Sea Anemones: Quiet Achievers in the Field of Peptide Toxins


1
School of Earth, Environmental and Biological Sciences, Queensland University of Technology (QUT), Brisbane, QLD 4001, Australia
2
Institute of Future Environments, Queensland University of Technology (QUT), Brisbane, QLD 4001, Australia
3
Faculty of Health, Queensland University of Technology (QUT), Brisbane, QLD 4001, Australia
4
Medicinal Chemistry, Monash Institute of Pharmaceutical Sciences, Monash University, Parkville, VIC 3052, Australia
*
Author to whom correspondence should be addressed.
Toxins 2018, 10(1), 36; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10010036
Submission received: 18 December 2017
/
Revised: 4 January 2018
/
Accepted: 4 January 2018
/
Published: 8 January 2018
(This article belongs to the Special Issue Selected Papers from the 2017 Venoms to Drugs Conference)
Abstract
:Sea anemones have been understudied as a source of peptide and protein toxins, with relatively few examined as a source of new pharmacological tools or therapeutic leads. This is surprising given the success of some anemone peptides that have been tested, such as the potassium channel blocker from Stichodactyla helianthus known as ShK. An analogue of this peptide, ShK-186, which is now known as dalazatide, has successfully completed Phase 1 clinical trials and is about to enter Phase 2 trials for the treatment of autoimmune diseases. One of the impediments to the exploitation of sea anemone toxins in the pharmaceutical industry has been the difficulty associated with their high-throughput discovery and isolation. Recent developments in multiple ‘omic’ technologies, including genomics, transcriptomics and proteomics, coupled with advanced bioinformatics, have opened the way for large-scale discovery of novel sea anemone toxins from a range of species. Many of these toxins will be useful pharmacological tools and some will hopefully prove to be valuable therapeutic leads.
Keywords:
sea anemone; peptide; ShK; potassium channel; autoimmune disease; genomics; transcriptomics; proteomics; evolutionKey Contribution:
The sea anemone peptide ShK highlights the potential of these venomous animals to produce valuable therapeutic leads, and the abundance in nature of peptides related to ShK suggests that this scaffold can support a range of functions. Current genomic, transcriptomic and proteomic studies of sea anemones promise to greatly expand the number of ShK analogues and to identify a range of novel peptide families.1. Introduction
Sea anemones are members of the phylum Cnidaria, class Anthozoa, subclass Hexacorallia and order Actiniaria, one of the oldest extant orders of venomous animals. In common with many other venomous animals, they produce venom that is a complex mixture of small molecules, peptides and proteins [1,2,3,4,5,6,7]. Unlike some of the better known groups of venomous animals such as snakes and spiders, however, or even their fellow cnidarians the Australian box jellyfish or Irukandji, which (quite appropriately) attract considerable public attention [8,9], the potentially harmful consequences of contact with sea anemones are relatively unknown. Indeed, many members of the public are not even aware that they are animals rather than plants, let alone venomous ones (Figure 1).
While most injuries caused by sea anemones are associated with skin rashes and oedema, more extreme reactions have been reported for several species, including Actinodendron plumosum and other species from the family Actinodendronidae (species from this family are collectively known as the hell’s fire anemones), Telmatactis species, Phyllodiscus semoni (night or wasp anemone), Actinia equina (beadlet anemone) and Anemonia sulcata (the snakelocks anemone, synonomy Anemonia viridis) [10,11]. For example, a swimmer lost consciousness and underwent cardiopulmonary arrest after being stung by the sea anemone, Actinia equina, although this may have been a consequence of an anaphylactic reaction following prior exposure to unknown sea anemones [12]. Sea anemones from the family Aliciideae are known to be particularly dangerous to humans, with severe reactions observed following contact with both Triactis producta [13] and P. semoni [10]. In fact, P. semoni is responsible for one of the few fatalities to result from sea anemone envenomation [14]. The venom from P. semoni has caused acute renal failure in humans, with a protein toxin (PsTX-115) from this venom causing severe kidney damage in rat models [15].
2. Venom Apparatus
Sea anemones, in common with other members of the phylum Cnidaria, possess numerous specialized stinging cells (cnidocytes) that are widely distributed throughout the body [16]. These stinging cells are equipped with organelles known as nematocysts (cnidae), which contain small threads that are forcefully everted when stimulated mechanically or chemically [17]. These nematocysts contain a complex cocktail of toxins that is used to envenomate predatory and prey species upon discharge [1,3,6]. Nematocysts show significant heterogeneity in their density and morphology across different structures within sea anemones [18]. For example, in Actinia tenebrosa (or its northern hemisphere relatives Actinia equina and Actinia fragacea), acrorhagi (which are used in aggressive intra-specific combat (Figure 2)) contain holotrichs and basitrichs [19] (Figure 3), while the tentacles (which are used in prey capture and defence) contain basitrichs and spirocysts; basitrichs are also found in the mesenteric filaments, column, pedal disc and actinopharynx [19] (Figure 3). The differences in cnidae composition are related to differences in the functional specialization of morphological structures, i.e., the capture of prey (crustaceans, small fish), defence against predators and intraspecific aggression [19,20,21,22].
3. Peptide Toxins from Anemones
While many families of sea anemone toxins have been described already [7,23], much still remains to be discovered about toxins in this group. In fact, there are only 236 peptide or protein toxins in the manually curated ToxProt database [24], which have been isolated from just 45 sea anemone species. This means that fewer than four percent (45 of more than 1100 species) of all sea anemones have had their venom peptides and proteins examined. Currently, anemone toxins can be classified into 15 known families (Table 1), but 19 toxin peptides and proteins remain largely uncharacterized. The four most common toxin protein families isolated from anemones are the actinoporin family, sea anemone subfamily (30 proteins found in the ToxProt database); sea anemone sodium channel inhibitory toxin family, type I subfamily (52 proteins found in the ToxProt database); sea anemone type 3 (BDS) potassium channel toxin family (32 proteins found in the ToxProt database), and venom Kunitz-type family, sea anemone type 2 potassium channel toxin subfamily (26 proteins found in the ToxProt database). Of these four toxin protein families, only the actinoporin family, sea anemone subfamily, has been found outside the Actinioidea superfamily of anemones based on sequences currently available in the ToxProt database. In fact, 206 of the 236 toxin proteins in the ToxProt database are from the Actinioidea superfamily. This indicates that the species that have been examined show a strong taxonomic bias (Table 1), and we therefore need to examine the venom peptide profile from sea anemone species from other superfamilies. Consequently, there is every reason for optimism that the remaining 96% of species, particularly those distantly related to the superfamily Actinioidea, will provide interesting new peptides, some of which will no doubt have therapeutic potential. Some of the more venomous species, such as Telmatactis australiensis, Dofleinia armata and Triactis producta, should be especially interesting in this context.
This article will focus mainly on ShK domains (which, in sea anemones, are members of the type 1 potassium channel toxin family, Type 1b subfamily, Table 1) because of their demonstrated therapeutic potential and broad distribution in nature, as documented in the next section. However, several other classes of peptide toxins from sea anemones have been investigated as therapeutic leads or pharmacological tools. For example, the sodium channel toxins anthopleurin-A and -B showed initial promise as positive inotropes for use in cardiovascular disease [25,26], although they and their homologues from other anemones, for example ATX-I and -II and ShI [27,28], have also proven to be valuable probes of site 3 on voltage-gated sodium channels [29], which mediates channel inactivation. APETx2, from Anthopleura elegantissima, inhibits the acid-sensing ion channel ASIC3, which is a proton-gated Na+ channel that has been implicated in pain transduction associated with acidosis in inflamed or ischemic tissues [30]. However, this peptide also inhibits NaV1.2 and NaV1.8 channels, and to a lesser extent NaV1.6 [31], as well as the human ether-a-go-go-related (hERG) potassium channel (KV11.1) [32]. APETx1 was identified initially as a gating modifier of the hERG channel [33], but is also promiscuous, inhibiting mammalian NaV1.2–NaV1.6 and NaV1.8 channels [31]. Recently, a new member of the APETx family, APETx4, was shown to have activity on a potential anti-cancer target, the human ether-à-go-go channel (hEag1 or KV10.1), but this peptide also inhibits other KV and NaV channels [34]. The demonstrated lack of target specificity for members of the APETx family limits their application as pharmacological tools, although analogues with specific mutations have the potential to overcome this limitation [31]. Peptides from the sea anemone Heteractis crispa (APHC1, APHC2 and APHC3) are active on TRPV1 receptors [35,36].
The structures of APETx1, APETx2 and BDS-I (which acts on channels containing KV3 subunits, including KV3.4 [37], but also modulates NaV channels [38]) are similar to those of the Na+-channel toxins such as AP-A although quite distinct from those of the ShK/BgK family of toxins (see below). As noted previously [5], sea anemones use common structural scaffolds to create blockers for distinct targets (AP-A, APETx1 and APETx2 act on VGSC, hERG and ASIC channels, respectively), while also using different scaffolds (all-β in APETx1 vs. all-α in ShK) to block similar channels (hERG and KV1, respectively).
Recent proteomic analyses of the venoms of Stichodactyla haddoni [39] and Aulactinia japonicus (designated Cnidopus japonicas in the title of the paper) [40] identified many new toxin families, a number of them with novel cysteine frameworks. A similar analysis of the mucus of Heteractis magnifica revealed the presence of hundreds of peptides [41]. No doubt some of these new peptide families will prove to have useful pharmacological properties and may eventually become therapeutic leads. The next section focuses on a peptide for which this clearly is the case.
4. Potassium Channel Blockers from Sea Anemones: Therapeutic Leads for the Treatment of Autoimmune Diseases
The voltage-gated K+ channel KV1.3 is involved in the activation of a sub-set of T lymphocytes known as effector memory T (TEM) cells as it regulates the membrane potential during activation by allowing K+ efflux to counterbalance the influx of Ca2+ from intracellular stores and through CRAC channels [42]. Blocking KV1.3 channels in TEM cells blocks their activation and proliferation. As TEM cells are key mediators of autoimmune diseases, KV1.3 blockers are attractive leads as a new class of therapeutic for these conditions [43,44].
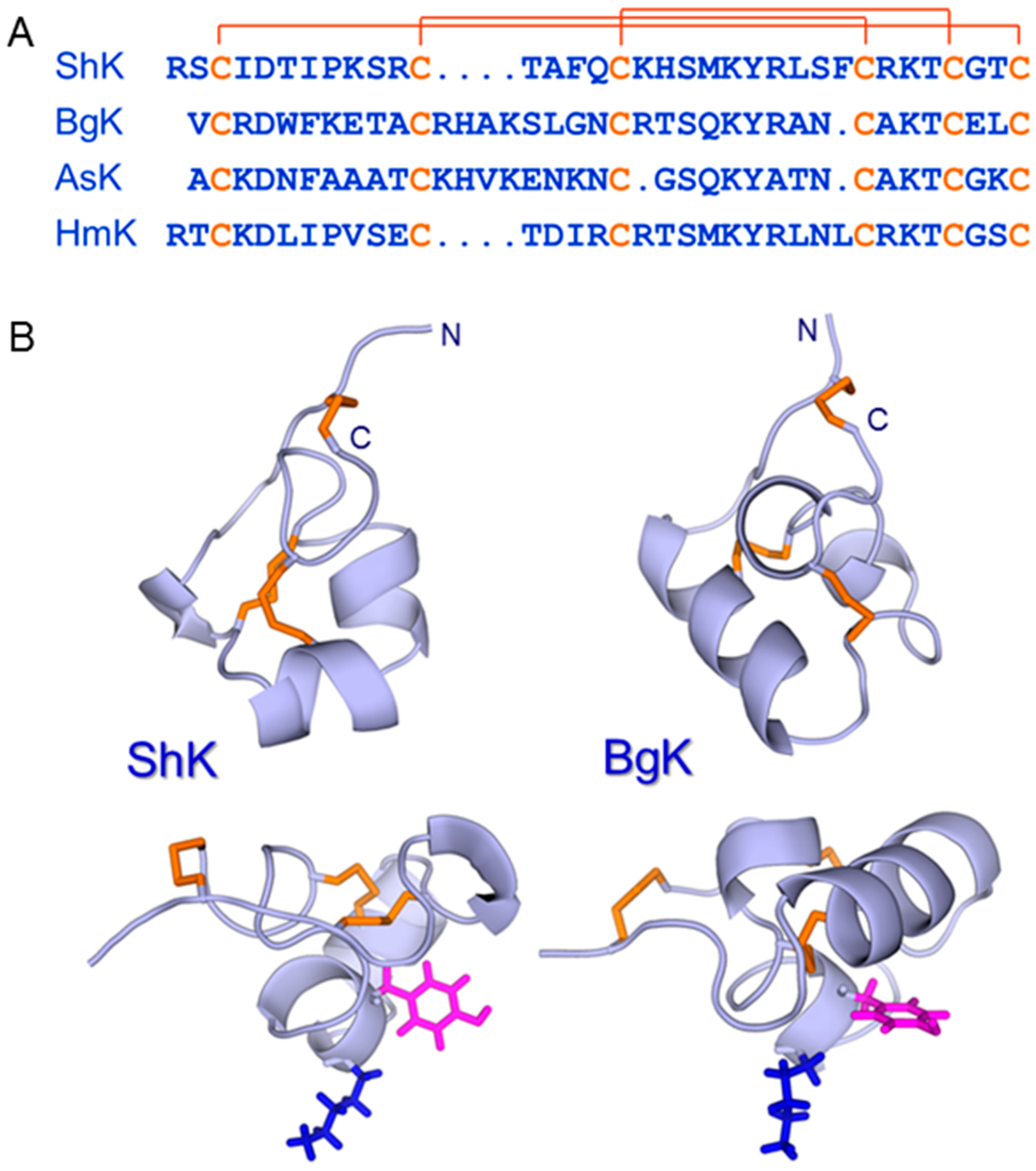
Several peptide toxins from scorpions were found to be potent blockers of KV1.3 [45,46], but did not progress to clinical trials for a variety of reasons, including the lack of desired specificity for this channel over related KV1 channels [47]. Around the same time, a novel peptide, ShK, was isolated from the Caribbean sun anemone, Stichodactyla helianthus [48]. This 35-residue peptide was found to be a potent competitive inhibitor of α-dendrotoxin binding to rat brain synaptosomes and blocked K+ current in dorsal root ganglion cells. Its amino acid sequence [48] (Figure 4), disulfide bonding pattern [49] and solution structure [50] were all very different from the scorpion toxins, although Lys22 and Tyr23 in ShK, the two key residues for KV1.3 blockade, are spatially conserved in an arrangement common to KV-channel blocking peptides from widely different species [51]. ShK has a very high affinity (Ki ~ 10 pM) for KV1.3 channels but also displays high pM affinity for KV1.1, KV1.4 and KV1.6, which are present in brain and cardiac tissues [52,53]. In order for this promising peptide toxin to progress to clinical trials, therefore, more selective analogues had to be developed [54]. This eventually led to ShK-186, which had a 100-fold improvement in selectivity for KV1.3 over KV1.1, KV1.4 and KV1.6 [55].
ShK and its analogues had already shown efficacy in animal models of human autoimmune diseases such as multiple sclerosis and rheumatoid arthritis [56,57], and preclinical testing of ShK-186 yielded favourable results in both rats and monkeys [58]. Unexpectedly, ShK-186 was found to have a long half-life at the site of sub-cutaneous injection, resulting in sustained high pM levels in plasma and a prolonged therapeutic efficacy [58]. ShK-186, which is now known as dalazatide, completed Phase 1a and 1b trials in 2016. The Phase 1b trial in mild-to-moderate plaque psoriasis patients showed that dalazatide was well tolerated and reduced psoriatic skin lesions [59]. It is expected to begin Phase 2a trails in 2018. Dalazatide is being advanced as a treatment for various autoimmune diseases, including inclusion body myositis, lupus, ANCA vasculitis, multiple sclerosis, psoriasis, psoriatic arthritis, rheumatoid arthritis, type 1 diabetes and inflammatory bowel diseases [44,45]. New analogues of ShK with good selectivity for KV1.3 over other KV1 channels have also been developed [60,61,62,63,64].
The progress of ShK analogues towards the clinic has stimulated interest in this class of peptides, driven partly by the potential for finding additional members of this family of peptides in sea anemones that might be selective for KV1.3 or the many other KV channels that are potential therapeutic targets [45]. As a result, it has become apparent that the ‘ShK fold’, typified by ShK [50] and BgK [51], is widely distributed not only in sea anemones and other cnidarians [68,69,70], but also throughout nature. The next section highlights this broad distribution.
5. ShK: A Privileged Scaffold in Nature?
In 2010, the Simple Modular Architecture Research Tool (SMART) database (http://smart.embl-heidelberg.de/) predicted the existence of a large superfamily of proteins that contain domains resembling ShK or BgK, which were referred to collectively as ShKT domains [71]. These domains were distributed across nearly 400 proteins from both the plant and animal kingdoms, including Viridiplantae, Arabidopsis thaliana, Oryza sativa, and green alga Ostreococcus sp.; Protozoa, Cryptosporidium parvum; Cnidaria, sea anemones, hydra, and jellyfish; Echinodermata, sea urchin; Mollusca, bivalve clams and oysters; Ciona, sea squirt Ciona intestinalis; Actinopterygii, zebrafish Danio rerio and pufferfish Takifugu rubripes; Caenorhabditis, C. elegans and C. brigssae; Rhabditida, rhabditid nematodes other than Caenorhabditis sp.; Ophidia, snakes; Xenopus, Xenopus tropicalis; Aves, chicken Gallus gallus; Mammalia. Many of these proteins (~70) were metallopeptidases, whereas others were prolyl-4-hydroxylases, tyrosinases, peroxidases, oxidoreductases, or proteins containing epidermal growth factor-like domains, thrombospondin-type repeats, or trypsin-like serine protease domains [71].
The only human protein containing a ShKT domain in the SMART data base was matrix metalloprotease 23 (MMP23) [72,73]. A second human protein with an ShKT domain, microfibril associated protein MFAP2, was not mentioned in the SMART database at that time [74]. The cysteine-rich secretory proteins (Crisp), which are found predominantly in the mammalian male reproductive tract as well as in the venom of reptiles, are two-domain proteins, one domain of which is an ShKT domain. For example, murine Tpx-1 (testis specific protein-1) contains two subdomains, one of which has a similar fold to BgK and ShK [75]. The Tpx-1 Crisp domain inhibited the cardiac ryanodine receptor (RyR2) and activated the skeletal RyR1.
By 2014, the SMART database identified 668 proteins that contained 1315 ShKT domains. The largest family was found in worms, with 276 of those 668 proteins coming from Caenorhabditis elegans, C. briggsae, Brugia malayi, B. pahangi, Ancylostoma ceylanicum, Schistosoma mansoni and Toxocara canis [76]. This prompted an investigation of whether worm ShKTs share structural similarity to ShK, block KV1.3, and exhibit immunomodulatory activity. Based on phylogenetic analysis, two worm peptides were selected for study: AcK1, a 51-residue peptide expressed in the anterior secretory glands of both the dog-infecting hookworm Ancylostoma caninum and the human-infecting hookworm Ancylostoma ceylanicum, and BmK1, the C-terminal domain of a metalloprotease from the filarial worm Brugia malayi. These peptides proved to have helical structures closely resembling that of ShK [76]. They also blocked KV1.3 channels, selectively suppressed the function of TEM lymphocytes, and inhibited delayed-type hypersensitivity. It was suggested that ShK-like worm peptides may be among the active principles that contribute to the well-known protective effect of parasitic worms in autoimmune diseases [76], although further work is required to confirm this hypothesis.
Recently, we have investigated several new ShK-like peptides identified in anemone transcriptomes. One such peptide is AsK132958, from Anemonia sulcata, which had an ShKT cysteine framework but was six amino acid residues shorter [77]. The disulfide connectivities and structural scaffold were very similar to those of ShK, although the structure was more constrained. However, AsK132958 showed no activity against grass shrimp, Artemia nauplii, or any of the KV channels tested, owing partly to the absence of a functional Lys-Tyr dyad. It appears that Lys19, which would be expected to occupy the pore of the channel, was not sufficiently accessible for binding, and therefore that AsK132958 must have a distinct functional role that does not involve KV channels. The evolutionary relationship between AsK132958 and other ShK-like amino acid sequences is unclear. AsK132958 may represent the shortest peptide sequence that can support a stable ShKT fold, although this remains to be confirmed.
Another peptide, identified in the transcriptome of an Oulactis species, was similar to BgK in terms of amino acid sequence and three-dimensional structure, and furthermore contained a Lys-Tyr dyad, but was inactive against KV channels tested to date (our unpublished results). These findings highlight the likely diversity of functions supported by this versatile scaffold.
At the time of writing, the SMART database includes 3345 ShKT domains spread across 1797 proteins, a huge increase over the numbers documented in 2010 [71] and 2014 [76]. These domains are found mainly in animals and plants, but also occur in fungi, viruses and undefined kingdoms. With the dramatic expansion in the number of genomes and transcriptomes spanning all kingdoms, this number is set to increase rapidly. The next section discusses these developments in more detail and outlines how we might begin to assess the functions of these domains.
6. New Methods for the Large-Scale Detection of Known and Novel Peptide Toxins
The large-scale identification of new peptide toxins, as well as the evolutionary analysis of known peptide toxins, in sea anemones has been limited by a lack of genomic resources for many species. This is changing rapidly, however, as transcriptome and genome assemblies are being generated using next generation sequencing platforms for a number of sea anemone species [39,78,79,80]. Currently, the genome sequences of Nematostella vectensis [81] and Exaiptasia pallida are publicly available, as well as transcriptomes of ~30 anemone species from a range of anemone superfamilies. These genomic resources have enabled the in-depth characterization of gene families in sea anemones, including immune gene families [82], and some detailed investigations of select toxin gene families (e.g., actinoporins, which are cytolytic proteins [83]). These genomic resources can be used to study the number, distribution and evolution of a range of candidate toxin gene families in anemones. For example, we were able to identify 71 and 90 non-redundant open reading frames with ShKT domains from previously-published, high-quality transcriptome assemblies for Calliactis polypus and Actinia tenebrosa [82], respectively (our unpublished results). From these peptide sequences, we were able to identify canonical ShK (Type I KTxs) toxin peptides with a single domain, as well as a many proteins with multiple ShKT domains, and in some instances ShKT domains with other toxin domains. Proteins with multiple ShKT domains have been identified previously in other anemone species, including Megalactis griffithsi and Anemonia sulcata [79]. Further information about the expression of toxin gene families can be derived by application of state-of-the-art transcriptomic based gene expression studies across nematocyst-rich tissues in anemone species.
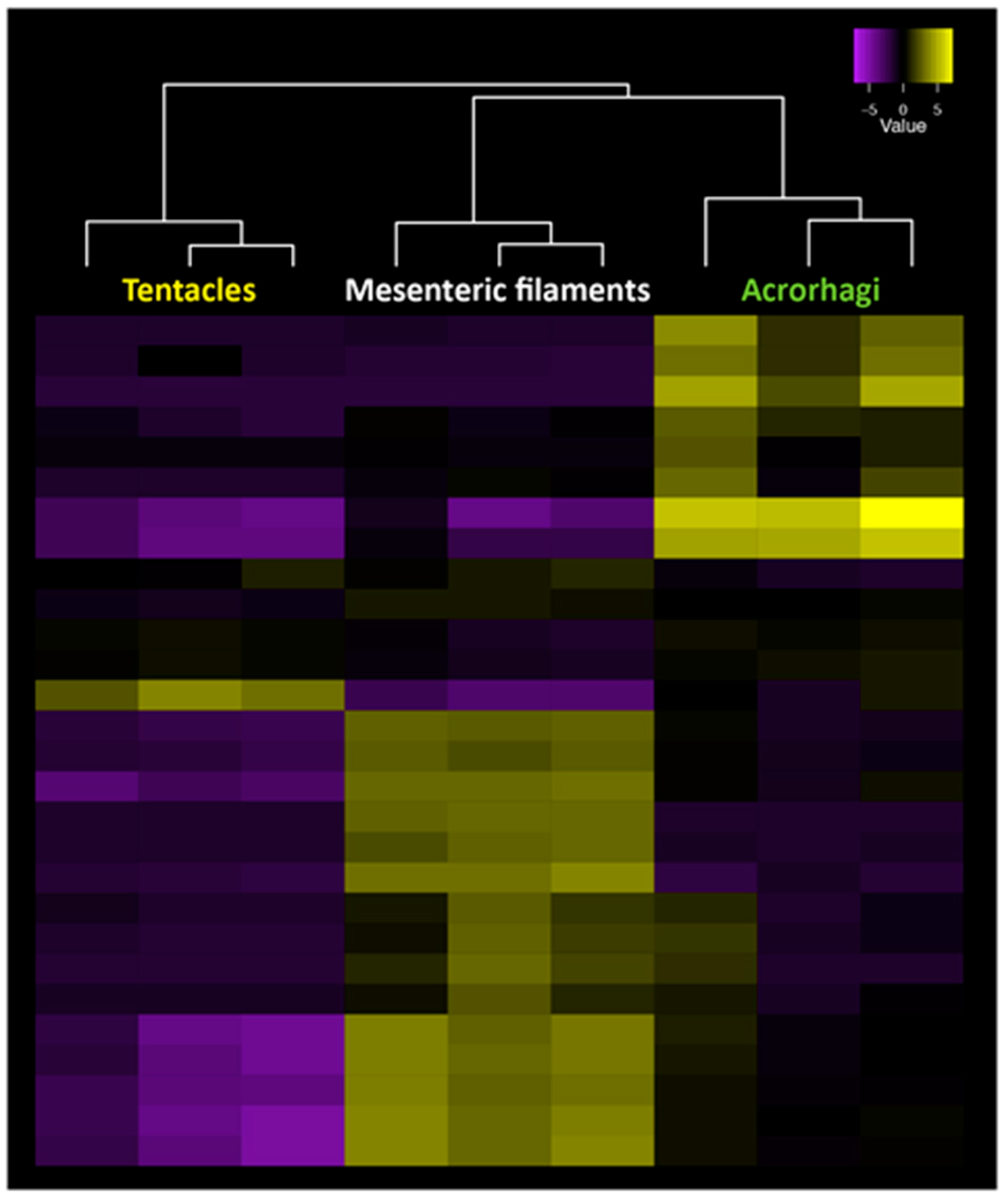
Gene expression analysis across morphological structures in sea anemones has shown that many toxin genes are differentially expressed across tissues, presumably resulting in the production of distinct venom combinations in different structures [79]. In fact, specific genes from toxin gene families are expressed precisely in a region-specific pattern, with different gene copies expressed in specific structures, such as tentacles, acrorhagi or mesenteric filaments, as exemplified for the venom Kunitz peptides in Figure 5. This highlights that different genes from known toxin gene families show strong regionalization in their expression patterns across nematocyst-rich tissues in a pattern consistent with changes in nematocyst morphology and density (our unpublished results). Testing whether different nematocyte populations are contributing directly to the changes in toxin gene expression across tissues will require an examination of single-cell gene expression changes in nematocyte populations isolated from specific tissues. The analysis of gene expression profiles from single cell populations has improved greatly over recent years [84] and has led to a precise understanding of the function of individual cell types in a range of species [85]. Determining single-cell venom gene expression patterns in nematocyte populations will provide unprecedented insight into the evolution of venom composition and how functionally different venoms can be produced by a single animal. While this information is useful in understanding the evolution and composition of venoms, the identification of peptide toxins with new modes of action will require proteomic analyses of the venom.
High-throughput proteomic analyses of sea anemone venoms have been greatly accelerated through the use of mass spectrometry [86] coupled with the availability of complete genome and transcriptome sequences for sea anemones species. MALDI imaging [87,88] also offers the prospect of mapping the tissue distributions of numerous peptides, and thus providing a clue to their possible function in the anemone. Recent proteomic research has assayed venom composition in the nematocytes of a few anemone species [69,86], revealing a high frequency of novel peptides and proteins in nematocysts, and in some cases detecting peptides that had not been identified in earlier proteomics studies [89,90,91]. Many of these represent novel candidate toxins and show little overlap among species, highlighting the potential for proteogenomic approaches to identify previously uncharacterized toxins. Another recent study used a combination of proteomic and transcriptomic techniques to analyse the protein composition of milked venom from the sea anemone Stichodactyla haddoni [39]; the milked venom contained 23 putative toxin families, 12 of which showed no identity to any known peptide or protein in current databases. The identification of these 12 cysteine-rich, but previously unknown, putative toxins further emphasizes that a proteogenomic approach can greatly accelerate the discovery of novel proteins with potential therapeutic applications.
Continual improvements in the efficiency of peptide synthesis and recombinant expression will facilitate the production of such peptides for biological evaluation, although the challenge will remain to define the native disulfide connectivities for novel sequences containing multiple cysteine residues. New peptides need to be assessed against a range of targets, and in multiple functional assays, in order to identify those with the potential to be new therapeutic leads or at least pharmacological tools. For the most part, these assays will focus on mammalian targets, leaving open the intriguing but often neglected question of what function(s) these peptides have in the anemones in which they are identified. As noted above, MALDI imaging studies of peptide distribution in the anemone should assist in identifying their endogenous functions.
It is to be hoped that new bioinformatic approaches, such as weighted gene co-expression network analysis, may provide preliminary information on the function of candidate peptides and proteins. Weighted gene co-expression analysis categorizes genes into groups whose expression levels are highly correlated across samples [92]. This approach has been used to identify gene clusters with functions in development and lipid metabolism, among others [93,94]. In sea anemones, identifying gene clusters with correlated expression levels that contain known toxins may offer a way to detect novel toxin genes with similar or novel functions.
7. Conclusions
Sea anemones remain relatively under-explored as a source of peptide toxins, although that is beginning to change as the transcriptomes and genomes of several species from different geographic regions and different habitats are being investigated. Proteomics studies on sea anemones [39] are more challenging than for other venomous species, where the venom apparatus can be readily milked or dissected, but ongoing enhancements in the resolution and sensitivity of liquid chromatography-mass spectrometry methods will enable characterization of the complete peptide repertoires of even limited quantities of venom when it can be obtained.
The progress of an analogue of ShK through clinical trials for autoimmune diseases emphasizes the promise of this scaffold and, more broadly, of sea anemone peptides. The abundance of ShKT domains in nature and their distribution across different phyla pose questions about the diversity of functions supported by this scaffold. Defining the relationships among sequence, structure and function remains one of the exciting challenges of ongoing studies.
Acknowledgments
R.S.N. acknowledges fellowship support from the Australian National Health and Medical Research Council. This work was supported in part by a grant from the Australian Research Council (LP150100621). We thank Michela Mitchell and Joachim Surm for helpful comments on the manuscript.
Author Contributions
All authors contributed to writing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Beress, L. Biologically active compounds from coelenterates. Pure Appl. Chem. 1982, 54, 1981–1994. [Google Scholar] [CrossRef]
- Norton, R.S. Structure and structure-function relationships of sea anemone proteins that interact with the sodium channel. Toxicon 1991, 29, 1051–1084. [Google Scholar] [CrossRef]
- Honma, T.; Shiomi, K. Peptide toxins in sea anemones: Structural and functional aspects. Mar. Biotechnol. 2006, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, K. Novel peptide toxins recently isolated from sea anemones. Toxicon 2009, 54, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Norton, R.S. Structures of sea anemone toxins. Toxicon 2009, 54, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Norton, R.S. Sea anemone venom peptides. In Handbook of Biologically Active Peptides; Kastin, A.J., Ed.; Elsevier (Academic): San Diego, CA, USA, 2013; pp. 430–436. [Google Scholar]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient venom systems: A review on cnidaria toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [PubMed]
- Tibballs, J. Australian venomous jellyfish, envenomation syndromes, toxins and therapy. Toxicon 2006, 48, 830–859. [Google Scholar] [CrossRef] [PubMed]
- Tibballs, J.; Li, R.; Tibballs, H.A.; Gershwin, L.A.; Winkel, K.D. Australian carybdeid jellyfish causing “Irukandji syndrome”. Toxicon 2012, 59, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Nishikawa, K.; Yuzawa, Y.; Kanie, T.; Mori, H.; Araki, Y.; Hotta, N.; Matsuo, S. Acute renal failure after a sea anemone sting. Am. J. Kidney Dis. 2000, 36, E10. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M. Envenomation by cnidarians and renal injuries. In The Cnidaria, Past, Present and Future: The World of Medusa and Her Sisters; Goffredo, S., Dubinsky, Z., Eds.; Springer: Berlin, Germany, 2016; pp. 623–636. [Google Scholar]
- Gracia Bara, M.T.; Iriarte, P.; Pineda, F. Allergy to Actinia equina and Anemona viridis. Allergy 2006, 61, 1151–1152. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.; Masry, D.; Halstead, B.W. Report of stingings by the sea anemone Triactis producta Klunzinger from Red Sea. Clin. Toxicol. 1970, 3, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, H.; Knop, D. Corals: Indo-Pacific Field Guide. Gorgonians, Soft Corals, Stony Corals, Sea Anemones; IKAN Unterwasserarchiv: ConchBooks, Frankfurt, Germany, 2005. [Google Scholar]
- Mizuno, M.; Nozaki, M.; Morine, N.; Suzuki, N.; Nishikawa, K.; Morgan, B.P.; Matsuo, S. A protein toxin from the sea anemone Phyllodiscus semoni targets the kidney and causes a severe renal injury with predominant glomerular endothelial damage. Am. J. Pathol. 2007, 171, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Fautin, D.G. Structural diversity, systematics, and evolution of cnidae. Toxicon 2009, 54, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.M.; Hessinger, D.A. Cnidocyte mechanoreceptors are tuned to the movements of swimming prey by chemoreceptors. Science 1989, 243, 1589–1591. [Google Scholar] [CrossRef] [PubMed]
- Kass-Simon, G.; Scappaticci, A.A. The behavioral and developmental physiology of nematocysts. Can. J. Zool. 2002, 80, 1772–1794. [Google Scholar] [CrossRef]
- Schama, R.; Mitchell, M.; Sole-Cava, A.M. Actinia ebhayiensis sp. nov., a new species of sea anemone (Anthozoa: Actiniaria: Actiniidae) from South Africa. J. Mar. Biol. Assoc. UK 2012, 92, 885–894. [Google Scholar] [CrossRef]
- Ayre, D.J. Inter-genotype aggression in the solitary sea anemone Actinia tenebrosa. Mar. Biol. 1982, 68, 199–205. [Google Scholar] [CrossRef]
- Reft, A.J. Understanding the Morphology and Distribution of Nematocysts in Sea Anemones and Their Relatives; The Ohio State University: Columbus, OH, USA, 2012. [Google Scholar]
- Macrander, J.; Brugler, M.R.; Daly, M. A RNA-seq approach to identify putative toxins from acrorhagi in aggressive and non-aggressive Anthopleura elegantissima polyps. BMC Genom. 2015, 16, 221. [Google Scholar] [CrossRef] [PubMed]
- Frazao, B.; Vasconcelos, V.; Antunes, A. Sea anemone (Cnidaria, Anthozoa, Actiniaria) toxins: An overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Jungo, F.; Bairoch, A. Tox-Prot, the toxin protein annotation program of the Swiss-Prot protein knowledgebase. Toxicon 2005, 45, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Reimer, N.S.; Yasunobu, C.L.; Yasunobu, K.T.; Norton, T.R. Amino acid sequence of the Anthopleura xanthogrammica heart stimulant, anthopleurin-B. J. Biol. Chem. 1985, 260, 8690–8693. [Google Scholar] [PubMed]
- Monks, S.A.; Pallaghy, P.K.; Scanlon, M.J.; Norton, R.S. Solution structure of the cardiostimulant polypeptide anthopleurin-B and comparison with anthopleurin-A. Structure 1995, 3, 791–803. [Google Scholar] [CrossRef]
- Schweitz, H.; Vincent, J.P.; Barhanin, J.; Frelin, C.; Linden, G.; Hugues, M.; Lazdunski, M. Purification and pharmacological properties of eight sea anemone toxins from Anemonia sulcata, Anthopleura xanthogrammica, Stoichactis giganteus, and Actinodendron plumosum. Biochemistry 1981, 20, 5245–5252. [Google Scholar] [CrossRef] [PubMed]
- Kem, W.R.; Parten, B.; Pennington, M.W.; Price, D.A.; Dunn, B.M. Isolation, characterization, and amino acid sequence of a polypeptide neurotoxin occurring in the sea anemone Stichodactyla helianthus. Biochemistry 1989, 28, 3483–3489. [Google Scholar] [CrossRef] [PubMed]
- Cestele, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Diochot, S.; Baron, A.; Rash, L.D.; Deval, E.; Escoubas, P.; Scarzello, S.; Salinas, M.; Lazdunski, M. A new sea anemone peptide, APETx2, inhibits ASIC3, a major acid-sensitive channel in sensory neurons. EMBO J. 2004, 23, 1516–1525. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Beress, L.; Moller, C.; Mari, F.; Forssmann, W.G.; Tytgat, J. A natural point mutation changes both target selectivity and mechanism of action of sea anemone toxins. FASEB J. 2012, 26, 5141–5151. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.E.; Cristofori-Armstrong, B.; Anangi, R.; Rosengren, K.J.; Lau, C.H.; Mobli, M.; Brust, A.; Alewood, P.F.; King, G.F.; Rash, L.D. Understanding the molecular basis of toxin promiscuity: The analgesic sea anemone peptide APETx2 interacts with acid-sensing ion channel 3 and hERG channels via overlapping pharmacophores. J. Med. Chem. 2014, 57, 9195–9203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, X.S.; Diochot, S.; Lazdunski, M.; Tseng, G.N. APETx1 from sea anemone Anthopleura elegantissima is a gating modifier peptide toxin of the human ether-a-go-go-related potassium channel. Mol. Pharmacol. 2007, 72, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Moreels, L.; Peigneur, S.; Galan, D.T.; De Pauw, E.; Beress, L.; Waelkens, E.; Pardo, L.A.; Quinton, L.; Tytgat, J. APETx4, a novel sea anemone toxin and a modulator of the cancer-relevant potassium channel KV10.1. Mar. Drugs 2017, 15, 287. [Google Scholar] [CrossRef] [PubMed]
- Andreev, Y.A.; Kozlov, S.A.; Koshelev, S.G.; Ivanova, E.A.; Monastyrnaya, M.M.; Kozlovskaya, E.P.; Grishin, E.V. Analgesic compound from sea anemone Heteractis crispa is the first polypeptide inhibitor of vanilloid receptor 1 (TRPV1). J. Biol. Chem. 2008, 283, 23914–23921. [Google Scholar] [CrossRef] [PubMed]
- Nikolaev, M.V.; Dorofeeva, N.A.; Komarova, M.S.; Korolkova, Y.V.; Andreev, Y.A.; Mosharova, I.V.; Grishin, E.V.; Tikhonov, D.B.; Kozlov, S.A. TRPV1 activation power can switch an action mode for its polypeptide ligands. PLoS ONE 2017, 12, e0177077. [Google Scholar] [CrossRef] [PubMed]
- Yeung, S.Y.; Thompson, D.; Wang, Z.; Fedida, D.; Robertson, B. Modulation of Kv3 subfamily potassium currents by the sea anemone toxin BDS: Significance for CNS and biophysical studies. J. Neurosci. 2005, 25, 8735–8745. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Jo, S.; Bean, B.P. Modulation of neuronal sodium channels by the sea anemone peptide BDS-I. J. Neurophysiol. 2012, 107, 3155–3167. [Google Scholar] [CrossRef] [PubMed]
- Madio, B.; Undheim, E.A.B.; King, G.F. Revisiting venom of the sea anemone Stichodactyla haddoni: Omics techniques reveal the complete toxin arsenal of a well-studied sea anemone genus. J. Proteom. 2017, 166, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Babenko, V.V.; Mikov, A.N.; Manuvera, V.A.; Anikanov, N.A.; Kovalchuk, S.I.; Andreev, Y.A.; Logashina, Y.A.; Kornilov, D.A.; Manolov, A.I.; Sanamyan, N.P.; et al. Identification of unusual peptides with new Cys frameworks in the venom of the cold-water sea anemone Cnidopus japonicus. Sci. Rep. 2017, 7, 14534. [Google Scholar] [CrossRef] [PubMed]
- Sintsova, O.; Gladkikh, I.; Chausova, V.; Monastyrnaya, M.; Anastyuk, S.; Chernikov, O.; Yurchenko, E.; Aminin, D.; Isaeva, M.; Leychenko, E.; et al. Peptide fingerprinting of the sea anemone Heteractis magnifica mucus revealed neurotoxins, Kunitz-type proteinase inhibitors and a new β-defensin α-amylase inhibitor. J. Proteom. 2018, 173, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Cahalan, M.D.; Chandy, K.G. The functional network of ion channels in T lymphocytes. Immunol. Rev. 2009, 231, 59–87. [Google Scholar] [CrossRef] [PubMed]
- Chandy, K.G.; Wulff, H.; Beeton, C.; Pennington, M.; Gutman, G.A.; Cahalan, M.D. K+ channels as targets for specific immunomodulation. Trends Pharmacol. Sci. 2004, 25, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Chandy, K.G.; Norton, R.S. Peptide blockers of Kv1.3 channels in T cells as therapeutics for autoimmune disease. Curr. Opin. Chem. Biol. 2017, 38, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Norton, R.S.; Chandy, K.G. Venom-derived peptide inhibitors of voltage-gated potassium channels. Neuropharmacology 2017, 127, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide therapeutics from venom: Current status and potential. Bioorg. Med. Chem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bartok, A.; Toth, A.; Somodi, S.; Szanto, T.G.; Hajdu, P.; Panyi, G.; Varga, Z. Margatoxin is a non-selective inhibitor of human Kv1.3 K+ channels. Toxicon 2014, 87, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, O.; Sotolongo, V.; Amor, A.M.; Stocklin, R.; Anderson, A.J.; Harvey, A.L.; Engstrom, A.; Wernstedt, C.; Karlsson, E. Characterization of a potassium channel toxin from the Caribbean Sea anemone Stichodactyla helianthus. Toxicon 1995, 33, 603–613. [Google Scholar] [CrossRef]
- Pohl, J.; Hubalek, F.; Byrnes, M.E.; Nielsen, K.R.; Woods, A.; Pennington, M.W. Assignment of the three disulfide bonds in ShK toxin, a potent potassium channel blocker from the sea anemone Stichodactyla helianthus. Lett. Pept. Sci. 1995, 1, 291–297. [Google Scholar] [CrossRef]
- Tudor, J.E.; Pallaghy, P.K.; Pennington, M.W.; Norton, R.S. Solution structure of ShK toxin, a novel potassium channel inhibitor from a sea anemone. Nat. Struct. Biol. 1996, 3, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Dauplais, M.; Lecoq, A.; Song, J.; Cotton, J.; Jamin, N.; Gilquin, B.; Roumestand, C.; Vita, C.; de Medeiros, C.L.; Rowan, E.G.; et al. On the convergent evolution of animal toxins. Conservation of a diad of functional residues in potassium channel-blocking toxins with unrelated structures. J. Biol. Chem. 1997, 272, 4302–4309. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Byrnes, M.E.; Zaydenberg, I.; Khaytin, I.; de Chastonay, J.; Krafte, D.S.; Hill, R.; Mahnir, V.M.; Volberg, W.A.; Gorczyca, W.; et al. Chemical synthesis and characterization of ShK toxin: A potent potassium channel inhibitor from a sea anemone. Int. J. Pept. Protein Res. 1995, 46, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Kalman, K.; Pennington, M.W.; Lanigan, M.D.; Nguyen, A.; Rauer, H.; Mahnir, V.; Paschetto, K.; Kem, W.R.; Grissmer, S.; Gutman, G.A.; et al. ShK-Dap22, a potent Kv1.3-specific immunosuppressive polypeptide. J. Biol. Chem. 1998, 273, 32697–32707. [Google Scholar] [CrossRef] [PubMed]
- Chi, V.; Pennington, M.W.; Norton, R.S.; Tarcha, E.J.; Londono, L.M.; Sims-Fahey, B.; Upadhyay, S.K.; Lakey, J.T.; Iadonato, S.; Wulff, H.; et al. Development of a sea anemone toxin as an immunomodulator for therapy of autoimmune diseases. Toxicon 2012, 59, 529–546. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Beeton, C.; Galea, C.A.; Smith, B.J.; Chi, V.; Monaghan, K.P.; Garcia, A.; Rangaraju, S.; Giuffrida, A.; Plank, D.; et al. Engineering a stable and selective peptide blocker of the Kv1.3 channel in T lymphocytes. Mol. Pharmacol. 2009, 75, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Beeton, C.; Wulff, H.; Barbaria, J.; Clot-Faybesse, O.; Pennington, M.; Bernard, D.; Cahalan, M.D.; Chandy, K.G.; Beraud, E. Selective blockade of T lymphocyte K+ channels ameliorates experimental autoimmune encephalomyelitis, a model for multiple sclerosis. Proc. Natl. Acad. Sci. USA 2001, 98, 13942–13947. [Google Scholar] [CrossRef] [PubMed]
- Beeton, C.; Wulff, H.; Standifer, N.E.; Azam, P.; Mullen, K.M.; Pennington, M.W.; Kolski-Andreaco, A.; Wei, E.; Grino, A.; Counts, D.R.; et al. Kv1.3 channels are a therapeutic target for T cell-mediated autoimmune diseases. Proc. Natl. Acad. Sci. USA 2006, 103, 17414–17419. [Google Scholar] [CrossRef] [PubMed]
- Tarcha, E.J.; Chi, V.; Munoz-Elias, E.J.; Bailey, D.; Londono, L.M.; Upadhyay, S.K.; Norton, K.; Banks, A.; Tjong, I.; Nguyen, H.; et al. Durable pharmacological responses from the peptide drug ShK-186, a specific Kv1.3 channel inhibitor that suppresses T cell mediators of autoimmune disease. J. Pharmacol. Exp. Ther. 2012, 342, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Tarcha, E.J.; Olsen, C.M.; Probst, P.; Peckham, D.; Munoz-Elias, E.J.; Kruger, J.G.; Iadonato, S.P. Safety and pharmacodynamics of dalazatide, a Kv1.3 channel inhibitor, in the treatment of plaque psoriasis: A randomized phase 1b trial. PLoS ONE 2017, 12, e0180762. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Harunur Rashid, M.; Tajhya, R.B.; Beeton, C.; Kuyucak, S.; Norton, R.S. A C-terminally amidated analogue of ShK is a potent and selective blocker of the voltage-gated potassium channel Kv1.3. FEBS Lett. 2012, 586, 3996–4001. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Heinzelmann, G.; Huq, R.; Tajhya, R.B.; Chang, S.C.; Chhabra, S.; Pennington, M.W.; Beeton, C.; Norton, R.S.; Kuyucak, S. A potent and selective peptide blocker of the Kv1.3 channel: Prediction from free-energy simulations and experimental confirmation. PLoS ONE 2013, 8, e78712. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Huq, R.; Chhabra, S.; Beeton, C.; Pennington, M.W.; Smith, B.J.; Norton, R.S. N-Terminally extended analogues of the K+ channel toxin from Stichodactyla helianthus as potent and selective blockers of the voltage-gated potassium channel Kv1.3. FEBS J. 2015, 282, 2247–2259. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Chang, S.C.; Chauhan, S.; Huq, R.; Tajhya, R.B.; Chhabra, S.; Norton, R.S.; Beeton, C. Development of highly selective Kv1.3-blocking peptides based on the sea anemone peptide ShK. Mar. Drugs 2015, 13, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.K.; Qian, Y.X.; Liu, B.; Elliott, R.; Aral, J.; Park, C.; Zhang, X.; Stenkilsson, M.; Salyers, K.; Rose, M.; et al. Pharmaceutical optimization of peptide toxins for ion channel targets: Potent, selective, and long-lived antagonists of Kv1.3. J. Med. Chem. 2015, 58, 6784–6802. [Google Scholar] [CrossRef] [PubMed]
- Cotton, J.; Crest, M.; Bouet, F.; Alessandri, N.; Gola, M.; Forest, E.; Karlsson, E.; Castaneda, O.; Harvey, A.L.; Vita, C.; et al. A potassium-channel toxin from the sea anemone Bunodosoma granulifera, an inhibitor for KV1 channels. Revision of the amino acid sequence, disulfide-bridge assignment, chemical synthesis, and biological activity. Eur. J. Biochem. 1997, 244, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Schweitz, H.; Bruhn, T.; Guillemare, E.; Moinier, D.; Lancelin, J.M.; Beress, L.; Lazdunski, M. Kalicludines and kaliseptine. Two different classes of sea anemone toxins for voltage sensitive K+ channels. J. Biol. Chem. 1995, 270, 25121–25126. [Google Scholar] [CrossRef] [PubMed]
- Gendeh, G.S.; Young, L.C.; de Medeiros, C.L.; Jeyaseelan, K.; Harvey, A.L.; Chung, M.C. A new potassium channel toxin from the sea anemone Heteractis magnifica: Isolation, cDNA cloning, and functional expression. Biochemistry 1997, 36, 11461–11471. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, O.; Harvey, A.L. Discovery and characterization of cnidarian peptide toxins that affect neuronal potassium ion channels. Toxicon 2009, 54, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Rachamim, T.; Morgenstern, D.; Aharonovich, D.; Brekhman, V.; Lotan, T.; Sher, D. The dynamically evolving nematocyst content of an anthozoan, a scyphozoan, and a hydrozoan. Mol. Biol. Evol. 2015, 32, 740–753. [Google Scholar] [CrossRef] [PubMed]
- Ponce, D.; Brinkman, D.L.; Potriquet, J.; Mulvenna, J. Tentacle transcriptome and venom proteome of the pacific sea nettle, Chrysaora fuscescens (Cnidaria: Scyphozoa). Toxins 2016, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Rangaraju, S.; Khoo, K.K.; Feng, Z.P.; Crossley, G.; Nugent, D.; Khaytin, I.; Chi, V.; Pham, C.; Calabresi, P.; Pennington, M.W.; et al. Potassium channel modulation by a toxin domain in matrix metalloprotease 23. J. Biol. Chem. 2010, 285, 9124–9136. [Google Scholar] [CrossRef] [PubMed]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Galea, C.A.; Nguyen, H.M.; George Chandy, K.; Smith, B.J.; Norton, R.S. Domain structure and function of matrix metalloprotease 23 (MMP23): Role in potassium channel trafficking. Cell. Mol. Life Sci. 2014, 71, 1191–1210. [Google Scholar] [CrossRef] [PubMed]
- Faraco, J.; Bashir, M.; Rosenbloom, J.; Francke, U. Characterization of the human gene for microfibril-associated glycoprotein (MFAP2), assignment to chromosome 1p36.1-p35, and linkage to D1S170. Genomics 1995, 25, 630–637. [Google Scholar] [CrossRef]
- Gibbs, G.M.; Scanlon, M.J.; Swarbrick, J.; Curtis, S.; Gallant, E.; Dulhunty, A.F.; O’Bryan, M.K. The cysteine-rich secretory protein domain of Tpx-1 is related to ion channel toxins and regulates ryanodine receptor Ca2+ signaling. J. Biol. Chem. 2006, 281, 4156–4163. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, S.; Chang, S.C.; Nguyen, H.M.; Huq, R.; Tanner, M.R.; Londono, L.M.; Estrada, R.; Dhawan, V.; Chauhan, S.; Upadhyay, S.K.; et al. Kv1.3 channel-blocking immunomodulatory peptides from parasitic worms: Implications for autoimmune diseases. FASEB J. 2014, 28, 3952–3964. [Google Scholar] [CrossRef] [PubMed]
- Krishnarjuna, B.; MacRaild, C.A.; Sunanda, P.; Morales, R.A.V.; Peigneur, S.; Macrander, J.; Yu, H.H.; Daly, M.; Raghothama, S.; Dhawan, V.; et al. Structure, folding and stability of a minimal homologue from Anemonia sulcata of the sea anemone potassium channel blocker ShK. Peptides 2018, 99, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Baumgarten, S.; Simakov, O.; Esherick, L.Y.; Liew, Y.J.; Lehnert, E.M.; Michell, C.T.; Li, Y.; Hambleton, E.A.; Guse, A.; Oates, M.E.; et al. The genome of Aiptasia, a sea anemone model for coral symbiosis. Proc. Natl. Acad. Sci. USA 2015, 112, 11893–11898. [Google Scholar] [CrossRef] [PubMed]
- Macrander, J.; Broe, M.; Daly, M. Tissue-specific venom composition and differential gene expression in sea anemones. Genome Biol. Evol. 2016, 8, 2358–2375. [Google Scholar] [CrossRef] [PubMed]
- Stewart, Z.K.; Pavasovic, A.; Hock, D.H.; Prentis, P.J. Transcriptomic investigation of wound healing and regeneration in the cnidarian Calliactis polypus. Sci. Rep. 2017, 7, 41458. [Google Scholar] [CrossRef] [PubMed]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V.; et al. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 2007, 317, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Van der Burg, C.A.; Prentis, P.J.; Surm, J.M.; Pavasovic, A. Insights into the innate immunome of actiniarians using a comparative genomic approach. BMC Genom. 2016, 17, 850. [Google Scholar] [CrossRef] [PubMed]
- Macrander, J.; Daly, M. Evolution of the cytolytic pore-forming proteins (actinoporins) in sea anemones. Toxins 2016, 8, 368. [Google Scholar] [CrossRef] [PubMed]
- Linnarsson, S.; Teichmann, S.A. Single-cell genomics: Coming of age. Genome Biol. 2016, 17, 97. [Google Scholar] [CrossRef] [PubMed]
- Gawad, C.; Koh, W.; Quake, S.R. Single-cell genome sequencing: Current state of the science. Nat. Rev. Genet. 2016, 17, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Praher, D.; Schlesinger, A.; Ayalon, A.; Tal, Y.; Technau, U. Analysis of soluble protein contents from the nematocysts of a model sea anemone sheds light on venom evolution. Mar. Biotechnol. 2013, 15, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.L.; Hamilton, B.R.; Madio, B.; Morales, R.A.V.; Tonkin-Hill, G.Q.; Papenfuss, A.T.; Purcell, A.W.; King, G.F.; Undheim, E.A.B.; Norton, R.S. The use of imaging mass spectrometry to study peptide toxin distribution in Australian sea anemones. Aust. J. Chem. 2017, 70, 1235–1237. [Google Scholar] [CrossRef]
- Undheim, E.A.; Hamilton, B.R.; Kurniawan, N.D.; Bowlay, G.; Cribb, B.W.; Merritt, D.J.; Fry, B.G.; King, G.F.; Venter, D.J. Production and packaging of a biological arsenal: Evolution of centipede venoms under morphological constraint. Proc. Natl. Acad. Sci. USA 2015, 112, 4026–4031. [Google Scholar] [CrossRef] [PubMed]
- Zaharenko, A.J.; Ferreira, W.A., Jr.; Oliveira, J.S.; Richardson, M.; Pimenta, D.C.; Konno, K.; Portaro, F.C.; de Freitas, J.C. Proteomics of the neurotoxic fraction from the sea anemone Bunodosoma cangicum venom: Novel peptides belonging to new classes of toxins. Comp. Biochem. Physiol. Part D 2008, 3, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.A.; Cassoli, J.S.; Sa, F.; Dong, Z.Q.; de Freitas, J.C.; Pimenta, A.M.; de Lima, M.E.; Konno, K.; Lee, S.M.; Garateix, A.; et al. Peptide fingerprinting of the neurotoxic fractions isolated from the secretions of sea anemones Stichodactyla helianthus and Bunodosoma granulifera. New members of the APETx-like family identified by a 454 pyrosequencing approach. Peptides 2012, 34, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Cassoli, J.S.; Verano-Braga, T.; Oliveira, J.S.; Montandon, G.G.; Cologna, C.T.; Peigneur, S.; Pimenta, A.M.; Kjeldsen, F.; Roepstorff, P.; Tytgat, J.; et al. The proteomic profile of Stichodactyla duerdeni secretion reveals the presence of a novel O-linked glycopeptide. J. Proteom. 2013, 87, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Castellani, L.W.; Zhou, Z.; Paul, E.; Davis, R.; Schadt, E.E.; Lusis, A.J.; Horvath, S.; Mehrabian, M. A systems genetic analysis of high density lipoprotein metabolism and network preservation across mouse models. Biochim. Biophys. Acta 2012, 1821, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Huang, K.; Cai, C.; Cai, L.; Jiang, C.Y.; Feng, Y.; Liu, Z.; Zeng, Q.; Cheng, L.; Sun, Y.E.; et al. Genetic programs in human and mouse early embryos revealed by single-cell RNA sequencing. Nature 2013, 500, 593–597. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sea anemone species that are locally abundant in the waters off south-eastern Queensland, Australia; these species have been largely unexplored for their toxin contents and are currently under investigation in our labs to identify new peptide and protein toxins with potentially novel functions. (A) Aulactinia veratra (B) undescribed species of Anthopleura (C) Calliactis polypus (D) Actinia australiensis. Photo credit: Ana Pavasovic.
Figure 1.
Sea anemone species that are locally abundant in the waters off south-eastern Queensland, Australia; these species have been largely unexplored for their toxin contents and are currently under investigation in our labs to identify new peptide and protein toxins with potentially novel functions. (A) Aulactinia veratra (B) undescribed species of Anthopleura (C) Calliactis polypus (D) Actinia australiensis. Photo credit: Ana Pavasovic.

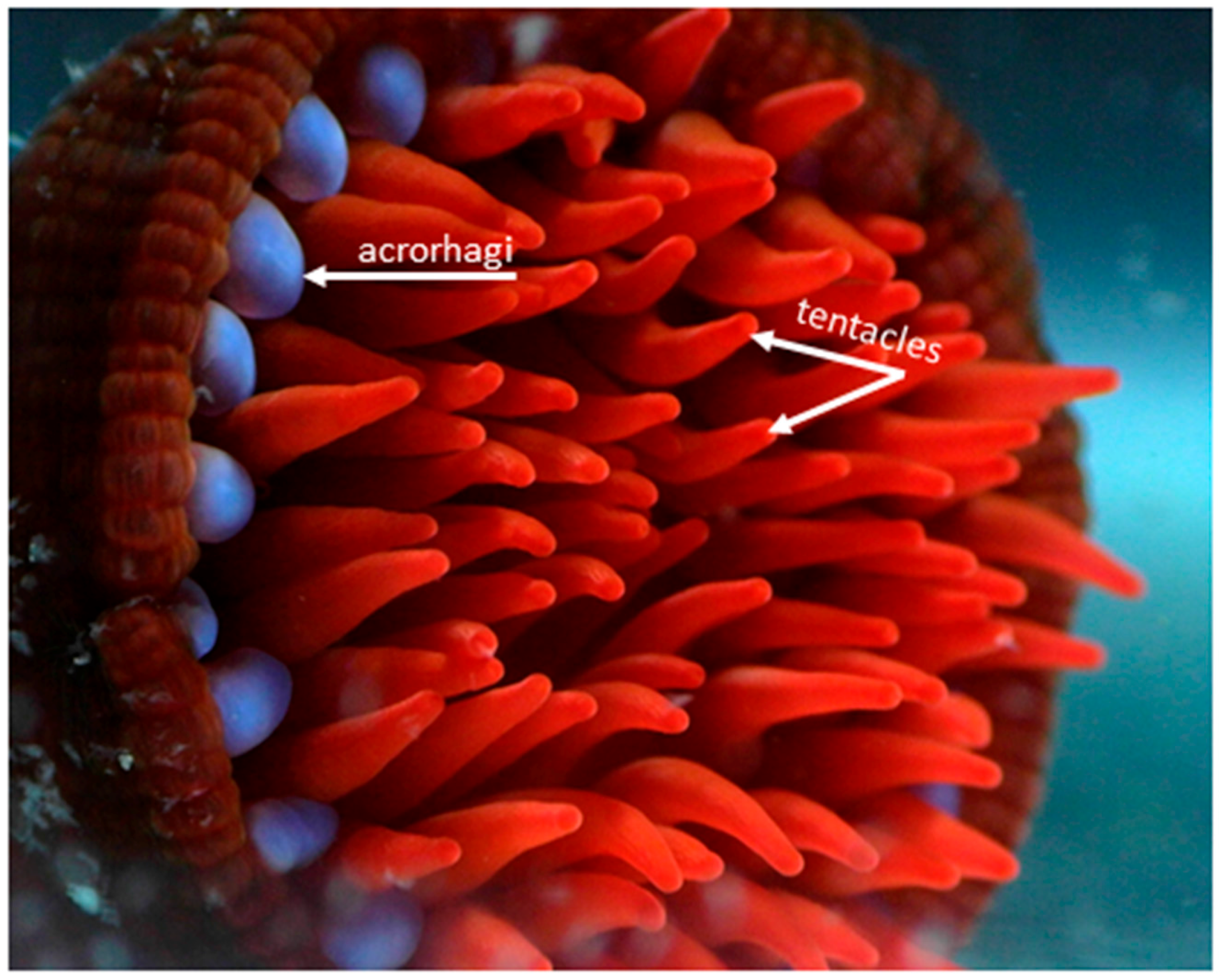
Figure 2.
This photograph shows the bright blue acrorhagi used in intraspecific combat [20] and red tentacles used in prey capture and defence against predators of the Australian sea anemone, Actinia tenebrosa. Both structures have a high density of nematocysts, but they have different nematocysts complements (for example, holotrichs are abundant in acrorhagi and basitrichs in tentacles). Photo credit: Chloe van der Burg.
Figure 2.
This photograph shows the bright blue acrorhagi used in intraspecific combat [20] and red tentacles used in prey capture and defence against predators of the Australian sea anemone, Actinia tenebrosa. Both structures have a high density of nematocysts, but they have different nematocysts complements (for example, holotrichs are abundant in acrorhagi and basitrichs in tentacles). Photo credit: Chloe van der Burg.

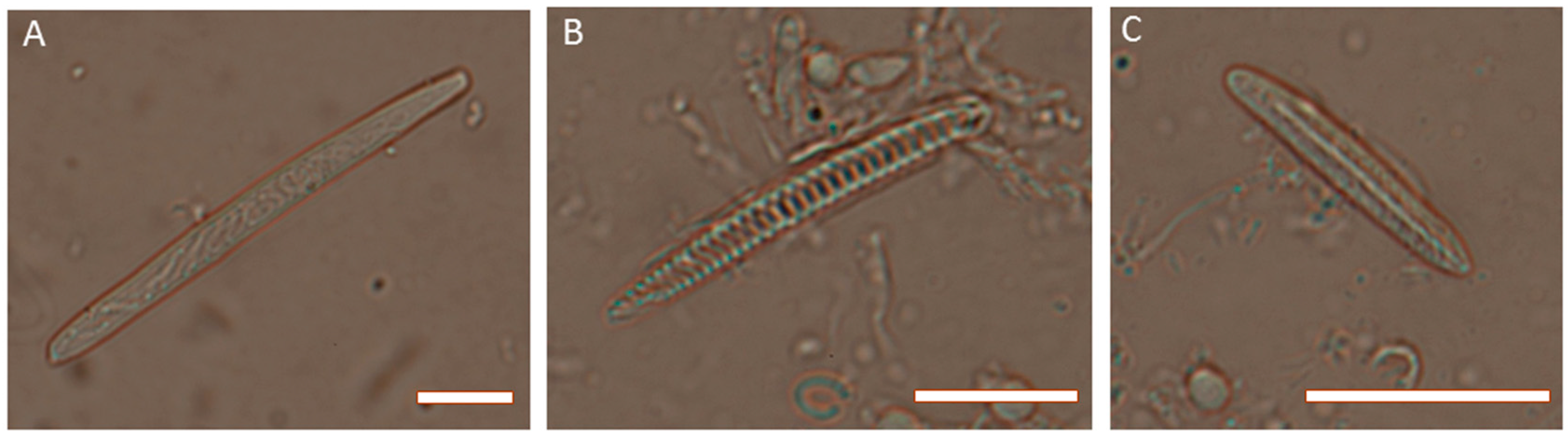
Figure 3.
Photomicrographs of cnidae from Actinia tenebrosa. (A) holotrich from acrorhagi; (B) spirocyst from tentacles; (C) basitrich from tentacles. Scale bars are 5 µm in each case. Photo credit: Michela Mitchell.
Figure 3.
Photomicrographs of cnidae from Actinia tenebrosa. (A) holotrich from acrorhagi; (B) spirocyst from tentacles; (C) basitrich from tentacles. Scale bars are 5 µm in each case. Photo credit: Michela Mitchell.
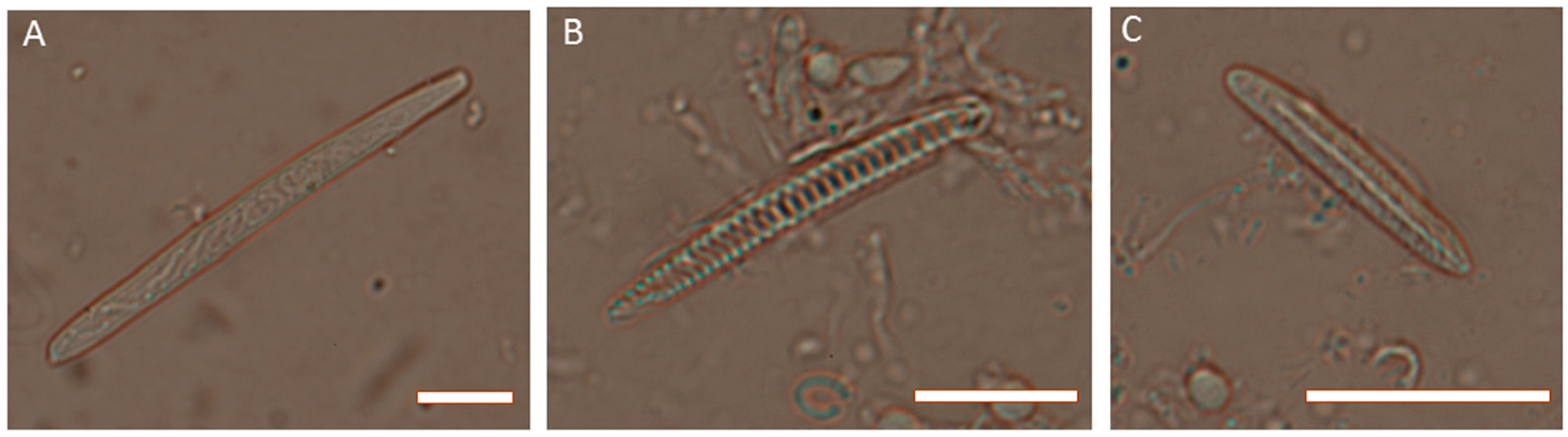
Figure 4.
(A) Amino acid sequences of ShK (UniProt entry P29187) [48], BgK (P29186) [65], AsK (Q9TWG1), also known as kaliseptine [66], and HmK (O16846) [67] with the three disulfide bonds indicated. (B) Structures of ShK (pdb id 1ROO) [50] and BgK (pdb id 1BGK) [51] are shown, with backbones in lightblue and disulfides in orange. The side chains of the Lys and Tyr that constitute the functional dyad [51] in each peptide are shown in blue and magenta, respectively. This dyad is displayed on a helical scaffold in ShK but a β-sheet in the scorpion toxins charybdotoxin and margatoxin (not shown).
Figure 4.
(A) Amino acid sequences of ShK (UniProt entry P29187) [48], BgK (P29186) [65], AsK (Q9TWG1), also known as kaliseptine [66], and HmK (O16846) [67] with the three disulfide bonds indicated. (B) Structures of ShK (pdb id 1ROO) [50] and BgK (pdb id 1BGK) [51] are shown, with backbones in lightblue and disulfides in orange. The side chains of the Lys and Tyr that constitute the functional dyad [51] in each peptide are shown in blue and magenta, respectively. This dyad is displayed on a helical scaffold in ShK but a β-sheet in the scorpion toxins charybdotoxin and margatoxin (not shown).

Figure 5.
Gene expression patterns for all gene copies of the venom Kunitz gene family across tentacles, mesenteric filaments and acrorhagi in A. tenebrosa. Tissue-specific expression can be observed among copies across the three tissues, with significantly upregulated transcripts in yellow and downregulated transcripts in purple (our unpublished data).
Figure 5.
Gene expression patterns for all gene copies of the venom Kunitz gene family across tentacles, mesenteric filaments and acrorhagi in A. tenebrosa. Tissue-specific expression can be observed among copies across the three tissues, with significantly upregulated transcripts in yellow and downregulated transcripts in purple (our unpublished data).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characterized toxin protein families identified in sea anemone species in the current ToxProt database, and the number of proteins from each toxin protein family identified in each of the sea anemone superfamilies. Act—Actinioidea; Edw—Edwardsioidea; Met—Metridioidea; Acti—Actinernoidea.
Table 1.
Characterized toxin protein families identified in sea anemone species in the current ToxProt database, and the number of proteins from each toxin protein family identified in each of the sea anemone superfamilies. Act—Actinioidea; Edw—Edwardsioidea; Met—Metridioidea; Acti—Actinernoidea.
| Toxin Protein Family | Anemone Superfamily | |||
|---|---|---|---|---|
| Act | Edw | Met | Acti | |
| Actinoporin family, Sea anemone subfamily | 25 | 0 | 5 | 0 |
| Cnidaria small cysteine-rich protein (SCRiP) family | 2 | 0 | 1 | 0 |
| Peptidase M12A family | 0 | 1 | 0 | 0 |
| Phospholipase A2 family | 3 | 0 | 1 | 0 |
| Sea anemone 8 toxin family | 5 | 0 | 0 | 0 |
| Sea anemone short toxin (type III) family | 8 | 0 | 0 | 0 |
| Sea anemone sodium channel inhibitory toxin family | 0 | 0 | 3 | 0 |
| Sea anemone sodium channel inhibitory toxin family, Type I subfamily | 52 | 0 | 0 | 0 |
| Sea anemone sodium channel inhibitory toxin family, Type II subfamily | 13 | 9 | 0 | 1 |
| Sea anemone structural class 9a family | 6 | 0 | 0 | 0 |
| Sea anemone type 1 potassium channel toxin family, Type 1a subfamily | 9 | 0 | 0 | 0 |
| Sea anemone type 1 potassium channel toxin family, Type 1b subfamily | 11 | 0 | 1 | 0 |
| Sea anemone type 3 (BDS) potassium channel toxin family | 32 | 0 | 0 | 0 |
| Sea anemone type 5 potassium channel toxin family | 1 | 1 | 1 | 0 |
| Venom Kunitz-type family, Sea anemone type 2 potassium channel toxin subfamily | 26 | 0 | 0 | 0 |
| Unknown | 15 | 0 | 4 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
J. Prentis, P.; Pavasovic, A.; S. Norton, R. Sea Anemones: Quiet Achievers in the Field of Peptide Toxins. Toxins 2018, 10, 36. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10010036
AMA Style
J. Prentis P, Pavasovic A, S. Norton R. Sea Anemones: Quiet Achievers in the Field of Peptide Toxins. Toxins. 2018; 10(1):36. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10010036
Chicago/Turabian StyleJ. Prentis, Peter, Ana Pavasovic, and Raymond S. Norton. 2018. "Sea Anemones: Quiet Achievers in the Field of Peptide Toxins" Toxins 10, no. 1: 36. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10010036
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.