Involvement of CD36 in Modulating the Decrease of NPY and AgRP Induced by Acute Palmitic Acid Stimulation in N1E-115 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatment
2.2. Cell ViabilityAssay
2.3. RNA Extraction and Real-Time Polymerase Chain Reaction (PCR) Analysis
2.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results
3.1. PA Treatment Does Not Affect N1E-115 Cell Viability
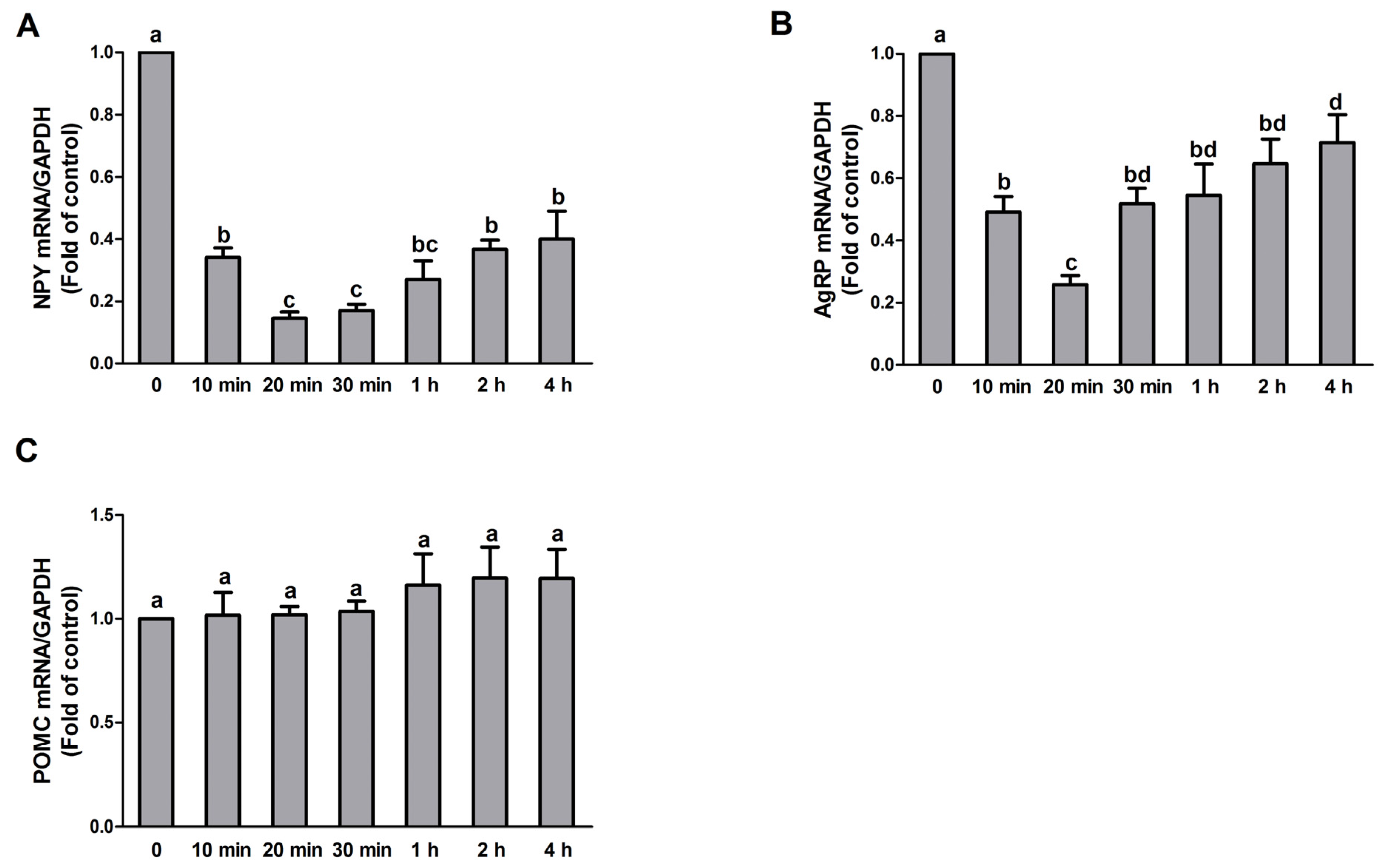
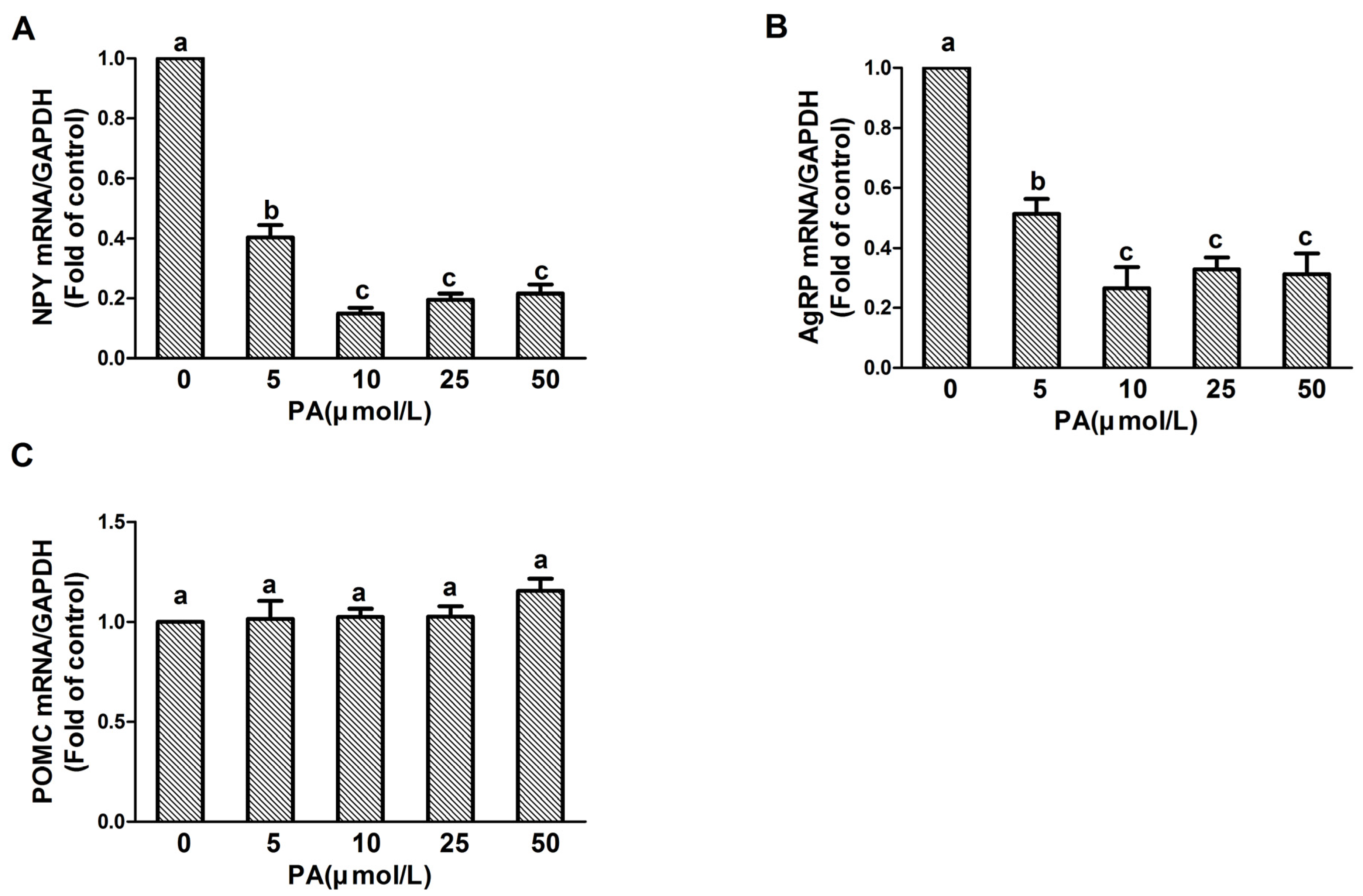
3.2. PA Treatment Affects NPY and AgRP but Not POMC Expression in N1E-115 Cells
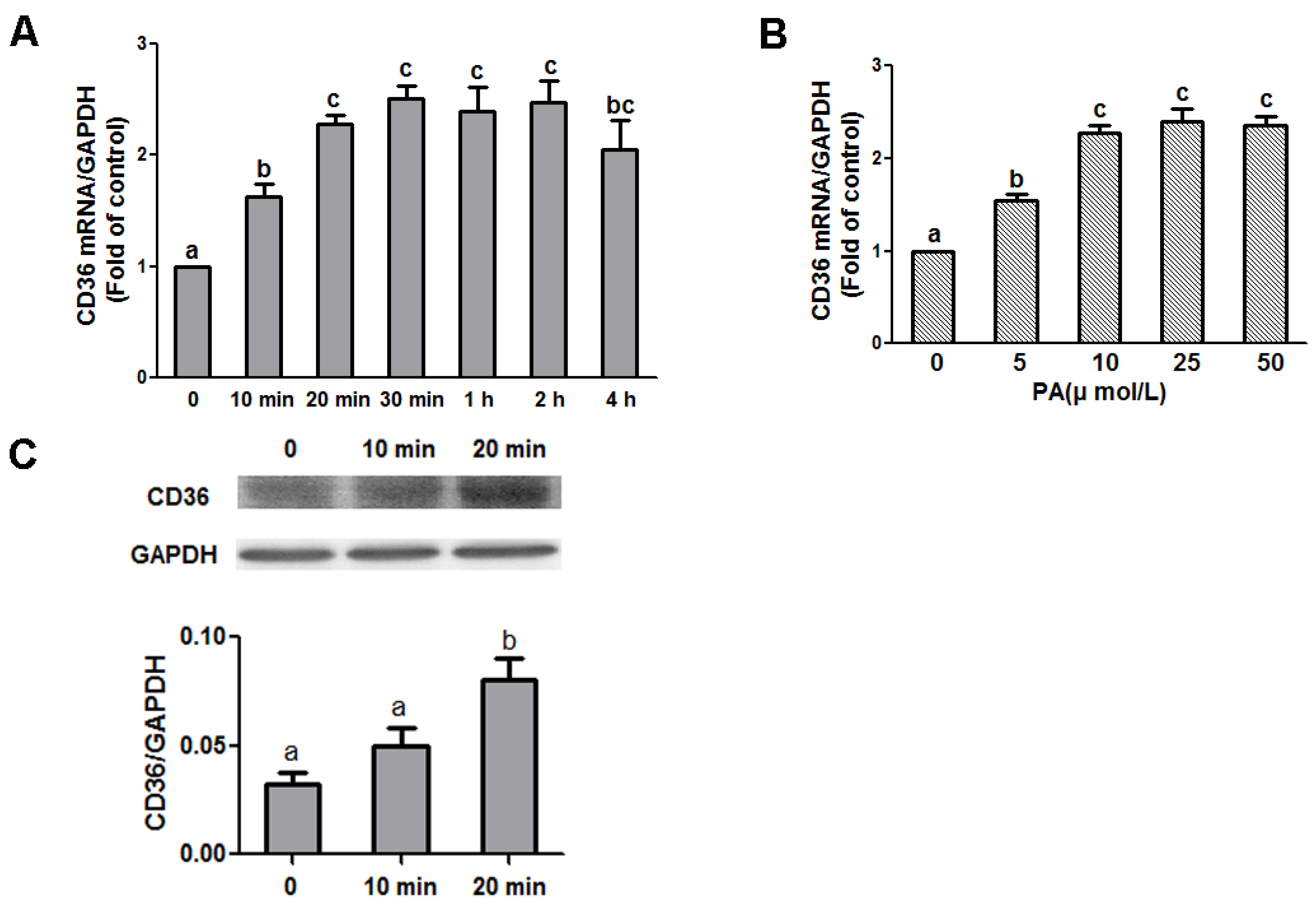
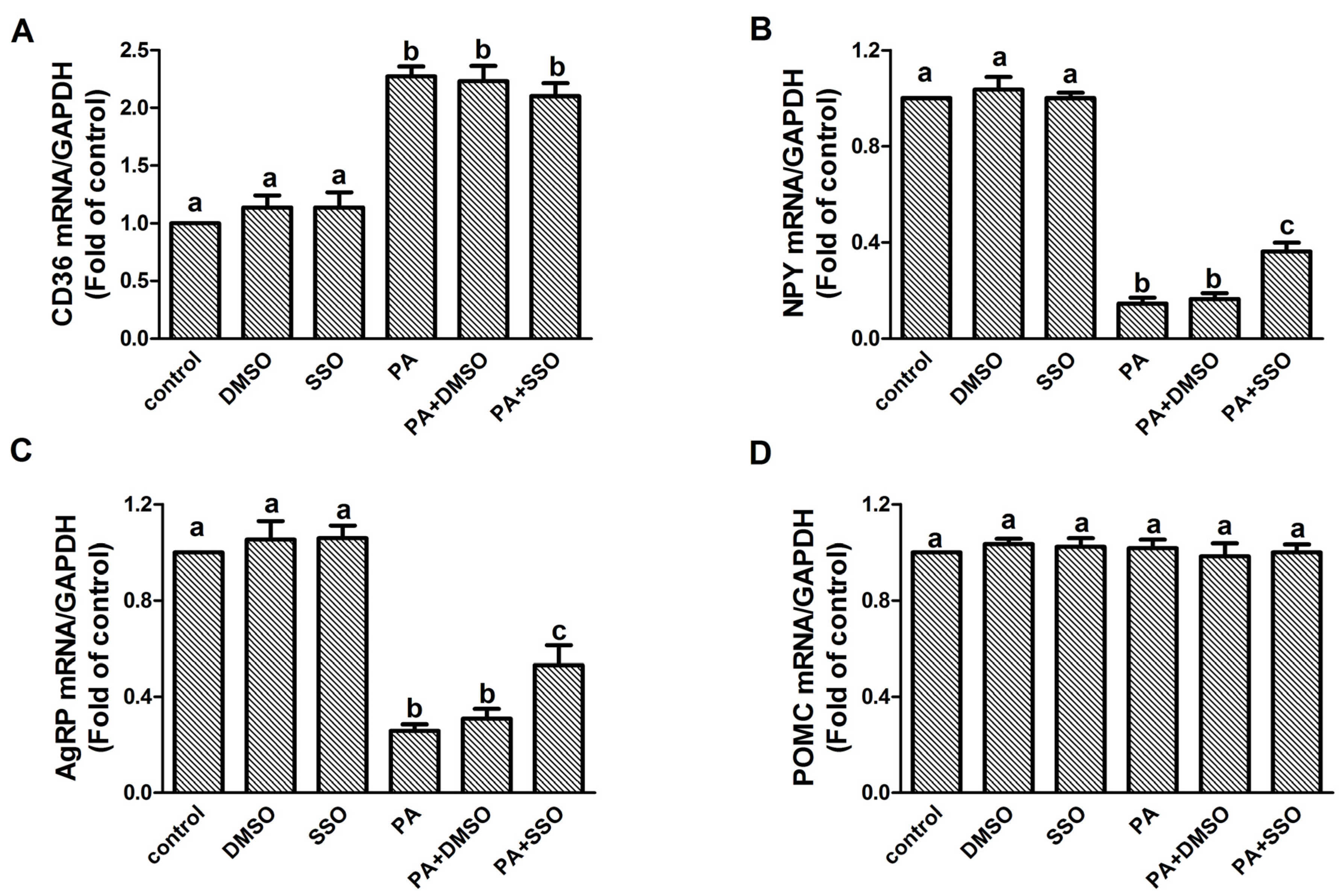
3.3. CD36 Is Involved in the PA-Dependent Regulation of NPY and AgRP Gene Expression
4. Discussion and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schwartz, M.W.; Woods, S.C.; Porte, D.J.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [PubMed]
- Blouet, C.; Schwartz, G.J. Hypothalamic nutrient sensing in the control of energy homeostasis. Behav. Brain Res. 2010, 209, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.E.; Magnan, C.; Dunn-Meynell, A.; Le Foll, C. Metabolic sensing and the brain: Who, what, where, and how? Endocrinology 2011, 152, 2552–2557. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.D.; Konner, A.C.; Bruning, J.C. Sensing the fuels: Glucose and lipid signaling in the cns controlling energy homeostasis. Cell. Mol. Life Sci. 2010, 67, 3255–3273. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Lelliott, C.J.; Vidal-Puig, A. Hypothalamic fatty acid metabolism: A housekeeping pathway that regulates food intake. Bioessays 2007, 29, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Gropp, E.; Shanabrough, M.; Borok, E.; Xu, A.W.; Janoschek, R.; Buch, T.; Plum, L.; Balthasar, N.; Hampel, B.; Waisman, A.; et al. Agouti-related peptide-expressing neurons are mandatory for feeding. Nat. Neurosci. 2005, 8, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Luquet, S.; Perez, F.A.; Hnasko, T.S.; Palmiter, R.D. Npy/agrp neurons are essential for feeding in adult mice but can be ablated in neonates. Science 2005, 310, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Hahn, T.M.; Breininger, J.F.; Baskin, D.G.; Schwartz, M.W. Coexpression of agrp and npy in fasting-activated hypothalamic neurons. Nat. Neurosci. 1998, 1, 271–272. [Google Scholar] [PubMed]
- Aponte, Y.; Atasoy, D.; Sternson, S.M. Agrp neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nat. Neurosci. 2011, 14, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Krashes, M.J.; Koda, S.; Ye, C.; Rogan, S.C.; Adams, A.C.; Cusher, D.S.; Maratos-Flier, E.; Roth, B.L.; Lowell, B.B. Rapid, reversible activation of agrp neurons drives feeding behavior in mice. J. Clin. Investig. 2011, 121, 1424–1428. [Google Scholar] [CrossRef] [PubMed]
- Posey, K.A.; Clegg, D.J.; Printz, R.L.; Byun, J.; Morton, G.J.; Vivekanandan-Giri, A.; Pennathur, S.; Baskin, D.G.; Heinecke, J.W.; Woods, S.C.; et al. Hypothalamic proinflammatory lipid accumulation, inflammation, and insulin resistance in rats fed a high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1003–E1012. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B.; Lee, H.K.; Querfurth, H.W. Oleate prevents palmitate-induced mitochondrial dysfunction, insulin resistance and inflammatory signaling in neuronal cells. BBA Mol. Cell Res. 2014, 1843, 1402–1413. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Chevrot, M.; Poirier, H.; Passilly-Degrace, P.; Niot, I.; Besnard, P. Cd36 as a lipid sensor. Physiol. Behav. 2011, 105, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Le Foll, C.; Dunn-Meynell, A.; Musatov, S.; Magnan, C.; Levin, B.E. Fat/cd36: A major regulator of neuronal fatty acid sensing and energy homeostasis in rats and mice. Diabetes 2013, 62, 2709–2716. [Google Scholar] [CrossRef] [PubMed]
- Luiken, J.J.; van Nieuwenhoven, F.A.; America, G.; van der Vusse, G.J.; Glatz, J.F. Uptake and metabolism of palmitate by isolated cardiac myocytes from adult rats: Involvement of sarcolemmal proteins. J. Lipid Res. 1997, 38, 745–758. [Google Scholar] [PubMed]
- Coort, S.L.; Willems, J.; Coumans, W.A.; van der Vusse, G.J.; Bonen, A.; Glatz, J.F.; Luiken, J.J. Sulfo-n-succinimidyl esters of long chain fatty acids specifically inhibit fatty acid translocase (fat/cd36)-mediated cellular fatty acid uptake. Mol. Cell. Biochem. 2002, 239, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Horvath, T.L.; Diano, S.; Tschop, M. Brain circuits regulating energy homeostasis. Neuroscientist 2004, 10, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Moulle, V.S.; Picard, A.; Le Foll, C.; Levin, B.E.; Magnan, C. Lipid sensing in the brain and regulation of energy balance. Diabetes Metab. 2014, 40, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Kleinridders, A.; Schenten, D.; Konner, A.C.; Belgardt, B.F.; Mauer, J.; Okamura, T.; Wunderlich, F.T.; Medzhitov, R.; Bruning, J.C. Myd88 signaling in the cns is required for development of fatty acid-induced leptin resistance and diet-induced obesity. Cell Metab. 2009, 10, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Moullé, V.S.F.; Cansell, C.; Luquet, S.; Cruciani-Guglielmacci, C. The multiple roles of fatty acid handling proteins in brain. Front. Physiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Le Foll, C.; Irani, B.G.; Magnan, C.; Dunn-Meynell, A.A.; Levin, B.E. Characteristics and mechanisms of hypothalamic neuronal fatty acid sensing. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R655–R664. [Google Scholar] [CrossRef] [PubMed]
- Kien, C.L. Dietary interventions for metabolic syndrome: Role of modifying dietary fats. Curr. Diab. Rep. 2009, 9, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.; Spitzer, J.J. Fatty acid profiles of various lipids in the cerebrospinal fluid. Proc. Soc. Exp. Biol. Med. 1971, 136, 1294–1296. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Richelson, E.; Nirenberg, M. Neurotransmitter synthesis by neuroblastoma clones (neuroblast differentiation-cell culture-choline acetyltransferase-acetylcholinesterase-tyrosine hydroxylase-axons-dendrites). Proc. Natl. Acad. Sci. USA 1972, 69, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Richelson, E. The use of cultured cells in the study of mood-normalizing drugs. Pharmacol. Toxicol. 1990, 66, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Obici, S.; Feng, Z.; Morgan, K.; Stein, D.; Karkanias, G.; Rossetti, L. Central administration of oleic acid inhibits glucose production and food intake. Diabetes 2002, 51, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Obici, S.; Zhang, B.B.; Karkanias, G.; Rossetti, L. Hypothalamic insulin signaling is required for inhibition of glucose production. Nat. Med. 2002, 8, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Morgan, K.; Obici, S.; Rossetti, L. Hypothalamic responses to long-chain fatty acids are nutritionally regulated. J. Biol. Chem. 2004, 279, 31139–31148. [Google Scholar] [CrossRef] [PubMed]
- Le Foll, C.; Dunn-Meynell, A.A.; Levin, B.E. Role of fat/cd36 in fatty acid sensing, energy, and glucose homeostasis regulation in dio and dr rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R188–R198. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Li, I.; Roberts, L.R.; Chan, C. Elevated free fatty acid uptake via cd36 promotes epithelial-mesenchymal transition in hepatocellular carcinoma. Sci. Rep. 2015, 5, 14752. [Google Scholar] [CrossRef] [PubMed]
- Pepino, M.Y.; Kuda, O.; Samovski, D.; Abumrad, N.A. Structure-function of cd36 and importance of fatty acid signal transduction in fat metabolism. Annu. Rev. Nutr. 2014, 34, 281–303. [Google Scholar] [CrossRef] [PubMed]
- Kuda, O.; Pietka, T.A.; Demianova, Z.; Kudova, E.; Cvacka, J.; Kopecky, J.; Abumrad, N.A. Sulfo-n-succinimidyl oleate (SSO) inhibits fatty acid uptake and signaling for intracellular calcium via binding cd36 lysine 164. J. Biol. Chem. 2013, 288, 15547–15555. [Google Scholar] [CrossRef] [PubMed]
- Dramane, G.; Abdoul-Azize, S.; Hichami, A.; Vogtle, T.; Akpona, S.; Chouabe, C.; Sadou, H.; Nieswandt, B.; Besnard, P.; Khan, N.A. Stim1 regulates calcium signaling in taste bud cells and preference for fat in mice. J. Clin. Investig. 2012, 122, 2267–2282. [Google Scholar] [CrossRef] [PubMed]
- Obici, S.; Feng, Z.; Arduini, A.; Conti, R.; Rossetti, L. Inhibition of hypothalamic carnitine palmitoyltransferase-1 decreases food intake and glucose production. Nat. Med. 2003, 9, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Bady, I.; Marty, N.; Dallaporta, M.; Emery, M.; Gyger, J.; Tarussio, D.; Foretz, M.; Thorens, B. Evidence from glut2-null mice that glucose is a critical physiological regulator of feeding. Diabetes 2006, 55, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Eny, K.M.; Wolever, T.M.; Fontaine-Bisson, B.; El-Sohemy, A. Genetic variant in the glucose transporter type 2 is associated with higher intakes of sugars in two distinct populations. Physiol. Genom. 2008, 33, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.Z.; Hulsey, M.G.; Martin, R.J. Intracerebroventricular administration of antisense oligodeoxynucleotide against glut2 glucose transporter mrna reduces food intake, body weight change and glucoprivic feeding response in rats. J. Nutr. 1998, 128, 287–291. [Google Scholar] [PubMed]
- Gremlich, S.; Bonny, C.; Waeber, G.; Thorens, B. Fatty acids decrease idx-1 expression in rat pancreatic islets and reduce glut2, glucokinase, insulin, and somatostatin levels. J. Biol. Chem. 1997, 272, 30261–30269. [Google Scholar] [CrossRef] [PubMed]
- Koch, L.; Wunderlich, F.T.; Seibler, J.; Konner, A.C.; Hampel, B.; Irlenbusch, S.; Brabant, G.; Kahn, C.R.; Schwenk, F.; Bruning, J.C. Central insulin action regulates peripheral glucose and fat metabolism in mice. J. Clin. Investig. 2008, 118, 2132–2147. [Google Scholar] [CrossRef] [PubMed]
- Theander-Carrillo, C.; Wiedmer, P.; Cettour-Rose, P.; Nogueiras, R.; Perez-Tilve, D.; Pfluger, P.; Castaneda, T.R.; Muzzin, P.; Schurmann, A.; Szanto, I.; et al. Ghrelin action in the brain controls adipocyte metabolism. J. Clin. Investig. 2006, 116, 1983–1993. [Google Scholar] [CrossRef] [PubMed]
- Minokoshi, Y.; Kim, Y.B.; Peroni, O.D.; Fryer, L.G.; Muller, C.; Carling, D.; Kahn, B.B. Leptin stimulates fatty-acid oxidation by activating amp-activated protein kinase. Nature 2002, 415, 339–343. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Wang, X.; Yang, H.; Zhang, X.; Yang, N. Involvement of CD36 in Modulating the Decrease of NPY and AgRP Induced by Acute Palmitic Acid Stimulation in N1E-115 Cells. Nutrients 2017, 9, 626. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9060626
Ma Y, Wang X, Yang H, Zhang X, Yang N. Involvement of CD36 in Modulating the Decrease of NPY and AgRP Induced by Acute Palmitic Acid Stimulation in N1E-115 Cells. Nutrients. 2017; 9(6):626. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9060626
Chicago/Turabian StyleMa, Yan, Xiaoyi Wang, Hongying Yang, Xu Zhang, and Nianhong Yang. 2017. "Involvement of CD36 in Modulating the Decrease of NPY and AgRP Induced by Acute Palmitic Acid Stimulation in N1E-115 Cells" Nutrients 9, no. 6: 626. https://0-doi-org.brum.beds.ac.uk/10.3390/nu9060626