Insect Antimicrobial Peptides, a Mini Review


1
College of Life Science, Yangtze University, Jingzhou 434025, China
2
Department of Chemistry, Faculty of Science, University of Hradec Kralove, 500 03 Hradec Kralove, Czech Republic
3
Department of Radiology and Toxicology, Faculty of Health and Social Studies, University of South Bohemia, 370 05 Ceske Budejovice, Czech Republic
4
Biomedical Research Centre, University Hospital, 500 03 Hradec Kralove, Czech Republic
*
Author to whom correspondence should be addressed.
Toxins 2018, 10(11), 461; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110461
Submission received: 20 September 2018
/
Revised: 1 November 2018
/
Accepted: 5 November 2018
/
Published: 8 November 2018
Abstract
:Antimicrobial peptides (AMPs) are crucial effectors of the innate immune system. They provide the first line of defense against a variety of pathogens. AMPs display synergistic effects with conventional antibiotics, and thus present the potential for combined therapies. Insects are extremely resistant to bacterial infections. Insect AMPs are cationic and comprise less than 100 amino acids. These insect peptides exhibit an antimicrobial effect by disrupting the microbial membrane and do not easily allow microbes to develop drug resistance. Currently, membrane mechanisms underlying the antimicrobial effects of AMPs are proposed by different modes: the barrel-stave mode, toroidal-pore, carpet, and disordered toroidal-pore are the typical modes. Positive charge quantity, hydrophobic property and the secondary structure of the peptide are important for the antibacterial activity of AMPs. At present, several structural families of AMPs from insects are known (defensins, cecropins, drosocins, attacins, diptericins, ponericins, metchnikowins, and melittin), but new AMPs are frequently discovered. We reviewed the biological effects of the major insect AMPs. This review will provide further information that facilitates the study of insect AMPs and shed some light on novel microbicides.
Keywords:
antimicrobial peptides; AMP; Structure-activity relationship; modification; mechanism of actionKey Contribution: The biological effects, especially the antibacterial activity of the major insect antimicrobial peptides (AMPs) are reviewed.
1. Introduction
Antimicrobial peptides (AMPs) are multifunctional components of the innate immune defense systems in prokaryotic and eukaryotic organisms [1]. Based on amino acid substitutions, AMPs are divided into several subgroups. They generally consist of between 12 and 50 amino acids and are divided into subgroups by their amino acid composition and structure. Some AMPs can be as short as 7 to 100 amino acids [2]. The hydrophobic part of their molecule generally takes up more than 50% of amino acids residues. The secondary structure of AMPs follows four themes: (1) α-helical due to the presence of coiled conformation; (2) β-stranded; (3) β-hairpin or loop; and (4) extended conformation [3].
AMPs have a range of antibacterial, antifungal, and antiviral activities. They have a promising capacity in the therapeutic and prophylactic applications [4,5]. Moreover, AMP-derived drugs are administered as topical formulations to treat skin and wound infections [6]. Some AMPs also show anticancer effects or have anticancer properties [7]. Aurein, for example, is highly effective against around 50 different cancer cell lines and displays little toxicity [8]. Bacteria do not develop resistance to AMPS as easily as to traditional antibiotics. These peptides can physically disrupt microbial cellular membranes and therefore kill a broad spectrum of pathogenic microorganisms. Thus, the microbial membrane is usually considered the primary target of AMPs [9,10]. Moreover, their outstanding membrane disruptive activity makes these peptides ideal candidates for combined therapies with conventional antibiotics [11]. AMPs can facilitate more antibiotic molecules entering the microorganism cytoplasm, where they can interact with their target (Figure 1).
AMPs kill bacteria via a variety of mechanisms including membrane disruption, interference with bacterial metabolism, and targeting of cytoplasmic components [5,6,10,12,13,14]. The primary contact between an AMP and the target bacterium occurs via an electrostatic or hydrophobic interaction, which is strongly dependent on the lipid composition of the bacterial membrane [15,16]. AMPs are capable of interacting with the surface of the cell membrane to alter the permeability of the membrane [10]. After AMPs interact with the cell membrane, the formed transmembrane potential affects the osmotic pressure balance [6]. In short, the interaction between the AMPs and the membrane is directly related to the antibacterial activity of the AMPs. At present, there are at least four modes of action commonly used to describe the membrane activity of AMPs: Barrel-stave, carpet, toroidal-pore, and disordered toroidal-pore [10,12]. For all these modes, a threshold concentration is required to conduct the antibacterial effect [10]. AMPs can also disrupt intracellular enzymes and DNA when they translocate into the pathogens [5]. The detailed explanation of these modes can be read in our recent review [6] as well other publications [5,10,12,13]. Regarding the membrane activity of AMPs, some issues need to be considered. For example, whether there is a specific membrane receptor, and whether there are other factors synergistically working in this context. The mechanism of action of different AMPs may be variable, and further research is needed.
AMPs can be classified into many types, based on their secondary structures in liquid media [17,18]. The β-sheet peptides contain a disulfide bond that stabilizes the structure, and helps the AMPs to cross the cell membrane. In addition to the β-sheet structure, AMPs also form an α-helical structure, and contain a cysteine in the peptide to form an intramolecular disulfide bridge [19,20]. Due to the presence of hydrophobic groups, the peptide chain forms a polymer by hydrophobic interaction to increase the affinity for cell membranes [6,21,22,23]. The optimum antibacterial activity appears to be a balance between charge density, hydrophobic character, and polymer chain length [24,25,26]. Increasing the number of positively charged amino acids or changing their position in the peptide chain can affect the secondary structure of the AMPs, thereby further affecting their antibacterial activity. Thus, the combination of charge, hydrophobicity, and length of the peptide is important for the antimicrobial activity of AMPs [27,28,29].
It is well known that insects are extremely resistant to bacterial infections. They can produce a wide range of proteins and peptides as a first line of defense against pathogen infection [14]. Insects activate immune systems, or directly target bacteria and viruses, to combat pathogens. We have previously reviewed the chemical and biological properties of marine AMPs [6]. In this review, we present characteristic and potential medical applications of insect origin peptides with antimicrobial activity. We especially focus on a large group of AMPs that are present both in ancient and recent insects: Defensins, cecropins, attacins, lebocins, dipterins, ponericins, jelleines, and others. By so doing, we will provide a new perspective on the function and biological effects of insect AMPs as well as their use in medicine.
2. Insect Antimicrobial Peptides
Insect AMPs are divided into three groups based on their amino acid sequence and structures: (a) Cecropins, the linear peptides with α-helix but lack cysteine residues; (b) Defensins with 6–8 conserved cysteine residues, have a stabilizing array of 3 or 4 disulfide bridges and 3 domains consisting in a flexible amino-terminal loop; and (c) peptides with an overrepresentation of Proline and/or Glycine residues [30]. The most explored insect AMPs are cecropins, drosocin, attacins, diptericins, defensins, ponericins, drosomycin and metchnikowin. However, more new peptides can still be discovered [31,32]. Most glycine-rich and proline-rich peptides are active against Gram-negative strains of bacteria [33]. Defensins can selectively kill Gram-positive bacteria, whereas cecropins are active against both types [12]. Iinsect AMPs are very potent since their IC50 ranges in the submicromolar or low micromolar range. Currently, there are still no insect-derived AMPs on the market yet. However, we have no doubt that insect AMPs can be exploited as an alternative to antibiotics [12].
2.1. Defensins
Defensins are a family of small, variable cationic arginine-rich peptides [34]. They are not specific to insects, and more than 300 defensins have been identified so far. Defensin peptides are ancient natural antibiotics with strong antimicrobial activity against a range of microorganisms [35,36]. They consist of 18–45 amino acids with 6–8 conserved cysteine residues [36]. Classic defensins (α-defensins) contain 29–35 amino acids, and the insect defensins contain 29–34 amino acids. The molecule of defensin is usually stabilized by three disulfide bonds and, a β-hairpin is their major structural feature [37]. Defensins bind to the cell membrane or form pore-like membrane defects to efflux of essential ions and nutrients [38,39].
Insect defensins are inducible antibacterial peptides with activity against both Gram-positive and Gram-negative bacteria [40]. They are highly effective against Gram-positive bacteria [41], including human pathogenic bacteria such as Staphylococcus aureus. However, these peptides are less effective against Gram-negative bacteria [42]. Insect defensins are isolated from insect orders such as Diptera, Hymenoptera, Coleoptera, Trichoptera, Hemiptera, and Odonata [1,40]. All types of AMPs are reported in lepidopteran insects, except for the insect defensins [43]. Defensin from rabbit neutrophils exerts potent bactericidal activity against the multi-drug-resistant (MDR) strains of Pseudomonas aeruginosa [44].
Royalisin is isolated from the royal jelly of Apis mellifera. It consists of 51 amino acids (VTCDLLSFKQVNDSACAANCLSLGKAGGHCEKGVCICRKTSFKDLWDKYF-NH2), in which six cysteine residues form three disulfide bonds to make the molecule a compact globular structure [45]. Royalisin is an amphipathic protein, and its C-terminal is rich in charged amino acids. This peptide inhibits the Gram-positive bacteria and fungi. It is particularly effective against the larvae of the bee pathogen Paenibacillus larvae, which causes American foulbrood [46].
2.2. Cecropins
Cecropins were first isolated from the hemolymph of the giant silk moth Hyalophora cecropia (cecropia moth), whence the term cecropin was derived [47]. These peptides are mainly structured by a large number of antibacterial and toxic peptides isolated from various lepidopteran and dipteran species, which constitute a major part of the cell-free immunity of insects. Cecropins are small proteins (around 35 amino acid residues) with activity against both Gram-positive and ram-negative bacteria. The principle insect cecropins (A, B and D) consist of 35–37 residues without cysteine [48,49]. Cecropins can lyse bacterial cellular membranes and can also inhibit proline uptake as well as cause leaky membranes [50,51]. Insect cecropins also have other names including bactericidin, lepidopteran, sarcotoxin, etc. [52]. These structurally related peptides are shown in Table 1.
Cecropin A is an AMP with a stabilized α-helical structure [55]. The precise antibacterial mechanism of cecropin A is unclear, but there is primary evidence showing that the cell membrane is the target [15]. Based on recent results, Yun and Lee [56] confirmed that an ion imbalance regulates cecropin A-induced apoptotic activity. Cecropin A can significantly reduce NADPH and glutathione levels to further induce oxidative stress by forming reactive oxygen species (ROS) [56,57]. Initially, cecropin peptides are arranged as antiparallel dimers with conserved residues of adjacent monomers in contact. The dimers may bind to the membrane with the NH2-terminal helices sunken into the head-group layer [57]. Cecropin A has promising activity against the fungus Beauveria bassiana in silkworm larvae [58].
Cecropin B is a naturally occurring linear cationic peptide consisting of 35 amino acids [59]. It is the member of the cecropin family with the highest antibacterial activity [59]. In a rat mode of septic shock, cecropin B significantly reduced the lethality of Escherichia coli load and plasma endotoxin levels [60]. Cecropin B attenuates the motility of the adult female nematode worm Brugia pahangi in adult females of Aedes aegypti and causes a significant decrease in the number of developing larvae [61]. Cecropin B also shows an antifungal capacity against Candida albicans [62]. Cecropin B, as well as other AMPs from the silkworm Bombyx mori including moricin (42 amino acids) [63,64], have a broad activity against porcine bacterial pathogens and is quite crucial in the porcine industry [65].
Cecropin C is present in very low quantities in the hemolymph of H. cecropia. Currently, the antibacterial activity of cecropin C is rarely reported. Compared with cecropin A, no C-terminal blocking group is present in cecropin C. Cecropin C is considered a precursor or degradation product of cecropin A [48].
Cecropin D is isolated from H. Cecropia and shows homology to cecropin A and cecropin B [48]. After bacterial infection, cecropin D appears in the hemolymph later than cecropin A or cecropin B [66]. A recombinant cecropin D has been successfully expressed in Pichia pastoris and showed antibacterial activity for both Gram-positive and Gram-negative bacteria [67]. The C-terminal lysine residue of cecropin D could increase antibacterial activity due to activated phosphorylation [68]. Cecropin D also inhibits porcine reproductive and respiratory syndrome virus (PRRSV) infection and replication in vitro [69].
Cecropin P1 is an antibacterial peptide from Ascaris suum, a parasitic nematode that resides in pig intestine [54]. Cecropin P1 could effectively inhibit the growth of enterotoxigenic E. coli with the minimal inhibitory concentration (MIC) of 1 mg/mL [70]. A tertiary structure study shows that cecropin P1 can form α-helical structures with the C-terminal region (Lys15–Gly29) in lipopolysaccharide (LPS) of the outer membrane of Gram-negative bacteria [71]. The concentration-dependent killing of E. coli by cecropin P1 can be driven through the extent of the immediate permeabilizing action of the peptide [72]. Cysteine-terminus modified cecropin P1 (CP1C) shows less antimicrobial activity, since the presence of polyethylene glycol (PEG) linker prevents CP1C from interacting with the bilayer [73]. Cecropin P1 also shows a significant inhibitory effect on human fungal pathogen C. albicans [74]. Cecropin P1 inhibits PRRSV by blocking attachment [75]. Cecropin P1 inhibits viral particle release and attenuates virus-induced apoptosis [75]. Currently, biosensors using cecropin P1 have been developed, and the peptide has been immobilized through different termini results in different functions and activities [76].
Lucilin, a 36-residue cecropin, is identified as a partial genetic sequence in Lucilia sericata maggots [77]. The fusion protein, GWLK-Lucilin-CPD-His8, shows a potential activity against multidrug resistant (MDR) bacteria E. coli [77]. Musca domestica cecropin is also a potential bactericidal agent against clinical isolates of E. coli [78].
2.3. Attacins
Attacins are glycine-rich proteins, belonging to the AMP group. Attacins were first discovered in Hyalophora cecropia [79]. They are effective against Gram-negative bacteria [80]. Attacins A–F are closely related antibacterial proteins, which are isolated from the hemolymph of immunized pupae of the cecropia moth (Hyalophora cecropia) [79]. They are a rather heterogeneous group of proteins, varying in size but rich in glycine residues (10–22%). Attacins A–F can be divided into two groups based on their amino acid composition: Attacins A–D constitute a basic group; and attacins E and F, which have acidic residues. Within each group, the forms are very similar (Figure 2).
Attacins act by blocking the synthesis of the major outer membrane proteins in dividing Gram-negative bacteria, thus disturbing the integrity of the cell wall and causing the bacteria to grow in long chains [81]. Attacins constitute an antibacterial active form of inducible immune protein P5. Attacins can effectively kill E. coli and other Gram-negative bacteria. In addition to cecropin and lysozyme, attacins are the third antibacterial protein in the humoral immune system of H. cecropia [82]. Some attacin and attacin-related proteins are isolated from Bombyx mori, Glossina morsitans (tse-tse fly), Heliothis virescens, Trichoplusia ni, Samia cynthia ricini (wild silkmoth) and Musca domestica (housefly) [83,84].
2.4. Lebocins
Lebocins are antibacterial peptides consisting of 32 amino acids, which were identified by Hara and Yamakawa [63] in the hemolymph of the silkworm Bombyx mori, immunized with E. coli [63]. Lebocin is a proline-rich and O-glycosylated peptide [85]. In total, 41% of the amino acid sequence of lebocin is identical with abaecin, a major 34 amino acid antibacterial peptide (YVPLPNVPQPGRRPFPTFPGQGPFNPKIKWPQGY-NH2) in the honeybee Apis mellifera [86]. The amino acid sequence in lebocin 1 is DLRFLYPRGKLPVPTPPPFNPKPIYIDMGNRY-NH2. The primary sequence of lebocins 1 and 2 differ only in their sugar moiety. Lebocin 3 has the same structure as lebocin 2, except that residue 16 is leucine instead of proline [87].
2.5. Drosocin
Drosocin is a peptide produced by Drosophila melanogaster [88]. This peptide contains 19 amino acids (GKPRPYSPRPTSHPRPIRV-NH2). The peptide is O-glycosylated, and this modification is required for maximum biological activity [89]. Glycosylation is an important post-translational modification for some proline-riched AMPs class [90]. In addition to the Gram-positive bacterium M. luteus, Drosocin is primarily resistant to Gram-negative bacteria. Deletion of the first five N-terminal residues completely abolishes the activity of drosocin [88,91]. Glycosylated drosocin is active against E. coli and fungi [92].
Apidaecin IB and drosocin show significant sequence homology and interaction mechanism but lack any pore-forming activity [93]. Apidaecins are the major components of the honeybee humoral defence against microbial invasion [93]. N-terminal mutation of apidaecins not only reinforces the interaction with unidentified intracellular target(s), but also promotes the cell-penetration efficiency [94]. Structure N-terminal Ile-Orn- and Trp-Orn-motif repeats increases the antimicrobial activity against Pseudomonas aeruginosa [95].
2.6. Diptericins
Diptericins constitute a family of related glycine-rich antibacterial peptides (about 8 kD) from Dipteran hemolymph proteins of about 82 amino acids [96]. Diptericins A–C have been isolated from immunized larvae of the dipteran Phormia terranovae [97]. Diptericin is also expressed in D. melanogaster [98], Sarcophaga peregrina (flesh fly) [99], and Mayetiola destructor (Hessian fly). The predominant member of this family of peptides is diptericin A (DDMTMKPTPPPQYPLNLQGGGGGQSGDGFGFAVQGHQKVWTSDNGRHEIGLNGGYGQHLGGPYGNSEPSWKVGSTYTYRFPNF-NH2). This peptide is active only against a limited number of Gram-negative bacteria (E. coli K12, Erwinia hericola T, and Erwinia carotovora 113). Diptericin A mainly acts on the cytoplasmic membrane of growing bacteria [100].
Prolixicin is a novel AMP from the family of diptericins. This peptide has been isolated from the hemipteran insect Rhodnius prolixus. Prolixicin is a peptide with 21 amino acids [101]. It has also two putative phosphorylation sites, but no glycosylation sites have been identified. Sequence analysis reveals that one region of prolixicin is related to the diptericin/attacin family of AMPs. This peptide can be produced by midgut tissues after the bacterial infection of the hemolymph [101].
2.7. Metchnikowin
In 1995, Russian scientists identified a novel 26-residue proline-rich immune-inducible linear peptide (HRHEGPIFNTRPSPFNPNEPRPGPIY-NH2) from D. melanogaster, which was remarkable in its unusual antimicrobial activity against Gram-positive bacteria and fungi. However, this peptide has no effect against Gram-negative bacteria [102]. This peptide was named Metchnikowin in honour of E. Metchnikow who initiated this field of research. There are two isoforms of Metchnikowin in the Oregon Drosophila strain, which differ by one residue (His compared to Arg) [103]. The target of Metchnikowin has been studied. Metchnikowin interacts with the fungal enzyme β(1,3)-glucanosyltransferase Gel1 (FgBGT), an important enzyme that is involved in fungal cell wall synthesis [104]. Metchnikowin also targets the iron-sulfur subunit (SdhB) of succinate-coenzyme Q reductase (SQR). In the study of Moghaddam et al. [104], Metchnikowin inhibited the SDH activity of Fusarium graminearum mitochondrial SQR by up to 52%.
2.8. Ponericins
Ponericins are peptides isolated from the venom of the predatory ant Pachycondyla goeldii. Members of the subfamily Ponerinae were isolated and their amino acid sequences were characterized by Orivel and coworkers [105]. Ponericins can be classified into three families based on their primary structure similarities: Ponericins G, W, and L (Table 2). Ponericins have high sequence similarities for the known peptides: Ponericin G has similarity with cecropin-like peptides; ponericin W is similar with gaegurins and melittin; and ponericin L is similar with dermaseptins. Ponericins also show hemolytic activities, as well as insecticidal activities against cricket larvae. Ponericins have α-helical structures in cell membranes [106].
2.9. Jelleines
Jelleines are a family of peptides isolated from Apis mellifera royal jelly [107]. They are composed of 8–9 amino acids and bear a +2 charge at the C-terminus [107]. Four AMPs were purified from royal jelly of honeybees: Jelleine-I (PFKLSLHL-NH2), jelleine-II (TPFKLSLHL-NH2), jelleine-III (EPFKLSLHL-NH2) and jelleine-IV (TPFKLSLH-NH2) [108]. Jelleines-I–III presented antimicrobial activities against yeast, fungi, Gram-positive and Gram-negative bacteria [108,109]. These peptides have no similarities with other AMPs from honeybees. Molecules of jelleines are still in the characterization phase [110].
2.10. Apisimin
Apisimin is a peptide consisting of 54 amino acids with a primary sequence of KTSISVKGESNVDVVSQINSLVSSIVSGANVSAVLLAQTLVNILQILIDANVFA-NH2, and is found in honeybee royal jelly [111], which stimulates the proliferation of human monocytes [112]. Apisimin is rich in valine and serine, and contains only one aromatic amino acid, phenylalanine [108]. Apisimin is a small peptide in royal jelly. High levels of small mRNA expression of apisimin are observed in the heads of nurse and foraging honeybees. Therefore, they may play a physiological role in honeybee colonies [111]. The study of Gannabathula et al. [113] provides evidence that apisimin and arabinogalactan proteins are present in honey and contribute to their immune active properties.
2.11. Pyrrhocoricin
Pyrrhocoricin, a proline-rich inducible AMP, was isolated from the hemolymph of the sap-sucking bug Pyrrhocoris apterus by Cociancich et al. [114]. This 20 amino acid peptide (VDKGSYLPRPTPPRPIYNRN-NH2) interacts with the heat shock protein DnaK, which is correlated with the antimicrobial activity [115]. Pyrrhocoricin can bind to and promote the ATPase activity of the molecular chaperone DnaK [116,117]. Boxell et al. [118] showed that pyrrhocoricin could act as a delivery vehicle in transducing peptides across the cell membrane of the parasite Cryptosporidium parvum. The successful transduction facilitates target validation. It will also help to deliver peptide-based drugs to this important human pathogen. Cyclization of pyrrhocoricin structural elements is important for the antimicrobial activity of the native peptide [119].
2.12. Persulcatusin
Persulcatusin has been identified in the midgut of Ixodes persulcatus. Its amino acid sequence is GFGCPFNQGACHRHCRSIGRRGGYCAGLFKQTCTCYSR-NH2 [120]. The complete structure of persulcatusin has not been identified yet. Its similarity with other known tick AMPs is from 71% to 88% [121,122]. The structural integrity of persulcatusin is maintained by three S-S bonds, which are energetically important for the stability and the formation of the structure of α-helix and β-sheet [123,124]. This peptide can inhibit the growth of methicillin-sensitive S. aureus (MSSA) and methicillin-resistant S. aureus (MRSA) with the MIC of 0.156–1.25 μg/mL and 0.625–2.5 μg/mL, respectively [31]. Very recently, persulcatusin exhibits strong antibacterial activity against MDR S. aureus strains, including VRSA [123]. The antimicrobial activity of persulcatusin against MRSA was stronger than that of other AMPs [124].
2.13. Melittin
Melittin is a peptide toxin found in bee venom and is effective against bacteria [125,126]. This peptide has a linear structure with 26 amino acid residues (GIGAVLKVLTTGLPALISWIKRKRQQ-NH2) [125,127]. Melittin has a strong antibacterial effect against a variety of bacteria including Borrelia burgdorferi [128], Listeria monocytogenes [129], S. aureus, and P. aeruginosa [130,131]. Melittin has antibacterial activity against Xanthomonas oryzae pv. oryzae, a destructive bacterial disease of rice, indicating that this peptide may have potential applications in plant protection [132]. RV-23, a melittin-related peptide, shows strong antibacterial activity against E. coli and S. aureus [133]. A new synthetic peptide, MelP5, is a gain-of-function variant of melittin [134]. Moreover, this peptide facilitates the passage of macromolecules across bilayers.
Melittin binds to membrane surfaces with a negative charge to disturb the integrity of phospholipid bilayers by forming pores, which subsequently induces the leakage of atomic ions and molecules and ultimately leads to cell lysis [135]. Proline residue is important in the antimicrobial activity of melittin [136]. Analogues lacking the poline residue and dimers decrease the cytotoxicity and minimize the inhibitor concentrations. However, there are ongoing debates regarding the molecular mechanism of melittin [137,138]. For decades, the equilibrium transmembrane pore mode has been considered the major mechanism of the antibacterial activity of melittin. However, emerging evidence shows that the transmembrane pore is not required in this context [139]. Notably, the mechanisms can be markedly influenced by experimental design in different studies [137]. Lee and Lee [140] further reported that melittin triggers apoptosis in C. albicans through the ROS-mediated mitochondria and caspase pathway.
Very recently, Akbari et al. [141] showed that there are highly synergistic effects of melittin with conventional antibiotics against MDR isolates of Acinetobacter baumannii and P. aeruginosa. In their study, the geometric means of MIC for melittin and doripenem after combination were reduced to 61.5- and 51.5-fold, respectively, against A. baumannii isolates. This working group [142] further showed that the new melittin-derived peptides MDP1 and MDP2 exhibited efficient antibacterial activity against MDR S. aureus, E. coli, and P. aeruginosa. Melittin also exhibits very effective antibacterial activity against MRSA strains [143]. MRSA-infected mice treated with melittin were successfully rescued from bacteraemia. The clinical application of melittin still needs a lot of work in the future, since most current work is in the preclinical phase [144].
3. Concluding Remarks
Insect AMPs are the main immune effector molecules. Therefore, there are large numbers of AMP resources in the huge insect world. In recent years, fruit fly and mosquito have been used as model organisms, and great progress has been made in the study of the natural immunity of insects. There is growing evidence that the natural immune system of insects is much more complex than one might expect. In the meantime, due to the conservation of biological evolution, certain molecules and signaling pathways in the natural immune system of insects have certain similarities with vertebrates (including humans). Studies of the natural immune system of insects helps to further understand the complexity of the human immune system. Currently, more bacteria have developed multidrug resistance due to the abuse of antibiotics, and some super antibiotic resistant bacteria have emerged, which pose a great threat to human health [145]. Finding and developing new antimicrobial drugs has become an urgent problem in the medical field.
AMPs not only have a broad-spectrum killing effect on bacteria and fungi, but also have a killing effect on viruses, protozoa and cancer cells. Compared with traditional antibiotics, their mechanism of action is unique, and it is not easy for AMPs to cause microbial resistance. Most of them do not damage or destroy normal cells of higher animals. For example, very recently, clavaspirin peptide, a peptide from tunicate Styela clava, exhibited the capacity to kill drug-resistant pathogens (S. aureus) without detectable resistance [146]. The above advantages and rich resource content of insect AMPs make them excellent templates for the development of new antimicrobial drugs. Indeed, some synergistic effects of AMPs with conventional antibiotics are observed against bacteria [141]. However, quantitative methods are rarely used to test this synergistic profile. Wu et al. [147] recently tested the synergistic effect of AMP DP7 and antibiotics on MDRs (S. aureus, E. coli) using quantitative polymerase chain reaction. Other researchers [148] also tested the synergistic antimicrobial activity of frog peptides via oriented circular dichroism and quantitative solid-state F-19-NMR analysis. In the future, more quantitative studies on the synergistic effects of AMPs with antibiotics should be encouraged. Currently, the market for peptide drugs is increasing steadily, and some products such as Bacitracin, polymyxin and Fuzeon are already on the market [5]. However, the clinical use of AMPs is still limited by some shortcomings, such as low bioavailability, potential hemolysis, instability to proteases, and unknown toxicity [10]. Investigations of natural peptides and nano-delivery systems from natural polymers are a new research focus area for the future [149]. However, there is one issue that needs to be considered: Despite intensive studies, we still do not fully understand the structure-activity relationship (SAR) and mechanisms underlying AMP activity. Therefore, more SAR studies of AMPs are required. Moreover, further investigations into the cellular and molecular mechanisms of AMP effects are warranted. Finally, a library of insect AMPs should be established in order to optimize them further and improve their antimicrobial activity and toxic properties.
Author Contributions
Q.W. and J.P. wrote the manuscript; K.K. reviewed and revised the manuscript. All of the authors critically reviewed the literature and contributed to drafting the manuscript.
Funding
This work was funded by National Natural Science Foundation of China (grant no. 31602114) and the Excellence project UHK and Long-term development plan (University Hospital, Hradec Kralove, Czech Republic).
Acknowledgments
Q.W. wishes to thank X.J. Zhang for her nice care during the hard working on this manuscript with K.K.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bulet, P.; Hetru, C.; Dimarcq, J.L.; Hoffmann, D. Abtimicrobial peptides in insects; structure and function. Dev. Comp. Immun. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uccelletti, D.; Zanni, E.; Marcellini, L.; Palleschi, C.; Barra, D.; Mangoni, M.L. Anti-Pseudomonas activity of frog skin antimicrobial peptides in a Caenorhabditis elegans infection model: A plausible mode of action in vitro and in vivo. Antimicrob. Agents Chemother. 2010, 54, 3853–3860. [Google Scholar] [CrossRef] [PubMed]
- Tonk, M.; Vilcinskas, A.; Rahnamaeian, M. Insect antimicrobial peptides: Potential tools for the prevention of skin cancer. Appl. Microbiol. Biotechnol. 2016, 100, 7397–7405. [Google Scholar] [CrossRef] [PubMed]
- Ongey, E.L.; Pflugmacher, S.; Neubauer, P. Bioinspired designs, molecular premise and tools for evaluating the ecological importance of antimicrobial peptides. Pharmaceuticals 2018, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Patocka, J.; Nepovimova, E.; Klimova, B.; Wu, Q.; Kuca, K. Antimicrobial peptides: Amphibian host defense peptides. Curr. Med. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Weinberg, A. Human antimicrobial peptides and cancer. Semin. Cell Dev. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Chernysh, S.; Gordya, N.; Suborova, T. Insect antimicrobial peptide complexes prevent resistance development in bacteria. PLoS ONE 2015, 10, e0130788. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; He, P.; Xiao, C.; Chen, X. From Antimicrobial Peptides to Antimicrobial Poly(α-amino acid)s. Adv. Healthc. Mater. 2018, 1, 1800354. [Google Scholar] [CrossRef] [PubMed]
- Hollmann, A.; Martinez, M.; Maturana, P.; Semorile, L.C.; Maffia, P.C. Antimicrobial peptides: Interaction with model and biological membranes and synergism with chemical antibiotics. Front. Chem. 2018, 6, 204. [Google Scholar] [CrossRef] [PubMed]
- Jozefiak, A.; Engberg, R.M. Insect proteins as a potential source of antimicrobial peptides in livestock production. A review. J. Anim. Feed Sci. 2017, 26, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Bechinger, B.; Gorr, S.U. Antimicrobial peptides: Mechanisms of action and resistance. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Oñate-Garzón, J.; Manrique-Moreno, M.; Trier, S.; Leidy, C.; Torres, R.; Patiño, E. Antimicrobial activity and interactions of cationic peptides derived from Galleria mellonella cecropin D-like peptide with model membranes. J. Antibiot. 2017, 70, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, L.; Axelsen, P.H. Membrane-induced folding of cecropin A. Biophys. J. 2000, 79, 1465–1477. [Google Scholar] [CrossRef]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.P.S.; Hancock, R.E.W. The relationship between peptide structure and antibacterial activity. Peptides 2003, 24, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Nan, Y.H.; Shin, S.Y. Effect of disulphide bond position on salt resistance and LPS-neutralizing activity of α-helical homo-dimeric model antimicrobial peptides. BMB Rep. 2011, 44, 747–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaakobi, K.; Liebes-Peer, Y.; Kushmaro, A.; Rapaport, H. Designed amphiphilic β-sheet peptides as templates for paraoxon adsorption and detection. Langmuir 2013, 29, 6840–6848. [Google Scholar] [CrossRef] [PubMed]
- Freudenthal, O.; Quilès, F.; Francius, G. Discrepancies between cyclic and linear antimicrobial peptide actions on the spectrochemical and nanomechanical fingerprints of a young biofilm. ACS Omega 2017, 2, 5861–5872. [Google Scholar] [CrossRef] [PubMed]
- Kindrachuk, J.; Napper, S. Structure-activity relationships of multifunctional host defence peptides. Mini-Rev. Med. Chem. 2010, 10, 596–614. [Google Scholar] [CrossRef] [PubMed]
- Suarez, M.; Haenni, M.; Canarelli, S.; Fisch, F.; Chodanowski, P.; Servis, C.; Michielin, O.; Freitag, R.; Moreillon, P.; Mermod, N. Structure-function characterization and optimization of a plant-derived antibacterial peptide. Antimicrob. Agents Chemother. 2005, 49, 3847–3857. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.; Mannion, M.; Pike, D.; Lewis, K.; Flynn, A.; Brannan, A.M.; Browne, M.J.; Jackman, D.; Madera, L.; Power Coombs, M.R.; Hoskin, D.W.; Rise, M.L.; Booth, V. Structure-function relationships in histidine-rich antimicrobial peptides from Atlantic cod. Biochim. Biophys. Acta 2015, 1848, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Locock, K.E.; Michl, T.D.; Valentin, J.D.; Vasilev, K.; Hayball, J.D.; Qu, Y.; Traven, A.; Griesser, H.J.; Meagher, L.; Haeussler, M. Guanylated polymethacrylates: A class of potent antimicrobial polymers with low hemolytic activity. Biomacromolecules 2013, 14, 4021–4031. [Google Scholar] [CrossRef] [PubMed]
- Leptihn, S.; Har, J.Y.; Wohland, T.; Ding, J.L. Correlation of charge, hydrophobicity, and structure with antimicrobial activity of S1 and MIRIAM peptides. Biochemistry 2010, 49, 9161–9170. [Google Scholar] [CrossRef] [PubMed]
- Naumenkova, T.V.; Antonov, M.I.u.; Nikolaev, I.N.; Shaĭtan, K.V. Effect of Pro11-Ala11 amino acid substitution on structural and functional properties of antimicrobial peptide buforin 2. Biofizika 2012, 57, 988–999. [Google Scholar] [PubMed]
- Derache, C.; Meudal, H.; Aucagne, V.; Mark, K.J.; Cadène, M.; Delmas, A.F.; Lalmanach, A.C.; Landon, C. Initial insights into structure-activity relationships of avian defensins. J. Biol. Chem. 2012, 287, 7746–7755. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.J.; Sohn, H.I.; Nan, Y.H.; Murugan, R.N.; Cheong, C.; Ryu, E.K.; Kim, E.H.; Kang, S.W.; Kim, E.J.; Shin, S.Y.; et al. Functional and structural characterization of Drosocin and its derivatives linked O-GalNAc at Thr 11 residue. Bull. Korean Chem. Soc. 2011, 32, 3327–3332. [Google Scholar] [CrossRef]
- Walkenhorst, W.F.; Klein, J.W.; Vo, P.; Wimley, W.C. pH Dependence of microbe sterilization by cationic antimicrobial peptides. Antimicrob. Agents Chemother. 2013, 57, 3312–3320. [Google Scholar] [CrossRef] [PubMed]
- Makarova, O.; Johnston, P.; Rodriguez-Rojas, A.; El Shazely, B.; Morales, J.M.; Rolff, J. Genomics of experimental adaptation of Staphylococcus aureus to a natural combination of insect antimicrobial peptides. Sci. Rep. 2018, 8, 15359. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, E.; Podsiadlowski, L.; Muhammed, M.; Vilcinskas, A. Diversity, evolution and medical applications of insect antimicrobial peptides. Philos. Trans. R. Soc. Lond. B 2016, 371, 20150290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Rozgonyi, F.; Szabo, D.; Kocsis, B.; Ostorházi, E.; Abbadessa, G.; Cassone, M.; Wade, J.D.; Otvos, L., Jr. The antibacterial effect of a proline-rich antibacterial peptide A3-APO. Curr. Med. Chem. 2009, 16, 3996–4002. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Lehrer, R.I. Defensins. Pharmacol. Ther. 1995, 66, 191–205. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.; Cammue, B.P.; Thevissen, K. Plant defensins. Planta 2002, 216, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Gao, B. Evolutionary origin of β-defensins. Dev. Comp. Immunol. 2013, 39, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Cederlund, A.; Gudmundsson, G.H.; Agerberth, B. Antimicrobial peptides important in innate immunity. FEBS J. 2011, 278, 3942–3951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, D.K.; Rajagopalan, G.; Li, X.; Chen, Y.; Lua, L.H.; Leong, S.S. A new bioproduction route for a novel antimicrobial peptide. Biotechnol. Bioeng. 2011, 108, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.A.; Hetru, C. Insect defensins: Inducible antibacterial peptides. Immunol. Today 1992, 13, 411–415. [Google Scholar] [CrossRef]
- Hetru, C.; Troxler, L.; Hoffmann, J.A. Drosophiola melanogaster antimicrobial defense. J. Infect. Dis. 2003, 187, S327–S343. [Google Scholar] [CrossRef] [PubMed]
- Gomes, P.S.; Fernandes, M.H. Defensins in the oral cavity: Distribution and biological role. J. Oral Pathol. Med. 2010, 39, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G. Antibacterial peptides: Key components needed in immunity. Cell 1991, 65, 205–207. [Google Scholar] [CrossRef]
- Zhao, W.Y.; Dong, B.R.; Zhou, Y. In vitro antimicrobial activity of defensins from rabbit neutrophils against Pseudomonas aeruginosa and its multiple-drug-resistance strains. Sichuan Da Xue Xue Bao Yi Xue Ban. 2005, 36, 83–85. [Google Scholar] [PubMed]
- Fujiwara, S.; Imai, J.; Fujiwara, M.; Yaeshima, T.; Kawashima, T.; Kobayashi, K. A potent antibacterial protein in royal jelly. J. Biol. Chem. 1990, 265, 11333–11337. [Google Scholar] [PubMed]
- Bılikova, K.; Gusu, W.; Simuth, J. Isolation of a peptide fraction from honeybee royal jelly as a potential antifoulbrood factor. Apidologie 2001, 32, 275–283. [Google Scholar] [CrossRef]
- Steiner, H.; Hultmark, D.; Engstrom, A.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D.; Engström, A.; Bennich, H.; Kapur, R.; Boman, H.G. Insect immunity. Isolation and structure of cecropin D and four minor antibacterial components from cecropia pupae. Eur. J. Biochem. 1982, 127, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Van Hofsten, P.; Faye, I.; Kockum, K.; Lee, J.Y.; Xanthopoulos, K.G.; Boman, I.A. Molecular cloning, cDNA sequencing, and chemical synthesis of cecropin B from Hyalophora cecropia. Proc. Nat. Acad. Sci. USA 1985, 82, 2240–2243. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.J.; Beazley, W.D.; Bibby, M.C.; Devine, D.A. Antimicrobial activity of cecropins. J. Antimicrobiol. Chemother. 1996, 37, 1077–1089. [Google Scholar] [CrossRef] [Green Version]
- Bechinger, B.; Lohner, K. Detergent-like actions of linear amphipathic cationic antimicrobial peptides. Biochim. Biophys. Acta 2006, 1758, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Xu, X.; Freed, S.; Gao, Y.; Yu, J.; Wang, S.; Ju, W.; Zhang, Y.; Jin, F. Cecropins from Plutella xylostella and Their Interaction with Metarhizium anisopliae. PLoS ONE 2015, 10, e0142451. [Google Scholar] [CrossRef] [PubMed]
- Srisailam, S.; Kumar, T.K.S.; Arunkumar, A.I.; Leung, K.W.; Yu, C.; Chen, H.M. Crumpled structure of the custom hydrophobic lytic peptide cecropin B3. Eur. J. Biochem. 2001, 268, 4278–4284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, A.; Ueno, S.; Zhang, H.; Lee, J.M.; Kato, Y. Cecropin P1 and novel nematode cecropins: A bacteria-inducible antimicrobial peptide family in the nematode Ascaris suum. Biochem. J. 2005, 390, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Björstad, A.; Dahlgren, C.; Bylund, J. A bacterial cecropin-A peptide with a stabilized α-helical structure possess an increased killing capacity but no proinflammatory activity. Inflammation 2004, 28, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Lee, D.G. Cecropin A-induced apoptosis is regulated by ion balance and glutathione antioxidant system in Candida albicans. IUBMB Life 2016, 68, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Durell, S.R.; Raghunathan, G.; Guy, H.R. Modeling the ion channel structure of cecropin. Biophys. J. 1992, 63, 1623–1631. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Geng, T.; Hou, C.; Huang, Y.; Qin, G.; Guo, X. Bombyx mori cecropin A has a high antifungal activity to entomopathogenic fungus Beauveria bassiana. Gene 2016, 583, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Srisailam, S.; Arunkumar, A.I.; Wang, W.; Yu, C.; Chen, H.M. Conformational study of a custom antibacterial peptide cecropin B1: Implications of the lytic activity. Biochim. Biophys. Acta 2000, 1479, 275–285. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Ghiselli, R.; Viticchi, C.; Mocchegiani, F.; Riva, A.; Saba, V.; Scalise, G. Effect of mono-dose intraperitoneal cecropins in experimental septic shock. Crit. Care Med. 2001, 29, 1666–1669. [Google Scholar] [CrossRef] [PubMed]
- Chalk, R.; Townson, H.; Ham, P.J. Brugia pahangi: The effects of cecropins on microfilariae in vitro and in Aedes aegypti. Exp. Parasitol. 1995, 80, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Andrä, J.; Berninghausen, O.; Leippe, M. Cecropins, antibacterial peptides from insects and mammals, are potently fungicidal against Candida albicans. Med. Microbiol. Immunol. 2001, 189, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Hara, S.; Yamakawa, M. A novel antibacterial peptide family isolated from the silkworm, Bombyx mori. Biochem. J. 1995, 310, 651–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, S.; Yamakawa, M. Moricin, a novel type of antibacterial peptide isolated from the silkworm, Bombyx mori. J. Biol. Chem. 1995, 270, 29923–29927. [Google Scholar] [PubMed]
- Hu, H.; Wang, C.; Guo, X.; Li, W.; Wang, Y.; He, Q. Broad activity against porcine bacterial pathogens displayed by two insect antimicrobial peptides moricin and cecropin B. Mol. Cells 2013, 35, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudmundsson, G.H.; Lidholm, D.A.; Asling, B.; Gan, R.; Boman, H.G. The cecropin locus. Cloning and expression of a gene cluster encoding three antibacterial peptides in Hyalophora cecropia. J. Biol. Chem. 1991, 266, 11510–11517. [Google Scholar] [PubMed]
- Guo, C.; Huang, Y.; Zheng, H.; Tang, L.; He, J.; Xiao, L.; Liu, D.; Jiang, H. Secretion and activity of antimicrobial peptide cecropin D expressed in Pichia pastoris. Exp. Ther. Med. 2012, 4, 1063–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.I.; An, H.S.; Chang, B.S.; Yoe, S.M. Expression, cDNA cloning, and characterization of the antibacterial peptide cecropin D from Agrius convolvuli. Anim. Cells Syst. 2013, 17, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Guo, C.; Huang, Y.; Zhang, X.; Chen, Y. Inhibition of porcine reproductive and respiratory syndrome virus by Cecropin D in vitro. Infect. Genet. Evol. 2015, 34, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.P.; Wang, W.J.; Liu, Z.Q.; Liu, G.; Souffrant, W.B.; Yin, Y.L. Effect of lactoferricin B and cecropin P1 against enterotoxigenic Escherichia coli in vitro. J. Food Agric. Environ. 2011, 9, 271–274. [Google Scholar]
- Baek, M.H.; Kamiya, M.; Kushibiki, T.; Nakazumi, T.; Tomisawa, S.; Abe, C.; Kumaki, Y.; Kikukawa, T.; Demura, M.; Kawano, K.; et al. Lipopolysaccharide-bound structure of the antimicrobial peptide cecropin P1 determined by nuclear magnetic resonance spectroscopy. J. Pept. Sci. 2016, 22, 214–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arcidiacono, S.; Soares, J.W.; Meehan, A.M.; Marek, P.; Kirby, R. Membrane permeability and antimicrobial kinetics of cecropin P1 against Escherichia coli. J. Pept. Sci. 2009, 15, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wei, P.H.; Zhu, X.; Wirth, M.J.; Bhunia, A.; Narsimhan, G. Effect of immobilization on the antimicrobial activity of a cysteine-terminated antimicrobial Peptide Cecropin P1 tethered to silica nanoparticle against E. coli O157:H7 E.D.L. Colloids Surf. B Biointerfaces 2017, 156, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Zakharchenko, N.S.; Belous, A.S.; Biryukova, Y.K.; Medvedeva, OA.; Belyakova, A.V.; Masgutova, G.A.; Trubnikova, E.V.; Buryanov, Y.I.; Lebedeva, A.A. Immunomodulating and Revascularizing Activity of Kalanchoe pinnata Synergize with Fungicide Activity of Biogenic Peptide Cecropin P1. J. Immunol. Res. 2017, 2017, 3940743. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Huang, Y.; Cong, P.; Liu, X.; Chen, Y.; He, Z. Cecropin P1 inhibits porcine reproductive and respiratory syndrome virus by blocking attachment. BMC Microbiol. 2014, 14, 273. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Han, X.; He, N.; Chen, Z.; Brooks, C.L., 3rd. Molecular structures of C- and N-terminus cysteine modified cecropin P1 chemically immobilized onto maleimide-terminated self-assembled monolayers investigated by molecular dynamics simulation. J. Phys. Chem. B 2014, 118, 5670–5680. [Google Scholar] [CrossRef] [PubMed]
- Téllez, G.A.; Castaño-Osorio, J.C. Expression and purification of an active cecropin-like recombinant protein against multidrug resistance Escherichia coli. Protein Expr. Purif. 2014, 100, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Shen, J.; Jin, X.; Ma, Y.; Huang, Y.; Mei, H.; Chu, F.; Zhu, J. Bactericidal activity of Musca domestica cecropin (Mdc) on multidrug-resistant clinical isolate of Escherichia coli. Appl. Microbiol. Biotechnol. 2012, 95, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D.; Engström, A.; Andersson, K.; Steiner, H.; Bennich, H.; Boman, H.G. Insect immunity. Attacins, a family of antibacterial proteins from Hyalophora cecropia. EMBO J. 1983, 2, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, A.; Engström, P.; Palva, E.T.; Bennich, H. Attacin, an antibacterial protein from Hyalophora cecropia, inhibits synthesis of outer membrane proteins in Escherichia coli by interfering with omp gene transcription. Infect. Immun. 1991, 59, 3040–3045. [Google Scholar] [PubMed]
- Carlsson, A.; Nyström, T.; de Cock, H.; Bennich, H. Attacin—An insect immune protein—Binds LPS and triggers the specific inhibition of bacterial outer-membrane protein synthesis. Microbiology 1998, 144, 2179–2188. [Google Scholar] [CrossRef] [PubMed]
- Kockum, K.; Faye, I.; Hofsten, P.V.; Lee, J.Y.; Xanthopoulos, K.G.; Boman, H.G. Insect immunity. Isolation and sequence of two cDNA clones corresponding to acidic and basic attacins from Hyalophora cecropia. EMBO J. 1984, 3, 2071–2075. [Google Scholar] [CrossRef] [PubMed]
- Dushay, M.S.; Roethele, J.B.; Chaverri, J.M.; Dulek, D.E.; Syed, S.K.; Kitami, T.; Eldon, E.D. Two attacin antibacterial genes of Drosophila melanogaster. Gene 2000, 246, 49–57. [Google Scholar] [CrossRef]
- Geng, H.; An, C.J.; Hao, Y.J.; Li, D.S.; Du, R.Q. Molecular cloning and expression of Attacin from housefly (Musca domestica). Yi Chuan Xue Bao 2004, 31, 1344–1350. [Google Scholar] [PubMed]
- Liu, G.; Kang, D.; Steiner, H. Trichoplusia ni lebocin, an inducible immune gene with a downstream insertion element. Biochem. Biophys. Res. Commun. 2000, 269, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Casteels, P.; Ampe, C.; Riviere, L.; Van Damme, J.; Elicone, C.; Fleming, M.; Jacobs, F.; Tempst, P. Isolation and characterization of abaecin, a major antibacterial response peptide in the honeybee (Apis mellifera). Eur. J. Biochem. 1990, 187, 381–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slocinska, M.; Marciniak, P.; Rosinski, G. Insects antiviral and anticancer peptides: New leads for the future? Protein Pept. Lett. 2008, 15, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Dimarcq, J.L.; Hetru, C.; Lagueux, M.; Charlet, M.; Hegy, G.; Van Dorsselaer, A.; Hoffmann, J.A. A novel inducible antibacterial peptide of Drosophila carries an O-glycosylated substitution. J. Biol. Chem. 1993, 268, 14893–14897. [Google Scholar] [PubMed]
- McManus, A.M.; Otvos, L., Jr.; Hoffmann, R.; Craik, D.J. Conformational studies by NMR of the antimicrobial peptide, drosocin, and its non-glycosylated derivative: Effects of glycosylation on solution conformation. Biochemistry 1999, 38, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Lele, D.S.; Talat, S.; Kumari, S.; Srivastava, N.; Kaur, K.J. Understanding the importance of glycosylated threonine and stereospecific action of Drosocin, a Proline rich antimicrobial peptide. Eur. J. Med. Chem. 2015, 92, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Urge, L.; Ohresser, S.; Hetru, C.; Otvos, L., Jr. Enlarged scale chemical synthesis and range of activity of drosocin, an O-glycosylated antibacterial peptide of Drosophila. Eur. J. Biochem. 1996, 238, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Imler, J.L.; Bulet, P. Antimicrobial peptides in Drosophila: Structures, activities and gene regulation. Chem. Immunol. Allergy 2005, 86, 1–21. [Google Scholar] [PubMed]
- Gobbo, M.; Biondi, L.; Filira, F.; Gennaro, R.; Benincasa, M.; Scolaro, B.; Rocchi, R. Antimicrobial peptides: Synthesis and antibacterial activity of linear and cyclic drosocin and apidaecin 1b analogues. J. Med. Chem. 2002, 45, 4494–4504. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Orikasa, Y.; Ichinohe, K.; Hashimoto, S.; Ooi, T.; Taguchi, S. Flow cytometric analysis of the contributing factors for antimicrobial activity enhancement of cell-penetrating type peptides: Case study on engineered apidaecins. Biochem. Biophys. Res. Commun. 2010, 395, 7–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bluhm, M.E.; Schneider, V.A.; Schäfer, I.; Piantavigna, S.; Goldbach, T.; Knappe, D.; Seibel, P.; Martin, L.L.; Veldhuizen, E.J.; Hoffmann, R. N-Terminal Ile-Orn- and Trp-Orn-Motif Repeats Enhance Membrane Interaction and Increase the Antimicrobial Activity of Apidaecins against Pseudomonas aeruginosa. Front. Cell Dev. Biol. 2016, 4, 39. [Google Scholar] [CrossRef] [PubMed]
- Cudic, M.; Bulet, P.; Hoffmann, R.; Craik, D.J.; Otvos, L., Jr. Chemical synthesis, antibacterial activity and conformation of diptericin, an 82-mer peptide originally isolated from insects. Eur. J. Biochem. 1999, 266, 549–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimarcq, J.L.; Keppi, E.; Dunbar, B.; Lambert, J.; Reichhart, J.M.; Hoffmann, D.; Rankine, S.M.; Fothergill, J.E.; Hoffmann, J.A. Insect immunity. Purification and characterization of a family of novel inducible antibacterial proteins from immunized larvae of the dipteran Phormia terranovae and complete amino-acid sequence of the predominant member, diptericin A. Eur. J. Biochem. 1988, 171, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Reichhart, J.M.; Meister, M.; Dimarcq, J.L.; Zachary, D.; Hoffmann, D.; Ruiz, C.; Richards, G.; Hoffmann, J.A. Insect immunity: Developmental and inducible activity of the Drosophila diptericin promoter. EMBO J. 1992, 11, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Kubo, T.; Natori, S. Purification and characterization of a diptericin homologue from Sarcophaga peregrina (flesh fly). Biochem. J. 1992, 287, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Keppi, E.; Pugsley, A.P.; Lambert, J.; Wicker, C.; Dimarcq, J.L.; Hoffmann, J.A.; Hoffmann, D. Mode of action of diptericin A, a bactericidal peptide induced in the hemolymph of Phormia terranovae larvae. Insect Biochem. Physiol. 1989, 10, 229–239. [Google Scholar] [CrossRef]
- Ursic-Bedoya, R.; Buchhop, J.; Joy, J.B.; Durvasula, R.; Lowenberger, C. Prolixicin: A novel antimicrobial peptide isolated from Rhodnius prolixus with differential activity against bacteria and Trypanosoma cruzi. Insect Mol. Biol. 2011, 20, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Levashina, E.A.; Ohresser, S.; Bulet, P.; Reichhart, J.M.; Hetru, C.; Hoffmann, J.A. Metchnikowin, a novel immune-inducible proline-rich peptide from Drosophila with antibacterial and antifungal properties. Eur. J. Biochem. 1995, 233, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Levashina, E.A.; Ohresser, S.; Lemaitre, B.; Imler, J.L. Two distinct pathways can control expression of the gene encoding the Drosophila antimicrobial peptide metchnikowin. J. Mol. Biol. 1998, 278, 515–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghaddam, M.R.B.; Gross, T.; Becker, A.; Vilcinskas, A.; Rahnamaeian, M. The selective antifungal activity of Drosophila melanogaster metchnikowin reflects the speciesdependent inhibition of succinate-coenzyme Q reductase. Sci. Rep. 2017, 7, 8192. [Google Scholar] [CrossRef] [PubMed]
- Orivel, J.; Redeker, V.; Le Caer, J.P.; Krier, F.; Revol-Junelles, A.M.; Longeon, A.; Chaffotte, A.; Dejean, A.; Rossier, J. Ponericins, new antibacterial and insecticidal peptides from the venom of the ant Pachycondyla goeldii. J. Biol. Chem. 2001, 276, 17823–17829. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.R.; Copello, J.A.; Evans, M.S.; Suarez, A.V. A biochemical characterization of the major peptides from the Venom of the giant Neotropical hunting ant Dinoponera australis. Toxicon 2010, 55, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, A.; Moggio, L.; Montella, R.C.; Campiglia, P.; Iannaccone, M.; Capuano, F.; Pedone, C.; Capparelli, R. Peptides from Royal Jelly: Studies on the antimicrobial activity of jelleins, jelleins analogs and synergy with temporins. J. Pept. Sci. 2011, 17, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Fontana, R.; Mendes, M.A.; de Souza, B.M.; Konno, K.; César, L.M.; Malaspina, O.; Palma, M.S. elleines: A family of antimicrobial peptides from the Royal Jelly of honeybees (Apis mellifera). Peptides 2004, 5, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Wang, J.; Peng, J; Zhao, P.; Kong, Z.; Wang, K.; Yan, W.; Wang, R. The in vitro, in vivo antifungal activity and the action mode of Jelleine-I against Candida species. Amino Acids 2018, 50, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Barnutiu, L.I.; Marghitas, L.A.; Dezmirean, D.S.; Bobis, O.; Mihai, C.M.; Pavel, C. Antimicrobial compounds of Royal Jelly. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca 2011, 68, 85–90. [Google Scholar]
- Bilikova, K.; Hanes, J.; Nordhoff, E.; Saenger, W.; Klaudiny, J.; Simuth, J. Apisimin. A new serine-valine-rich peptide from honeybee (Apis mellifera L.) royal jelly: Purification and molecular characterization. FEBS Lett. 2002, 528, 125–129. [Google Scholar] [CrossRef]
- Shen, L.L.; Xing, Y.Y.; Gao, Q. Sequence analysis of functional Apisimin-2 cDNA from royal jelly of Chinese honeybee and its expression in Escherichia coli. Asia Pac. J. Clin. Nutr. 2007, 16, 222–226. [Google Scholar] [PubMed]
- Gannabathula, S.; Krissansen, G.W.; Skinner, M.; Steinhorn, G.; Schlothauer, R. Honeybee apisimin and plant arabinogalactans in honey costimulate monocytes. Food Chem. 2015, 168, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Cociancich, S.; Dupont, A.; Hegy, G.; Lanot, R.; Holder, F.; Hetru, C.; Hoffmann, J.A.; Bulet, P. Novel inducible antibacterial peptides from a hemipteran insect, the sap-sucking bug Pyrrhocoris apterus. Biochem. J. 1994, 300, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kragol, G.; Lovas, S.; Varadi, G.; Condie, B.A.; Hoffmann, R.; Otvos, L., Jr. The antibacterial peptide pyrrhocoricin inhibits the ATPase actions of DnaK and prevents chaperone-assisted protein folding. Biochemistry 2001, 40, 3016–3026. [Google Scholar] [CrossRef]
- Chesnokova, L.S.; Slepenkov, S.V.; Witt, S.N. The insect antimicrobial peptide, L-pyrrhocoricin, binds to and stimulates the ATPase activity of both wild-type and lidless DnaK. FEBS Lett. 2004, 565, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Kragol, G.; Hoffmann, R.; Chattergoon, M.A.; Lovas, S.; Cudic, M.; Bulet, P.; Condie, B.A.; Rosengren, K.J.; Montaner, L.J.; Otvos, L., Jr. Identification of crucial residues for the antibacterial activity of the proline-rich peptide, pyrrhocoricin. Eur. J. Biochem. 2002, 269, 4226–4237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boxell, A.; Lee, S.H.; Jefferies, R.; Watt, P.; Hopkins, R.; Reid, S.; Armson, A.; Ryan, U. Pyrrhocoricin as a potential drug delivery vehicle for Cryptosporidium parvum. Exp. Parasitol. 2008, 119, 301–303. [Google Scholar] [CrossRef] [PubMed]
- Rosengren, K.J.; Göransson, U.; Otvos, L., Jr.; Craik, D.J. Cyclization of pyrrhocoricin retains structural elements crucial for the antimicrobial activity of the native peptide. Biopolymers 2004, 76, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Konnai, S.; Yamada, S.; Imamura, S.; Nishikado, H.; Ito, T.; Onuma, M.; Ohashi, K. Identification and characterization of antimicrobial peptide, defensin, in the taiga tick, Ixodes persulcatus. Insect Mol. Biol. 2009, 18, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L. Gene organization of a novel defensin of Ixodes ricinus: First annotation of an intron/exon structure in a hard tick defensin gene and first evidence of the occurrence of two isoforms of one member of the arthropod defensin family. Insect Mol. Biol. 2007, 16, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, N.; Battsetseg, B.; Boldbaatar, D.; Miyoshi, T.; Xuan, X.; Oliver, J.H., Jr.; Fujisaki, K. Babesial vector tick defensin against Babesia sp. parasites. Infect. Immun. 2007, 75, 3633–3640. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, N.; Isogai, E.; Hiramatsu, K.; Sasaki, T. Activity of tick antimicrobial peptide from Ixodes persulcatus (persulcatusin) against cell membranes of drug-resistant Staphylococcus aureus. J. Antibiot. 2017, 70, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, N.; Saito, T.; Ohmura, T.; Kuroda, K.; Suita, K.; Ihara, K.; Isogai, E. Functional structure and antimicrobial activity of persulcatusin, an antimicrobial peptide from the hard tick Ixodes persulcatus. Parasites Vectors 2016, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Jamasbi, E.; Lucky, S.S.; Li, W.; Hossain, M.A.; Gopalakrishnakone, P.; Separovic, F. Effect of dimerized melittin on gastric cancer cells and antibacterial activ ity. Amino Acids 2018. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Guan, S.M.; Sun, W.; Fu, H. Melittin, the major pain-producing substance of bee venom. Neurosci. Bull. 2016, 32, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Bae, H. Anti-Inflammatory applications of melittin, a major component of bee venom: Detailed mechanism of action and adverse effects. Molecules 2016, 21, E616. [Google Scholar] [CrossRef] [PubMed]
- Socarras, K.M.; Theophilus, P.A.S.; Torres, J.P.; Gupta, K.; Sapi, E. Antimicrobial activity of bee venom and melittin against borrelia burgdorferi. Antibiotics 2017, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Singh, A.K.; Wu, X.; Lyu, Y.; Bhunia, A.K.; Narsimhan, G. Characterization of antimicrobial activity against Listeria and cytotoxicity of native melittin and its mutant variants. Colloids. Surf. B Biointerfaces 2016, 143, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Leandro, L.F.; Mendes, C.A.; Casemiro, L.A.; Vinholis, A.H.; Cunha, W.R.; de Almeida, R.; Martins, C.H. Antimicrobial activity of apitoxin, melittin and phospholipase A₂ of honey bee (Apis mellifera) venom against oral pathogens. Anais Acad. Bras. Ciênc. 2015, 87, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Picoli, T.; Peter, C.M.; Zani, J.L.; Waller, S.B.; Lopes, M.G.; Boesche, K.N.; Vargas, G.D.Á.; Hübner, S.O.; Fischer, G. Melittin and its potential in the destruction and inhibition of the biofilm formation by Staphylococcus aureus, Escherichia coli and Pseudomonas aeruginosa isolated from bovine milk. Microb. Pathog. 2017, 112, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Li, C.; Li, M.; Zong, X.; Han, D.; Chen, Y. Antimicrobial peptide melittin against Xanthomonas oryzae pv. oryzae, the bacterial leaf blight pathogen in rice. Appl. Microbiol. Biotechnol. 2016, 100, 5059–5067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.K.; Ma, Q.; Li, S.B.; Gao, H.W.; Tan, Y.X.; Gong, F.; Ji, S.P. RV-23, a melittin-related peptide with cell-selective antibacterial activity and high hemocompatibility. J. Microbiol. Biotechnol. 2016, 26, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Wiedman, G.; Fuselier, T.; He, J.; Searson, P.C.; Hristova, K.; Wimley, W.C. Highly efficient macromolecule-sized poration of lipid bilayers by a synthetically evolved peptide. J. Am. Chem. Soc. 2014, 136, 4724–4731. [Google Scholar] [CrossRef] [PubMed]
- Rady, I.; Siddiqui, I.A.; Rady, M.; Mukhtar, H. Melittin, a major peptide component of bee venom, and its conjugates in cancer therapy. Cancer Lett. 2017, 402, 16–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamasbi, E.; Mularski, A.; Separovic, F. Model membrane and cell studies of antimicrobial activity of melittin analogues. Curr. Top. Med. Chem. 2016, 16, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Wimley, W.C. How does melittin permeabilize membranes? Biophys. J. 2018, 114, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Sun, T.L.; Hung, W.C.; Huang, H.W. Process of inducing pores in membranes by melittin. Proc. Natl. Acad. Sci. USA 2013, 110, 14243–14248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiedman, G.; Herman, K.; Searson, P.; Wimley, W.C.; Hristova, K. The electrical response of bilayers to the bee venom toxin melittin: Evidence for transient bilayer permeabilization. Biochim. Biophys. Acta-Biomembr. 2013, 1828, 1357–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Lee, D.G. Melittin triggers apoptosis in Candida albicans through the reactive oxygen species-mediated mitochondria/caspasedependent pathway. FEMS Microbiol. Lett. 2014, 355, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Akbari, R.; Hakemi Vala, M.; Pashaie, F.; Bevalian, P.; Hashemi, A.; Pooshang Bagheri, K. Highly synergistic effects of melittin with conventional antibiotics against multidrug-resistant isolates of acinetobacter baumannii and pseudomonas aeruginosa. Microb. Drug Resist. 2018. [Google Scholar] [CrossRef] [PubMed]
- Akbari, R.; Hakemi Vala, M.; Hashemi, A.; Aghazadeh, H.; Sabatier, J.M.; Pooshang Bagheri, K. Action mechanism of melittin-derived antimicrobial peptides, MDP1 and MDP2, de novo designed against multidrug resistant bacteria. Amino Acids 2018, 50, 1231–1243. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jang, A.Y.; Lin, S.; Lim, S.; Kim, D.; Park, K.; Han, S.M.; Yeo, J.H.; Seo, H.S. Melittin, a honeybee venom-derived antimicrobial peptide, may target methicillin-resistant Staphylococcus aureus. Mol. Med. Rep. 2015, 12, 6483–6490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hou, C.; Shamsi, I.H.; Ali, E.; Muhammad, N.; Shah, J.M.; Abid, A.A. Identification of super antibiotic-resistant bacteria in diverse soils. Int. J. Agric. Biol. 2015, 17, 1133–1140. [Google Scholar] [CrossRef]
- Lee, J.K.; Luchian, T.; Park, Y. New antimicrobial peptide kills drug-resistant pathogens without detectable resistance. Oncotarget 2018, 9, 15616–15634. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, Z.; Li, X.; Tian, Y.; Fan, Y.; Yu, C.; Zhou, B.; Liu, Y.; Xiang, R.; Yang, L. Synergistic effects of antimicrobial peptide DP7 combined with antibiotics against multidrug-resistant bacteria. Drug Des. Devel. Ther. 2017, 11, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Zerweck, J.; Strandberg, E.; Burck, J. Reichert, J.; Wadhwani, P.; Kukharenko, O.; Ulrich, A.S. Homo- and heteromeric interaction strengths of the synergistic antimicrobial peptides PGLa and magainin 2 in membranes. Eur. Biophys. J. Biophy. Lett. 2016, 45, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Biswaro, L.S.; da Costa Sousa, M.G.; Rezende, T.M.B.; Dias, S.C.; Franco, O.L. Antimicrobial peptides and nanotechnology, recent advances and challenges. Front. Microbiol. 2018, 9, 855. [Google Scholar] [CrossRef] [PubMed]
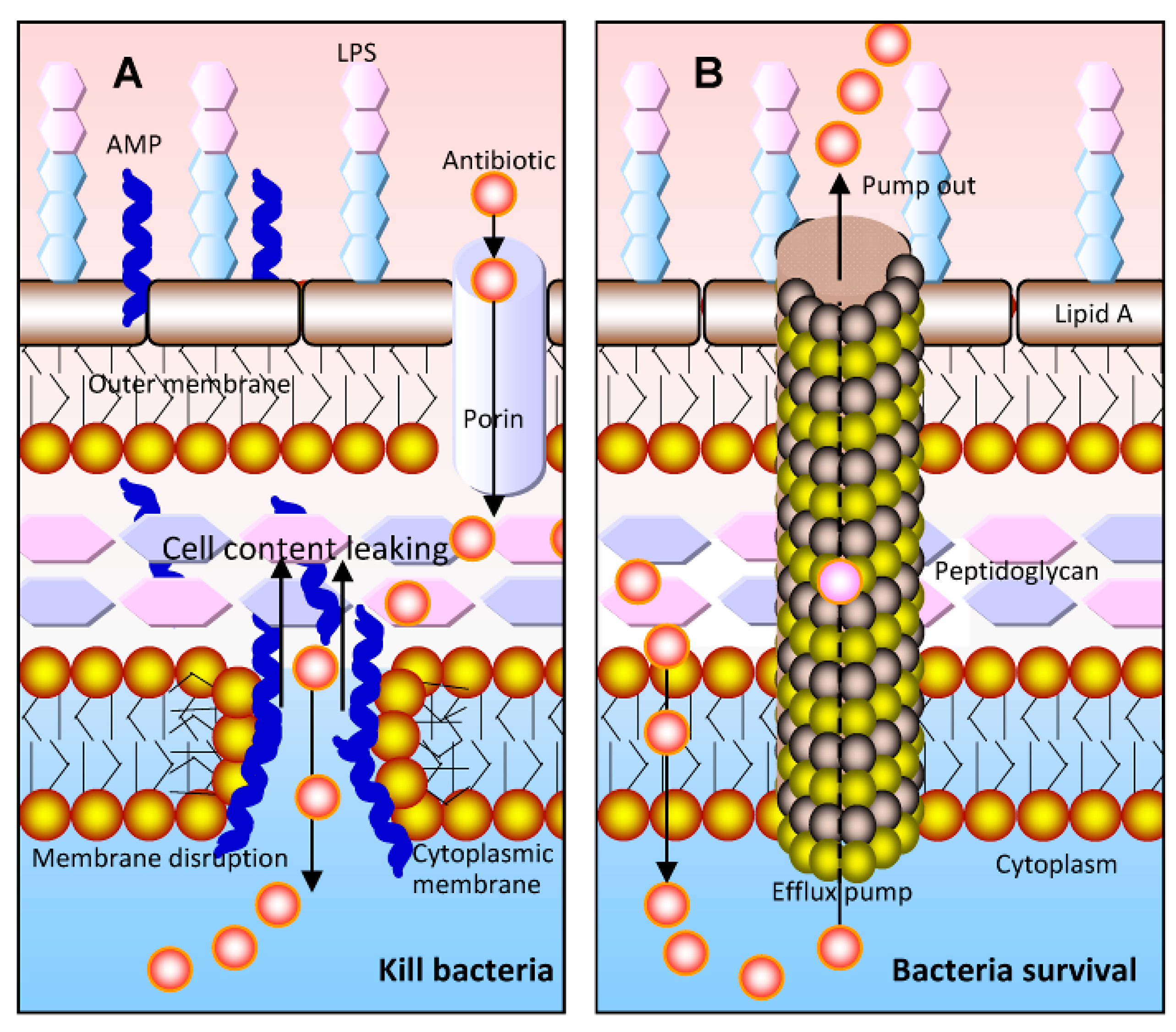
Figure 1.
Combined effects of antimicrobial peptides (AMPs) and antibiotics on bacteria. (A) AMPs can disrupt the bacterial membrane to cause the leakage of the cell content into the extracellular medium and kill the bacteria. The AMPs can facilitate more antibiotics to enter the cytoplasm of bacteria and finally interact with their target. However, the leakage of the antibiotics from the cytoplasm should not be ignored; (B) in bacterial cells, antibiotics are pumped out of the cells by the multidrug efflux pumps, which is how bacteria exert their resistance properties (adapted from [11]).
Figure 1.
Combined effects of antimicrobial peptides (AMPs) and antibiotics on bacteria. (A) AMPs can disrupt the bacterial membrane to cause the leakage of the cell content into the extracellular medium and kill the bacteria. The AMPs can facilitate more antibiotics to enter the cytoplasm of bacteria and finally interact with their target. However, the leakage of the antibiotics from the cytoplasm should not be ignored; (B) in bacterial cells, antibiotics are pumped out of the cells by the multidrug efflux pumps, which is how bacteria exert their resistance properties (adapted from [11]).
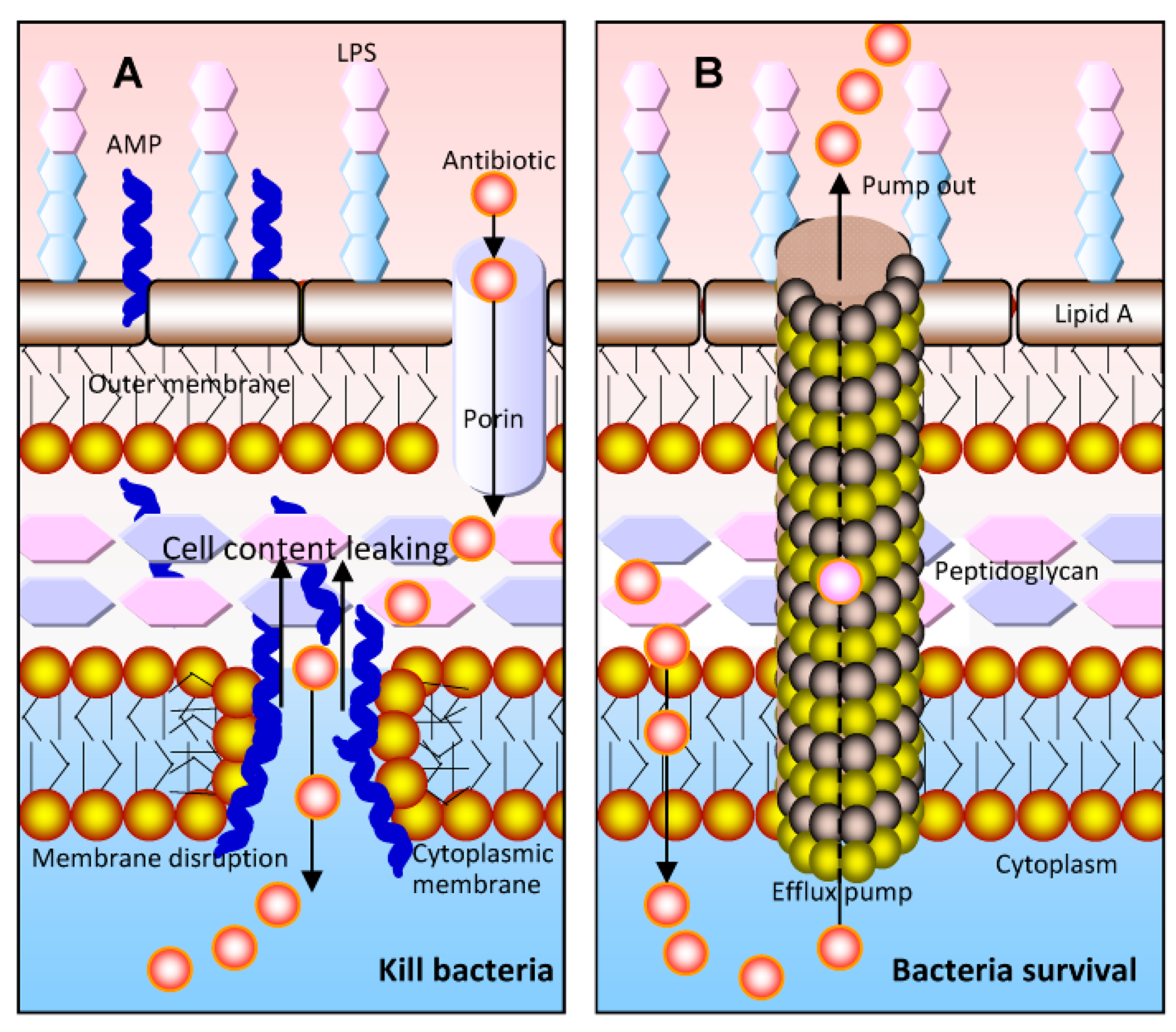
Figure 2.
Amino-terminal sequence of attacins A–F. Their sequence difference can be observed from the highlighted residues.
Figure 2.
Amino-terminal sequence of attacins A–F. Their sequence difference can be observed from the highlighted residues.


{kind=link}
{kind=link}
Table 1.
The amino acid sequences of cecropins.
| Name | Amino Acid Sequence | Reference |
|---|---|---|
| Cecropin A | GGLKKLGKKLEGVGKRVFKASEKALPVAVGIKALG-NH2 | [47] |
| Cecropin B | KWKVFKKIEKMGRNIRNGIVKAGPAIAVLGEAKAL-NH2 | [47] |
| Cecropin B1 | KWKVFKKIEKMGRNIRNGIVKAGPKWKVFKKIEK-NH2 | [53] |
| Cecropin B3 | AIAVLGEAKALMGRNIRNGIVKAGPAIAVLGEAKAL-NH2 | [53] |
| Cecropin C | GWLKKLGKRIERIGQHTRDATIQGLGIAQQAANVAATAR-NH2 | [48] |
| Cecropin D | WNPFKELEKVGQRVRDAVISAGPAVATVAQATALAK-NH2 | [48] |
| Cecropin P1 | SWLSKTAKKLENSAKKRISEGIAIAIQGGPR-NH2 | [54] |
Table 2.
The amino acid sequences of ponericins, which are antibacterial insect peptides (According to [106]).
Table 2.
The amino acid sequences of ponericins, which are antibacterial insect peptides (According to [106]).
| Name | Amino Acid Sequence |
|---|---|
| Ponericin G1 | GWKDWAKKAGGWLKKKGPGMAKAALKAAMQ-NH2 |
| Ponericin G2 | GWKDWLKKGKEWLKAKGPGIVKAALQAATQ-NH2 |
| Ponericin G3 | GWKDWLNKGKEWLKKKGPGIMKAALKAATQ-NH2 |
| Ponericin G4 | DFKDWMKTAGEWLKKKGPGILKAAMAAAT-NH2 |
| Ponericin G5 | GLKDWVKIAGGWLKKGPGILKAAMAAATQ-NH2 |
| Ponericin G6 | GLVDVLGKVGGLIKKLLP-NH2 |
| Ponericin G7 | GLVDVLGKVG GLIKKLLPG-NH2 |
| Ponericin W1 | WLGSALKIGAKLLPSVVGLFKKKKQ-NH2 |
| Ponericin W2 | WLGSALKIGAKLLPSVVGLFQKKKK-NH2 |
| Ponericin W3 | GIWGTLAKIGIKAVPRVISMLKKKKQ-NH2 |
| Ponericin W4 | GIWGTALKWGVKLLPKLVGMAQTKKQ-NH2 |
| Ponericin W5 | FWGALIKGAAKLIPSVVGLFKKKQ-NH2 |
| Ponericin W6 | FIGTALGIASAIPAIVKLFK-NH2 |
| Ponericin L1 | LLKELWTKMKGAGKAVLGKI-NH2 |
| Ponericin L2 | LLKELWTKIKGAGKAVLGKIKGLL-NH2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, Q.; Patočka, J.; Kuča, K. Insect Antimicrobial Peptides, a Mini Review. Toxins 2018, 10, 461. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110461
AMA Style
Wu Q, Patočka J, Kuča K. Insect Antimicrobial Peptides, a Mini Review. Toxins. 2018; 10(11):461. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110461
Chicago/Turabian StyleWu, Qinghua, Jiří Patočka, and Kamil Kuča. 2018. "Insect Antimicrobial Peptides, a Mini Review" Toxins 10, no. 11: 461. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110461
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.