Intravenous Arginine Administration Promotes Proangiogenic Cells Mobilization and Attenuates Lung Injury in Mice with Polymicrobial Sepsis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Procedures
2.3. Flow Cytometric Analysis Of Proangiogenic Cells in Blood
2.4. Measurements of Proangiogenic Cell-Mobilizing Factors in Plasma
2.5. Measurement of Cytokines in Lung Tissue
2.6. Angpt1, Angpt2, and Tie-2 Messenger (m)RNA Extraction and Analysis by Quantitative Real-Time Reverse-Transcription (RT) Polymerase Chain Reaction (PCR)
2.7. Histopathology of Lung Tissue
2.8. Statistical Analysis
3. Results
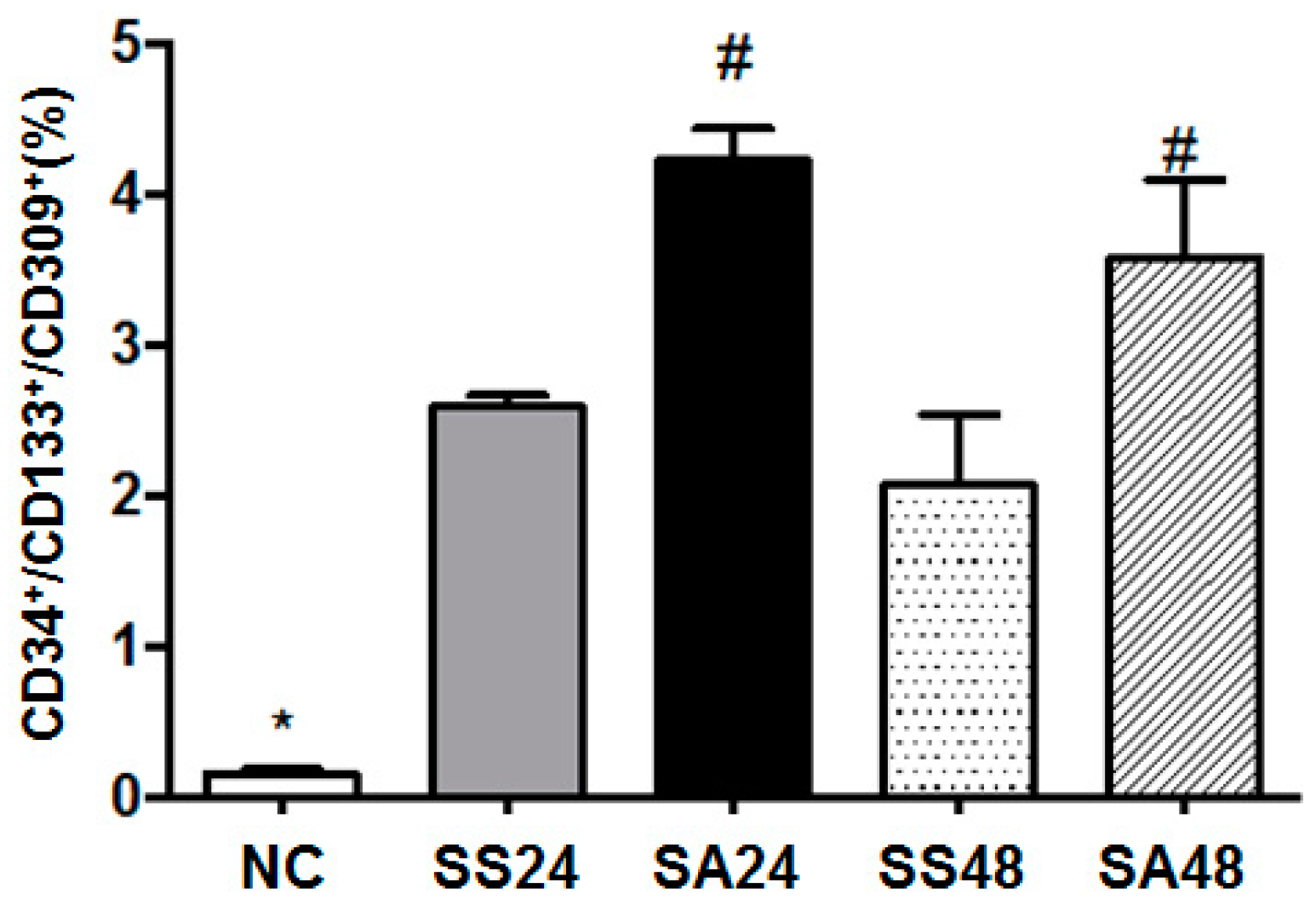
3.1. Differences in Proangiogenic Cell Population in Blood after CLP
3.2. Expression of Proangiogenic Cell-Related Proteins in Plasma
3.3. Concentration of Cytokines in Lung Homogenates
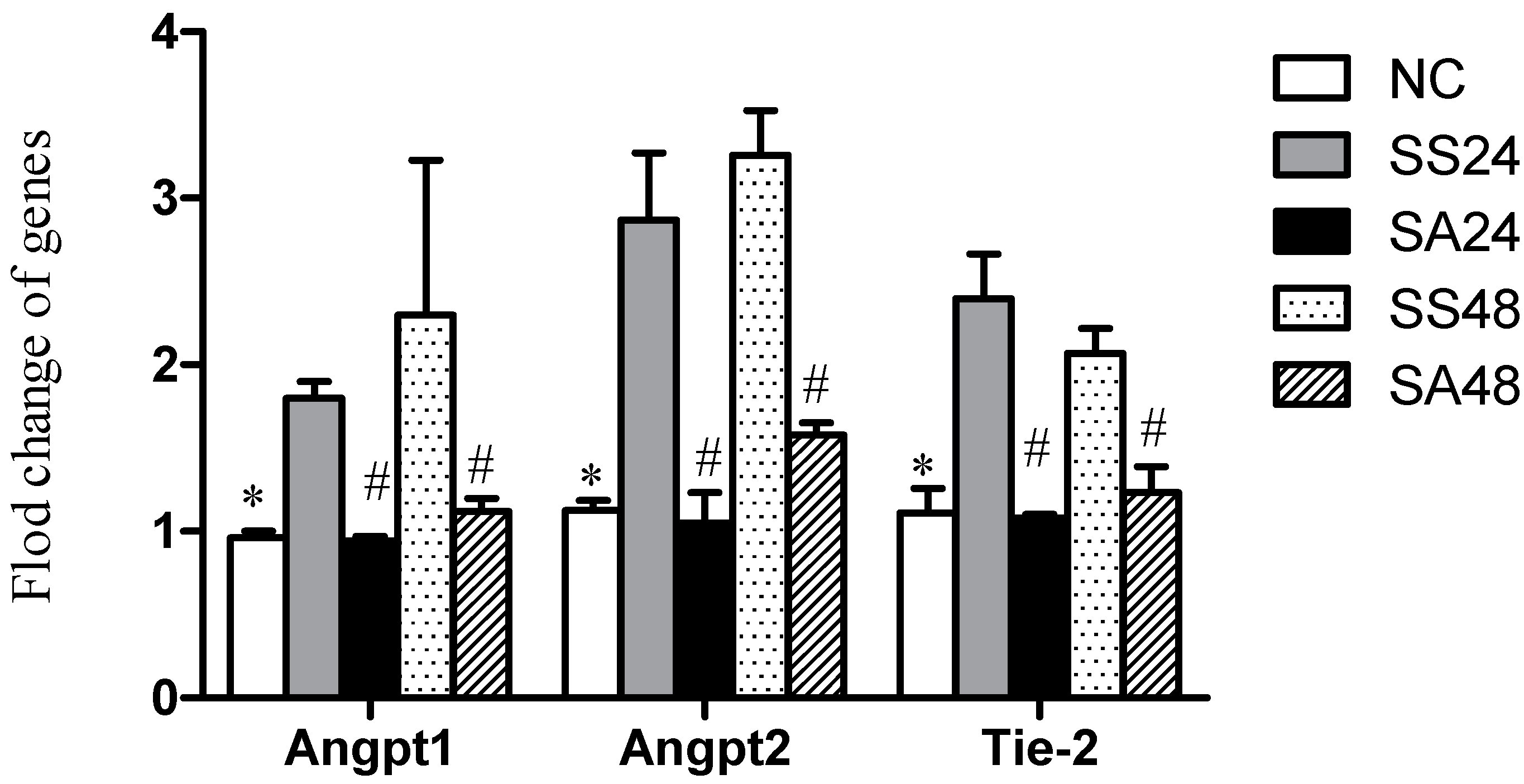
3.4. Expression of Angpt-1, Angpt-2 and Tie-2 mRNAs in Lung Tissues
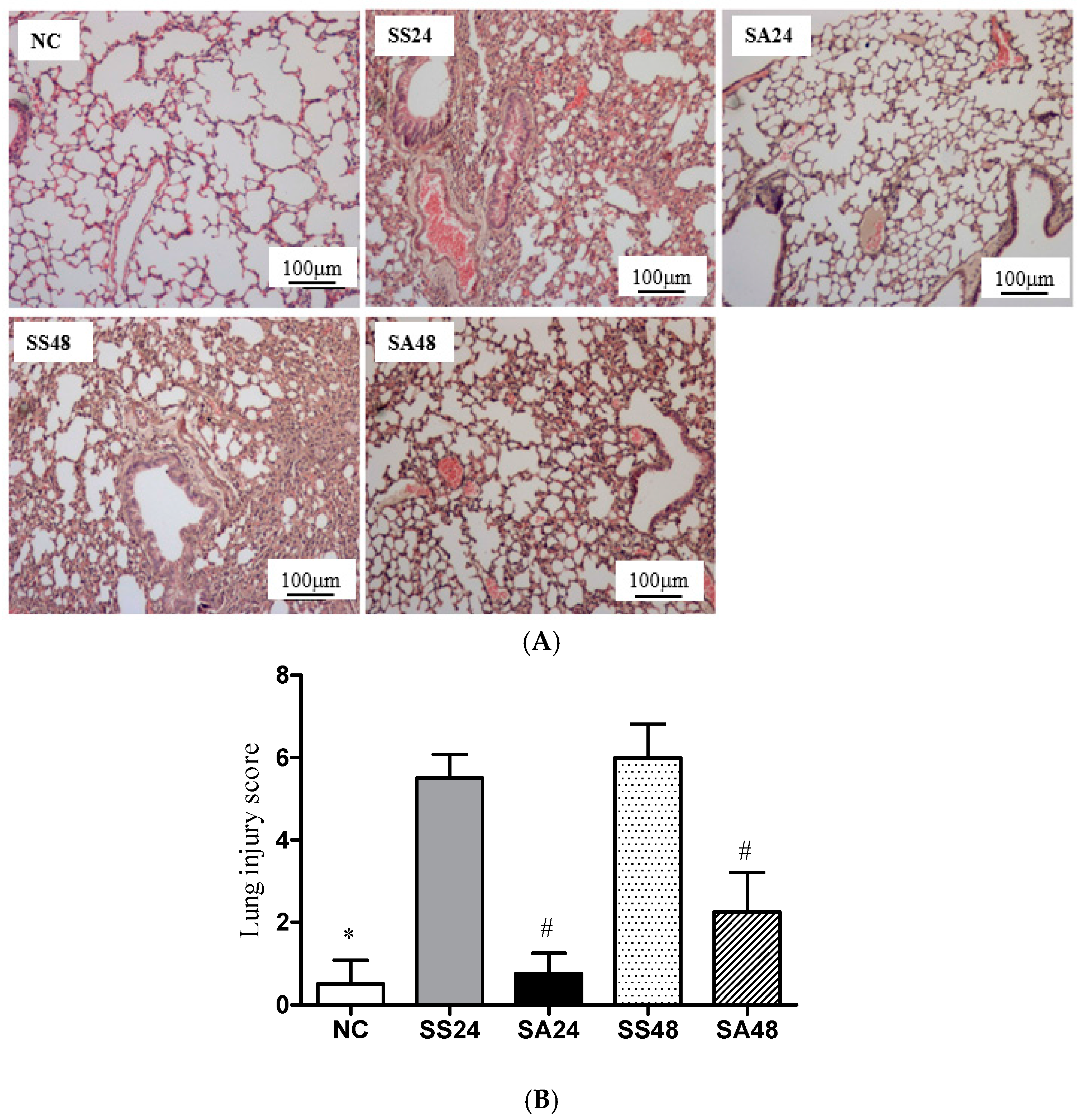
3.5. Lung Histology
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gustot, T. Multiple organ failure in sepsis: Prognosis and role of systemic inflammatory response. Curr. Opin. Crit. Care 2011, 17, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Boisrame-Helms, J.; Kremer, H.; Schini-Kerth, V.; Meziani, F. Endothelial dysfunction in sepsis. Curr. Vasc. Pharmacol. 2013, 11, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak, S.; Cinel, I.; Phillip Dellinger, R.; Shapiro, N.I.; Arnold, R.C.; Parrillo, J.E.; Hollenberg, S.M. Microcirculatory alterations in resuscitation and shock (mars) investigators. Resuscitating the microcirculation in sepsis: The central role of nitric oxide, emerging concepts for novel therapies, and challenges for clinical trials. Acad. Emerg. Med. 2008, 15, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Skibsted, S.; Jones, A.E.; Puskarich, M.A.; Arnold, R.; Sherwin, R.; Trzeciak, S.; Schuetz, P.; Aird, W.C.; Shapiro, N.I. Biomarkers of endothelial cell activation in early sepsis. Shock 2013, 39, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Volk, T.; Kox, W.J. Endothelium function in sepsis. Inflamm. Res. 2000, 49, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yang, J.; Li, N.; Wu, R.; Tian, H.; Song, H.; Wang, H. Role of endothelial progenitor cell transplantation in rats with sepsis. Transplant. Proc. 2015, 47, 2991–3001. [Google Scholar] [CrossRef] [PubMed]
- Adams, V.; Lenk, K.; Linke, A.; Lenz, D.; Erbs, S.; Sandri, M.; Tarnok, A.; Gielen, S.; Emmrich, F.; Schuler, G.; et al. Increase of circulating endothelial progenitor cells in patients with coronary artery disease after exercise-induced ischemia. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Malli, F.; Koutsokera, A.; Paraskeva, E.; Zakynthinos, E.; Papagianni, M.; Makris, D.; Tsilioni, I.; Molyvdas, P.A.; Gourgoulianis, K.I.; Daniil, Z. Endothelial progenitor cells in the pathogenesis of idiopathic pulmonary fibrosis: an evolving concept. PLoS ONE 2013, 8, e53658. [Google Scholar] [CrossRef] [PubMed]
- Marrotte, E.J.; Chen, D.D.; Hakim, J.S.; Chen, A.F. Manganese superoxide dismutase expression in endothelial progenitor cells accelerates wound healing in diabetic mice. J. Clin. Investig. 2010, 120, 4207–4219. [Google Scholar] [CrossRef] [PubMed]
- Rafat, N.; Hanusch, C.; Brinkkoetter, P.T.; Schulte, J.; Brade, J.; Zijlstra, J.G.; van der Woude, F.J.; van Ackern, K.; Yard, B.A.; Beck, G.C. Increased circulating endothelial progenitor cells in septic patients: Correlation with survival. Crit. Care Med. 2007, 35, 1677–1684. [Google Scholar] [CrossRef] [PubMed]
- Duong, H.; Erzurum, S.; Asosingh, K. Pro-angiogenic hematopoietic progenitor cells and endothelial colony forming cells in pathological angiogenesis of bronchial and pulmonary circulation. Angiogenesis 2011, 14, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.J.; Miller, K.R.; Rosenthal, C.; Rosenthal, M.D. When is it appropriate to use arginine in critical illness? Nutr. Clin. Pract. 2016, 31, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Evoy, D.; Lieberman, M.D.; Fahey, T.J., 3rd; Daly, J.M. Immunonutrition: The role of arginine. Nutrition 1998, 14, 611–617. [Google Scholar] [CrossRef]
- Casas-Rodera, P.; Gomez-Candela, C.; Benitez, S.; Mateo, R.; Armero, M.; Castillo, R.; Culebras, J.M. Immunoenhanced enteral nutrition formulas in head and neck cancer surgery: A prospective, randomized clinical trial. Nutr. Hosp. 2008, 23, 105–110. [Google Scholar] [PubMed]
- Acquaviva, R.; Lanteri, R.; Li Destri, G.; Caltabiano, R.; Vanella, L.; Lanzafame, S.; Di Cataldo, A.; Li Volti, G.; Di Giacomo, C. Beneficial effects of rutin and l-arginine coadministration in a rat model of liver ischemia-reperfusion injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G664–G670. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336 Pt 1, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ge, K.; Lu, S.L.; Qing, C.; Xie, T.; Rong, L.; Niu, Y.W.; Wang, M.J.; Liao, Z.J.; Shi, J.X. The influence of l-arginine on the angiogenesis in burn wounds in diabetic rats. Chin. J. Burns 2004, 20, 210–213. [Google Scholar]
- Evrard, S.M.; d’Audigier, C.; Mauge, L.; Israel-Biet, D.; Guerin, C.L.; Bieche, I.; Kovacic, J.C.; Fischer, A.M.; Gaussem, P.; Smadja, D.M. The profibrotic cytokine transforming growth factor-beta1 increases endothelial progenitor cell angiogenic properties. J. Thromb. Haemost. 2012, 10, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Moss, A. The angiopoietin: Tie 2 interaction: a potential target for future therapies in human vascular disease. Cytokine Growth Factor Rev. 2013, 24, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.H.; Shih, Y.M.; Shih, J.M.; Yeh, C.L. Glutamine administration modulates endothelial progenitor cell and lung injury in septic mice. Shock 2016, 46, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Vaschetto, R.; Kuiper, J.W.; Chiang, S.R.; Haitsma, J.J.; Juco, J.W.; Uhlig, S.; Plotz, F.B.; Corte, F.D.; Zhang, H.; Slutsky, A.S. Inhibition of poly(adenosine diphosphateribose) polymerase attenuates ventilator-induced lung injury. Anesthesiology 2008, 108, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Luiking, Y.C.; Poeze, M.; Deutz, N.E. Arginine infusion in patients with septic shock increases nitric oxide production without haemodynamic instability. Clin. Sci. 2015, 128, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, M.D.; Carrott, P.W.; Patel, J.; Kiraly, L.; Martindale, R.G. Parenteral or enteral arginine supplementation safety and efficacy. J. Nutr. 2016, 146, 2594S–2600S. [Google Scholar] [CrossRef] [PubMed]
- Lucas, M.L.; Rhoden, C.R.; Rhoden, E.L.; Zettler, C.G.; Mattos, A.A. Effects of l-Arginine and l-NAME on ischemia-reperfusion in rat liver. Acta Cir. Bras. 2015, 30, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Braulio, V.B.; Ten Have, G.A.; Vissers, Y.L.; Deutz, N.E. Time course of nitric oxide production after endotoxin challenge in mice. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E912–E918. [Google Scholar] [CrossRef] [PubMed]
- Lortie, M.J.; Satriano, J.; Gabbai, F.B.; Thareau, S.; Khang, S.; Deng, A; Pizzo, D.P.; Thomson, S.C.; Blantz, R.C.; Munger, K.A. Production of arginine by the kidney is impaired in a model of sepsis: Early events following LPS. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R1434–R1440. [Google Scholar] [CrossRef] [PubMed]
- Cepinskas, G.; Wilson, J.X. Inflammatory response in microvascular endothelium in sepsis: Role of oxidants. J. Clin. Biochem. Nutr. 2008, 42, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Patschan, S.A.; Patschan, D.; Temme, J.; Korsten, P.; Wessels, J.T.; Koziolek, M.; Henze, E.; Muller, G.A. Endothelial progenitor cells (EPC) in sepsis with acute renal dysfunction (ARD). Crit. Care 2011, 15, R94. [Google Scholar] [CrossRef] [PubMed]
- George, A.L.; Bangalore-Prakash, P.; Rajoria, S.; Suriano, R.; Shanmugam, A.; Mittelman, A.; Tiwari, R.K. Endothelial progenitor cell biology in disease and tissue regeneration. J. Hematol. Oncol. 2011, 4, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Dery, M.A.; Michaud, M.D.; Richard, D.E. Hypoxia-inducible factor 1: Regulation by hypoxic and non-hypoxic activators. Int. J. Biochem. Cell Biol. 2005, 37, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Takahashi, T.; Masuda, H.; Kalka, C.; Chen, D.; Iwaguro, H.; Inai, Y.; Silver, M.; Isner, J.M. VEGF contributes to postnatal neovascularization by mobilizing bone marrow-derived endothelial progenitor cells. EMBO J. 1999, 18, 3964–3972. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.G.; Zhong, Z.Y.; Sun, G.F.; Zhou, Y.X.; Zhao, Y. Effects of tumour necrosis factor-alpha on activity and nitric oxide synthase of endothelial progenitor cells from peripheral blood. Cell Prolif. 2011, 44, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Duda, D.G.; Fukumura, D.; Jain, R.K. Role of eNOS in neovascularization: NO for endothelial progenitor cells. Trends Mol. Med. 2004, 10, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Thum, T.; Fraccarollo, D.; Schultheiss, M.; Froese, S.; Galuppo, P.; Widder, J.D.; Tsikas, D.; Ertl, G.; Bauersachs, J. Endothelial nitric oxide synthase uncoupling impairs endothelial progenitor cell mobilization and function in diabetes. Diabetes 2007, 56, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Tadié, J.M.; Cynober, L.; Peigne, V.; Caumont-Prim, A.; Neveux, N.; Gey, A.; Guerot, E.; Diehl, J.L.; Fagon, J.Y.; Tartour, E.; et al. Arginine administration to critically ill patients with a low nitric oxide fraction in the airways: A pilot study. Intensive Care Med. 2013, 39, 1663–1665. [Google Scholar] [CrossRef] [PubMed]
- Carraway, M.S.; Piantadosi, C.A.; Jenkinson, C.P.; Huang, Y.C. Differential expression of arginine and iNOS in the lung in sepsis. Exp. Lung. Res. 1998, 24, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Parikh, S.M. Dysregulation of the angiopoietin-Tie-2 axis in sepsis and ARDS. Virulence 2013, 4, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, U.; Scharpfenecker, M.; Koidl, S.; Hegen, A.; Grunow, V.; Schmidt, J.M.; Kriz, W.; Thurston, G.; Augustin, H.G. The Tie-2 ligand angiopoietin-2 is stored in and rapidly released upon stimulation from endothelial cell Weibel-Palade bodies. Blood 2004, 103, 4150–4156. [Google Scholar] [CrossRef] [PubMed]
- Senbel, A.M.; Omar, A.G.; Abdel-Moneim, L.M.; Mohamed, H.F.; Daabees, T.T. Evaluation of l-arginine on kidney function and vascular reactivity following ischemic injury in rats: Protective effects and potential interactions. Pharmacol. Rep. 2014, 66, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Wang, L.; Qian, L. The role of eNOS in the migration and proliferation of bone-marrow derived endothelial progenitor cells and in vitro angiogenesis. Cell Biol. Int. 2015, 39, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.M.; Yeh, C.L.; Pai, M.H.; Lee, W.Y.; Yeh, S.L. Glutamine administration modulates lung gammadelta T lymphocyte expression in mice with polymicrobial sepsis. Shock 2014, 41, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Rittirsch, D.; Huber-Lang, M.S.; Flierl, M.A.; Ward, P.A. Immunodesign of experimental sepsis by cecal ligation and puncture. Nat. Protoc. 2009, 4, 31–36. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| CXCL-12 | MMP-9 | VEGF | TNF-α | NO | |
|---|---|---|---|---|---|
| ng/mL | pg/mL | μmol/L | |||
| NC | 4.6 ± 0.3 * | 22.0 ± 1.4 * | 65.0 ± 6.8 * | 1.8 ± 0.6 * | 4.1 ± 0.5 * |
| SS24 | 18.0 ± 1.2 | 48.8 ± 4.7 | 92.1 ± 2.4 | 16.3 ± 1.9 | 11.3 ± 0.5 |
| SA24 | 17.1 ± 1.1 | 33.4 ± 4.0 # | 108.8 ± 1.0 # | 10.5 ± 0.8 # | 38.8 ± 1.2 # |
| SS48 | 21.2 ± 1.1 | 46.3 ± 2.0 | 112.6 ± 6.3 | 10.1 ± 1.9 | 8.9 ± 0.2 |
| SA48 | 27.9 ± 1.6 # | 30.6 ± 1.4 # | 130.1 ± 3.4 # | 4.96 ± 0.6 # | 15.7 ± 0.7 # |
| IL-1 | IL-6 | TNF-α | IL-10 | TGF-β1 | |
|---|---|---|---|---|---|
| pg/mg | |||||
| NC | 10.3 ± 1.2 * | 12.5 ± 2.1 * | 20.7 ± 2.3 * | 265.8 ± 17.9 | 16.4 ± 1.8 |
| SS24 | 92.6 ± 4.5 | 1275.6 ± 18.3 | 80.4 ± 3.4 | 235.8 ± 4.1 | 12.8 ± 1.6 |
| SA24 | 44.3 ± 2.6 # | 662.3 ± 13.5 # | 42.9 ± 1.9 # | 367.8 ± 9.2 # | 55.3 ± 2.6 # |
| SS48 | 83.2 ± 5.4 | 834.3 ± 17.3 | 67.8 ± 3.6 | 214.2 ± 5.6 | 16.6 ± 5.6 |
| SA48 | 32.3 ± 2.3 # | 385.4 ± 12.8 # | 38.8 ± 3.2 # | 229.4 ± 7.7 | 22.4 ± 3.4 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, C.-L.; Pai, M.-H.; Shih, Y.-M.; Shih, J.-M.; Yeh, S.-L. Intravenous Arginine Administration Promotes Proangiogenic Cells Mobilization and Attenuates Lung Injury in Mice with Polymicrobial Sepsis. Nutrients 2017, 9, 507. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9050507
Yeh C-L, Pai M-H, Shih Y-M, Shih J-M, Yeh S-L. Intravenous Arginine Administration Promotes Proangiogenic Cells Mobilization and Attenuates Lung Injury in Mice with Polymicrobial Sepsis. Nutrients. 2017; 9(5):507. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9050507
Chicago/Turabian StyleYeh, Chiu-Li, Man-Hui Pai, Yao-Ming Shih, Juey-Ming Shih, and Sung-Ling Yeh. 2017. "Intravenous Arginine Administration Promotes Proangiogenic Cells Mobilization and Attenuates Lung Injury in Mice with Polymicrobial Sepsis" Nutrients 9, no. 5: 507. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9050507