A High-Fructose-High-Coconut Oil Diet Induces Dysregulating Expressions of Hippocampal Leptin and Stearoyl-CoA Desaturase, and Spatial Memory Deficits in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Morris Water Maze (MWM) Task
2.3. Blood Biochemical Analysis, and Fasting Serum Insulin, Fructosamine, and Leptin Levels
2.4. Collection of Tissues
2.5. Fatty Acid Analysis of the Brain
2.6. Western Blot Analysis for Brain Leptin
2.7. Real-Time Reverse-Transcription Polymerase Chain Reaction (RT-PCR) for Leptin Receptor and SCD1 Genes
2.8. Statistical Analysis
3. Results
3.1. Blood Parameters and Body Weights
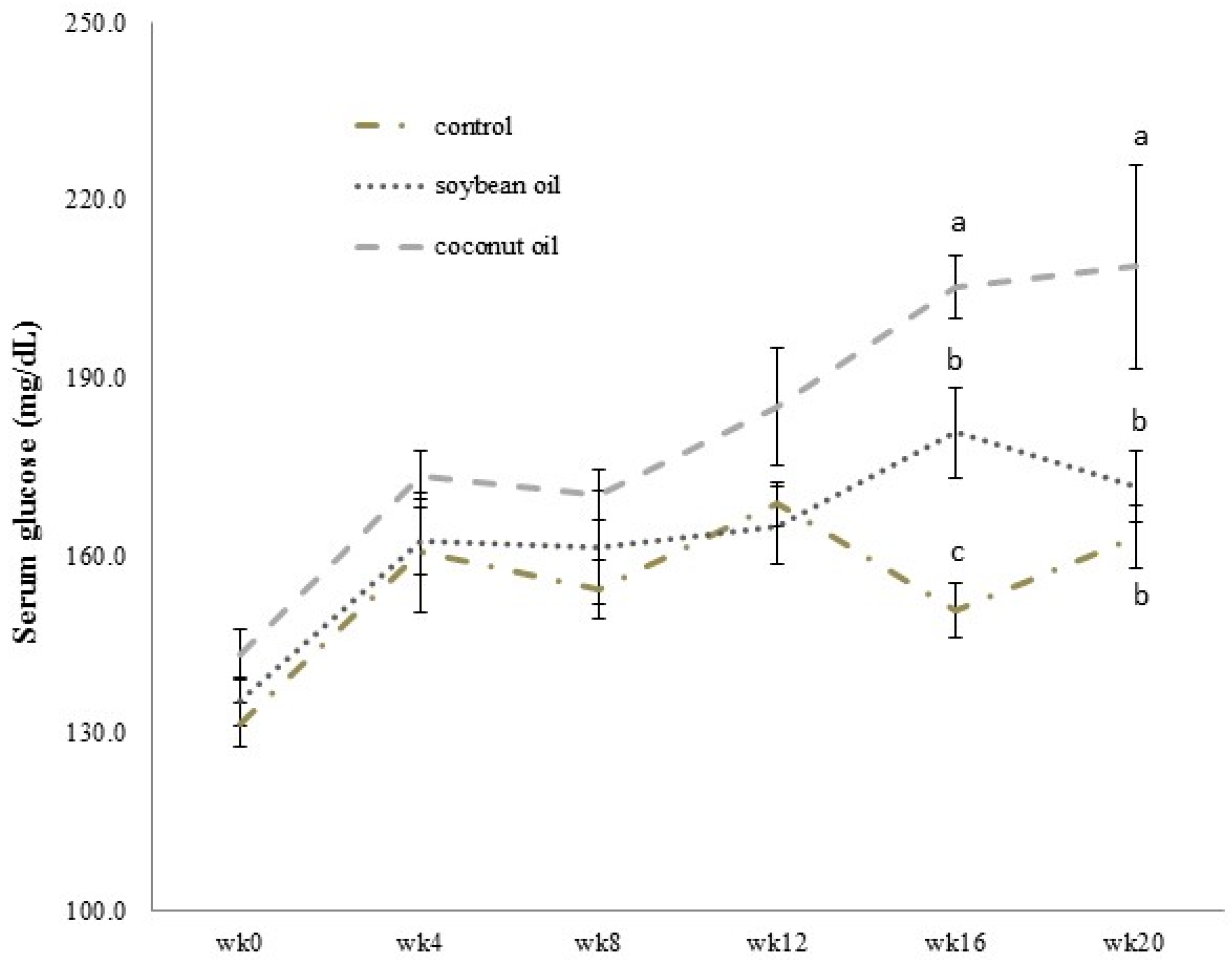
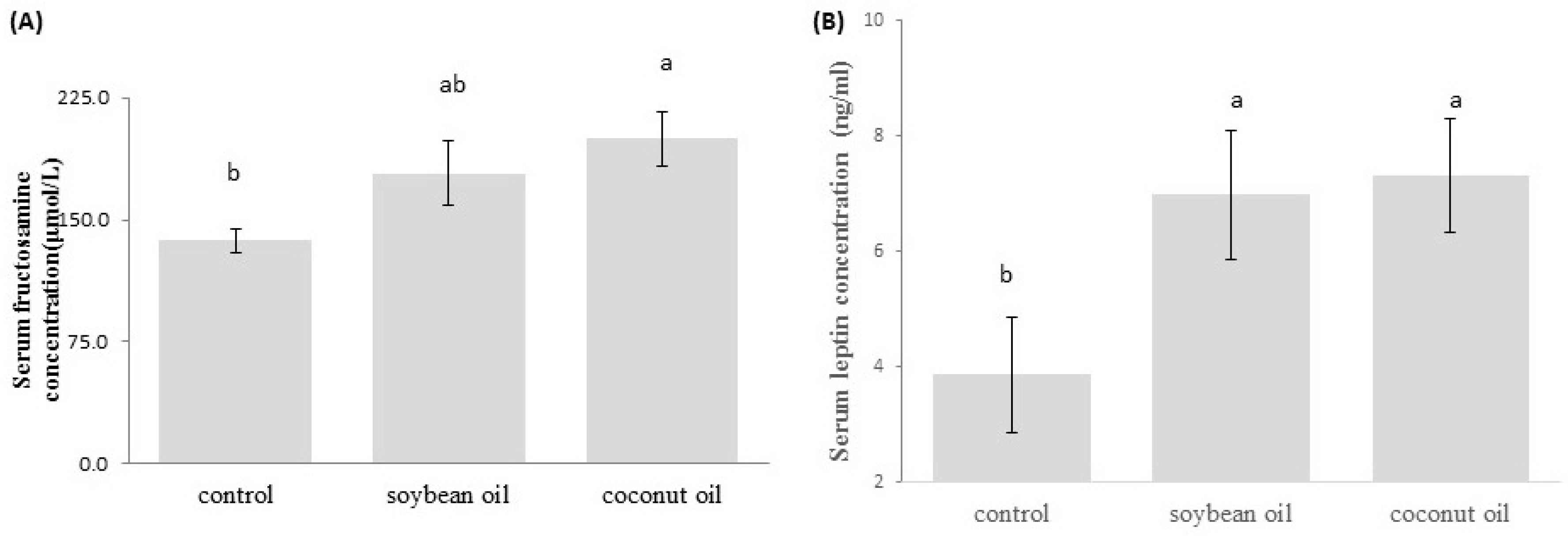
3.2. Levels of Blood Fasting Glucose, Fructosamine and Leptin
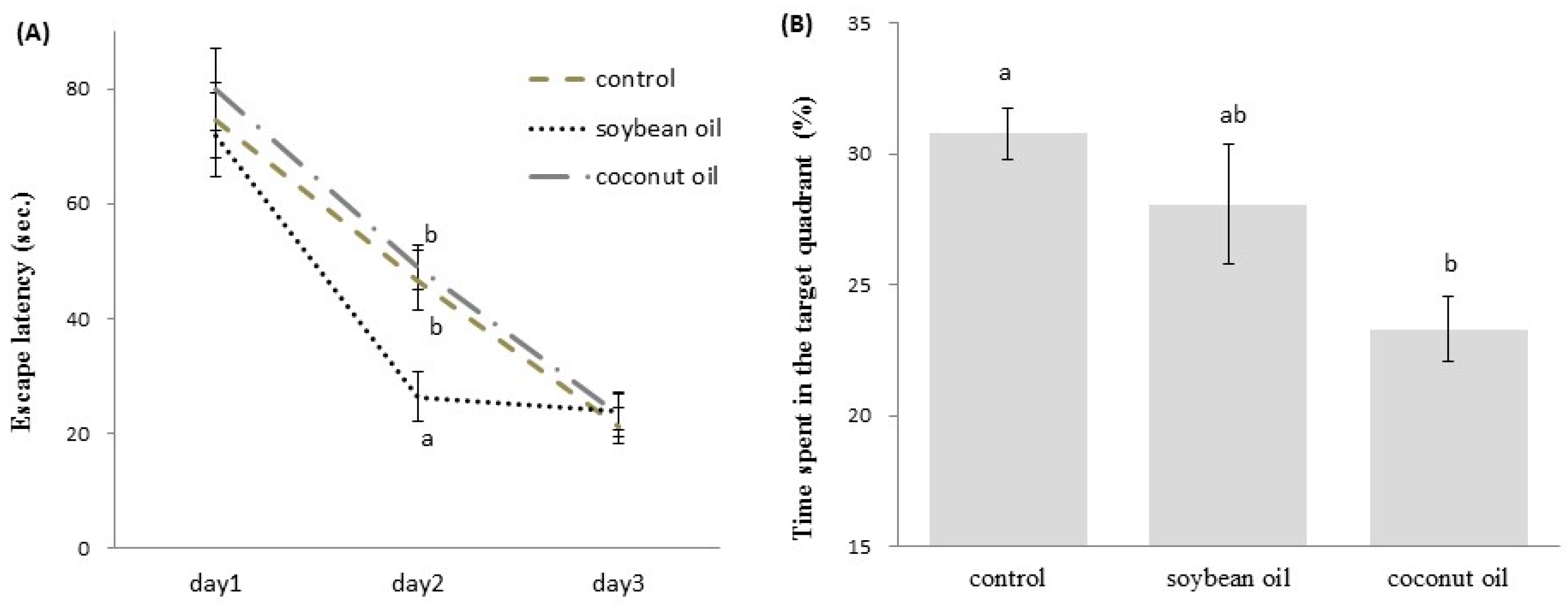
3.3. MWM Task for Spatial Learning and Memory
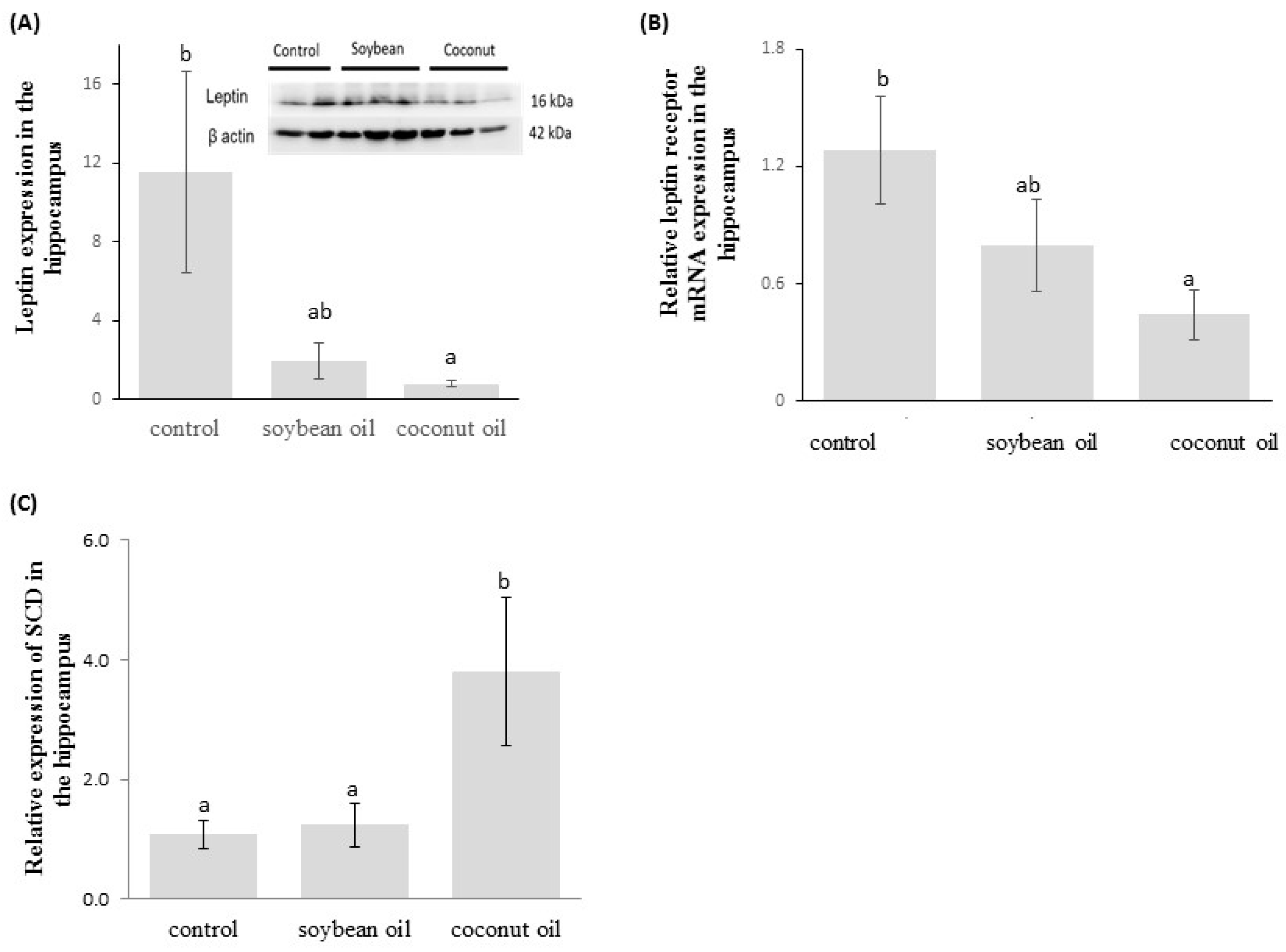
3.4. Levels of Hippocampal Leptin, and mRNA Expressions of Leptin Receptor and SCD1
3.5. Fatty Acid Composition in the Brain
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Phillips, C.M. Nutrigenetics and metabolic disease: Current status and implications for personalised nutrition. Nutrients 2013, 5, 32–57. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.M.; Harrison, F.E. Shared neuropathological characteristics of obesity, type 2 diabetes and alzheimer’s disease: Impacts on cognitive decline. Nutrients 2015, 7, 7332–7357. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Scafato, E.; Capurso, C.; D’Introno, A.; Colacicco, A.M.; Frisardi, V.; Vendemiale, G.; Baldereschi, M.; Crepaldi, G.; Di Carlo, A.; et al. Metabolic syndrome, mild cognitive impairment, and progression to dementia. The italian longitudinal study on aging. Neurobiol. Aging 2011, 32, 1932–1941. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Tangney, C.C. Dietary fat composition and dementia risk. Neurobiol. Aging 2014, 35 (Suppl. 2), S59–S64. [Google Scholar] [CrossRef] [PubMed]
- Francis, H.; Stevenson, R. The longer-term impacts of western diet on human cognition and the brain. Appetite 2013, 63, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Kanoski, S.E.; Davidson, T.L. Western diet consumption and cognitive impairment: Links to hippocampal dysfunction and obesity. Physiol. Behav. 2011, 103, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Winocur, G.; Greenwood, C.E. Studies of the effects of high fat diets on cognitive function in a rat model. Neurobiol. Aging 2005, 26 (Suppl. 1), 46–49. [Google Scholar] [CrossRef] [PubMed]
- Pistell, P.J.; Morrison, C.D.; Gupta, S.; Knight, A.G.; Keller, J.N.; Ingram, D.K.; Bruce-Keller, A.J. Cognitive impairment following high fat diet consumption is associated with brain inflammation. J. Neuroimmunol. 2010, 219, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Lakhan, S.E.; Kirchgessner, A. The emerging role of dietary fructose in obesity and cognitive decline. Nutr. J. 2013, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.M.; Konanur, V.R.; Taing, L.; Usui, R.; Kayser, B.D.; Goran, M.I.; Kanoski, S.E. Effects of sucrose and high fructose corn syrup consumption on spatial memory function and hippocampal neuroinflammation in adolescent rats. Hippocampus 2015, 25, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Lozano, I.; Van der Werf, R.; Bietiger, W.; Seyfritz, E.; Peronet, C.; Pinget, M.; Jeandidier, N.; Maillard, E.; Marchioni, E.; Sigrist, S.; et al. High-fructose and high-fat diet-induced disorders in rats: Impact on diabetes risk, hepatic and vascular complications. Nutr. Metab. 2016, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Dearborn, J.L.; Knopman, D.; Sharrett, A.R.; Schneider, A.L.; Jack, C.R., Jr.; Coker, L.H.; Alonso, A.; Selvin, E.; Mosley, T.H.; Wagenknecht, L.E.; et al. The metabolic syndrome and cognitive decline in the atherosclerosis risk in communities study (aric). Dement. Geriatr. Cogn. Disord. 2014, 38, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Crescenzo, R.; Bianco, F.; Coppola, P.; Mazzoli, A.; Cigliano, L.; Liverini, G.; Iossa, S. The effect of high-fat--high-fructose diet on skeletal muscle mitochondrial energetics in adult rats. Eur. J. Nutr. 2015, 54, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Korivi, M.; Tsai, C.H.; Yang, J.H.; Tsai, Y.C. Supplementation of lactobacillus plantarum k68 and fruit-vegetable ferment along with high fat-fructose diet attenuates metabolic syndrome in rats with insulin resistance. Evid. Based Complement. Altern. Med. 2013, 2013, 943020. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Burrington, C.M.; Graff, E.C.; Zhang, J.; Judd, R.L.; Suksaranjit, P.; Kaewpoowat, Q.; Davenport, S.K.; O’Neill, A.M.; Greene, M.W. Metabolic phenotype and adipose and liver features in a high-fat western diet-induced mouse model of obesity-linked nafld. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E418–E439. [Google Scholar] [CrossRef] [PubMed]
- Manrique, C.; DeMarco, V.G.; Aroor, A.R.; Mugerfeld, I.; Garro, M.; Habibi, J.; Hayden, M.R.; Sowers, J.R. Obesity and insulin resistance induce early development of diastolic dysfunction in young female mice fed a western diet. Endocrinology 2013, 154, 3632–3642. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, M.A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate, high-fat diet-induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.; Tumer, N.; Gao, Y.; Cheng, K.Y.; Scarpace, P.J. Prevention and reversal of diet-induced leptin resistance with a sugar-free diet despite high fat content. Br. J. Nutr. 2011, 106, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Bonda, D.J.; Stone, J.G.; Torres, S.L.; Siedlak, S.L.; Perry, G.; Kryscio, R.; Jicha, G.; Casadesus, G.; Smith, M.A.; Zhu, X.; et al. Dysregulation of leptin signaling in alzheimer disease: Evidence for neuronal leptin resistance. J. Neurochem. 2014, 128, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Ntambi, J.M.; Miyazaki, M. Recent insights into stearoyl-coa desaturase-1. Curr. Opin. Lipidol. 2003, 14, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Kim, Y.C.; Ntambi, J.M. A lipogenic diet in mice with a disruption of the stearoyl-coa desaturase 1 gene reveals a stringent requirement of endogenous monounsaturated fatty acids for triglyceride synthesis. J. Lip. Res. 2001, 42, 1018–1024. [Google Scholar]
- Cohen, P.; Miyazaki, M.; Socci, N.D.; Hagge-Greenberg, A.; Liedtke, W.; Soukas, A.A.; Sharma, R.; Hudgins, L.C.; Ntambi, J.M.; Friedman, J.M. Role for stearoyl-coa desaturase-1 in leptin-mediated weight loss. Science 2002, 297, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Astarita, G.; Jung, K.M.; Vasilevko, V.; Dipatrizio, N.V.; Martin, S.K.; Cribbs, D.H.; Head, E.; Cotman, C.W.; Piomelli, D. Elevated stearoyl-coa desaturase in brains of patients with alzheimer’s disease. PLoS ONE 2011, 6, e24777. [Google Scholar] [CrossRef] [PubMed]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Madison, D.V.; Edson, E.B. Preparation of hippocampal brain slices. Curr. Protoc. Neurosci. 2001. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Parafati, M.; Lascala, A.; Morittu, V.M.; Trimboli, F.; Rizzuto, A.; Brunelli, E.; Coscarelli, F.; Costa, N.; Britti, D.; Ehrlich, J.; et al. Bergamot polyphenol fraction prevents nonalcoholic fatty liver disease via stimulation of lipophagy in cafeteria diet-induced rat model of metabolic syndrome. J. Nutr. Biochem. 2015, 26, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Gomez-Pinilla, F. ‘Metabolic syndrome’ in the brain: Deficiency in omega-3 fatty acid exacerbates dysfunctions in insulin receptor signalling and cognition. J. Physiol. 2012, 590, 2485–2499. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, D.A. Fructosamine: Structure, analysis, and clinical usefulness. Clin. Chem. 1987, 33, 2153–2163. [Google Scholar] [PubMed]
- von Frankenberg, A.D.; Marina, A.; Song, X.; Callahan, H.S.; Kratz, M.; Utzschneider, K.M. A high-fat, high-saturated fat diet decreases insulin sensitivity without changing intra-abdominal fat in weight-stable overweight and obese adults. Eur. J. Nutr. 2017, 56, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Crescenzo, R.; Bianco, F.; Coppola, P.; Mazzoli, A.; Tussellino, M.; Carotenuto, R.; Liverini, G.; Iossa, S. Fructose supplementation worsens the deleterious effects of short-term high-fat feeding on hepatic steatosis and lipid metabolism in adult rats. Exp. Physiol. 2014, 99, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Ruderman, N.; Chisholm, D.; Pi-Sunyer, X.; Schneider, S. The metabolically obese, normal-weight individual revisited. Diabetes 1998, 47, 699–713. [Google Scholar] [CrossRef] [PubMed]
- Seppala-Lindroos, A.; Vehkavaara, S.; Hakkinen, A.M.; Goto, T.; Westerbacka, J.; Sovijarvi, A.; Halavaara, J.; Yki-Jarvinen, H. Fat accumulation in the liver is associated with defects in insulin suppression of glucose production and serum free fatty acids independent of obesity in normal men. J. Clin. Endocrinol. Metab. 2002, 87, 3023–3028. [Google Scholar] [CrossRef] [PubMed]
- Boitard, C.; Cavaroc, A.; Sauvant, J.; Aubert, A.; Castanon, N.; Laye, S.; Ferreira, G. Impairment of hippocampal-dependent memory induced by juvenile high-fat diet intake is associated with enhanced hippocampal inflammation in rats. Brain Behav. Immun. 2014, 40, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.P.; Bartness, T.J.; Mielke, J.G.; Parent, M.B. A high fructose diet impairs spatial memory in male rats. Neurobiol. Learn. Mem. 2009, 92, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Freeman, L.R.; Haley-Zitlin, V.; Rosenberger, D.S.; Granholm, A.-C. Damaging effects of a high-fat diet to the brain and cognition: A review of proposed mechanisms. Nutr. Neurosci. 2014, 17, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Haring, S.J.; Harris, R.B. The relation between dietary fructose, dietary fat and leptin responsiveness in rats. Physiol. Behav. 2011, 104, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A. Role of the blood-brain barrier in the evolution of feeding and cognition. Ann. N. Y. Acad. Sci. 2012, 1264, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Farr, S.A.; Banks, W.A.; Morley, J.E. Effects of leptin on memory processing. Peptides 2006, 27, 1420–1425. [Google Scholar] [CrossRef] [PubMed]
- Valladolid-Acebes, I.; Fole, A.; Martin, M.; Morales, L.; Cano, M.V.; Ruiz-Gayo, M.; Del Olmo, N. Spatial memory impairment and changes in hippocampal morphology are triggered by high-fat diets in adolescent mice. Is there a role of leptin? Neurobiol. Learn. Mem. 2013, 106, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhu, B.; Hua, F. Leptin deficiency is involved in the cognitive impairment of streptozocin-induced diabetic rats undergoing cardiopulmonary bypass. Int. J. Clin. Exp. Med. 2014, 7, 2571–2577. [Google Scholar] [PubMed]
- Li, X.L.; Aou, S.; Oomura, Y.; Hori, N.; Fukunaga, K.; Hori, T. Impairment of long-term potentiation and spatial memory in leptin receptor-deficient rodents. Neuroscience 2002, 113, 607–615. [Google Scholar] [CrossRef]
- Park, E.I.; Paisley, E.A.; Mangian, H.J.; Swartz, D.A.; Wu, M.X.; O’Morchoe, P.J.; Behr, S.R.; Visek, W.J.; Kaput, J. Lipid level and type alter stearoyl coa desaturase mrna abundance differently in mice with distinct susceptibilities to diet-influenced diseases. J. Nutr. 1997, 127, 566–573. [Google Scholar] [PubMed]
- Ntambi, J.M. The regulation of stearoyl-coa desaturase (scd). Prog. Lip. Res. 1995, 34, 139–150. [Google Scholar] [CrossRef]
- Mauvoisin, D.; Prevost, M.; Ducheix, S.; Arnaud, M.P.; Mounier, C. Key role of the erk1/2 mapk pathway in the transcriptional regulation of the stearoyl-coa desaturase (scd1) gene expression in response to leptin. Mol. Cell. Endocrinol. 2010, 319, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Biddinger, S.B.; Miyazaki, M.; Boucher, J.; Ntambi, J.M.; Kahn, C.R. Leptin suppresses stearoyl-coa desaturase 1 by mechanisms independent of insulin and sterol regulatory element-binding protein-1c. Diabetes 2006, 55, 2032–2041. [Google Scholar] [CrossRef] [PubMed]
- Jicha, G.A.; Markesbery, W.R. Omega-3 fatty acids: Potential role in the management of early alzheimer’s disease. Clin. Interv. Aging 2010, 5, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Amtul, Z.; Westaway, D.; Cechetto, D.F.; Rozmahel, R.F. Oleic acid ameliorates amyloidosis in cellular and mouse models of alzheimer’s disease. Brain Pathol. 2011, 21, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Fabelo, N.; Santpere, G.; Puig, B.; Marin, R.; Ferrer, I.; Diaz, M. Lipid alterations in lipid rafts from alzheimer’s disease human brain cortex. J. Alzheimer’s Dis. 2010, 19, 489–502. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Parameters | Control | Soybean Oil | Coconut Oil |
|---|---|---|---|
| Glucose (mg/dL) | 131.3 ± 4.36 | 135.3 ± 4.14 | 142.3 ± 4.72 |
| Insulin (μg/L) | 2.44 ± 0.12 | 2.58 ± 0.13 | 2.52 ± 0.17 |
| TG (mg/dL) | 75.9 ± 3.31 | 80.5 ± 7.77 | 78.6 ± 6.47 |
| TC (mg/dL) | 75.6 ± 3.31 | 78.3 ± 3.00 | 79.4 ± 1.77 |
| HDL (mg/dL) | 21.0 ± 0.97 | 21.2 ± 0.82 | 21.7 ± 1.07 |
| LDL (mg/dL) | 10.3 ± 1.23 | 9.42 ± 0.73 | 9.70 ± 0.52 |
| BW (g) | 260.6 ± 2.87 | 267.9 ± 2.05 | 267.2 ± 2.54 |
| Biochemical Parameters | Control | Soybean Oil | Coconut Oil |
|---|---|---|---|
| Glucose (mg/dL) | 159.8 ± 4.96 b | 169.1 ± 6.30 ab | 208.8 ± 17.2 a |
| Insulin (μg/L) | 1.29 ± 0.29 b | 1.88 ± 2.4 ab | 3.26 ± 0.44 a |
| TG (mg/dL) | 169.4 ± 23.9 b | 134.0 ± 11.3 c | 316.5 ± 20.6 a |
| TC (mg/dL) | 85.6 ± 4.44 | 75.7 ± 2.79 | 84.3 ± 2.89 |
| HDL (mg/dL) | 18.9 ± 1.90 b | 16.9 ± 0.48 c | 27.0 ± 0.97 a |
| LDL (mg/dL) | 6.14 ± 0.46 | 6.89 ± 0.39 | 7.27 ± 0.47 |
| BW (g) | 626.9 ± 26.12 ab | 662.9 ± 19.33 a | 604.6 ± 13.12 b |
| Adiposity index # (%) | 5.3 ± 0.8 c | 6.6 ± 0.5 b | 8.2 ± 0.9 a |
| Fatty Acid (% of Total Fatty Acids) | Control | Soybean Oil | Coconut Oil |
|---|---|---|---|
| Myristic acid (C14:0) | 0.65 ± 0.09 | 0.54 ± 0.14 | 0.72 ± 0.10 |
| Palmitic acid (C16:0) | 5.54 ± 1.19 | 4.23 ± 1.35 | 5.19 ± 2.09 |
| Palmitoleic acid (C16:1) | 1.58 ± 1.51 | 4.47 ± 1.59 | 3.75 ± 1.64 |
| Stearic acid (C18:0) | 1.52 ± 1.09 | 0.98 ± 0.61 | 0.27 ± 0.08 |
| Oleic acid (C18:1) | 6.51 ± 0.70 b | 6.22 ± 0.39 b | 8.98 ± 1.03 a |
| Linoleic acid (C18:2) | 1.94 ± 1.00 b | 4.48 ± 1.50 a | 1.00 ± 0.70 b |
| Linolenic acid (C18:3) | 0.14 ± 0.04 | 0.33 ± 0.12 | 0.18 ± 0.07 |
| Eicosenoic acid (C20:1) | 0.26 ± 0.06 b | 0.25 ± 0.07 b | 0.70 ± 0.13 a |
| Eicosadienoic acid (C20:2) | 1.31 ± 0.61 | 1.57 ± 0.55 | 0.61 ± 0.45 |
| Arachidonic acid (C20:4) | 2.37 ± 0.30 b | 2.31 ± 0.46 b | 4.10 ± 0.73 a |
| Docosapentaenoic acid (C22:5) | 1.59 ± 0.76 | 1.57 ± 0.64 | 0.54 ± 0.49 |
| Docosahexaenoic acid (C22:6) | 2.69 ± 0.33 b | 2.35 ± 0.49 b | 4.25 ± 0.69 a |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-I.; Shen, C.-F.; Hsu, T.-H.; Lin, S.-H. A High-Fructose-High-Coconut Oil Diet Induces Dysregulating Expressions of Hippocampal Leptin and Stearoyl-CoA Desaturase, and Spatial Memory Deficits in Rats. Nutrients 2017, 9, 619. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9060619
Lin C-I, Shen C-F, Hsu T-H, Lin S-H. A High-Fructose-High-Coconut Oil Diet Induces Dysregulating Expressions of Hippocampal Leptin and Stearoyl-CoA Desaturase, and Spatial Memory Deficits in Rats. Nutrients. 2017; 9(6):619. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9060619
Chicago/Turabian StyleLin, Ching-I, Chu-Fu Shen, Tsui-Han Hsu, and Shyh-Hsiang Lin. 2017. "A High-Fructose-High-Coconut Oil Diet Induces Dysregulating Expressions of Hippocampal Leptin and Stearoyl-CoA Desaturase, and Spatial Memory Deficits in Rats" Nutrients 9, no. 6: 619. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9060619