Effective Biodegradation of Aflatoxin B1 Using the Bacillus licheniformis (BL010) Strain

 , , and
, , and
Abstract
:1. Introduction
2. Results
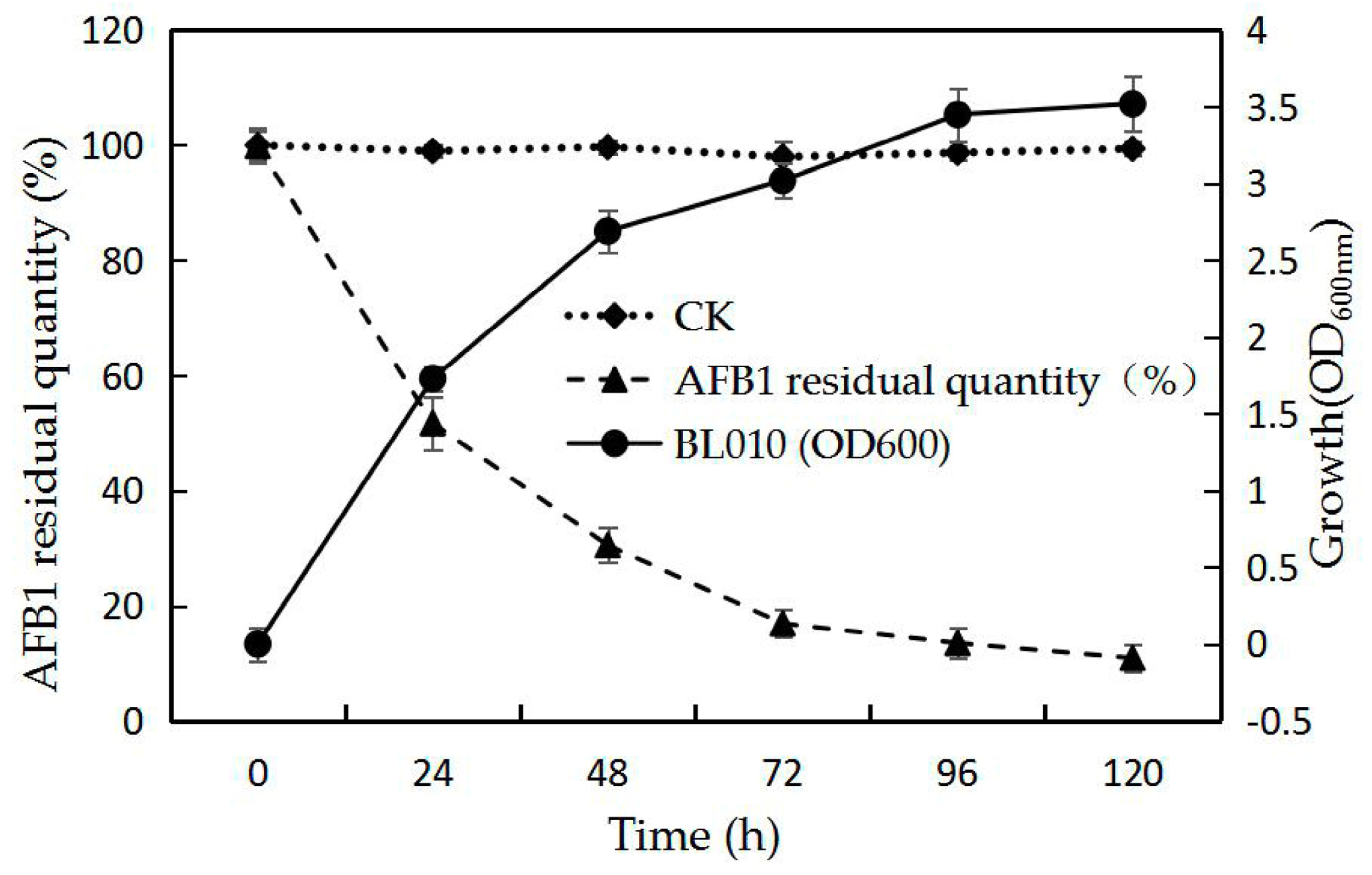
2.1. Biodegradation Kinetics of BL010 in ABS
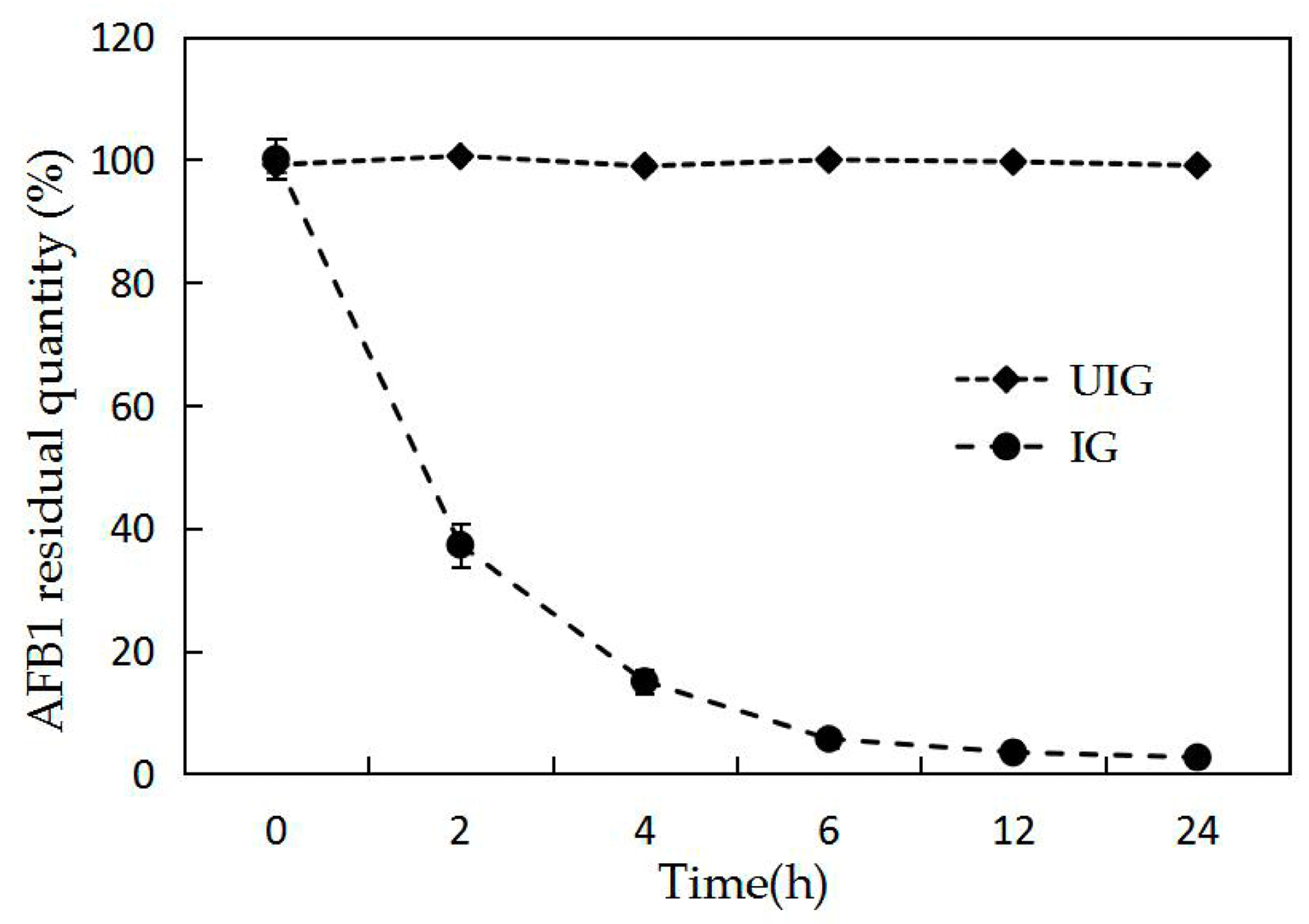
2.2. Effect of Induction on Degradation Enzymes
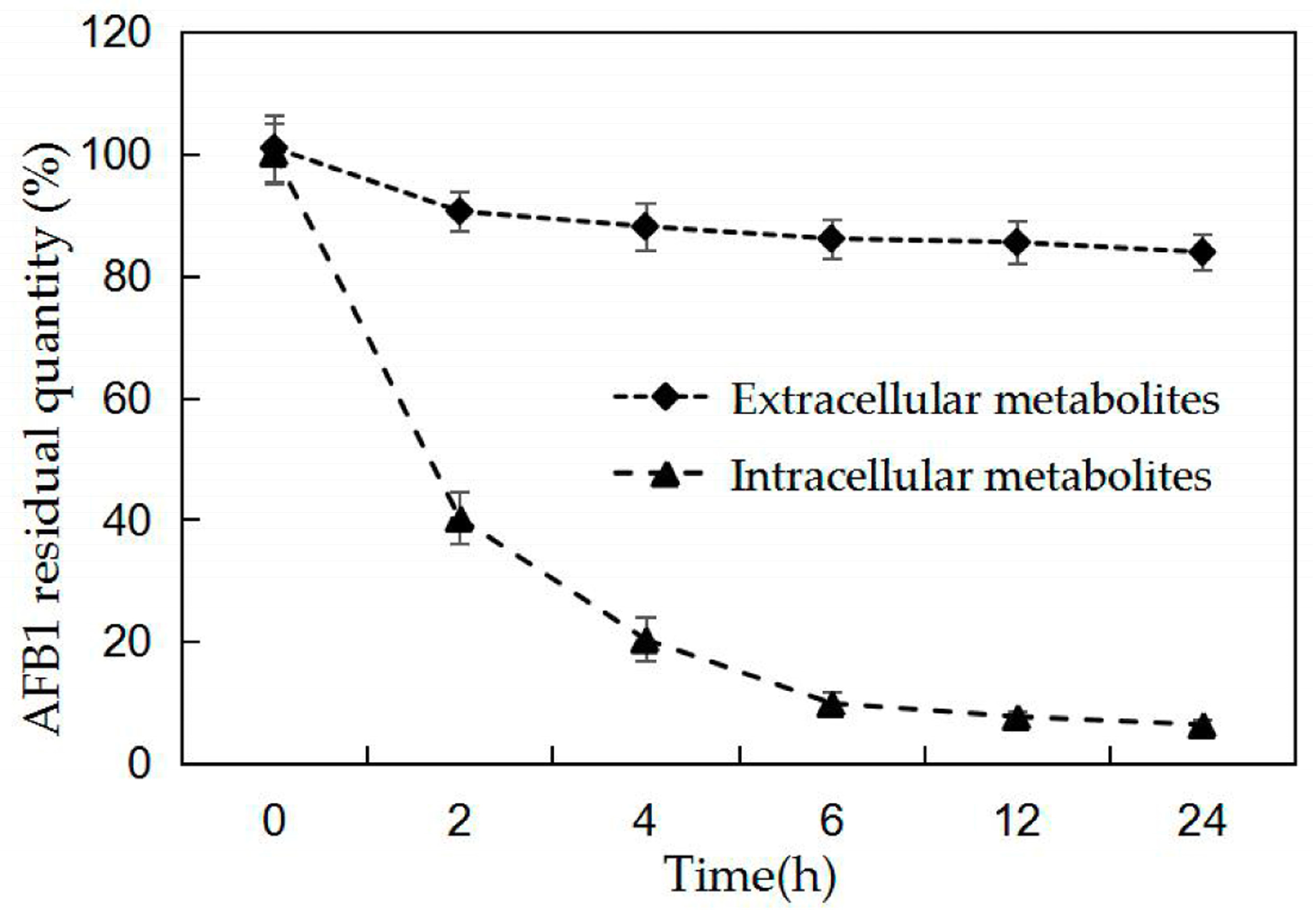
2.3. AFB1 Degradation by BL010 Cell Culture Fluid and Cell-Free Extract
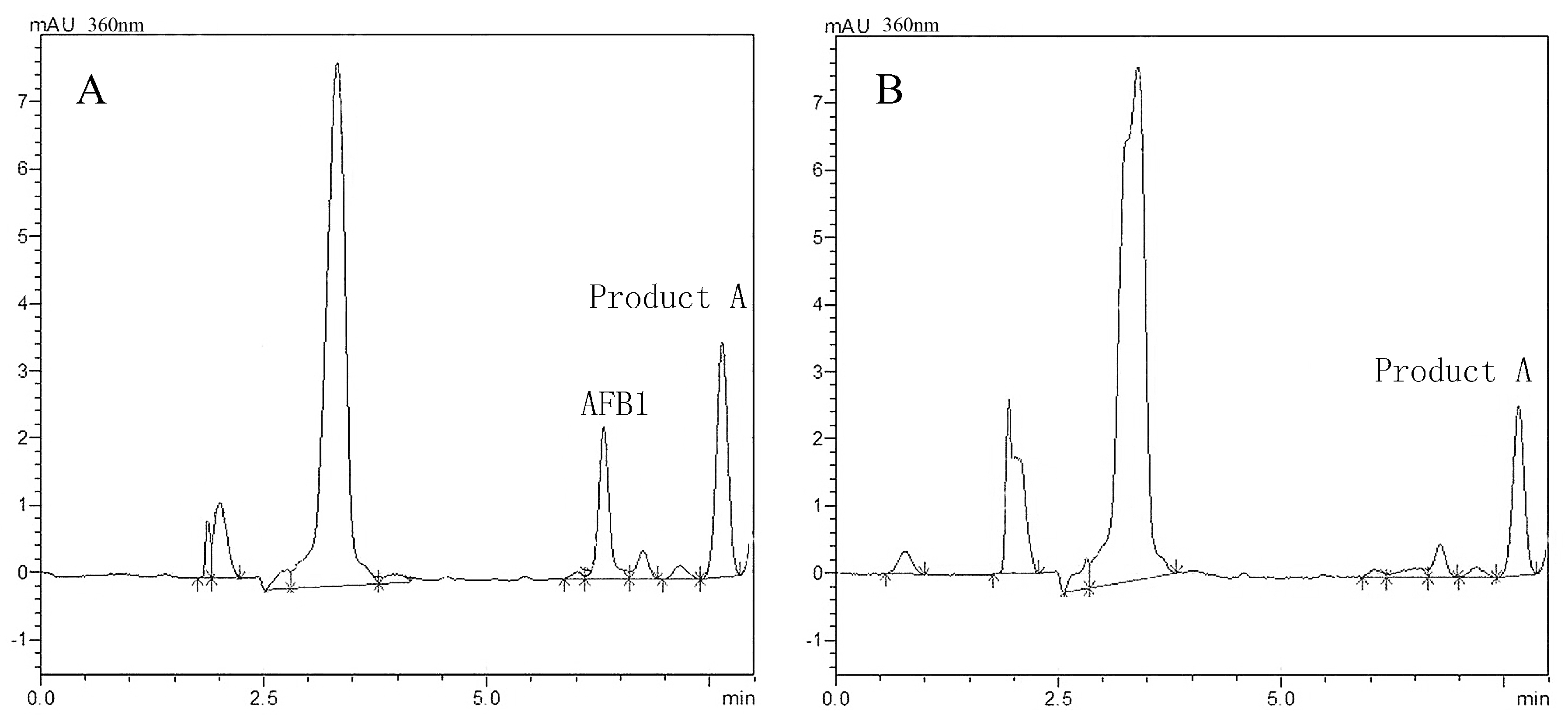
2.4. AFB1 Degradation Product Analysis
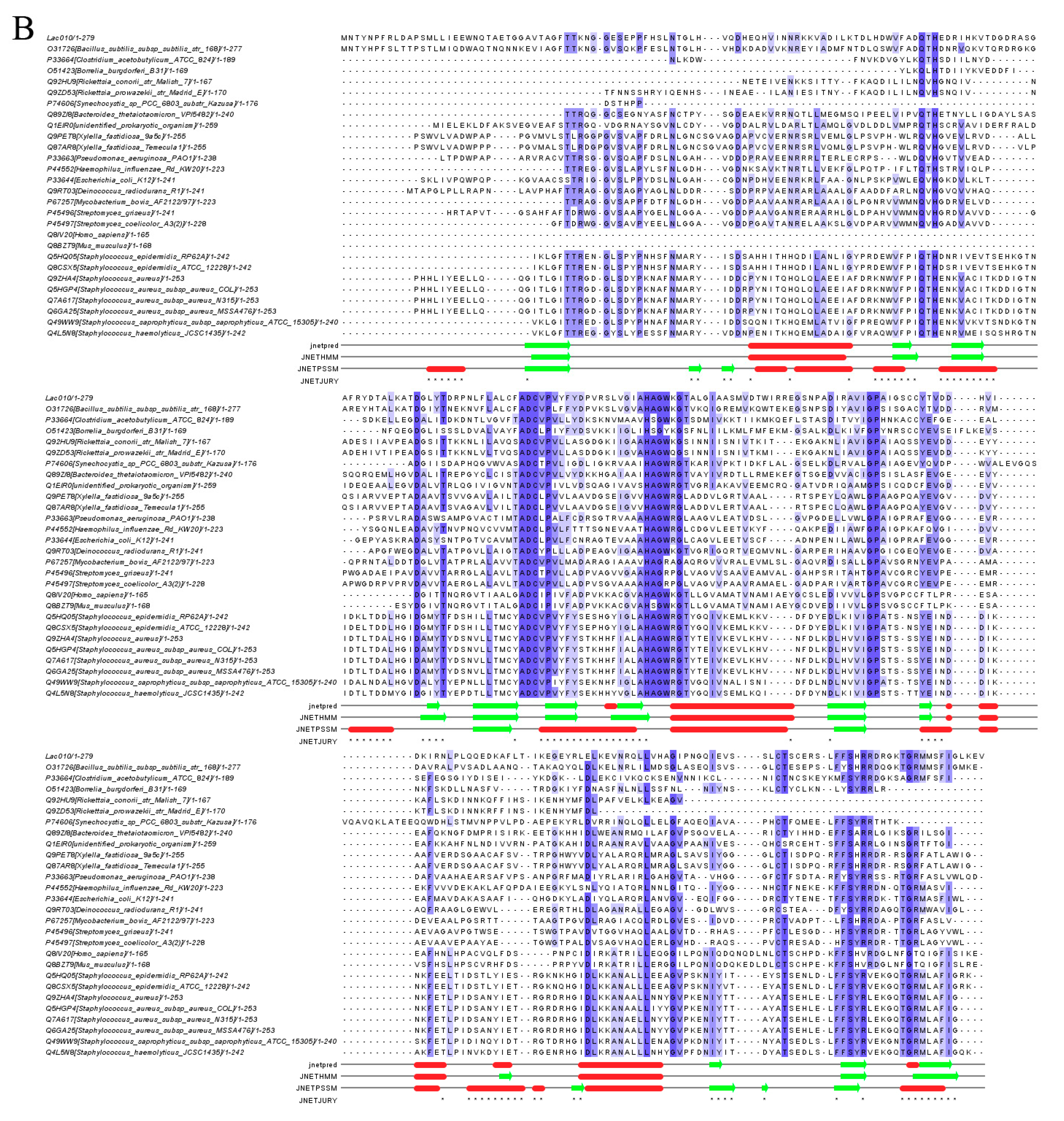
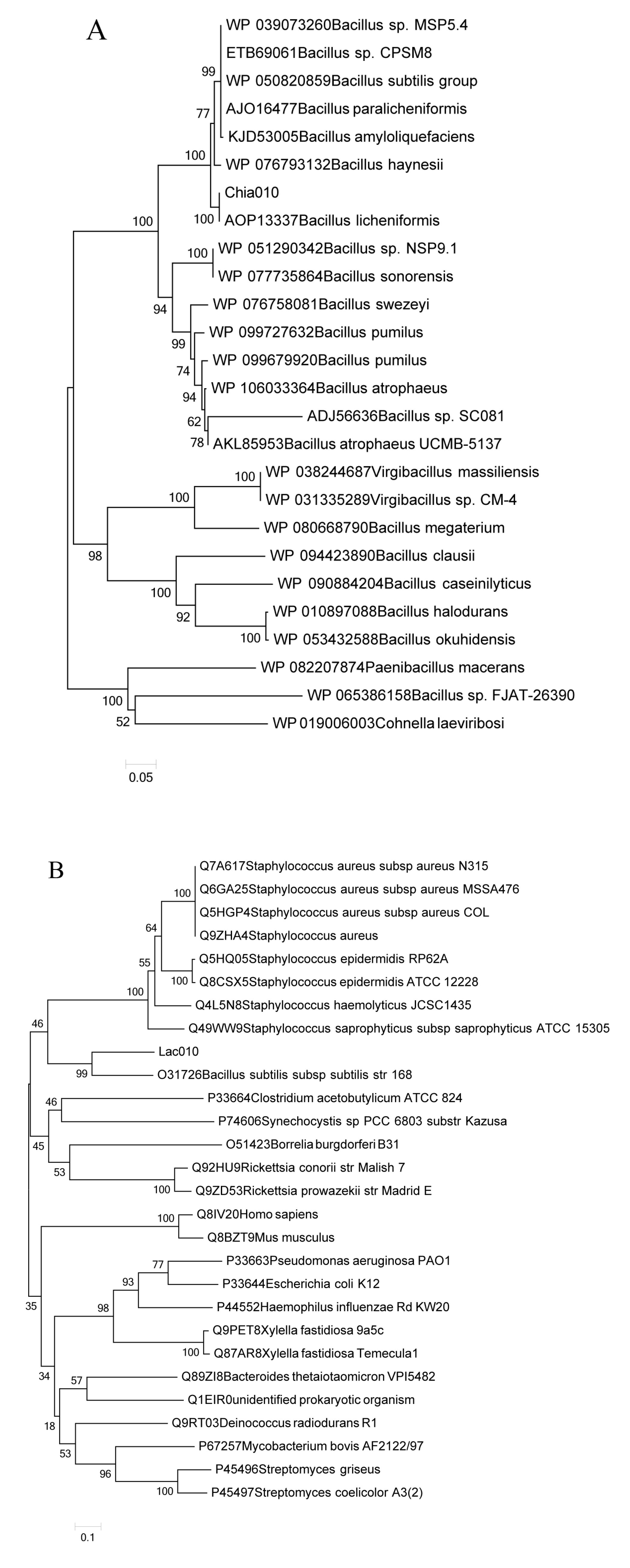
2.5. Genome Identification and Phylogenetic Relationships with Laccase and Chitinase Genes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents and Materials
5.2. Strain and Media
5.3. Biodegradation of AFB1 in ABS
5.4. Effect of Induction on Degradation Enzymes
5.5. Biodegradation Activity Test of BL010 Cell Culture Fluid and Cell-Free Extract
5.6. Analysis of Aflatoxin B1 Concentration and Degradation Products
5.7. Identification and Bioinformatics Analysis of BL010 Degradation Enzymes Genes
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wu, Q.; Jezkova, A.; Yuan, Z.; Pavlikova, L.; Dohnal, V.; Kuca, K. Biological degradation of aflatoxins. Drug Metab. Rev. 2009, 41, 1. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T. Threat of indoor fungi-health risk associated with uncertain mycotoxin exposure: An overview. JSM Mycotoxins 2008, 58, 41–46. [Google Scholar] [CrossRef]
- Mishra, H.N.; Das, C. A Review on Biological Control and Metabolism of Aflatoxin. Crit. Rev. Food Technol. 2003, 43, 245–264. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Peng, H.M.; Wang, X.; Li, B.Q.; Long, M.Y.; Tian, S.P. Biodegradation Mechanisms of Patulin in Candida guilliermondii: An iTRAQ-Based Proteomic Analysis. Toxins 2017, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Hassan, Y.I.; Watts, C.; Zhou, T. Innovative technologies for the mitigation of mycotoxins in animal feed and ingredients—A review of recent patents. Anim. Feed Sci. Technol. 2016, 216, 19–29. [Google Scholar] [CrossRef]
- Leslie, J.F.; Logrieco, A.F. 9. Physical and Chemical Methods for Mycotoxin Decontamination in Maize; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 116–129. [Google Scholar]
- Zhao, L.H.; Guan, S.; Gao, X.; Ma, Q.G.; Lei, Y.P.; Bai, X.M.; Ji, C. Preparation, purification and characteristics of an aflatoxin degradation enzyme from Myxococcus fulvus ANSM068. J. Appl. Microbiol. 2015, 110, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, Z.; Hui, W.; Qiu, H.; Zhang, M.; Shuang, L.; Luo, X.; Song, Y.; Hao, Z.; Ma, W. Rapid Biodegradation of Aflatoxin B1 by Metabolites of Fusarium sp. WCQ3361 with Broad Working Temperature and Excellent Thermostability. J. Sci. Food Agric. 2016, 97, 1342. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, W.; Liu, J.; Pan, D.; Tu, X.; Lv, P.; Wang, Y.; Cao, H.; Wang, Y.; Hua, R. Rapid Biodegradation of the Herbicide 2,4-Dichlorophenoxyacetic Acid by Cupriavidus gilardii T-1. J. Agric. Food Chem. 2017, 65, 3711–3720. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; He, S.; Liang, Z.; Li, Q.X.; Yan, H.; Hu, J.; Liu, X. Biodegradation of pyraclostrobin by two microbial communities from Hawaiian soils and metabolic mechanism. J. Hazard. Mater. 2018, 354, 225. [Google Scholar] [CrossRef] [PubMed]
- Shantha, T. Fungal degradation of aflatoxin B1. Neurogastroenterol. Motil. 2010, 7, 175–178. [Google Scholar] [CrossRef]
- Zjalic, S.; Reverberi, M.; Ricelli, A.; Mario, G.V.; Fanelli, C.; Adele, F.A. Trametes versicolor: A possible tool for aflatoxin control. Int. J. Food Microbiol. 2006, 107, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Smiley, R.D.; Draughon, F.A. Preliminary evidence that degradation of aflatoxin B1 by Flavobacterium aurantiacum is enzymatic. J. Food Prot. 2000, 63, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Teniola, O.D.; Addo, P.A.; Brost, I.M.; Färber, P.; Jany, K.D.; Alberts, J.F.; van Zyl, W.H.; Steyn, P.S.; Holzapfel, W.H. Degradation of aflatoxin B(1) by cell-free extracts of Rhodococcus erythropolis and Mycobacterium fluoranthenivorans sp. nov. DSM44556(T). Int. J. Food Microbiol. 2005, 105, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.S.; Sivaramakrishna, A.; Mehta, A. Degradation and detoxification of aflatoxin B1 by Pseudomonas putida. Int. Biodeter. Biodegr. 2014, 86, 202–209. [Google Scholar] [CrossRef]
- Dowd, P.F.; Vega, F.E.; Nelsen, T.C.; Richard, J.L. Dusky sap beetle mediated dispersal of Bacillus subtilis to inhibit Aspergillus flavus and aflatoxin production in maize Zea mays L. Biocontrol. Sci. Technol. 1998, 8, 221–235. [Google Scholar] [CrossRef]
- Hormisch, D.; Brost, I.; Kohring, G.W.; Giffhorn, F.; Kroppenstedt, R.M.; Stackebrandt, E.; Färber, P.; Holzapfel, W.H. Mycobacterium fluoranthenivorans sp. nov., a fluoranthene and aflatoxin B1 degrading bacterium from contaminated soil of a former coal gas plant. Syst. Appl. Microbiol. 2004, 27, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Elsanhoty, R.M.; Al-Turki, I.A.; Ramadan, M.F. Application of lactic acid bacteria in removing heavy metals and aflatoxin B1 from contaminated water. Water Sci. Technol. 2016, 74, 625–638. [Google Scholar] [CrossRef] [PubMed]
- El-Deeb, B.; Altalhi, A.; Khiralla, G.; Hassan, S.; Gherbawy, Y. Isolation and Characterization of Endophytic Bacilli Bacterium from Maize Grains Able to Detoxify Aflatoxin B1. Food Biotechnol. 2013, 27, 199–212. [Google Scholar] [CrossRef]
- Cao, H.; Liu, D.; Mo, X.; Xie, C.; Yao, D. A fungal enzyme with the ability of aflatoxin B1 conversion: Purification and ESI-MS/MS identification. Microbiol. Res. 2011, 166, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ogata, M.; Hirai, H.; Kawagishi, H. Detoxification of aflatoxin B1 by manganese peroxidase from the white-rot fungus Phanerochaete sordida YK-624. FEMS Microbiol. Lett. 2011, 314, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Dellafiora, L.; Galaverna, G.; Reverberi, M.; Dall’Asta, C. Degradation of Aflatoxins by Means of Laccases from Trametes versicolor: An In Silico Insight. Toxins 2017, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Fanelli, F.; Zucca, P.; Liuzzi, V.C.; Quintieri, L.; Cimmarusti, M.T.; Monaci, L.; Haidukowski, M.; Logrieco, A.F.; Sanjust, E. Aflatoxin B1 and M1 Degradation by Lac2 from Pleurotus pulmonarius and Redox Mediators. Toxins 2016, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Yan, P.S.; Cao, L.X.; Ding, Q.L.; Shao, C.; Zhao, T.F. Potential of chitinolytic Serratia marcescens strain JPP1 for biological control of Aspergillus parasiticus and aflatoxin. BioMed Res. Int. 2013, 2013, 397142. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Feng, K.; Ding, T.; Huang, K.; Yan, H.; Liu, X.; Zhang, Z. Complete genome sequence of Bacillus licheniformis BL-010. Microb. Pathogenesis 2018, 118, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Mochidzuki, K.; Cheng, W.; Zhou, M.; Gao, H.; Zheng, D.; Wang, X.; Cui, Z. Effects of different pretreatment strategies on corn stalk acidogenic fermentation using a microbial consortium. Bioresour. Technol. 2011, 102, 7526–7531. [Google Scholar] [CrossRef] [PubMed]
- Haruta, S.; Cui, Z.; Huang, Z.; Li, M.; Ishii, M.; Igarashi, Y. Construction of a stable microbial community with high cellulose-degradation ability. Appl. Microbiol. Biotechnol. 2002, 59, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Prettl, Z.; Dési, E.; Lepossa, A.; Kriszt, B.; Kukolya, J.; Nagy, E. Biological degradation of aflatoxin B 1 by a Rhodococcus pyridinivorans strain in by-product of bioethanol. Anim. Feed Sci. Technol. 2017, 224. [Google Scholar] [CrossRef]
- Mahunu, G.K.; Zhang, H.; Yang, Q.; Li, C.; Zheng, X. Biological Control of Patulin by Antagonistic Yeast: A case study and possible model. Crit. Rev. Microbiol. 2015, 42, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, C.; Zhang, D.; Zhao, M.; Zheng, D.; Lyu, Y.; Cheng, W.; Guo, P.; Cui, Z. Effective degradation of aflatoxin B1 using a novel thermophilic microbial consortium TADC7. Bioresour. Technol. 2017, 224, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, M.; Shi, Z.Q.; Ghassempour, A.; Sedaghat, N.; Ahmadzadeh, M.; Mirabolfathy, M.; Javan-Nikkhah, M. Aflatoxin B1 degradation by Bacillus subtilis UTBSP1 isolated from pistachio nuts of Iran. Food Control 2012, 23, 100–106. [Google Scholar] [CrossRef]
- Guan, S.; Ji, C.; Zhou, T.; Li, J.; Ma, Q.; Niu, T. Aflatoxin B1 Degradation by Stenotrophomonas maltophilia and Other Microbes Selected Using Coumarin Medium. Int. J. Mol. Sci. 2008, 9, 1489–1503. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.H.; Jun-Xia, L.I.; Yun-Long, H.E.; Guan, S.; Wang, N.; Cheng, J.I.; Niu, T.G. AFB1 Bio-Degradation by a New Strain—Stenotrophomonas sp. J. Integr. Agric. 2008, 7, 1433–1437. [Google Scholar] [CrossRef]
- Sumbu, Z.L.; Thonart, P.; Bechet, J. Action of patulin on a yeast. Appl. Environ. Microbiol. 1983, 45, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Feussner, K.; Wu, T.; Yan, F.; Karlovsky, P.; Zheng, X. Detoxification of mycotoxin patulin by the yeast Rhodosporidium paludigenum. Food Chem. 2015, 179, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Eisa Ahmed, M.F.; Sangare, L.; Zhao, Y.; Selvaraj, J.N.; Xing, F.; Wang, Y.; Yang, H.; Liu, Y. Novel Aflatoxin-Degrading Enzyme from Bacillus shackletonii L7. Toxins 2017, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Risa, A.; Divinyi, D.M.; Baka, E.; Krifaton, C. Aflatoxin B1 detoxification by cell-free extracts of Rhodococcus strains. Acta Microbiol. Immunol. Hung. 2017, 64, 423. [Google Scholar] [CrossRef] [PubMed]
- Sangare, L.; Zhao, Y.; Folly, Y.M.; Chang, J.; Li, J.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Liu, Y. Aflatoxin B1 Degradation by a Pseudomonas Strain. Toxins 2014, 6, 3028. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Yang, Q.; Zhang, H.; Cao, J.; Zhang, X.; Apaliya, M.T. The Possible Mechanisms Involved in Degradation of Patulin by Pichia caribbica. Toxins 2016, 8, 289. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.R.; Vipin, A.V.; Hariprasad, P.; Appaiah, K.A.A.; Venkateswaran, G. Biological detoxification of Aflatoxin B1 by Bacillus licheniformis CFR1. Food Control 2017, 71, 234–241. [Google Scholar] [CrossRef]
- Xia, X.; Zhang, Y.; Li, M.; Garba, B.; Zhang, Q.; Wang, Y.; Zhang, H.; Li, P. Isolation and characterization of a Bacillus subtilis strain with aflatoxin B 1 biodegradation capability. Food Control 2017, 75, 92–98. [Google Scholar] [CrossRef]
- Alberts, J.F.; Engelbrecht, Y.; Steyn, P.S.; Holzapfel, W.H.; van Zyl, W.H. Biological degradation of aflatoxin B1 by Rhodococcus erythropolis cultures. Int. J. Food Microbiol. 2006, 109, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Gao, H.; Gao, Z.; Zhao, H.; Yang, Y.; Wu, Q.; Wu, B.; Jiang, C. Identification and molecular characterization of a metagenome-derived L-lysine decarboxylase gene from subtropical soil microorganisms. PLoS ONE 2017, 12, e185060. [Google Scholar] [CrossRef] [PubMed]
- Drozdetskiy, A.; Cole, C.; Procter, J.; Barton, G.J. JPred4: A protein secondary structure prediction server. Nucleic Acids Res. 2015, 43, W389. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, H.; Zhu, Z.; Ji, F.; Yin, X.; Hong, Q.; Shi, J. Isolation and characterization of Bacillus amyloliquefaciens ZDS-1: Exploring the degradation of Zearalenone by Bacillus spp. Food Control 2016, 68, 244–250. [Google Scholar] [CrossRef]
- Kobayashi, H.; Nagahama, T.; Arai, W.; Sasagawa, Y.; Umeda, M.; Hayashi, T.; Nikaido, I.; Watanabe, H.; Oguri, K.; Kitazato, H. Polysaccharide hydrolase of the hadal zone amphipods Hirondellea gigas. Biosci. Biotechnol. Biochem. 2018, 82, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Philippe, F.; Pelloux, J.; Rayon, C. Plant pectin acetylesterase structure and function: New insights from bioinformatic analysis. BMC Genom. 2017, 18, 456. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Condition |
|---|---|
| Column | Diamonsil C18 5 μm (250 × 4.6 mm) |
| Mobile phase | Acetonitrile (solvent B)/water (solvent A) = 45:55 (v/v) |
| Velocity | 1 mL/min |
| Injection volume | 20 μL |
| Detection wavelength | 360 nm |
| Aflatoxin B1 Retention time | 6.5 min |
| Product A Retention time | 7.7 min |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, H.; Yan, H.; Yin, C.; Liu, Y.; Xu, Q.; Liu, X.; Zhang, Z. Effective Biodegradation of Aflatoxin B1 Using the Bacillus licheniformis (BL010) Strain. Toxins 2018, 10, 497. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120497
Wang Y, Zhang H, Yan H, Yin C, Liu Y, Xu Q, Liu X, Zhang Z. Effective Biodegradation of Aflatoxin B1 Using the Bacillus licheniformis (BL010) Strain. Toxins. 2018; 10(12):497. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120497
Chicago/Turabian StyleWang, Ye, Haiyang Zhang, Hai Yan, Chunhua Yin, Yang Liu, Qianqian Xu, Xiaolu Liu, and Zhongbao Zhang. 2018. "Effective Biodegradation of Aflatoxin B1 Using the Bacillus licheniformis (BL010) Strain" Toxins 10, no. 12: 497. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120497