day walnuts added to a low-fat diet vs. | ↓ TC after walnut plus a low-fat diet compared with habitual diet or low-fat diet | ||||
| habitual diet vs. low-fat diet | ↓ LDL-C (p < 0.01) after walnut plus a low-fat diet compared with a low-fat diet | ||||
| McManus et al., 2001 [79] | 101 overweight subjects | Randomized controlled/18 months | Peanuts, almonds, cashews, hazelnuts, macadamias, pecans, pistachios, walnuts | Moderate fat low energy diet (fat: 35% of energy; predominantly PUFAs; nut-enriched diet) vs. low fat, low-energy diet (fat: 20% of energy) | ↓ 4.1 kg body weight, ↓ 1.6 kg/m2 BMI, ↓ 6.9 cm waist circumference in moderate fat group, while ↑ 2.9 kg body weight, ↑ 1.4 kg/m2 BMI, ↑ 2.6 cm waist circumference in low fat group |
| Fraser et al., 2002 [50] | 81 male and female subjects | Randomized crossover/6 months | Almonds | 42–70 g/day almond supplementation (320 kcal/day) vs. without almond supplementation | ↑ 0.40 kg body weight |
| Jenkins et al., 2002 [62] | 27 subjects with hypercholesterolemia | Randomised crossover, dietary advice/4 weeks | Almonds | 3 isoenergetic (mean 423 kcal/day) supplement periods each for 4 weeks; - full-dose almonds (73 g/day), - half-dose almonds plus half-dose muffins, - full-dose muffins (control) Supplements provided 22.2% of energy | ↔ body weight, ↔ energy intake |
| ↓ LDL-C (p < 0.001), ↓ LDL-C:HDL-C (p < 0.001) on both almonds | |||||
| ↓ oxidised LDL (p < 0.001), ↓ lipoprotein (a) on full -dose almonds | |||||
| ↔ pulmonary nitric oxide between control, half-dose, and full-dose almond diet | |||||
| Hyson et al., 2002 [65] | 22 subjects with normal lipid levels | Randomised crossover, pre-post, supplemental/6 weeks | Almonds | 50% of fat energy of habitual diet replaced with almonds (~66 ± 5 g/day), vs. 50% of fat energy of habitual diet replaced with almond oil (~35 ± 2 g/day) | ↔ body weight, ↔ energy intake |
| ↓ TC, ↓ LDL-C, ↓ TG, ↑ HDL compared with baseline | |||||
| Morgan et al., 2002 [69] | 42 subjects with borderline high total cholesterol | Randomised crossover, dietary advice/6 weeks | Walnuts | 64 g/day of walnuts added to low-fat, low-cholesterol diet vs. low-fat, low-cholesterol diet | ↔ body weight, ↔ energy intake |
| ↓ TC, ↓ LDL-C, ↓ TG, ↑ HDL-C | |||||
| ↔ homocysteine ,↓ tPA, ↓PAI-1 | |||||
| Lovejoy et al., 2002 [37] | 20 healthy subjects | Pre-post supplemental study, dietary advice/4 weeks | Almonds | 100 g/day of almond added to habitual diet vs. baseline | ↑ body weight (↑ 0.9 kg; male, ↑ 0.3 kg: female), ↑ ~51 kJ energy intake, ↓ LDL-C (p = 0.0004), ↔ insulin sensitivity compared with baseline |
| Alper et al., 2002 [49] | 15 healthy subjects with normal weight | Crossover, 3 treatment phases (free-feeding, addition and substitution)/30 weeks | Peanuts | In the free-feeding phase, 50% of dietary fat energy from peanuts for 8 weeks with no restriction of the background diet. No dietary advice given. | ↔ body weight during the substitution phase |
| In the addition phase, peanuts added to their habitual diet for 3 weeks with dietary instructions | ↑ 1 kg body weight during the free-feeding phase | ||||
| In a substitution phase, an equivalent quantity of fat from peanuts substituted for fat in the diet for 8 weeks. Peanuts consumed at average of 89 ± 21 g/day equivalent to 2113 ± 494 kJ/day (505 ± 118 kcal/day) during the 3 treatment phases. 50% dietary fat energy provided by peanuts. | ↑ resting energy expenditure by 11% after adjustment for changes in body weight after peanut consumption for 19 weeks (p < 0.01) | ||||
| Wien et al., 2003 [26] | 65 overweight and obese subjects | Randomised prospective/24 weeks | Almonds | Formula-based low-calorie diet enriched with 84 g/day of almonds vs. complex carbohydrates matching calories and protein | ↓ 62% body weight/BMI, |
| ↓ 50% waist circumference, | |||||
| ↓ 56% greater reduction in fat mass compared with the complex carbohydrate diet | |||||
| ↓ glucose, ↓ insulin, ↓ HOMA-IR,↓ TC, ↓ TG, ↓ LDL-C, ↓ LDL-C/HDL-C in both diets | |||||
| Pelkman et al., 2004 [80] | 53 overweigh and obese subjects | Parallel-arm/10 weeks (6 weeks for weight loss and additional 4-week for weigh maintenance) | Peanuts | Moderate-fat (33% of energy) diet vs. low-fat (18% of energy) diet | ↓ 1.2 ± 0.05 kg/week body weight within the moderate-fat group |
| ↓ 1.09 ± 0.06 kg/week body weight within the low-fat diet groups | |||||
| No difference in weight loss between two groups. | |||||
| ↔ HDL-C, ↓ triacylglycerol, | |||||
| non-HDL C/HDL-C within the moderate-fat diet group for 6 weeks | |||||
| ↑ triacylglycerol, ↓ HDL-C, ↔ non-HDL-C/HDL-C within the low-fat diet group for 6 weeks | |||||
| Sabate et al., 2005 [66] | 90 healthy subjects | Randomized cross-over/6 months | Walnuts | 28–56 g/day walnut-supplementation (12% energy intake) vs. control diet without nuts | ↑ energy intake (133 kcal), ↑ 0.4 kg body weight |
| Kocyigit et al., 2006 [71] | 44 health subjects | Randomised parallel arm/3 weeks | Pistachios | 20% energy of regular diet replaced with pistachios vs. regular diet | ↔ body weight, ↔ energy intake |
| ↓ TC, ↓ MDA, ↓ TC/HDL-C (p < 0.001) | |||||
| ↓ LDL-C/HDL-C (p < 0.01), ↑ HDL-C (p < 0.001), ↑AOP, ↑AOP/MDA | |||||
| Hollis et al., 2007 [51] | 20 healthy female subjects | Randomised crossover, no dietary advice/10 weeks | Almonds | 1440 kJ portion of raw, unsalted almonds added to a habitual diet vs. habitual diet | ↔ body weight, ↔ energy expenditure |
| Wang et al., 2012 [73] | 90 subjects with metabolic syndrome | Randomised parallel arm, dietary advice/12 weeks | Pistachios | 42 g/day pistachios plus AHA Step 1 diet vs. 70 g/day pistachio plus AHA Step 1 diet vs. AHA step 1 diet | ↔ body weight,↔ fasting glucose, ↔ 2-h postprandial glucose among treatments |
| ↓ 2-h postprandial glucose within the pistachio groups | |||||
| ↔ TC, ↔ TG (p = 0.018), ↔ LDL-C, ↔HDL-C | |||||
| Holligan et al., 2014 [74] | 28 subjects with elevated LDL levels | Randomised, cross-over, controlled-/4 weeks | Pistachios | 20% energy of a diet replaced with 63–126 g/day of pistachios vs. 10% energy of a diet replaced with 32–63 g/day of pistachios vs. low-fat control diet | ↔ body weight, ↔ energy intake |
| ↓ small, dense LDL-C levels compared with the low-fat control diet. | |||||
| ↓ triacylglycerol/HDL-C on 63–126 g/day of pistachio diet compared with the control diet | |||||
| Razquin et al., 2017 [78] | 4242 subjects | Randomised controlled/3 years | Walnuts, Almonds, Hazelnuts | Mediterranean diets enriched with 30 g/day of nuts (15 g/day walnuts, 7.5 g/day hazelnuts, and 7.5 g/day almonds) vs. Mediterranean diets enriched with 50 mL/day of extra virgin olive oil vs. low-fat diet | ↓ body weight |
| (↓ ~5 kg in the two lowest quintiles of energy density | |||||
| ↓ ~4 kg the highest quintile of energy density | |||||
| ↓ ~2 kg in the two middle quintiles of energy density) |
AOP, antioxidant potential index; AHA, American Heart Association; BMI, body mass index; BP, blood pressure; HDL-C, high-density lipoprotein cholesterol; LDL-C, low density lipoprotein; MDA, malondialdehyde; MUFA, monounsaturated fatty acids; PAI-1, plasminogen activator inhibitor-1; SFA, saturated fatty acid; TC, total cholesterol; TG, triglyceride; tPA, tissue plasminogen activator; ↑, increase; ↓, decrease; ↔, no effect.
Table 4.
Summary of clinical trials examining the effect of nut consumption on oxidative stress.
| Reference | Subjects | Study Design/Period | Nut Type | Diet Intervention | Results |
|---|---|---|---|---|---|
| Durak et al., 1999 [77] | 30 healthy subjects | Pre-post supplemental, dietary advice | Hazelnuts | 1 g/kg body weight/day of hazelnuts supplemented with a habitual diet vs. baseline | ↑ 0.5 kg body weight |
| ↓ TC (p < 0.005) , ↓ LDL-C (p < 0.0005), ↑ HDL-C, ↑ TG (p< 0.001), ↑ HDL-C/LDL-C, (p < 0.0005) | |||||
| ↓ MDA(p < 0.0005), ↑ AOP (p < 0.0005) after hazelnut supplementation | |||||
| Kocyigit et al., 2006 [71] | 44 health subjects | Randomised parallel arm/3 weeks | Pistachios | 20% energy of regular diet replaced with pistachio vs. regular diet | ↔ body weight, ↔ energy intake |
| ↓ TC, ↓ MDA, ↓ TC/HDL-C (p < 0.001) | |||||
| ↓ LDL-C/HDL-C (p < 0.01), ↑ HDL-C (p < 0.001), ↑ AOP, ↑ AOP/MDA | |||||
| Jia et al., 2006 [205] | 30 healthy adult male regular smokers | Parallel arm, control/4 weeks | Almonds | 84 g/day almond group (n = 10) vs. 168 g/day almond group (n = 10) vs. Control group without nuts (n = 10) | ↓ urinary 8-OH-dG, ↓ single strand DNA breaks, |
| ↓MDA, ↔ SOD, ↔ GSH-Px in almond groups compared with the control group | |||||
| Haddad et al., 2006 [212] | 24 healthy subjects | Randomized, controlled, crossover/4 weeks | Pecans | 20% energy of a diet replaced with pecans vs. a control diet without nuts | ↓ MDA, ↔ferric-reducing ability, ↔Trolox equivalent antioxidant capacity |
| Davis et al., 2007 [206] | 65 subjects with metabolic syndrome | Randomised, parallel arm, control/8 weeks | Walnuts | 20% energy of a habitual diet replaced with walnuts (63–108 g/day) vs. 20% energy of a habitual diet replaced with cashews (63–108 g/day) vs. Control diet without nuts | ↓ GSSG, ↑ ORAC compared with baseline |
| Garg et al., 2007 [207] | 17 male hypercholesterolemic subjects | Pre-post supplemental/4 weeks | Macadamias | 15% energy of a habitual diet replaced with macadamias (40–90 g/day) vs. baseline | ↓ oxidative status (8-isoprostane) |
| ↓ inflammation (leukotriene, LTB4) | |||||
| ↔ TXB2/PGI2 ratio | |||||
| Li et al., 2007 [208] | 60 healthy male habitual smokers | Randomized, crossover/4 weeks | Almonds | Smokers; 84 g/day almonds vs. 120 g/day of pork (for smokers) | ↑ serum a-tocopherol, ↑ SOD, ↑ GPX, |
| 30 healthy male | ↔ caltalase after almond intake compared with no change in smokers after pork intake | ||||
| non-smokers | Non-smokers; 120 g/day of pork for reference comparison | ↓ DNA strand breaks, ↓ 8-OHdG, ↓ MDA, in smokers after almond intake compared with baseline | |||
| Jenkins et al., 2008 [209] | 27 hyperlipidemic subjects‘ | Randomized, crossover/4 weeks | Almonds | 73 g/day almonds added to self-selected low-fat therapeutic diet vs. 36.5 g/day almonds added to self-selected low-fat therapeutic diet vs. self-selected low-fat therapeutic diet without nuts (control) | ↓ MDA, ↓ urinary isoprostane on 73 g/day almonds compared with control |
| ↔ α-or γ-tocopherol, | |||||
| Thomson et al., 2008 [203] | 59 healthy subjects | Randomized controlled/12 weeks | Brazil nuts | 2 Brazil nuts (providing ≈100 μg Se, 100 μg Se as selenomethionine) vs. Placebo | ↑ selenium (p < 0.0001), ↑ GPx (p < 0.001) |
| McKay et al., 2010 [213] | 21 non-smoking man and postmenopausal women aged over 50 years | Randomized crossover/6 weeks | Walnuts | 42 g/day walnuts vs. 21 g/day walnuts | ↔ antioxidant activity (ORAC, ORAC with perchloric acid (pca) precipitation, FRAP and TAP), ↔ biomarkers of antioxidant status (total thiols, phenols, carotenoids and GPx), ↔ MDA |
| Lopez-Uriarte et al., 2010 [121] | 50 subjects with metabolic syndrome | Randomised, controlled, parallel/4 weeks | Mixed nuts | 30 g/day mix nuts (15 g walnuts, 7.5 g almonds and 7.5 g hazelnuts) vs. control diet without nuts | ↓ DNA damage (p < 0.001), |
| ↔ Antioxidant capacity, ↔oxidized LDL, ↔ conjugated diene, | |||||
| ↔ 8-iso-prostanes, ↔ endothelial function | |||||
| Kay et al., 2010 [210] | 28 hypercholesterolemic subjects | Randomized, crossover controlled/4 weeks | Pistachios | 20% energy of a low-fat diet replaced with 63–126 g/day pistachios vs. 20% energy of a low-fat diet replaced with 32–63 g/day pistachios vs. a low-fat control diet without pistachios | ↓ oxidized-LDL compared with a low-fat control diet |
| ↑ lutein (p < 0.0001), ↑ α-carotene, ↑ β-carotene (p < 0.01)compared with baseline | |||||
| Sari et al., 2010 [24] | 32 healthy young men | Dietary intervention/8 weeks | Pistachios | Subjects consumed a Mediterranean diet for 4 weeks and in the next 4 weeks they supplemented the Mediterranean diet with pistachios for 4 weeks replacing 32% of the energy obtained from MUFAs in the control diet (20% of energy) | ↓ glucose levels (−8.8 ± 8.5% p < 0.001), |
| ↓ TC ( p < 0.001) ↓ LDL-C (p < 0.001), ↓ triacylglycerol, ↔ HDL-C, ↓ TC-C/HDL-C ↓ LDL-C/HDL-C ( p < 0.001), | |||||
| ↑ endothelium-dependent vasodilation (p = 0.002) | |||||
| ↓ lipid hydroperoxide (p < 0.001), | |||||
| ↓ MDA (p < 0.001), ↓ IL-6 (p < 0.001), | |||||
| ↑ SOD (p < 0.001), ↔ hs CRP, ↔ TNF-α | |||||
| compared with a Mediterranean diet | |||||
| Haddad et al., 2014 [211] | healthy young subjects. | Randomized, crossover, controlled, acute 5-h test | Walnuts | A walnut meal containing 90 g walnuts and a isocaloric control meal (50% carbohydrate; 20% protein; 30% fat) | ↓ oxidative stress, ↑ iAUC for hydrophilic and lipophilic ORAC, ↓ iAUC for MDA |
| ↓ oxidized LDL, ↑ epicatechin gallate | |||||
| ↑ epicallocatechin gallate, ↑ urolithin-A in urine |
AOP, antioxidant potential index; 8-OH-dG, urinary 8-hydroxy-2′-deoxyguanosine; FRAP, ferric reducing antioxidant power; GSH-Px, glutathione peroxidase; GSSG, oxidized glutathione; MDA, malondialdehyde; ORAC, oxygen radical absorbance capacity; PGI2, prostacyclin; SOD, superoxide dismutase; TAP, total antioxidant performance; TXB2, plasma thromboxane; ↑, increase; ↓, decrease; ↔, no effect.
Table 5.
Summary of clinical trials examining the effect of nut consumption on inflammation.
| Reference | Subjects | Study Design/Period | Nut Type | Diet Intervention | Results |
|---|---|---|---|---|---|
| Zhao et al., 2004 & 2007 [222,223] | 23 hypercholesterolemic subjects | Randomized, controlled cross-over/6 weeks | Walnuts | Three diets; - AAD—13% energy from SFA, 13% energy from MUFAs and 8.7% energy from PUFAs (7.7% LA; 0.8% ALA) - The LA diet consisted of 16.4% energy from PUFAs (12.6% LA; 3.6% ALA) - The ALA diet consisted of 17% energy from PUFAs (10.5% LA; 6.5% ALA). Walnuts and walnut oil were used for half of total fat in the LA and ALA diets. The ratios of LA (n-6) to ALA (n-3) for the AAD, LA and ALA diet were 10:1, 4:1, and 2:1 | ↓ hs-CRP (p < 0.01) on ALA diet compared with the AAD |
| ↓ IL-6, ↓ IL-1β, ↓ TNF-α cultured in PBMCs on the ALA diet than on the LA diet | |||||
| ↓ TC, ↓ TG, ↓ LDL-C ↓ ICAM-1 on both ALA and LA diets compared with AAD diet | |||||
| ↓ VCAM-1 (p < 0.01), ↓ E-selectin (p < 0.01) on the ALA diet than on the LA diet | |||||
| ↓ HDL-C and apolipoprotein AI on ALA diet compared with AAD diet | |||||
| Garg et al., 2007 [207] | 17 male hypercholesterolemic subjects | Pre-post supplemental/4 weeks | Macadamias | 15% energy of a habitual diet replaced with macadamias (40–90 g/day) vs. baseline | ↓ oxidative status (8-isoprostane) |
| ↓ inflammation (leukotriene, LTB4) | |||||
| ↔ TXB2/PGI2 ratio | |||||
| Rajaram et al., 2010 [224] | 25 health subjects | Randomised, controlled, crossover/4 weeks | Almonds | High-almond diet (20% energy of a control diet replaced with almonds) vs. Low-almond diet (10% energy of a control diet replaced with almonds) vs. Heart-healthy control diet without nuts | ↓ hs -CRP ↓ E-selectin (p < 0.0001) |
| Sari et al., 2010 [24] | 32 healthy young men | Dietary intervention/8 weeks | Pistachios | Subjects consumed a Mediterranean diet for 4 weeks and in the next 4 weeks they supplemented the Mediterranean diet with pistachios for 4 weeks replacing 32% of the energy obtained from MUFAs in the control diet (20% of energy) | ↓ glucose levels (−8.8 ± 8.5% p < 0.001) |
| ↓ TC, ↓ LDL-C, ↓ triacylglycerol, ↔ HDL-C, ↓ TC-C/HDL-C ↓ LDL-C/HDL-C | |||||
| ↑ endothelium-dependent vasodilation | |||||
| ↓ lipid hydroperoxide, ↓ MDA, ↓ IL-6, | |||||
| ↑ SOD, ↔ hs CRP, ↔ TNF-α | |||||
| compared with a Mediterranean diet | |||||
| Hernandez-Alonso et al., 2014 [225] | 54 subjects with prediabetes | Randomized, controlled cross-over/4 months | Pistachios | A diet supplemented with 57 g/day pistachio vs. a control diet | ↓ IL-6 mRNA, ↓ resistin gene expression |
| ↑ SLC2A4 expression | |||||
| ↓ fasting glucose, ↓ insulin, ↓ HOMA-IR | |||||
| ↓ fibrinogen, ↓ oxidized LDL, ↓ platelet factor 4 | |||||
| ↑ GLP-1 | |||||
| Sauder et al., 2015 [22] | 30 subjects with well-controlled T2DM | Randomized, crossover, controlled/4 weeks | Pistachios | Healthy diet with pistachios contributing 20% of total energy. Low-fat or fat-free snacks (e.g., pretzels) in the control diet were substituted with roasted pistachios providing 20% of daily energy (59 to 128 g/day of pistachios). Half of amount of pistachio was ingested unsalted vs. AHA Therapeutic Lifestyle Changes diet (26.9% total fat, 6.7% SFA, 186 mg/day cholesterol). | ↓ fructosamine, ↓ TC, ↓ ratio of TC to HDL-C, ↓ TG (p = 0.003), ↔ HbA1c, |
| ↔ fasting glucose, ↔ insulin, ↔ hs-CRP, | |||||
| ↔ ICAM, ↔ VCAM, ↔endothelia function | |||||
| Arpon et al., 2017 [221] | 36 subjects at high cardiovascular risks | Randomised, controlled, parallel/5 years | Mixed nuts | Mediterranean diet supplemented with nuts (30 g/day of nuts) vs. Mediterranean diet supplemented with extra virgin olive oil (1 L/week of virgin olive oil) vs. Low-fat diet (control) | ↑ association between methylation changes of 8 genes (EEF2, COL18A1, IL4I1, LEPR, PLAGL1, IFRD1, MAPKAPK2, and PPARGC1B) related to inflammation and Mediterranean diets ↑ association between EEF2 methylation and TNF-α and hs-CRP on Mediterranean diets compared with baselines. |
| Gulati et al., 2017 [226] | 52 subjects with T2DM | Pre-post supplemental study, dietary advice, physical activity (45 min of walking at least 5 days some week)/24 weeks | Almonds | 20% of total energy of a diet replaced with almonds vs. baseline | ↓ hs-CRP (p < 0.01), |
| ↓ waist circumference, | |||||
| ↓ waist-to-height ratio, | |||||
| ↓ TC, ↓ TG, ↓ LDL-C, ↓ HbA1c |
AAD, average American diet; ALA, α-linolenic acid; GLP-1, glucagon-like peptide-1; HbA1c, glycosylated haemoglobin; HDL-C, high density lipoprotein cholesterol; HOMA-IR, homeostasis model assessment of insulin resistance; hs-CRP, high sensitivity C-reactive protein; ICAM-1, intercellular cell adhesion molecule-1; LA, linoleic acid; LDL-C, low density lipoprotein cholesterol; MUFA, monounsaturated fatty acids; PBMC, peripheral blood mononuclear cell; PUFA, polyunsaturated fatty acids; TC, total cholesterol; TG, triglyceride; VCAM-1, vascular cell adhesion molecule-1; ↑, increase; ↓, decrease; ↔, no effect.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, Y.; Keogh, J.B.; Clifton, P.M. Benefits of Nut Consumption on Insulin Resistance and Cardiovascular Risk Factors: Multiple Potential Mechanisms of Actions. Nutrients 2017, 9, 1271. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9111271
AMA Style
Kim Y, Keogh JB, Clifton PM. Benefits of Nut Consumption on Insulin Resistance and Cardiovascular Risk Factors: Multiple Potential Mechanisms of Actions. Nutrients. 2017; 9(11):1271. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9111271
Chicago/Turabian StyleKim, Yoona, Jennifer B. Keogh, and Peter M. Clifton. 2017. "Benefits of Nut Consumption on Insulin Resistance and Cardiovascular Risk Factors: Multiple Potential Mechanisms of Actions" Nutrients 9, no. 11: 1271. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9111271
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.