Delayed LY333013 (Oral) and LY315920 (Intravenous) Reverse Severe Neurotoxicity and Rescue Juvenile Pigs from Lethal Doses of Micrurus fulvius (Eastern Coral Snake) Venom

 ,
,
Abstract
:1. Introduction
2. Results
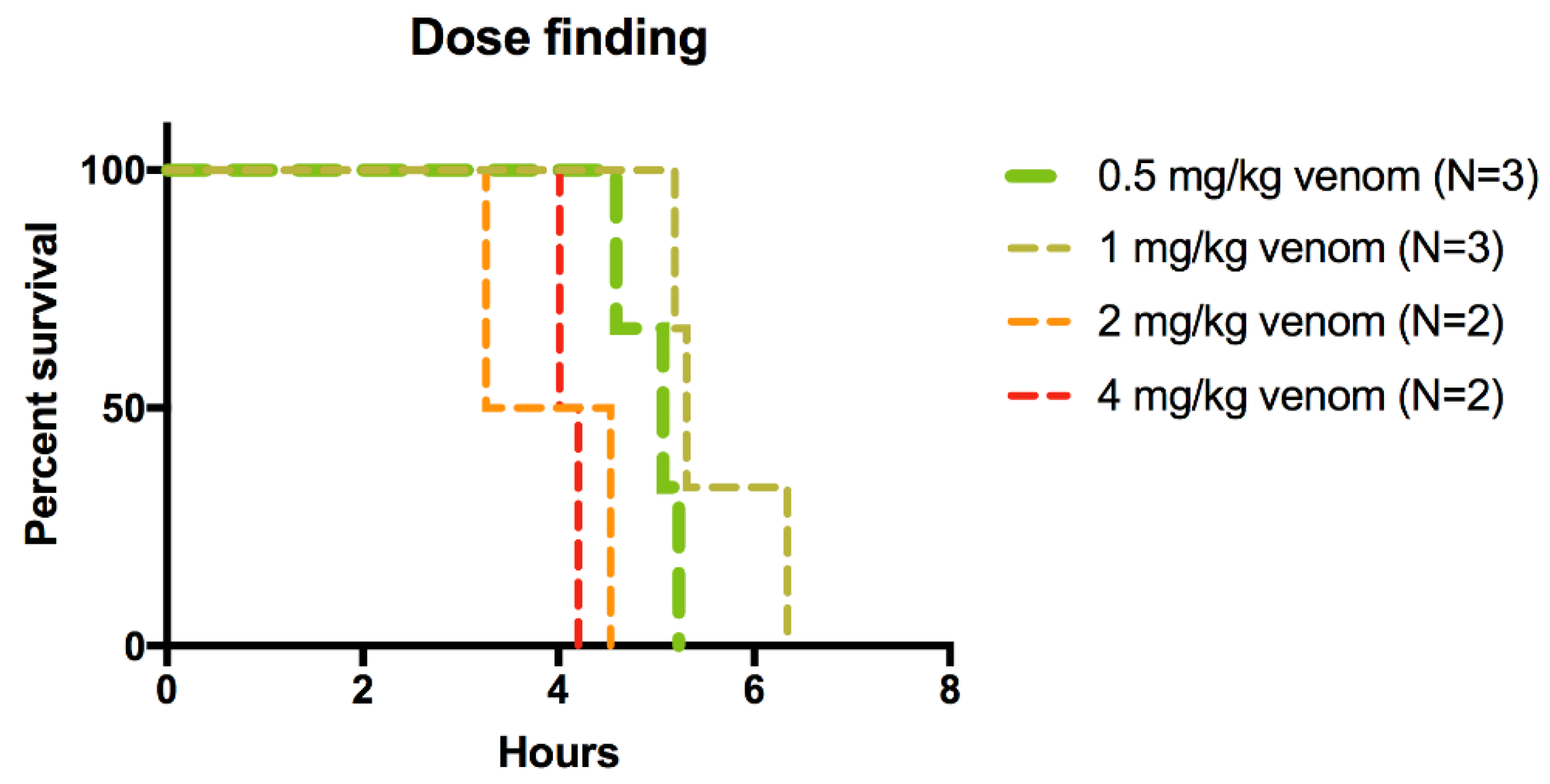
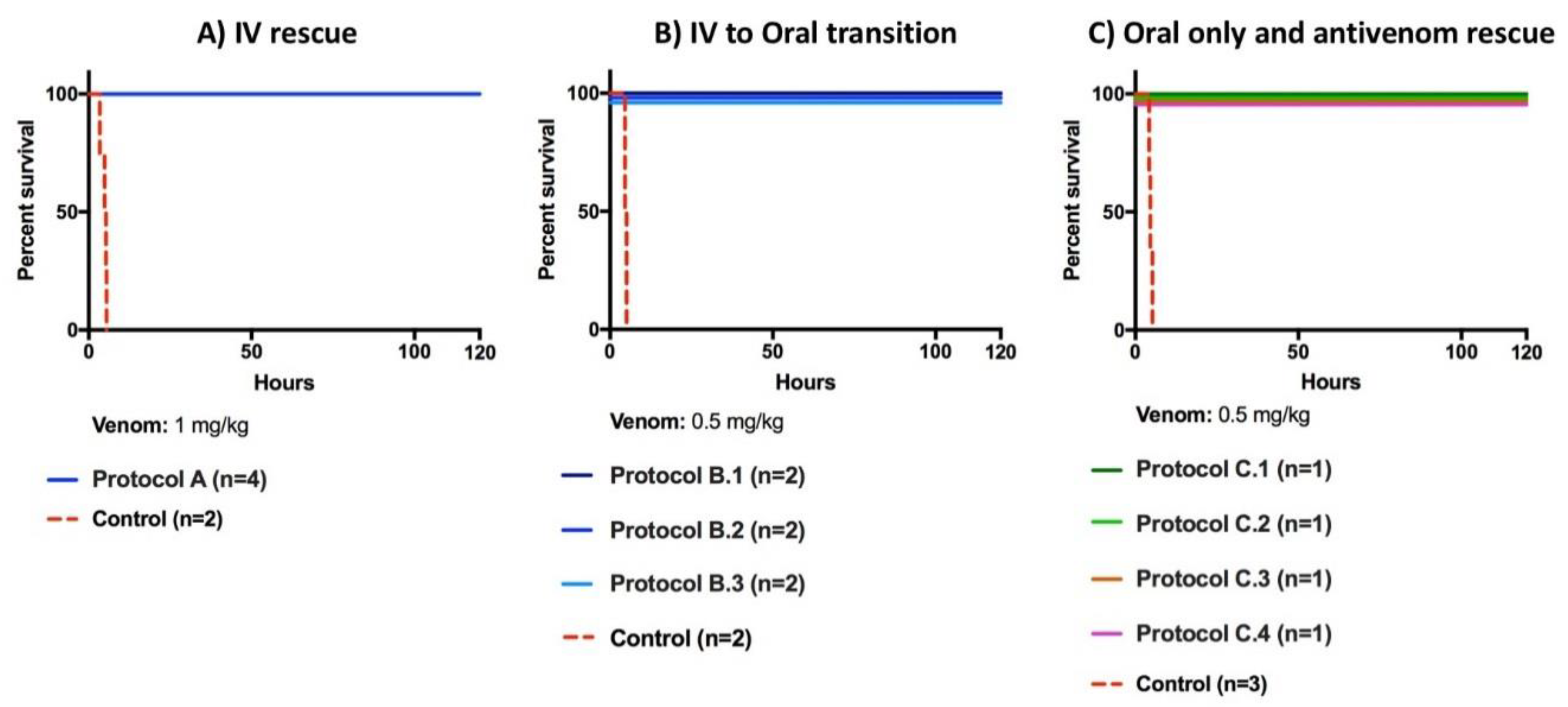
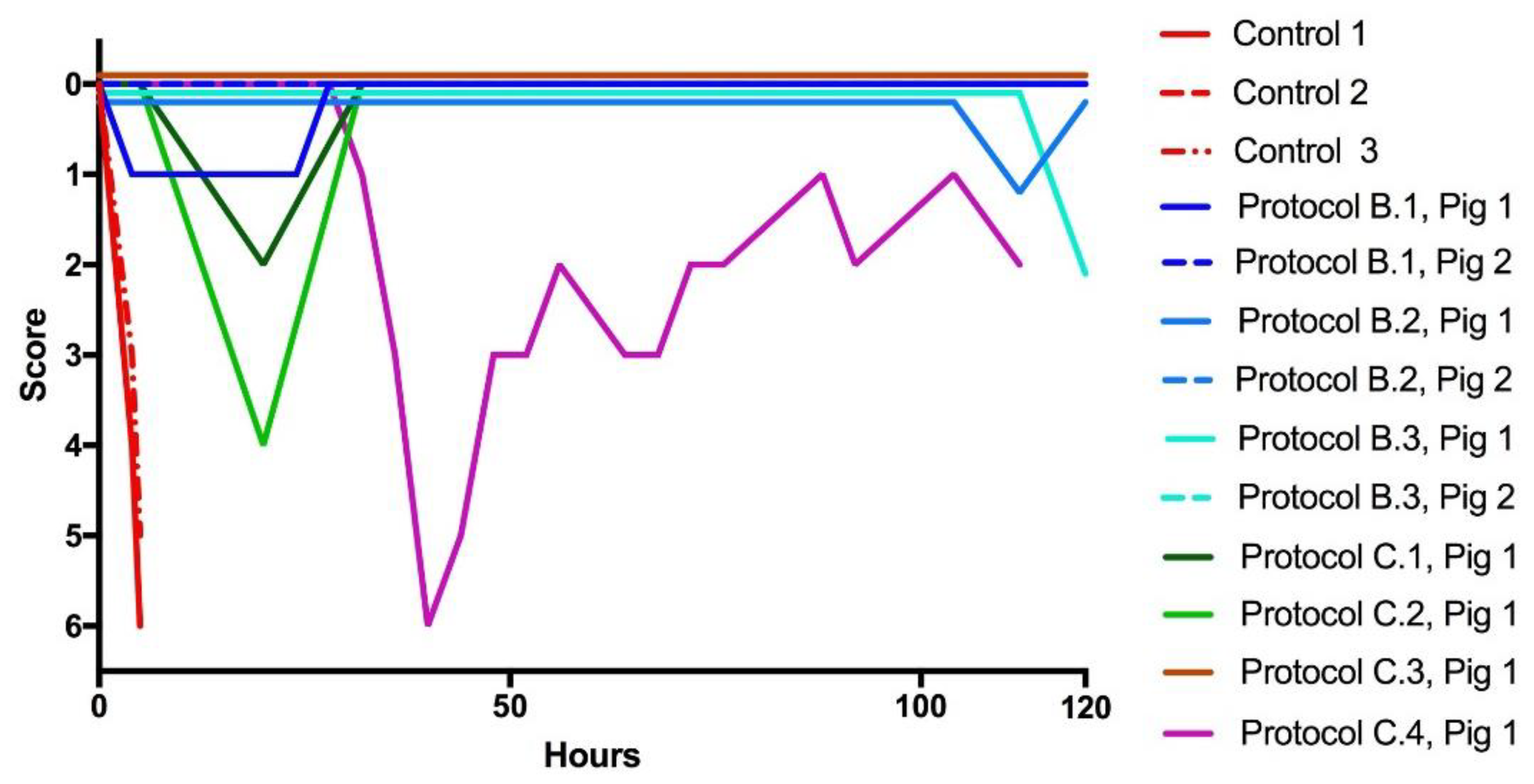
2.1. Rescue Experiments
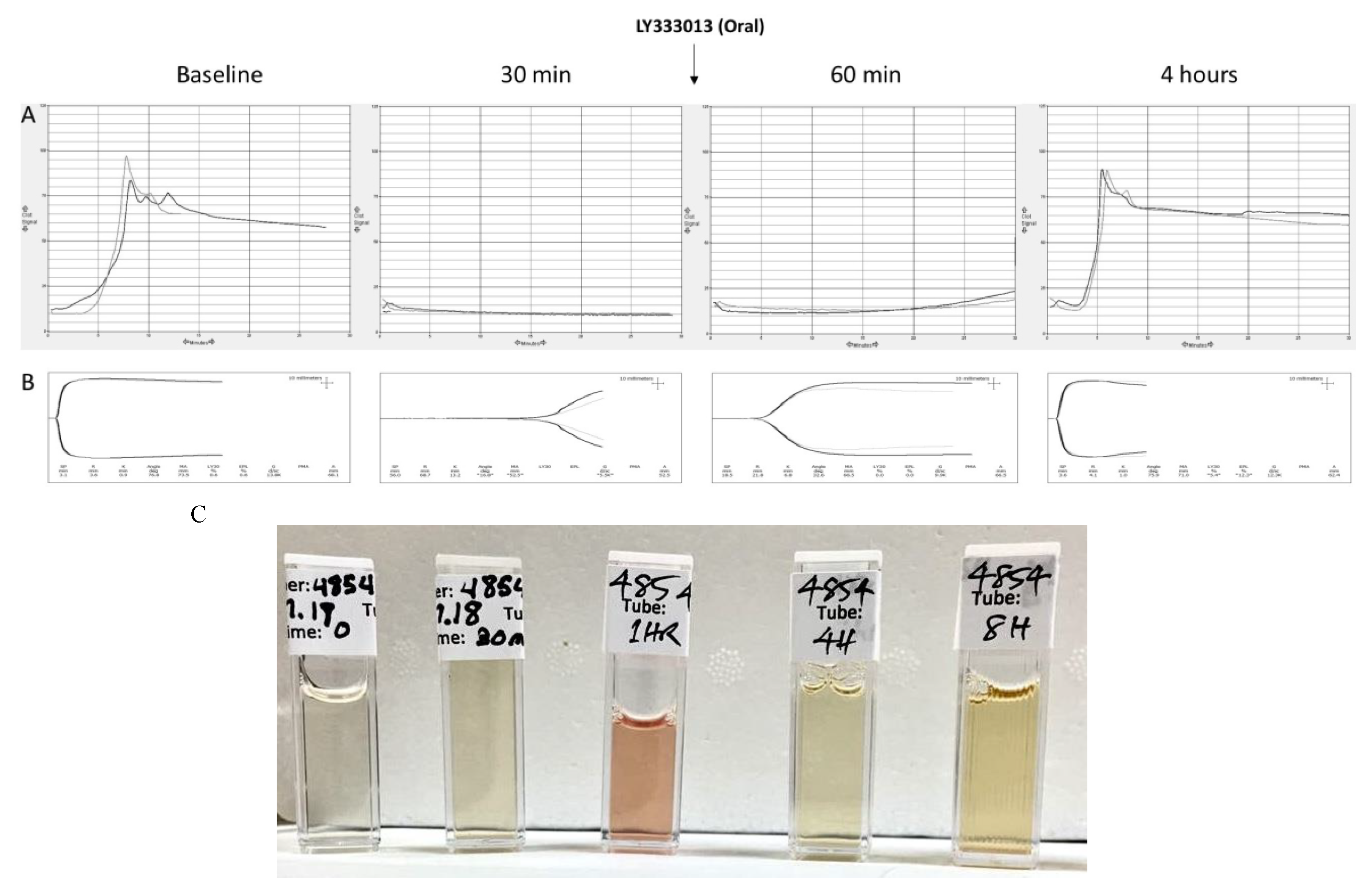
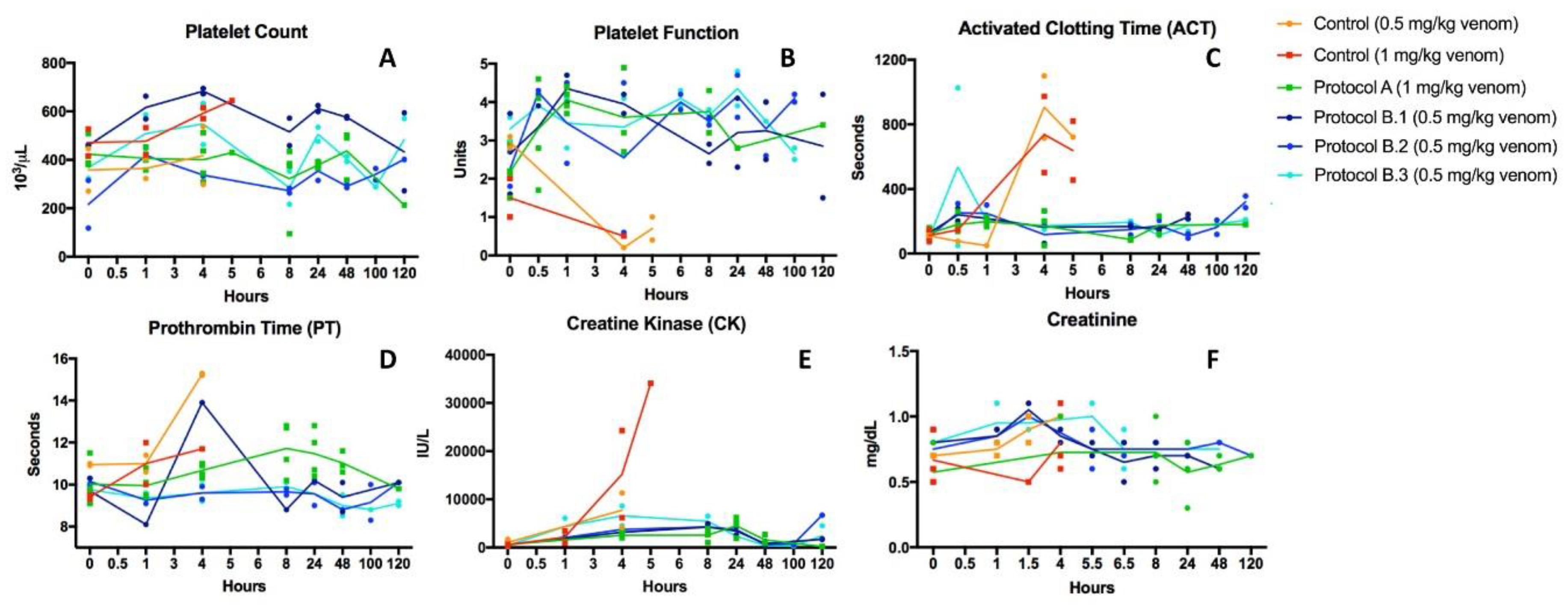
2.2. Recovery from Coagulopathy
2.3. Recovery from Neurotoxicity
2.4. Limitations
3. Discussion
4. Materials and Methods
4.1. Venom
4.2. Antidotes, Excipients, and Antivenom
4.3. Animals
4.4. Ethical Approval
4.5. Study Period
4.6. Envenoming and Rescue Experiment Protocols
4.7. Clinical Assessment of the Severity of Envenoming
4.8. Laboratory Tests
5. Materials and Methods Detail
- (1)
- Every 15 min post venom administration for the first 4 h following recovery from general anesthesia.
- (2)
- From 4 h to 8 h post venom administration, pigs were given a clinical and lameness (veterinary surrogate for pain) score every 30 min.
- (3)
- From 8 h to 48 h post venom administration pigs were given a clinical and lameness score every hour.
- (4)
- From 48 h to 96 h pigs were given a clinical and lameness score every 6 h.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Editorial Board. Snake-bite envenoming: A priority neglected tropical disease. Lancet (London, England) 2017, 390, 2. [Google Scholar] [CrossRef]
- Sharma, S.K.; Chappuis, F.F.; Jha, N.; Bovier, P.A.; Loutan, L.; Koirala, S. Impact of snake bites and determinants of fatal outcomes in Southeastern Nepal. Am. J. Trop. Med. Hyg. 2004, 71, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Longbottom, J.; Shearer, F.M.; Devine, M.; Alcoba, G.; Chappuis, F.; Weiss, D.J.; Ray, S.E.; Ray, N.; Warrell, D.A.; Ruiz de Castañeda, R.; et al. Vulnerability to snakebite envenoming: A global mapping of hotspots. Lancet (London, England) 2018, 392, 673–684. [Google Scholar] [CrossRef]
- Vaiyapuri, S.; Vaiyapuri, R.; Ashokan, R.; Ramasamy, K.; Nattamaisundar, K.; Jeyaraj, A.; Chandran, V.; Gajjeraman, P. Snakebite and Its Socio-Economic Impact on the Rural Population of Tamil Nadu, India. PLoS ONE 2013, 8, e80090. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.A.; Hargreaves, A.; Wagstaff, S.C.; Faragher, B.; Lalloo, D.G. Snake Envenoming: A Disease of Poverty. PLoS Negl. Trop. Dis. 2009, 3, e569. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Prim. 2017, 3, 17063. [Google Scholar] [CrossRef]
- Lewin, M.; Samuel, S.; Merkel, J.; Bickler, P. Varespladib (LY315920) Appears to Be a Potent, Broad-Spectrum, Inhibitor of Snake Venom Phospholipase A2 and a Possible Pre-Referral Treatment for Envenomation. Toxins (Basel) 2016, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Engmark, M.; Milbo, C.; Johannesen, J.; Lomonte, B.; Gutierrez, J.M.; Lohse, B. From Fangs to Pharmacology: The Future of Snakebite Envenoming Therapy. Curr. Pharm. Des. 2016, 22, 5270–5293. [Google Scholar] [CrossRef] [PubMed]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins (Basel) 2017, 9, 290. [Google Scholar] [CrossRef]
- Calvete, J.J. Proteomic tools against the neglected pathology of snake bite envenoming. Expert Rev. Proteomics 2011, 8, 739–758. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.B.; Grubb, B.D.; Maltin, C.A.; Dixon, R. The neurotoxicity of the venom phospholipases A(2), notexin and taipoxin. Exp. Neurol. 2000, 161, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Ownby, C.L. Skeletal muscle degeneration induced by venom phospholipases A2: Insights into the mechanisms of local and systemic myotoxicity. Toxicon 2003, 42, 915–931. [Google Scholar] [CrossRef]
- Clemetson, K.J.; Lu, Q.; Clemetson, J.M. Snake venom proteins affecting platelets and their applications to anti-thrombotic research. Curr. Pharm. Des. 2007, 13, 2887–2892. [Google Scholar] [CrossRef] [PubMed]
- Stief, T.W. Phospholipase A(2) activates hemostasis. Drug Target Insights 2007, 2, 83–96. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Greenhalgh, A.D.; López-Vales, R. Role of phospholipase A2s and lipid mediators in secondary damage after spinal cord injury. Cell Tissue Res. 2012, 349, 249–267. [Google Scholar] [CrossRef] [PubMed]
- Titsworth, W.L.; Liu, N.-K.; Xu, X.-M. Role of secretory phospholipase a(2) in CNS inflammation: Implications in traumatic spinal cord injury. CNS Neurol. Disord. Drug Targets 2008, 7, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Uhl, W.; Büchler, M.; Nevalainen, T.J.; Deller, A.; Beger, H.G. Serum phospholipase A2 in patients with multiple injuries. J. Trauma 1990, 30, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.-K.; Titsworth, W.L.; Zhang, Y.P.; Xhafa, A.I.; Shields, C.B.; Xu, X.-M. Characterizing phospholipase A2-induced spinal cord injury-a comparison with contusive spinal cord injury in adult rats. Transl. Stroke Res. 2011, 2, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Lomonte, B.; Sanz, L.; Calvete, J.J.; Pla, D. Immunological profile of antivenoms: Preclinical analysis of the efficacy of a polyspecific antivenom through antivenomics and neutralization assays. J. Proteomics 2014, 105, 340–350. [Google Scholar] [CrossRef]
- Sanhajariya, S.; Duffull, S.B.; Isbister, G.K. Pharmacokinetics of Snake Venom. Toxins (Basel) 2018, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Gutiérrez, J.M.; Knudsen, C.; Johansen, K.H.; Bermúdez-Méndez, E.; Cerni, F.A.; Jürgensen, J.A.; Ledsgaard, L.; Martos-Esteban, A.; Øhlenschlæger, M.; et al. Pros and cons of different therapeutic antibody formats for recombinant antivenom development. Toxicon 2018, 146, 151–175. [Google Scholar] [CrossRef] [PubMed]
- Adis R&D Profile Varespladib. Am. J. Cardiovasc. Drugs 2011, 11, 137–143.
- Wang, Y.; Zhang, J.; Zhang, D.; Xiao, H.; Xiong, S.; Huang, C. Exploration of the Inhibitory Potential of Varespladib for Snakebite Envenomation. Molecules 2018, 23, 391. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Li, H.; Zhang, D.; Li, Y.; Sun, S.; Huang, C.; Xiao, H.; Li, H.; Zhang, D.; Li, Y.; Sun, S.; Huang, C. Inactivation of Venom PLA2 Alleviates Myonecrosis and Facilitates Muscle Regeneration in Envenomed Mice: A Time Course Observation. Molecules 2018, 23, 1911. [Google Scholar] [CrossRef]
- Dennis, E.A.; Cao, J.; Hsu, Y.-H.; Magrioti, V.; Kokotos, G. Phospholipase A2 Enzymes: Physical Structure, Biological Function, Disease Implication, Chemical Inhibition, and Therapeutic Intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed]
- Lewin, M.R.; Gutiérrez, J.M.; Samuel, S.P.; Herrera, M.; Bryan-Quirós, W.; Lomonte, B.; Bickler, P.E.; Bulfone, T.C.; Williams, D.J. Delayed Oral LY333013 Rescues Mice from Highly Neurotoxic, Lethal Doses of Papuan Taipan (Oxyuranus scutellatus) Venom. Toxins (Basel) 2018, 10, 380. [Google Scholar] [CrossRef] [PubMed]
- Vergara, I.; Pedraza-Escalona, M.; Paniagua, D.; Restano-Cassulini, R.; Zamudio, F.; Batista, C.V.F.; Possani, L.D.; Alagón, A. Eastern coral snake Micrurus fulvius venom toxicity in mice is mainly determined by neurotoxic phospholipases A2. J. Proteomics 2014, 105, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Arce-Bejarano, R.; Lomonte, B.; Gutiérrez, J.M. Intravascular hemolysis induced by the venom of the Eastern coral snake, Micrurus fulvius, in a mouse model: Identification of directly hemolytic phospholipases A2. Toxicon 2014, 90, 26–35. [Google Scholar] [CrossRef] [PubMed]
- De Roodt, A.R.; Lago, N.R.; Stock, R.P. Myotoxicity and nephrotoxicity by Micrurus venoms in experimental envenomation. Toxicon 2012, 59, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Smyrnioudis, M.E.; O’Rourke, D.P.; Rosenbaum, M.D.; Brewer, K.L.; Meggs, W.J. Long-term efficacy of pressure immobilization bandages in a porcine model of coral snake envenomation. Am. J. Emerg. Med. 2014, 32, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- German, B.T.; Hack, J.B.; Brewer, K.; Meggs, W.J. Pressure-Immobilization Bandages Delay Toxicity in a Porcine Model of Eastern Coral Snake (Micrurus fulvius fulvius) Envenomation. Ann. Emerg. Med. 2005, 45, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Parker-Cote, J.L.; O’Rourke, D.P.; Brewer, K.L.; Lertpiriyapong, K.; Punja, M.; Bush, S.P.; Miller, S.N.; Meggs, W.J. Efficacy of Trypsin in Treating Coral Snake Envenomation in the Porcine Model. J. Med. Toxicol. 2015, 11, 430–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hack, J.B.; Deguzman, J.M.; Brewer, K.L.; Meggs, W.J.; O’Rourke, D.; O’rourke, D. A Localizing Circumferential Compression Device Increases Survival after Coral Snake Envenomation to the Torso of an Animal Model. JEM 2011, 41, 102–107. [Google Scholar] [CrossRef] [PubMed]
- LD50 and Venom Yields|Snakedatabase.org. Available online: http://snakedatabase.org/pages/ld50.php (accessed on 28 August 2018).
- Silva, A.; Johnston, C.; Kuruppu, S.; Kneisz, D.; Maduwage, K.; Kleifeld, O.; Smith, A.I.; Siribaddana, S.; Buckley, N.A.; Hodgson, W.C.; et al. Clinical and Pharmacological Investigation of Myotoxicity in Sri Lankan Russell’s Viper (Daboia russelii) Envenoming. PLoS Negl. Trop. Dis. 2016, 10, e0005172. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Hodgson, W.; Isbister, G. Cross-Neutralisation of In Vitro Neurotoxicity of Asian and Australian Snake Neurotoxins and Venoms by Different Antivenoms. Toxins (Basel) 2016, 8, 302. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.L.; Quartino, P.Y.; Arce-Bejarano, R.; Fernández, J.; Camacho, L.F.; Gutiérrez, J.M.; Kuemmel, D.; Fidelio, G.; Lomonte, B. Intravascular hemolysis induced by phospholipases A 2 from the venom of the Eastern coral snake, Micrurus fulvius: Functional profiles of hemolytic and non-hemolytic isoforms. Toxicol. Lett. 2018, 286, 39–47. [Google Scholar] [CrossRef]
- Davis, S.S.; Illum, L.; Hinchcliffe, M. Gastrointestinal transit of dosage forms in the pig. J. Pharm. Pharmacol. 2001, 53, 33–39. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Lomonte, B.; Rey-Suárez, P.; Fernández, J.; Sasa, M.; Pla, D.; Vargas, N.; Bénard-Valle, M.; Sanz, L.; Corrêa-Netto, C.; Núñez, V.; et al. Venoms of Micrurus coral snakes: Evolutionary trends in compositional patterns emerging from proteomic analyses. Toxicon 2016, 122, 7–25. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Structure-function relationships and mechanism of anticoagulant phospholipase A2 enzymes from snake venoms. Toxicon 2005, 45, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D. Snakes in Myth, Magic, and History: The Story of a Human Obsession; Praeger: Santa Barbara, CA, USA, 2008; ISBN 0313352925. [Google Scholar]
- Arias, A.S.; Rucavado, A.; Gutierrez, J.-M. Peptidomimetic hydroxamate metalloproteinase inhibitors abrogate local and systemic toxicity induced by Echis ocellatus (saw-scaled) snake venom. Toxicon 2017, 132, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Escalante, T.; Franceschi, A.; Rucavado, A.; Gutiérrez, J.M. Effectiveness of batimastat, a synthetic inhibitor of matrix metalloproteinases, in neutralizing local tissue damage induced by BaP1, a hemorrhagic metalloproteinase from the venom of the snake bothrops asper. Biochem. Pharmacol. 2000, 60, 269–274. [Google Scholar] [CrossRef]
- Herzel, B.; Samuel, S.P.; Bulfone, T.C.; Raj, C.S.; Lewin, M.R.; Kahn, J.G. Snakebite: An Exploratory Cost-Effectiveness Analysis of Adjunct Treatment Strategies. Am. J. Trop. Med. Hyg. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Hessel, M.M.; McAninch, S.A. Coral Snake Toxicity; StatPearls: Tampa, FL, USA, 2018. [Google Scholar]
- Sheikh, S.; Leffers, P. Emergency department management of North American snake envenomations. Emerg. Med. Pract. 2018, 20, 1–26. [Google Scholar] [PubMed]
- Sasaki, J.; Khalil, P.A.; Chegondi, M.; Raszynski, A.; Meyer, K.G.; Totapally, B.R. Coral Snake Bites and Envenomation in Children. Pediatr. Emerg. Care 2014, 30, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Kitchens, C.S.; Van Mierop, L.H. Envenomation by the Eastern coral snake (Micrurus fulvius fulvius). A study of 39 victims. JAMA 1987, 258, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.; Naum, C.; Bandi, V.; Gervich, D.; Lowry, S.F.; Wunderink, R.; Schein, R.M.; Macias, W.; Skerjanec, S.; Dmitrienko, A.; et al. Efficacy and safety of LY315920Na/S-5920, a selective inhibitor of 14-kDa group IIA secretory phospholipase A2, in patients with suspected sepsis and organ failure. Crit. Care Med. 2003, 31, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Rojas, G.; Jiménez, J.M.; Gutiérrez, J.M. Caprylic acid fractionation of hyperimmune horse plasma: Description of a simple procedure for antivenom production. Toxicon 1994, 32, 351–363. [Google Scholar] [CrossRef]
- Arce, V.; Rojas, E.; Ownby, C.L.; Rojas, G.; Gutiérrez, J.M. Preclinical assessment of the ability of polyvalent (Crotalinae) and anticoral (Elapidae) antivenoms produced in Costa Rica to neutralize the venoms of North American snakes. Toxicon 2003, 41, 851–860. [Google Scholar] [CrossRef]
- Flournoy, W.S.; Mani, S. Percutaneous external jugular vein catheterization in piglets using a triangulation technique. Lab. Anim. 2009, 43, 344–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larréché, S.; Jean, F.-X.; Benois, A.; Mayet, A.; Bousquet, A.; Vedy, S.; Clapson, P.; Dehan, C.; Rapp, C.; Kaiser, E.; et al. Thromboelastographic study of the snakebite-related coagulopathy in Djibouti. Blood Coagul. Fibrinol. 2018, 29, 196–204. [Google Scholar] [CrossRef]
- Hett, D.A.; Walker, D.; Pilkington, S.N.; Smith, D.C. Sonoclot analysis. Br. J. Anaesth. 1995, 75, 771–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number Animals per Group | Venom Dose (mg/kg) |
|---|---|---|
| 1 | 2 | 4 mg/kg |
| 2 | 2 | 2 mg/kg |
| 3 | 3 | 1 mg/kg |
| 4 | 3 | 0.5 mg/kg |
| Protocol | Number Animals per Group | Treatment | Venom Dose Level | Drug (and/or Antivenom) Dose |
|---|---|---|---|---|
| Protocol A | 4 | Venom | 1 mg/kg | -Loading dose: 5 mg/kg i.v. over 15 min. -2.5 mg/kg i.v. bolus q6 h × 24 h -1 mg/kg i.v. prn weakness/coagulopathy |
| + | ||||
| LY315920 (i.v.) | ||||
| Protocol B.1 | 2 | Venom | 0.5 mg/kg | -Loading dose: 1 mg/kg i.v. immediately -CRI i.v. 0.67 mg/kg/h × 24 h -Oral 200 mg q6 h × 4 then 100 mg PO q6 h prn weakness/coagulopathy |
| + | ||||
| LY315920 (i.v.) then LY333013 (p.o) | ||||
| Protocol B.2 | 2 | Venom | 0.5 mg/kg | -Loading dose: 1 mg/kg i.v. (15 min after envenoming) -CRI i.v. 0.67 mg/kg/h × 24 h -Oral loading dose of 400 mg PO 4 h prior to ending the CRI followed by 200 mg q6 h for 12 h then 100 mg PO q6 h prn weakness/coagulopathy |
| + | ||||
| LY315920 (i.v.) then LY333013 (p.o) | ||||
| Protocol B.3 | 2 | Venom | 0.5 mg/kg | -400 mg PO loading dose (15 min delay) -200 mg PO q6 h for 12 h -100 mg PO q6 h for 36 h -100 mg PO q6 h prn weakness/coagulopathy |
| + | ||||
| LY333013 (p.o) | ||||
| Protocol C.1 | 1 | Venom | 0.5 mg/kg | -46.4 mL antivenom i.v. (1 mL antivenom/0.125 mg venom) (15 min post-venom) |
| + | ||||
| Antivenom | ||||
| Protocol C.2 | 1 | Venom | 0.5 mg/kg | -67.2 mL antivenom i.v. (1 mL antivenom/0.125 mg venom) (>45 min post-venom) -67.2 mL antivenom i.v. prn weakness/coagulopathy (1 mL antivenom/0.125 mg venom) -Rescue with LY333013 200 mg PO × q6 h prn weakness/coagulopathy |
| + | ||||
| Antivenom | ||||
| + | ||||
| LY333013 (p.o) | ||||
| Protocol C.3 | 1 | Venom | 0.5 mg/kg | -200 mg PO (>45 min post-venom) -200 mg PO q6 24 h -100 mg PO q6 h for 24 h -200 mg PO weakness/coagulopathy |
| + | ||||
| LY333013 (p.o) | ||||
| Protocol C.4 | 1 | Venom | 0.5 mg/kg | -200 mg PO (>45 min delay) -200 mg PO q6 h for 12 h Drug withdrawn -Rescue with antivenom i.v. (1 mL/0.125 mg of venom) prn weakness/coagulopathy If no response to antivenom: -Rescue with 200 mg PO q6 h prn weakness/coagulopathy |
| + | ||||
| LY333013 (p.o) | ||||
| + | ||||
| Antivenom |
| Score | Clinical Picture |
|---|---|
| 0 | Normal activity, interested in food/water/toys, normal grunting, curious about environment, responds normally to stimulation by moving away, rises easily and quickly from recumbency when stimulated. |
| 1 | Normal movement around pen, reduced interest in food/water/toys, reduced interaction with environment or caretakers, rises from recumbency and moves away when stimulated. |
| 2 | Evidence of weakness in one or more limbs, reduced interest in food/water/toys, reduced responsiveness to stimulation but still able to rise normally from recumbency and remain standing >3 min. |
| 3 | Significant evidence of weakness (dog-like sitting, treading in back legs after rising) but able to rise unassisted when stimulated and stays standing >15 s but <3 min, some interest in food/water/toys. |
| 4 | Significant evidence of weakness (dog-like sitting, sternal or lateral, treading in back legs after rising), requires assistance to rise but can stand longer than 10 s on their own once assisted to stand. |
| 5 | Significant evidence of weakness (dog-like sitting, sternal or lateral, treading in back legs after rising), unable to rise without assistance, remains standing <10 s. |
| 6 | Marked evidence of weakness (sternal or lateral recumbency) unable to rise even with assistance. Meets criteria for Euthanasia |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lewin, M.R.; Gilliam, L.L.; Gilliam, J.; Samuel, S.P.; Bulfone, T.C.; Bickler, P.E.; Gutiérrez, J.M. Delayed LY333013 (Oral) and LY315920 (Intravenous) Reverse Severe Neurotoxicity and Rescue Juvenile Pigs from Lethal Doses of Micrurus fulvius (Eastern Coral Snake) Venom. Toxins 2018, 10, 479. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110479
Lewin MR, Gilliam LL, Gilliam J, Samuel SP, Bulfone TC, Bickler PE, Gutiérrez JM. Delayed LY333013 (Oral) and LY315920 (Intravenous) Reverse Severe Neurotoxicity and Rescue Juvenile Pigs from Lethal Doses of Micrurus fulvius (Eastern Coral Snake) Venom. Toxins. 2018; 10(11):479. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110479
Chicago/Turabian StyleLewin, Matthew R., Lyndi L. Gilliam, John Gilliam, Stephen P. Samuel, Tommaso C. Bulfone, Philip E. Bickler, and José María Gutiérrez. 2018. "Delayed LY333013 (Oral) and LY315920 (Intravenous) Reverse Severe Neurotoxicity and Rescue Juvenile Pigs from Lethal Doses of Micrurus fulvius (Eastern Coral Snake) Venom" Toxins 10, no. 11: 479. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10110479